

Tissue Inhibitor of Matrix Metalloproteinases-1 (TIMP-1) and Pulmonary Involvement in COVID-19 Pneumonia

 , , ,
, , ,  , , , , , and
, , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Chest CT and Image Analysis
2.3. Microfluidic Next Generation Enzyme-Linked Immunosorbent Assay (ELISA)
2.4. Gelatinase Activity by Zymography
2.5. Statistical Analysis
3. Results
3.1. Study Population
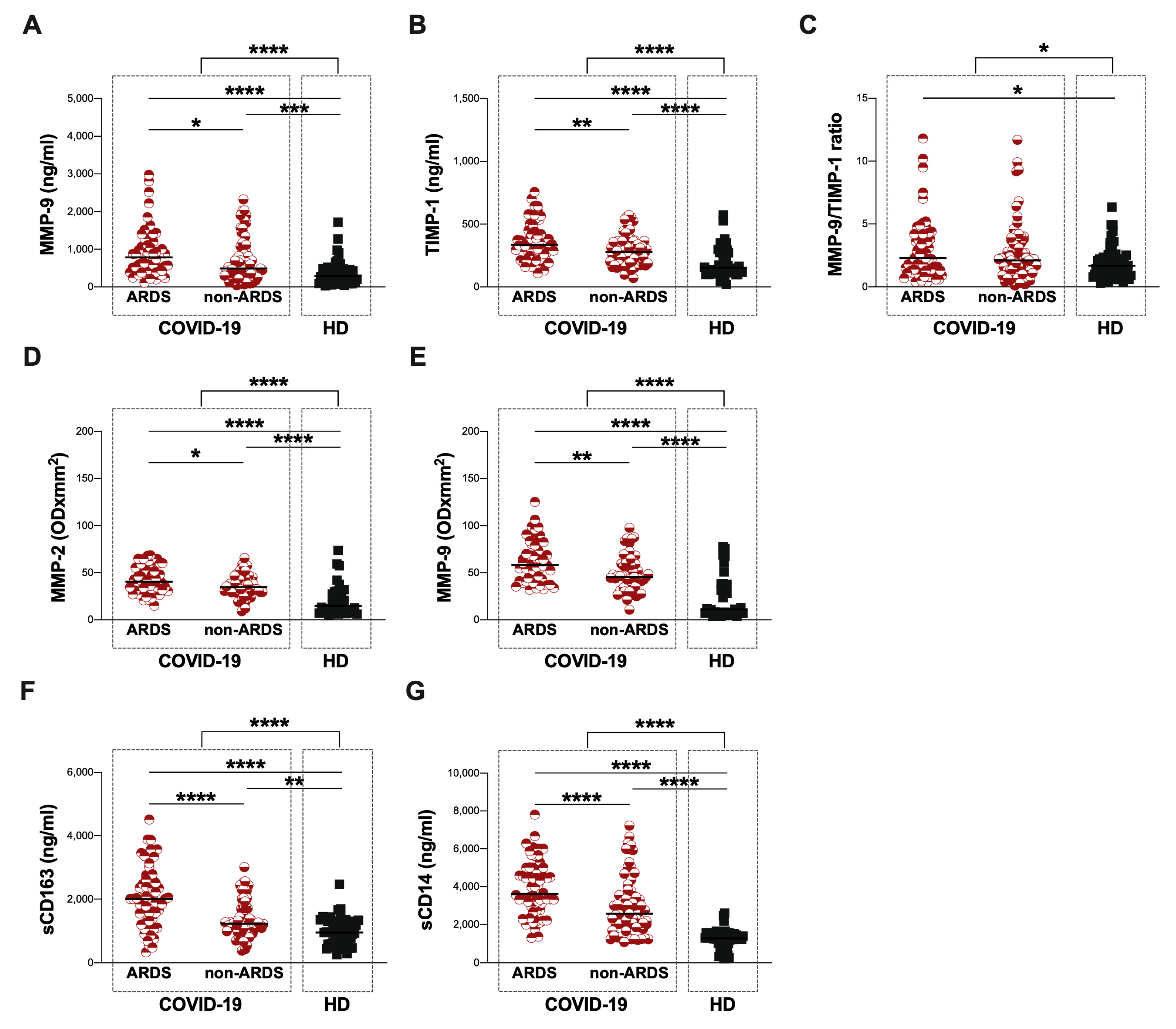
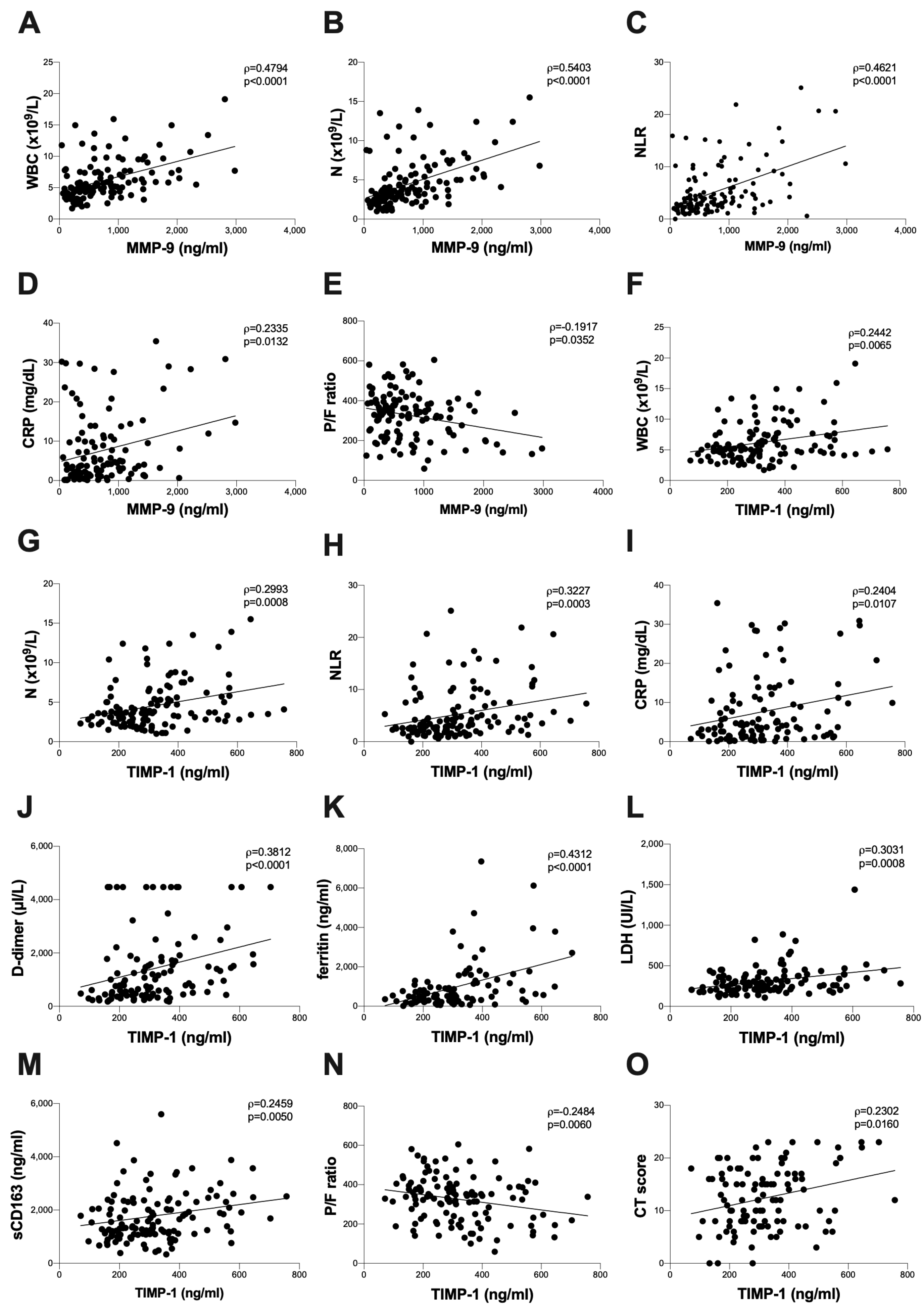
3.2. Baseline Assessment of Plasma Levels of MMP-9 and TIMP-1 and Plasma Activity of MMP-2 and MMP-9
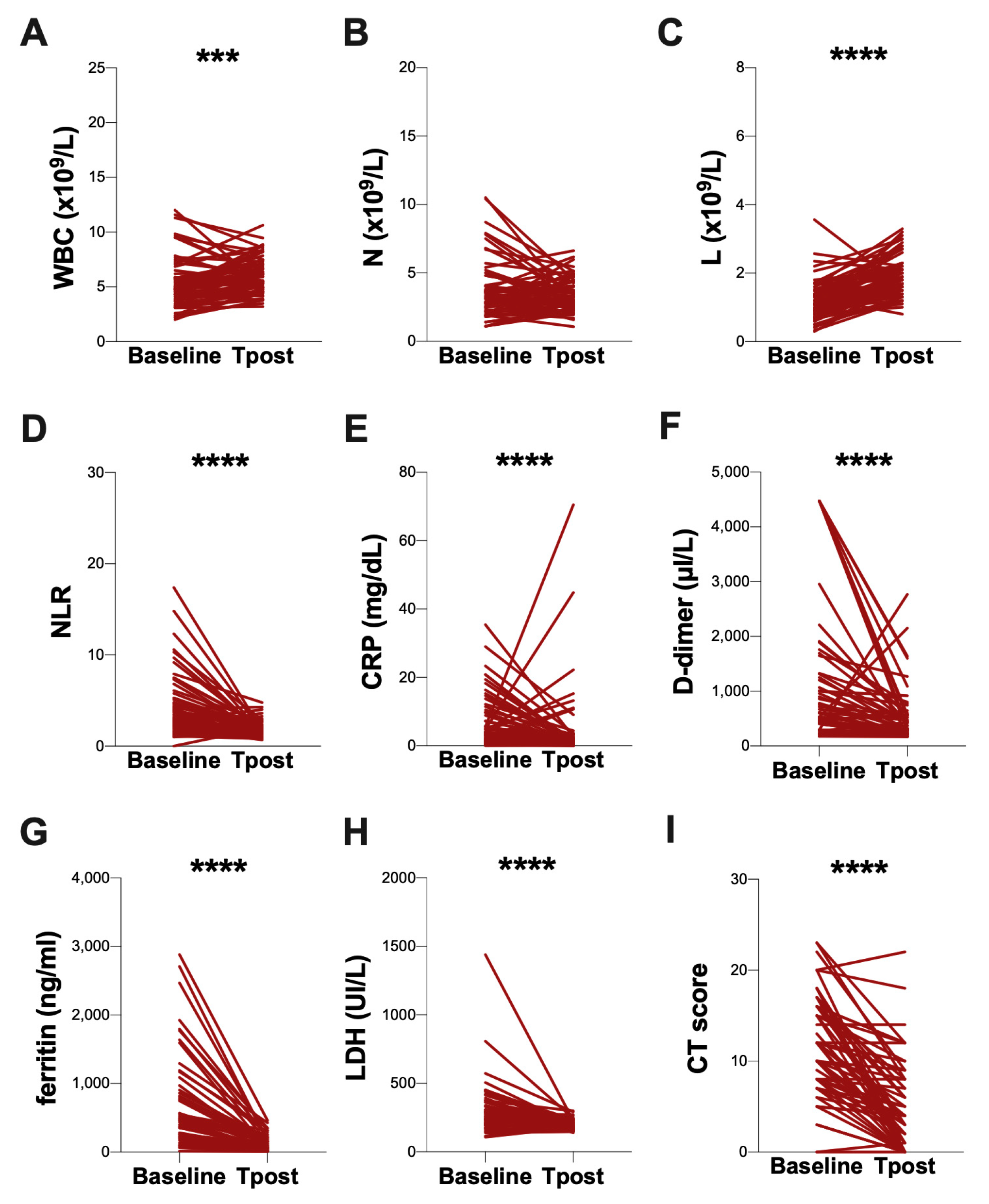
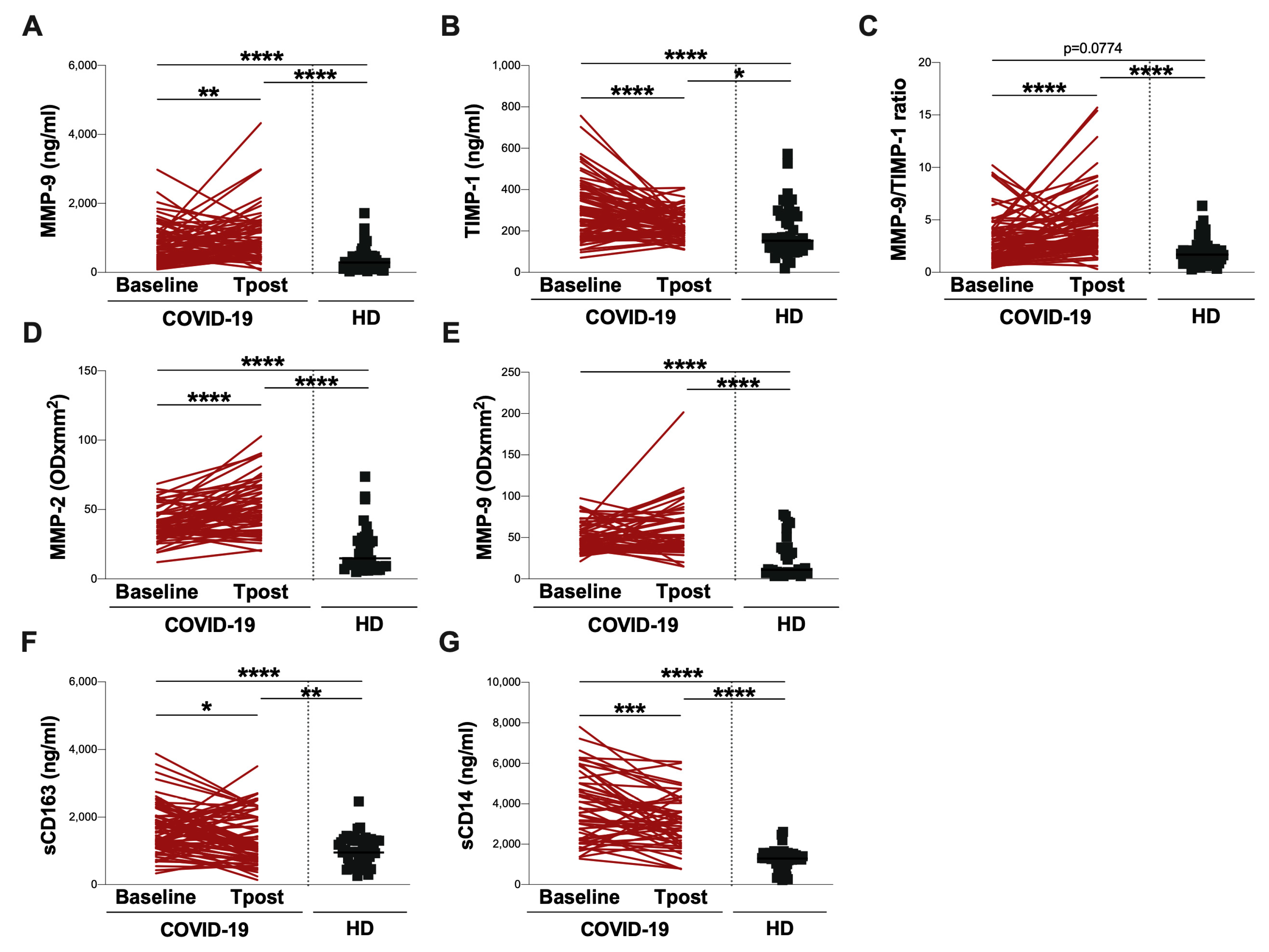
3.3. Evaluation at Post-COVID Clinic
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McGonagle, D.; Sharif, K.; O’Regan, A.; Bridgewood, C. The Role of Cytokines Including Interleukin-6 in COVID-19 Induced Pneumonia and Macrophage Activation Syndrome-Like Disease. Autoimmun. Rev. 2020, 19, 102537. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y.; et al. Clinical Characteristics of 138 Hospitalized Patients With 2019 Novel Coronavirus-Infected Pneumonia in Wuhan, China. JAMA 2020, 323, 1061–1069. [Google Scholar] [CrossRef]
- Zingaropoli, M.A.; Perri, V.; Pasculli, P.; Cogliati Dezza, F.; Nijhawan, P.; Savelloni, G.; La Torre, G.; D’Agostino, C.; Mengoni, F.; Lichtner, M.; et al. Major Reduction of NKT Cells in Patients with Severe COVID-19 Pneumonia. Clin. Immunol. 2021, 222, 108630. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.H.; Li, T.Y.; He, Z.C.; Ping, Y.F.; Liu, H.W.; Yu, S.C.; Mou, H.M.; Wang, L.H.; Zhang, H.R.; Fu, W.J.; et al. A pathological report of three COVID-19 cases by minimal invasive autopsies. Zhonghua Bing Li Xue Za Zhi 2020, 49, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Fox, S.E.; Akmatbekov, A.; Harbert, J.L.; Li, G.; Quincy Brown, J.; Vander Heide, R.S. Pulmonary and Cardiac Pathology in African American Patients with COVID-19: An Autopsy Series from New Orleans. Lancet Respir. Med. 2020, 8, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Middleton, E.A.; He, X.-Y.; Denorme, F.; Campbell, R.A.; Ng, D.; Salvatore, S.P.; Mostyka, M.; Baxter-Stoltzfus, A.; Borczuk, A.C.; Loda, M.; et al. Neutrophil Extracellular Traps Contribute to Immunothrombosis in COVID-19 Acute Respiratory Distress Syndrome. Blood 2020, 136, 1169–1179. [Google Scholar] [CrossRef] [PubMed]
- Veras, F.P.; Pontelli, M.C.; Silva, C.M.; Toller-Kawahisa, J.E.; de Lima, M.; Nascimento, D.C.; Schneider, A.H.; Caetité, D.; Tavares, L.A.; Paiva, I.M.; et al. SARS-CoV-2-Triggered Neutrophil Extracellular Traps Mediate COVID-19 Pathology. J. Exp. Med. 2020, 217, e20201129. [Google Scholar] [CrossRef]
- Santos-Martínez, M.J.; Medina, C.; Jurasz, P.; Radomski, M.W. Role of Metalloproteinases in Platelet Function. Thromb. Res. 2008, 121, 535–542. [Google Scholar] [CrossRef]
- Goetzl, E.J.; Banda, M.J.; Leppert, D. Matrix Metalloproteinases in Immunity. J. Immunol. 1996, 156, 1–4. [Google Scholar] [CrossRef]
- Parks, W.C.; Wilson, C.L.; López-Boado, Y.S. Matrix Metalloproteinases as Modulators of Inflammation and Innate Immunity. Nat. Rev. Immunol. 2004, 4, 617–629. [Google Scholar] [CrossRef]
- Parsons, S.L.; Watson, S.A.; Brown, P.D.; Collins, H.M.; Steele, R.J. Matrix Metalloproteinases. Br. J. Surg. 1997, 84, 160–166. [Google Scholar]
- Overall, C.M. Regulation of Tissue Inhibitor of Matrix Metalloproteinase Expression. Ann. N. Y. Acad. Sci. 1994, 732, 51–64. [Google Scholar] [CrossRef]
- Spinale, F.G. Myocardial Matrix Remodeling and the Matrix Metalloproteinases: Influence on Cardiac Form and Function. Physiol. Rev. 2007, 87, 1285–1342. [Google Scholar] [CrossRef]
- Wang, X.; Khalil, R.A. Matrix Metalloproteinases, Vascular Remodeling, and Vascular Disease. Adv. Pharmacol. 2018, 81, 241–330. [Google Scholar] [CrossRef]
- Aasen, A.O.; Wang, J.E. Mediator Responses in Surgical Infections. Surg. Infect. 2006, 7 (Suppl. S2), S3–S4. [Google Scholar] [CrossRef]
- Krane, S.M. Clinical Importance of Metalloproteinases and Their Inhibitors. Ann. N. Y. Acad. Sci. 1994, 732, 1–10. [Google Scholar] [CrossRef]
- Tokito, A.; Jougasaki, M. Matrix Metalloproteinases in Non-Neoplastic Disorders. Int. J. Mol. Sci. 2016, 17, 1178. [Google Scholar] [CrossRef] [PubMed]
- Klein, T.; Bischoff, R. Physiology and Pathophysiology of Matrix Metalloproteases. Amino Acids 2011, 41, 271–290. [Google Scholar] [CrossRef]
- Gueders, M.M.; Foidart, J.-M.; Noel, A.; Cataldo, D.D. Matrix Metalloproteinases (MMPs) and Tissue Inhibitors of MMPs in the Respiratory Tract: Potential Implications in Asthma and Other Lung Diseases. Eur. J. Pharmacol. 2006, 533, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Busceti, M.T.; Grande, R.; Amato, B.; Gasbarro, V.; Buffone, G.; Amato, M.; Gallelli, L.; Serra, R.; De Franciscis, S. Pulmonary Embolism, Metalloproteinases and Neutrophil Gelatinase Associated Lipocalin—Acta Phlebologica 2013 December. Acta Phlebol. 2013, 14, 115–121. [Google Scholar]
- Beltrán-García, J.; Osca-Verdegal, R.; Pallardó, F.V.; Ferreres, J.; Rodríguez, M.; Mulet, S.; Ferrando-Sánchez, C.; Carbonell, N.; García-Giménez, J.L. Sepsis and Coronavirus Disease 2019: Common Features and Anti-Inflammatory Therapeutic Approaches. Crit. Care Med. 2020, 48, 1841–1844. [Google Scholar] [CrossRef] [PubMed]
- Duda, I.; Krzych, Ł.; Jędrzejowska-Szypułka, H.; Lewin-Kowalik, J. Plasma Matrix Metalloproteinase-9 and Tissue Inhibitor of Matrix Metalloproteinase-1 as Prognostic Biomarkers in Critically Ill Patients. Open Med. 2020, 15, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, A.; Blázquez-Prieto, J.; Amado-Rodriguez, L.; López-Alonso, I.; Batalla-Solís, E.; González-López, A.; Sánchez-Pérez, M.; Mayoral-Garcia, C.; Gutiérrez-Fernández, A.; Albaiceta, G.M. Matrix Metalloproteinase-14 Triggers an Anti-Inflammatory Proteolytic Cascade in Endotoxemia. J. Mol. Med. 2017, 95, 487–497. [Google Scholar] [CrossRef]
- Hazra, S.; Chaudhuri, A.G.; Tiwary, B.K.; Chakrabarti, N. Matrix Metallopeptidase 9 as a Host Protein Target of Chloroquine and Melatonin for Immunoregulation in COVID-19: A Network-Based Meta-Analysis. Life Sci. 2020, 257, 118096. [Google Scholar] [CrossRef]
- Ueland, T.; Holter, J.C.; Holten, A.R.; Müller, K.E.; Lind, A.; Bekken, G.K.; Dudman, S.; Aukrust, P.; Dyrhol-Riise, A.M.; Heggelund, L. Distinct and Early Increase in Circulating MMP-9 in COVID-19 Patients with Respiratory Failure. J. Infect. 2020, 81, e41–e43. [Google Scholar] [CrossRef]
- Abers, M.S.; Delmonte, O.M.; Ricotta, E.E.; Fintzi, J.; Fink, D.L.; de Jesus, A.A.A.; Zarember, K.A.; Alehashemi, S.; Oikonomou, V.; Desai, J.V.; et al. An Immune-Based Biomarker Signature Is Associated with Mortality in COVID-19 Patients. JCI Insight 2021, 6, e144455. [Google Scholar] [CrossRef] [PubMed]
- D’Avila-Mesquita, C.; Couto, A.E.S.; Campos, L.C.B.; Vasconcelos, T.F.; Michelon-Barbosa, J.; Corsi, C.A.C.; Mestriner, F.; Petroski-Moraes, B.C.; Garbellini-Diab, M.J.; Couto, D.M.S.; et al. MMP-2 and MMP-9 Levels in Plasma Are Altered and Associated with Mortality in COVID-19 Patients. Biomed. Pharmacother. 2021, 142, 112067. [Google Scholar] [CrossRef] [PubMed]
- da Silva-Neto, P.V.; do Valle, V.B.; Fuzo, C.A.; Fernandes, T.M.; Toro, D.M.; Fraga-Silva, T.F.C.; Basile, P.A.; de Carvalho, J.C.S.; Pimentel, V.E.; Pérez, M.M.; et al. Matrix Metalloproteinases on Severe COVID-19 Lung Disease Pathogenesis: Cooperative Actions of MMP-8/MMP-2 Axis on Immune Response through HLA-G Shedding and Oxidative Stress. Biomolecules 2022, 12, 604. [Google Scholar] [CrossRef]
- Brusa, S.; Terracciano, D.; Bruzzese, D.; Fiorenza, M.; Stanziola, L.; Pinchera, B.; Valente, V.; Gentile, I.; Cittadini, A.; Mormile, I.; et al. Circulating Tissue Inhibitor of Metalloproteinases 1 (TIMP-1) at COVID-19 Onset Predicts Severity Status. Front. Med. 2022, 9, 1034288. [Google Scholar] [CrossRef]
- Francone, M.; Iafrate, F.; Masci, G.M.; Coco, S.; Cilia, F.; Manganaro, L.; Panebianco, V.; Andreoli, C.; Colaiacomo, M.C.; Zingaropoli, M.A.; et al. Chest CT Score in COVID-19 Patients: Correlation with Disease Severity and Short-Term Prognosis. Eur. Radiol. 2020, 30, 6808–6817. [Google Scholar] [CrossRef] [PubMed]
- Di Carlo, D.; Mazzuti, L.; Sciandra, I.; Guerrizio, G.; Oliveto, G.; Riveros Cabral, R.J.; Zingaropoli, M.A.; Turriziani, O. Comparison of FTD SARS-CoV-2 Assay and RealStar RT-PCR Kit 1.0 for the Detection of SARS-CoV-2. J. Virol. Methods 2021, 298, 114276. [Google Scholar] [CrossRef] [PubMed]
- Pasculli, P.; Zingaropoli, M.A.; Masci, G.M.; Mazzuti, L.; Perri, V.; Paribeni, F.; Russo, G.; Arcari, G.; Iafrate, F.; Vullo, F.; et al. Chest Computed Tomography Score, Cycle Threshold Values and Secondary Infection in Predicting COVID-19 Mortality. New Microbiol. 2021, 44, 145–154. [Google Scholar] [PubMed]
- Zingaropoli, M.A.; Iannetta, M.; Piermatteo, L.; Pasculli, P.; Latronico, T.; Mazzuti, L.; Campogiani, L.; Duca, L.; Ferraguti, G.; De Michele, M.; et al. Neuro-Axonal Damage and Alteration of Blood-Brain Barrier Integrity in COVID-19 Patients. Cells 2022, 11, 2480. [Google Scholar] [CrossRef]
- Ferrazzano, G.; Zingaropoli, M.A.; Costanzo, M.; Belvisi, D.; Dominelli, F.; Pasculli, P.; Ciardi, M.R.; Fabbrini, G.; Defazio, G.; Berardelli, A.; et al. Neurofilament Assessment in Patients with Cervical Dystonia. Park. Relat. Disord. 2022, 98, 70–71. [Google Scholar] [CrossRef]
- Iannetta, M.; Zingaropoli, M.A.; Latronico, T.; Pati, I.; Pontecorvo, S.; Prezioso, C.; Pietropaolo, V.; Cortese, A.; Frontoni, M.; D’Agostino, C.; et al. Dynamic Changes of MMP-9 Plasma Levels Correlate with JCV Reactivation and Immune Activation in Natalizumab-Treated Multiple Sclerosis Patients. Sci. Rep. 2019, 9, 311. [Google Scholar] [CrossRef] [PubMed]
- Petrella, C.; Zingaropoli, M.A.; Ceci, F.M.; Pasculli, P.; Latronico, T.; Liuzzi, G.M.; Ciardi, M.R.; Angeloni, A.; Ettorre, E.; Menghi, M.; et al. COVID-19 Affects Serum Brain-Derived Neurotrophic Factor and Neurofilament Light Chain in Aged Men: Implications for Morbidity and Mortality. Cells 2023, 12, 655. [Google Scholar] [CrossRef]
- Han, X.; Fan, Y.; Alwalid, O.; Li, N.; Jia, X.; Yuan, M.; Li, Y.; Cao, Y.; Gu, J.; Wu, H.; et al. Six-Month Follow-up Chest CT Findings after Severe COVID-19 Pneumonia. Radiology 2021, 299, E177–E186. [Google Scholar] [CrossRef]
- George, P.M.; Wells, A.U.; Jenkins, R.G. Pulmonary Fibrosis and COVID-19: The Potential Role for Antifibrotic Therapy. Lancet Respir. Med. 2020, 8, 807–815. [Google Scholar] [CrossRef]
- Bazdyrev, E.; Rusina, P.; Panova, M.; Novikov, F.; Grishagin, I.; Nebolsin, V. Lung Fibrosis after COVID-19: Treatment Prospects. Pharmaceuticals 2021, 14, 807. [Google Scholar] [CrossRef]
- Howard, E.W.; Bullen, E.C.; Banda, M.J. Regulation of the Autoactivation of Human 72-KDa Progelatinase by Tissue Inhibitor of Metalloproteinases-2. J. Biol. Chem. 1991, 266, 13064–13069. [Google Scholar] [CrossRef] [PubMed]
- Davey, A.; McAuley, D.F.; O’Kane, C.M. Matrix Metalloproteinases in Acute Lung Injury: Mediators of Injury and Drivers of Repair. Eur. Respir. J. 2011, 38, 959–970. [Google Scholar] [CrossRef]
- Fligiel, S.E.G.; Standiford, T.; Fligiel, H.M.; Tashkin, D.; Strieter, R.M.; Warner, R.L.; Johnson, K.J.; Varani, J. Matrix Metalloproteinases and Matrix Metalloproteinase Inhibitors in Acute Lung Injury. Hum. Pathol. 2006, 37, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Apparailly, F.; Noël, D.; Millet, V.; Baker, A.H.; Lisignoli, G.; Jacquet, C.; Kaiser, M.J.; Sany, J.; Jorgensen, C. Paradoxical Effects of Tissue Inhibitor of Metalloproteinases 1 Gene Transfer in Collagen-Induced Arthritis. Arthritis Rheum. 2001, 44, 1444–1454. [Google Scholar] [CrossRef] [PubMed]
- Mostafa Mtairag, E.; Chollet-Martin, S.; Oudghiri, M.; Laquay, N.; Jacob, M.P.; Michel, J.B.; Feldman, L.J. Effects of Interleukin-10 on Monocyte/Endothelial Cell Adhesion and MMP-9/TIMP-1 Secretion. Cardiovasc. Res. 2001, 49, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Murate, T.; Hayakawa, T. Multiple Functions of Tissue Inhibitors of Metalloproteinases (TIMPs): New Aspects in Hematopoiesis. Platelets 1999, 10, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Lambert, E.; Dassé, E.; Haye, B.; Petitfrère, E. TIMPs as Multifacial Proteins. Crit. Rev. Oncol. Hematol. 2004, 49, 187–198. [Google Scholar] [CrossRef]
- Ward, N.S.; Waxman, A.B.; Homer, R.J.; Mantell, L.L.; Einarsson, O.; Du, Y.; Elias, J.A. Interleukin-6-Induced Protection in Hyperoxic Acute Lung Injury. Am. J. Respir. Cell Mol. Biol. 2000, 22, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Veness-Meehan, K.A.; Cheng, E.R.; Mercier, C.E.; Blixt, S.L.; Johnston, C.J.; Watkins, R.H.; Horowitz, S. Cell-Specific Alterations in Expression of Hyperoxia-Induced MRNAs of Lung. Am. J. Respir. Cell Mol. Biol. 1991, 5, 516–521. [Google Scholar] [CrossRef]
- Walsh, J.; Absher, M.; Kelley, J. Variable Expression of Platelet-Derived Growth Factor Family Proteins in Acute Lung Injury. Am. J. Respir. Cell Mol. Biol. 1993, 9, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Pardo, A.; Barrios, R.; Maldonado, V.; Meléndez, J.; Pérez, J.; Ruiz, V.; Segura-Valdez, L.; Sznajder, J.I.; Selman, M. Gelatinases A and B Are Up-Regulated in Rat Lungs by Subacute Hyperoxia: Pathogenetic Implications. Am. J. Pathol. 1998, 153, 833–844. [Google Scholar] [CrossRef]
- Metzemaekers, M.; Cambier, S.; Blanter, M.; Vandooren, J.; de Carvalho, A.C.; Malengier-Devlies, B.; Vanderbeke, L.; Jacobs, C.; Coenen, S.; Martens, E.; et al. Kinetics of Peripheral Blood Neutrophils in Severe Coronavirus Disease 2019. Clin. Transl. Immunol. 2021, 10, e1271. [Google Scholar] [CrossRef] [PubMed]
- Yao, P.M.; Maitre, B.; Delacourt, C.; Buhler, J.M.; Harf, A.; Lafuma, C. Divergent Regulation of 92-KDa Gelatinase and TIMP-1 by HBECs in Response to IL-1beta and TNF-Alpha. Am. J. Physiol. 1997, 273, L866–L874. [Google Scholar] [CrossRef] [PubMed]
- Schnaper, H.W.; Grant, D.S.; Stetler-Stevenson, W.G.; Fridman, R.; D’Orazi, G.; Murphy, A.N.; Bird, R.E.; Hoythya, M.; Fuerst, T.R.; French, D.L. Type IV Collagenase(s) and TIMPs Modulate Endothelial Cell Morphogenesis in Vitro. J. Cell. Physiol. 1993, 156, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Corry, D.B.; Rishi, K.; Kanellis, J.; Kiss, A.; Song Lz, L.; Xu, J.; Feng, L.; Werb, Z.; Kheradmand, F. Decreased Allergic Lung Inflammatory Cell Egression and Increased Susceptibility to Asphyxiation in MMP2-Deficiency. Nat. Immunol. 2002, 3, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Campbell, L.G.; Ramachandran, S.; Liu, W.; Shipley, J.M.; Itohara, S.; Rogers, J.G.; Moazami, N.; Senior, R.M.; Jaramillo, A. Different Roles for Matrix Metalloproteinase-2 and Matrix Metalloproteinase-9 in the Pathogenesis of Cardiac Allograft Rejection. Am. J. Transplant. 2005, 5, 517–528. [Google Scholar] [CrossRef]
- Atkinson, J.J.; Senior, R.M. Matrix Metalloproteinase-9 in Lung Remodeling. Am. J. Respir. Cell Mol. Biol. 2003, 28, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Zymography—An Overview. In Encyclopedia of Biological Chemistry, 3rd ed.; Science Direct Topics; Elsevier: Amsterdam, The Netherlands, 2021; Available online: https://www.sciencedirect.com/topics/biochemistry-genetics-and-molecular-biology/zymography (accessed on 17 May 2023).
- Gómez-Rial, J.; Currás-Tuala, M.J.; Rivero-Calle, I.; Gómez-Carballa, A.; Cebey-López, M.; Rodríguez-Tenreiro, C.; Dacosta-Urbieta, A.; Rivero-Velasco, C.; Rodríguez-Núñez, N.; Trastoy-Pena, R.; et al. Increased Serum Levels of SCD14 and SCD163 Indicate a Preponderant Role for Monocytes in COVID-19 Immunopathology. Front. Immunol. 2020, 11, 560381. [Google Scholar] [CrossRef]
- Zingaropoli, M.A.; Nijhawan, P.; Carraro, A.; Pasculli, P.; Zuccalà, P.; Perri, V.; Marocco, R.; Kertusha, B.; Siccardi, G.; Del Borgo, C.; et al. Increased SCD163 and SCD14 Plasmatic Levels and Depletion of Peripheral Blood Pro-Inflammatory Monocytes, Myeloid and Plasmacytoid Dendritic Cells in Patients With Severe COVID-19 Pneumonia. Front. Immunol. 2021, 12, 627548. [Google Scholar] [CrossRef]
- Marocco, R.; Carraro, A.; Zingaropoli, M.A.; Nijhawan, P.; Tortellini, E.; Guardiani, M.; Mengoni, F.; Zuccalà, P.; Belvisi, V.; Kertusha, B.; et al. Role of Tocilizumab in Down Regulating SCD163 Plasmatic Levels in a Cohort of COVID-19 Patients. Front. Immunol. 2022, 13, 871592. [Google Scholar] [CrossRef]
- Yang, S.-F.; Chu, S.-C.; Chiang, I.-C.; Kuo, W.-F.; Chiou, H.-L.; Chou, F.-P.; Kuo, W.-H.; Hsieh, Y.-S. Excessive Matrix Metalloproteinase-9 in the Plasma of Community-Acquired Pneumonia. Clin. Chim. Acta Int. J. Clin. Chem. 2005, 352, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Lichtinghagen, R.; Pietsch, D.; Bantel, H.; Manns, M.P.; Brand, K.; Bahr, M.J. The Enhanced Liver Fibrosis (ELF) Score: Normal Values, Influence Factors and Proposed Cut-off Values. J. Hepatol. 2013, 59, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Eckfeld, C.; Schoeps, B.; Häußler, D.; Frädrich, J.; Bayerl, F.; Böttcher, J.P.; Knolle, P.; Heisz, S.; Prokopchuk, O.; Hauner, H.; et al. TIMP-1 Is a Novel Ligand of Amyloid Precursor Protein and Triggers a Proinflammatory Phenotype in Human Monocytes. J. Cell Biol. 2023, 222, e202206095. [Google Scholar] [CrossRef] [PubMed]
- Hsu, A.T.; Barrett, C.D.; DeBusk, G.M.; Ellson, C.D.; Gautam, S.; Talmor, D.S.; Gallagher, D.C.; Yaffe, M.B. Kinetics and Role of Plasma Matrix Metalloproteinase-9 Expression in Acute Lung Injury and the Acute Respiratory Distress Syndrome. Shock 2015, 44, 128–136. [Google Scholar] [CrossRef]
- Moore, R.; Hawley, A.; Sigler, R.; Farris, D.; Wrobleski, S.; Ramacciotti, E.; Myers, D. Tissue Inhibitor of Metalloproteinase-1 Is an Early Marker of Acute Endothelial Dysfunction in a Rodent Model of Venous Oxidative Injury. Ann. Vasc. Surg. 2009, 23, 498–505. [Google Scholar] [CrossRef]
- Brew, K.; Nagase, H. The Tissue Inhibitors of Metalloproteinases (TIMPs): An Ancient Family with Structural and Functional Diversity. Biochim. Biophys. Acta 2010, 1803, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.-Y.; Huang, H.-Y.; He, J.-R.; Huang, T.-T.; Heh, C.-C.; Sheng, T.-F.; Chung, K.F.; Kuo, H.-P.; Wang, C.-H. Increased Matrix Metalloproteinase-9 to Tissue Inhibitor of Metalloproteinase-1 Ratio in Smokers with Airway Hyperresponsiveness and Accelerated Lung Function Decline. Int. J. Chron. Obstruct. Pulmon. Dis. 2018, 13, 1135–1144. [Google Scholar] [CrossRef] [PubMed]
- Vignola, A.M.; Riccobono, L.; Mirabella, A.; Profita, M.; Chanez, P.; Bellia, V.; Mautino, G.; D’accardi, P.; Bousquet, J.; Bonsignore, G. Sputum Metalloproteinase-9/Tissue Inhibitor of Metalloproteinase-1 Ratio Correlates with Airflow Obstruction in Asthma and Chronic Bronchitis. Am. J. Respir. Crit. Care Med. 1998, 158, 1945–1950. [Google Scholar] [CrossRef] [PubMed]
- Gelzo, M.; Cacciapuoti, S.; Pinchera, B.; De Rosa, A.; Cernera, G.; Scialò, F.; Comegna, M.; Mormile, M.; Fabbrocini, G.; Parrella, R.; et al. Matrix Metalloproteinases (MMP) 3 and 9 as Biomarkers of Severity in COVID-19 Patients. Sci. Rep. 2022, 12, 1212. [Google Scholar] [CrossRef]
- Cacciola, R.; Gentilini Cacciola, E.; Vecchio, V.; Cacciola, E. Cellular and Molecular Mechanisms in COVID-19 Coagulopathy: Role of Inflammation and Endotheliopathy. J. Thromb. Thrombolysis 2022, 53, 282–290. [Google Scholar] [CrossRef]
- Ramírez-Martínez, G.; Jiménez-Álvarez, L.A.; Cruz-Lagunas, A.; Ignacio-Cortés, S.; Gómez-García, I.A.; Rodríguez-Reyna, T.S.; Choreño-Parra, J.A.; Zúñiga, J. Possible Role of Matrix Metalloproteinases and TGF-β in COVID-19 Severity and Sequelae. J. Interferon Cytokine Res. 2022, 42, 352–368. [Google Scholar] [CrossRef]
- Gutman, H.; Aftalion, M.; Melamed, S.; Politi, B.; Nevo, R.; Havusha-Laufer, S.; Achdout, H.; Gur, D.; Israely, T.; Dachir, S.; et al. Matrix Metalloproteinases Expression Is Associated with SARS-CoV-2-Induced Lung Pathology and Extracellular-Matrix Remodeling in K18-HACE2 Mice. Viruses 2022, 14, 1627. [Google Scholar] [CrossRef] [PubMed]
- Hardy, E.; Fernandez-Patron, C. Targeting MMP-Regulation of Inflammation to Increase Metabolic Tolerance to COVID-19 Pathologies: A Hypothesis. Biomolecules 2021, 11, 390. [Google Scholar] [CrossRef] [PubMed]
- Stetler-Stevenson, W.G. Tissue Inhibitors of Metalloproteinases in Cell Signaling: Metalloproteinase-Independent Biological Activities. Sci. Signal. 2008, 1, re6. [Google Scholar] [CrossRef]
- Grünwald, B.; Schoeps, B.; Krüger, A. Recognizing the Molecular Multifunctionality and Interactome of TIMP-1. Trends Cell Biol. 2019, 29, 6–19. [Google Scholar] [CrossRef] [PubMed]
- Schoeps, B.; Frädrich, J.; Krüger, A. Cut Loose TIMP-1: An Emerging Cytokine in Inflammation. Trends Cell Biol. 2022, 33, P413–P426. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| COVID-19 (n = 129) | ARDS (n = 60) | non-ARDS (n = 69) | p Value * | |
|---|---|---|---|---|
| Male/Female | 57/72 | 44/16 | 28/41 | 0.0013 |
| Age (years) | 64 (55–77) | 70 (63–81) | 58 (50–66) | <0.0001 |
| Deaths/Alive | 24/105 | 22/38 | 2/67 | <0.0001 |
| Comorbidities | ||||
| Any | 86 | 47 | 39 | 0.0407 |
| Hypertension | 52 | 27 | 25 | ns |
| Cardiovascular | 28 | 17 | 11 | ns |
| Diabetes | 23 | 13 | 10 | ns |
| Pulmonary | 23 | 12 | 11 | ns |
| Cancer | 13 | 11 | 2 | 0.0071 |
| Renal | 6 | 5 | 1 | ns |
| Symptoms | ||||
| Fever | 103 | 51 | 52 | ns |
| Cough | 58 | 28 | 30 | ns |
| Shortness of breath | 43 | 29 | 14 | 0.0026 |
| Myalgia or arthralgia | 30 | 8 | 22 | 0.0225 |
| Diarrhea | 16 | 6 | 10 | ns |
| Anosmia and ageusia | 7 | 2 | 5 | ns |
| Sputum production | 3 | 2 | 1 | ns |
| COVID-19 treatment | ||||
| Lopinavir/ritonavir | 32 | 16 | 16 | ns |
| Hydroxychloroquine | 90 | 42 | 48 | ns |
| Azithromycin | 70 | 35 | 35 | ns |
| Enoxaparin | 54 | 32 | 22 | 0.0339 |
| Corticosteroids | 40 | 27 | 13 | 0.0041 |
| Tocilizumab | 54 | 36 | 18 | 0.0004 |
| Laboratory findings | ||||
| WBC (×109/L) | 5.3 (4.3–7.5) | 5.8 (4.4–7.9) | 4.9 (3.9–6.7) | 0.0414 |
| Neutrophils (×109/L) | 3.7 (2.5–5.8) | 4.3 (3.1–6.9) | 3.6 (2.3–5.0) | 0.0254 |
| Lymphocytes (×109/L) | 1.0 (0.7–1.4) | 0.8 (0.6–1.3) | 1.2 (0.8–1.6) | 0.0015 |
| NLR | 3.4 (2.1–7.3) | 4.4 (2.9–10.2) | 2.7 (1.7–4.8) | 0.0006 |
| CRP (mg/dl) | 4.1 (1.4–10.7) | 9.8 (3.0–17.4) | 2.8 (0.7–5.6) | <0.0001 |
| D-dimer (µg/mL) | 899 (443–1762) | 1512 (630–3352) | 590 (423–1319) | 0.0004 |
| Ferritin (ng/mL) | 562 (278–1181) | 999 (347–1800) | 452 (234–712) | 0.0002 |
| LDH (U/L) | 271 (206–373) | 309 (255–436) | 236 (198–303) | <0.0001 |
| P/F ratio | 343 (255–398) | 293 (209–340) | 384 (343–438) | <0.0001 |
| Chest CT score (0–25) | 12 (8–16) | 15 (10–20) | 10 (7–15) | <0.0001 |
| COVID-19 (n = 129) | ARDS (n = 60) | non-ARDS (n = 69) | HD (n = 53) | |
|---|---|---|---|---|
| Next generation ELISA | ||||
| MMP-9 (ng/mL) | 606 (347–1067) | 785 (401–1179) | 489 (309–982) | 287 (124–464) |
| TIMP-1 (ng/mL) | 300 (206–386) | 335 (345–437) | 278 (191–361) | 153 (115–273) |
| MMP-9/TIMP-1 ratio | 2.2 (1.3–3.8) | 2.3 (1.3–4.2) | 2.1 (1.2–3.6) | 1.7 (1.0–2.3) |
| sCD163 (ng/mL) | 1553 (1110–2300) | 2007 (1353–2559) | 1226 (1005–1713) | 952 (588–1300) |
| sCD14 (ng/mL) | 3210 (2198–4548) | 3717 (2920–4996) | 2581 (2006–3557) | 1294 (813–1511) |
| Zymography data | ||||
| MMP-2 (ODxmm2) | 37 (30–48) | 40 (32–53) | 35 (29–45) | 15 (9–28) |
| MMP-9 (ODxmm2) | 49 (37–68) | 58 (42–80) | 46 (34–60) | 11 (7–38) |
| Baseline (n = 78) | T Post (n = 78) | |
|---|---|---|
| Laboratory findings | ||
| WBC (×109/L) | 4.9 (3.9–5.9) | 5.6 (4.7–7.1) |
| Neutrophils (×109/L) | 3.3 (2.4–4.1) | 3.2 (2.5–4.3) |
| Lymphocytes (×109/L) | 1.1 (0.8–1.4) | 1.8 (1.5–2.3) |
| NLR | 2.9 (1.9–4.8) | 1.7 (1.4–2.2) |
| CRP (mg/dl) | 3.2 (1.4–8.9) | 1.1 (0.8–2.4) |
| D-dimer (µg/mL) | 613 (411–1408) | 325 (239–566) |
| Ferritin (ng/mL) | 467 (227–885) | 88 (41–188) |
| LDH (U/L) | 252 (203–363) | 188 (164–218) |
| Chest CT score (0–25) | 12 (8–16) | 4 (0–8) |
| Next generation ELISA | ||
| MMP-9 (ng/mL) | 558 (329–1031) | 805 (465–1224) |
| TIMP-1 (ng/mL) | 292 (201–382) | 211 (177–272) |
| MMP-9/TIMP-1 ratio | 2.3 (1.2–3.7) | 3.6 (2.5–5.4) |
| sCD163 (ng/mL) | 1542 (1121–2027) | 1141 (797–2066) |
| sCD14 (ng/mL) | 3564 (2235–5023) | 3080 (2090–4084) |
| Zymography data | ||
| MMP-2 (ODxmm2) | 37 (31–47) | 50 (35–65) |
| MMP-9 (ODxmm2) | 48 (37–64) | 50 (36–73) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zingaropoli, M.A.; Latronico, T.; Pasculli, P.; Masci, G.M.; Merz, R.; Ciccone, F.; Dominelli, F.; Del Borgo, C.; Lichtner, M.; Iafrate, F.; et al. Tissue Inhibitor of Matrix Metalloproteinases-1 (TIMP-1) and Pulmonary Involvement in COVID-19 Pneumonia. Biomolecules 2023, 13, 1040. https://doi.org/10.3390/biom13071040
Zingaropoli MA, Latronico T, Pasculli P, Masci GM, Merz R, Ciccone F, Dominelli F, Del Borgo C, Lichtner M, Iafrate F, et al. Tissue Inhibitor of Matrix Metalloproteinases-1 (TIMP-1) and Pulmonary Involvement in COVID-19 Pneumonia. Biomolecules. 2023; 13(7):1040. https://doi.org/10.3390/biom13071040
Chicago/Turabian StyleZingaropoli, Maria Antonella, Tiziana Latronico, Patrizia Pasculli, Giorgio Maria Masci, Roberta Merz, Federica Ciccone, Federica Dominelli, Cosmo Del Borgo, Miriam Lichtner, Franco Iafrate, and et al. 2023. "Tissue Inhibitor of Matrix Metalloproteinases-1 (TIMP-1) and Pulmonary Involvement in COVID-19 Pneumonia" Biomolecules 13, no. 7: 1040. https://doi.org/10.3390/biom13071040
APA StyleZingaropoli, M. A., Latronico, T., Pasculli, P., Masci, G. M., Merz, R., Ciccone, F., Dominelli, F., Del Borgo, C., Lichtner, M., Iafrate, F., Galardo, G., Pugliese, F., Panebianco, V., Ricci, P., Catalano, C., Ciardi, M. R., Liuzzi, G. M., & Mastroianni, C. M. (2023). Tissue Inhibitor of Matrix Metalloproteinases-1 (TIMP-1) and Pulmonary Involvement in COVID-19 Pneumonia. Biomolecules, 13(7), 1040. https://doi.org/10.3390/biom13071040