Abstract
Ectopic calcification and dysregulated extracellular matrix remodeling are prominent hallmarks of the complex heterogenous pathobiochemistry of pseudoxanthoma elasticum (PXE). The disease arises from mutations in ABCC6, an ATP-binding cassette transporter expressed predominantly in the liver. Neither its substrate nor the mechanisms by which it contributes to PXE are completely understood. The fibroblasts isolated from PXE patients and Abcc6−/− mice were subjected to RNA sequencing. A group of matrix metalloproteinases (MMPs) clustering on human chromosome 11q21-23, respectively, murine chromosome 9, was found to be overexpressed. A real-time quantitative polymerase chain reaction, enzyme-linked immunosorbent assay and immunofluorescent staining confirmed these findings. The induction of calcification by CaCl2 resulted in the elevated expression of selected MMPs. On this basis, the influence of the MMP inhibitor Marimastat (BB-2516) on calcification was assessed. PXE fibroblasts (PXEFs) exhibited a pro-calcification phenotype basally. PXEF and normal human dermal fibroblasts responded with calcium deposit accumulation and the induced expression of osteopontin to the addition of Marimastat to the calcifying medium. The raised MMP expression in PXEFs and during cultivation with calcium indicates a correlation of ECM remodeling and ectopic calcification in PXE pathobiochemistry. We assume that MMPs make elastic fibers accessible to controlled, potentially osteopontin-dependent calcium deposition under calcifying conditions.
1. Introduction
Patients in whom the rare multisystemic disorder pseudoxanthoma elasticum (PXE, OMIM 264800) has been diagnosed are mostly suffering from symptoms evoked by ectopic calcification, a hallmark of the disease. In most cases, the onset of the disease is characterized by the appearance of dermal yellow papular lesions and excessive wrinkling due to the calcification of elastic fibers [1,2]. Mineralization deposits especially in small vessels elicit atherosclerotic plaques in the cardiovascular system. Hypertension, Claudication intermittent and cardiomyopathy are further complications associated with the disease [3,4]. Calcification of the retinal Bruch’s membrane finally leads to fracturing, observed as angioid streaks, and the formation of hemorrhage in the ocular fundus. In the case of extreme damage to the retina, visual function may be restricted or lost [5,6].
It is assumed that a local imbalance between calcification inhibiting and promoting factors is the cause of the symptoms. Extracellular inorganic pyrophosphate (PPi) has been identified as restraining the local deposition of calcium and phosphate [7,8]. The hydrolytic activity of ectonucleotide phosphodiesterase 1 (ENPP1) leads to the conversion of ATP and, thus, an increasing pyrophosphate level in the extracellular space. In this way, ENPP1 represents another parameter in the calcification regulation network [9]. Patients with PXE feature a diminished plasma PPi level [10,11,12]. Fibroblast cultures from PXE patients contain less cytosolic and extracellular PPi and are characterized by a lower expression of ENPP1 [13,14]. The administration of PPi prevents ectopic calcification in mice [12]. Bisphosphonates, which are stable metabolic PPi analoga, are currently under investigation for therapeutic approaches in PXE [15,16]. The expression of osteopontin (OPN, also known as secreted phosphoprotein 1; gene name SPP1) is coupled with the presence of pyrophosphates [17,18]. OPN is a mineralization-associated protein for which an inhibitory effect on the growth of apatite crystals, consisting of calcium and phosphate, has been demonstrated [19,20].
The ectopic calcification is closely related to another hallmark of PXE: the defects in the extracellular matrix (ECM). Mineral apatite crystals in the core of elastic fibers were identified as disease-defining even before pathologic details or the genetic background were known [8,21,22,23]. Subsequently, several macromolecules were identified to colocalize to these deposits, including OPN [22]. Histologically, the calcification of the mid-dermal elastic fibers manifests in their fragmentation. Moreover, collagen fibers are deformed, and proteoglycans accumulate [24,25]. Matrix metalloproteinases (MMPs) are discussed to relate to the initiation of these ECM defects. The 23 MMP family members in humans and mice degrade a broad spectrum of ECM molecules, thereby contributing to a dynamic ECM remodeling [26,27]. An elevated serum concentration of MMP2 and MMP9 is measurable for PXE patients, while their fibroblasts exhibit a higher expression of MMP2 [28,29].
PXE is inherited autosomal-recessively [30]. Genetic defects in ATP-binding cassette transporter C6 (ABCC6) were associated with the disease for the first time in 2000 [31]. Since then, approximately 400 mutations resulting in the exhibition of PXE symptoms have been described [32]. Considering the systemic expression profile, ABCC6 is mainly expressed in the liver, more specifically in the basolateral plasma membrane of hepatocytes. Its expression in the affected peripheral tissues, such as the skin, is markedly lower [33]. Furthermore, the fibroblasts isolated from PXE patients show a disease-related phenotype even outside the systemic context. This includes a higher responsiveness to calcification stimuli, abnormal ECM remodeling, an altered metabolic lipid profile and cellular senescence [13,34,35,36]. A better understanding of the pathobiochemistry encompassing the whole organism was gained using an Abcc6−/− mouse model. The model was generated by introducing a hygromycin resistance gene into exon 18 and disrupting the correct coding sequence of mAbcc6. These Abcc6−/−-mice resembled the clinical symptoms of PXE patients in large part [37].
This study aimed at a more detailed analysis of the deficits in human and murine fibroblasts with ABCC6-deficiency focusing on the extracellular matrix (ECM) and MMPs. Consequently, we analyzed ectopic calcification and ECM remodeling by treating human fibroblasts with the broad range MMP inhibitor Marimastat under calcifying conditions.
2. Materials and Methods
2.1. Human Cell Culture
Normal human dermal fibroblasts (NHDFs) were obtained from the Coriell Institute of Medical Research (Camden, NJ, USA). PXE fibroblasts (PXEFs), isolated from skin biopsies, were received as described previously [38]. The relevant information is summarized in Table 1.

Table 1.
Characteristics of fibroblasts used in this study. SSM, splice site mutation; hm, homozygous; cht, compound heterozygous; wt, wild type; n.s., not specified.
Primary human dermal fibroblasts were cultured in Dulbecco’s modified Eagle’s medium (Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 4 mM L-glutamine (PAN Biotech, Aidenbach, Germany), 1% (v/v) Penicilin-Streptomycin-Amphotericin B solution (100×; PAN Biotech, Aidenbach, Germany) and 10 % (v/v) fetal calf serum (Gibco, Waltham, MA, USA). The medium was changed twice per week. Meanwhile, fibroblasts were kept under standard conditions of 37 °C and 5 % CO2. Upon confluence, cells were subcultured using 0.05% (v/v) trypsin (PAN Biotech, Aidenbach, Germany).
In terms of the experimental procedure, human fibroblasts between passage eight and eleven were seeded at a density of 177 cells per mm2 and cultivated for 24 h in the complete medium supplemented with 10% fetal calf serum. The fibroblasts were cultured for a further 72 h prior to harvest for the basal expression analysis. According to the calcification experimental set-up, the exchange medium contained DMSO, 10 µM Marimastat (diluted in DMSO; Tocris, Bristol, UK), 8 mM CaCl2 or 10 µM Marimastat plus 8 mM CaCl2 for the calcification experiments. The concentration for the Marimastat application was determined in accordance with several publications [39,40,41]. The application of 8 mM CaCl2 for 21 days to trigger calcification was performed following our previously established protocol [42]. Studies on human fibroblasts were performed in biological and technical triplicates.
2.2. Murine Cell Culture
The Abcc6−/− mouse model generated by Gorgels et al. [37], based on a hybrid model of C57BL/6 and 129/Ola, was backcrossed to C57BL/g for more than five generations. We used Abcc6+/+ C57BL/6 mice as the control animals. Housing, care and breeding were taken over by the central animal facility of Bielefeld University (Germany). The mice were kept with water and food ad libitum. In the present study, the animals were 4 to 6 weeks old. Fibroblasts were isolated from four male and four female animals for each genotype. The mice were anesthetized with isoflurane inhalation (1.5–2%) and killed by cervical dislocation. The animal preparation is in accordance with the law on animal welfare for scientific purposes in Germany because the animals were killed solely for tissue dissection without prior burden. Therefore, ethical approval of this part of the study was not required.
A 1 cm2 piece of skin was cut from the tail of the dead animals. The isolation of adult murine dermal fibroblasts was carried out according to Takashima et al. [43]. In brief, the skin was kept in a 4 mg/mL dispase in phosphate-buffered saline (PBS; Gibco, Waltham, MA, USA) for 4 to 6 h at room temperature. Following a washing step in PBS, the epidermis was deducted from the dermis. The latter was cut in 2 mm2 pieces and incubated in a 1 mg/mL collagenase solution (Sigma, St. Louis, MO, USA) in Dulbecco’s modified Eagle’s medium for 2 h at 37 °C. Subsequently, the reaction was stopped by adding the same amount of the complete murine fibroblast cultivation medium. The latter consisted of Dulbecco’s modified Eagle’s medium supplemented with 1 % (v/v) Penicillin-Streptomycin-Amphotericin, 4 mM L-glutamine, 20 % fetal calf serum, 1 % (v/v) non-essential amino acids (Gibco, Waltham, MA, USA) and 1 % (v/v) HEPES buffer (Gibco, Waltham, MA, USA). After filtering the cell suspension through a 100 µm strainer (Corning, Corning, NY, USA), cells were seeded at a density of 40 cells per mm2. The typical spindle-shaped morphology was confirmed via microscopy. Upon confluence, the murine fibroblasts were seeded for the experiment without further subculture at a density of 177 cells per mm2. Cells were harvested after 96 h. Studies on murine fibroblasts were performed in technical replicates.
2.3. Nucleic Acid Isolation
The RNA isolation from fibroblasts’ cultures was achieved using the NucleoSpin RNA Kit (Macherey-Nagel, Düren, Germany). The DNA for normalization was extracted utilizing the NucleoSpin Blood Extraction Kit (Macherey-Nagel, Düren, Germany). In both cases, the procedure was carried out in accordance with the manufacturer’s instructions. Nucleic acid concentrations were determined on a NanoDrop200 spectrophotometer (Peqlab, Erlangen, Germany).
2.4. Gene Expression Analysis
Quality control, RNA library preparation and sequencing on Illumina NovaSeq6000 for RNA sequencing were taken over by Novogene Co. (Beijing, China). The RNA from two cell lines per sex in biological triplicates was pooled in equal amounts for the analysis.
Regarding the real-time quantitative polymerase chain reaction (RT-qPCR), 1 µg of isolated RNA was initially transcribed into cDNA using SuperScript II Reverse Transcriptase (Thermo Fisher Scientific, Waltham, MA, USA). Measurements were performed with mixtures of 2.5 µL cDNA (1:10 diluted), 0.25 µL forward and reverse primers (Biomers, Ulm, Germany), 2.0 µL water and 5.0 µL LightCycler 480 SYBR Green I Master reaction mixture (Roche, Penzberg, Germany) in a LightCycler 480 system (Roche, Penzberg, Germany). Following an initial incubation for 5 min at 95 °C, the PCR program continued with 45 cycles of denaturation (95 °C, 10 s), annealing (specific annealing temperature, 15 s) and elongation (72 °C, 20 s). Finally, a melting curve analysis was conducted. The target-specific mRNA gene expression was relativized to the gene expression of a selection of three housekeeping genes. These were glyceraldehyde-3-phosphate-dehydrogenase (hGAPDH), hydroxymethylbilane synthase (hHMBS), ribosomal protein L13 (hRPL13) and succinate dehydrogenase complex flavoprotein subunit A (hSDHA) for studies in human cell lines. Eukaryotic translation initiation factor 3 subunit A (mEif3a), glyceraldehyde-3-phosphate dehydrogenase (mGapdh) and hypoxanthine phosphor-ribosyltransferase 1 (mHprt) gene expression were measured for relativization in studies in murine cell lines. The delta-delta Ct method considering PCR efficiency and internal calibration was applied. Studies on human fibroblasts were performed in biological and technical triplicates. Information and sequences of forward and reverse primers can be found in Table 2 and Table 3.
Table 2.
Sequences, annealing temperatures (TA), efficiencies and resulting product sizes of primers used for qRT-PCR in studies based on human fibroblasts.
Table 3.
Sequences, annealing temperatures (TA), efficiencies and resulting product sizes of primers used for qRT-PCR in studies based on murine fibroblasts.
2.5. Enzyme-Linked Immunosorbent Assay
The protein concentrations of MMP3 and OPN in the cell culture supernatant were determined using commercially available ELISA Kits (DMP300 and DOST00; R&D Systems, Abingdon, UK). The results were normalized to the DNA content in the cell lysate.
2.6. hMMP12 Immunofluorescent Staining and Fluorescence Microscopy
Cells were seeded onto coverslips (Ø 13 mm) coated with 5 µg/cm2 rat collagen (ibidi, Gräfelfing Germany) for the fluorescence microscopy experiments. Following the cultivation procedure described above, the cells were washed twice in PBS, fixed for 20 min with 4% paraformaldehyde and washed twice again with PBS. Permeabilization was accomplished by 10 min incubation with 0.1% Triton® X 100 (Roth, Karlsruhe, Germany) in PBS and another two washing steps. The blocking of unspecific binding sites was achieved by the application of 5% bovine serum albumin in PBS for 1 h. Cells were washed twice again with PBS. A primary antibody dilution in 1% bovine serum albumin in PBS was applied (ab137443, 1:100; Abcam, Cambridge, UK) for 2 h. Subsequently, another double washing step was completed before the cells were incubated for 1 h in the presence of a secondary antibody dilution in 1% BSA in PBS (ab150078, 1:400; Abcam, Cambridge, UK). After washing twice, the cells were counterstained with a 0.25 µM DAPI solution (Abcam, Cambridge, UK). After a final double wash step, the coverslips were mounted in ROTI® Mount FluorCare mounting media (Roth, Karlsruhe, Germany). A BZ-X810 microscope (Keyence, Osaka, Japan) was used to capture fluorescence images.
2.7. Alizarin Red Staining
Cells were cultured in 24-well cell culture plates according to the experiment procedure stated above. The fibroblasts were washed twice with PBS and fixed by the addition of 70% ethanol and incubation for 30 s for staining. Subsequently, the cells were incubated for 5 min in a 40 mM alizarin red staining (ARS) solution (Sigma, St. Louis, MO, USA) and washed five times to remove the excess dye. The ARS and quantification were performed according to Gregory et al. [44]. In brief, the dye was dissolved out through incubation with 10% acetic acid for 30 min, and the contents of the well transferred into a microcentrifuge tube which was centrifuged at 18,000× g for 15 min. The vortexed mixture was then heated to 85 °C for 10 min and cooled down on ice for 5 min. Following another centrifugation step, 500 µL of the supernatant was neutralized with 200 µL 10 % (v/v) ammonium hydroxide. The absorbance of the mixture was measured in duplicates at 405 nm using a Tecan Reader infinite 200 Pro (Tecan, Männedorf, Switzerland).
2.8. ENPP1 Activity Assay
The ENPP1 activity was traced by a medium color change due to the conversion of its substrate thymidine 5′-monophosphate p-nitrophenyl ester. After fibroblasts were cultured, as stated above, the assay was executed as published by Lau et al. [45]. In summary, the cultivation medium was replaced by a medium containing 1 mg/mL substrate following a 1 h incubation at 37 °C. The absorbance of the cell culture supernatant at 415 nm was measured in duplicate using a Tecan Reader infinite 200 Pro (Tecan, Männedorf, Switzerland).
2.9. Statistical Analysis
The data of the mRNA gene expression analysis and protein concentration determination by ELISA were presented as mean ± standard error (SEM). The data concerning the ARS quantification and ENPP1 activity test were presented as box and whiskers with bars marking the 10–90 percentile. Statistical significance levels were calculated with non-parametric two-tailed Mann–Whitney U tests. Probability (p) values equal to or below 0.05 were assumed to be statistically significant.
3. Results
3.1. Upregulated Matrix Metalloproteinases Expression upon ABCC6-Deficiency
The degradation and fragmentation of ECM compounds in peripheral tissue are a hallmark of PXE. The grade and symptomatic manifestation of the resulting defects appear to be heterogenous between patients. Moreover, the exact underlying pathomechanism linking ECM irregularity to other hallmarks of this disease, such as the calcification, remains poorly understood.
Fibroblasts are the main cell type involved in the ECM maintenance of the affected peripheral tissue. Hence, we performed an RNA sequencing approach to obtain a better impression of intracellular dysregulations in female and male ABCC6-deficient fibroblasts. For this purpose, we used normal NHDFs as the healthy control as well as PXEFs. Moreover, fibroblasts isolated from a well-established mouse model [37] were utilized to further confirm our findings.
The RNA sequencing data revealed several gene ontology entities with differential expression patterns between NHDFs and PXEFs in female and male fibroblasts. Four of the twenty most significantly dysregulated terms in female PXEFs were directly associated with the ECM. In the case of the male human fibroblasts, this was true for half of the entities (Figure 1A). By further breaking down the list of differentially expressed genes, the group of MMPs stands out.
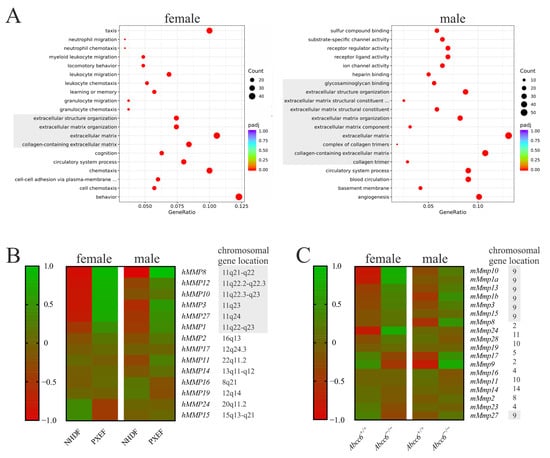
Figure 1.
RNA sequencing data from human and murine fibroblasts with or without ABCC6 deficiency. (A) Differential expression pattern according to the gene ontology database in PXEFs compared to NHDFs assessed separately for female (NHDF n = 2, PXEF n = 2) and male cell lines (NHDF n = 2, PXEF n = 2). Entities directly associated with the extracellular matrix are greyed. (B) Heat map of the differential expression in female and male human NHDFs and PXEFs regarding MMPs plus the specification of the respective gen locus. A location on the gene cluster Chr11q21-24 is highlighted in grey. (C) Heat map of differential expression in murine Abcc6+/+ and Abcc6−/− fibroblasts from female (Abcc6+/+ n = 4, Abcc6−/− n = 4) and male (Abcc6+/+ n = 4, Abcc6−/− n = 4) animals regarding MMPs plus the specification of the respective gen locus. A location on the gene cluster Chr9 is highlighted in grey.
Fourteen MMPs were found to be expressed in significant amounts in human fibroblasts (fragments per kilobase of transcript per million fragments mapped (fpkm) > 0.1). hMMP8, hMMP12, hMMP10, hMMP3, hMMP27, hMMP1 and hMMP2 were overexpressed at least 1.5-fold in the male and female PXEFs compared to NDHFs. Except for hMMP2, the genes of these MMPs were located on a cluster at Chr11q21-24. Additionally, it should be mentioned that hMMP24 and hMMP15 RNA expressions were reduced by at least 50% in female PXEFs in relation to female NHDFs. hMMP17, hMMP11, hMMP14, hMMP16 and hMMP19 RNA expressions were neither diminished nor increased by more than 50% in male or female PXEFs compared to NHDFs (Figure 1B).
Eighteen MMPs were expressed in significant amounts (fpkm > 0.1) in the murine fibroblasts. mMmp10, mMmp1a, mMmp13, mMmp1b and mMmp3 were overexpressed at least 1.5-fold in Abcc6−/− fibroblasts in comparison to Abcc6+/+ fibroblasts. mMmp8 and mMmp17 expressions were upregulated exclusively in Abcc6+/+ fibroblasts derived from male animals, whereas the overexpression of mMmp24 and mMmp28 was observed only in female Abcc6−/− fibroblasts. Surprisingly, mMmp9 RNA expression was increased in male but diminished in female Abcc6−/− fibroblasts relative to its expression in Abcc6+/+ fibroblasts. A difference in the expressions of mMmp15, mMmp19, mMmp16, mMmp11, mMmp14, mMmp2, mMmp23 and mMmp27 with more than 50% deviation was not detected when taking the Abcc6 genotype into account. Similar to the human gene cluster, the genes for mMmp10, mMmp1a, mMmp13, mMmp1b, mMmp3, mMmp15 and mMmp27 clustered on a locus on chromosome 9 (Figure 1C).
Subsequently, the mRNA expressional levels of the selected MMPs observed in the RNA sequencing data were validated via RT-qPCR. The selection was based on (a) the overall expression level, (b) the extent of differential expression between healthy and ABCC6-deficient fibroblasts and (c) the similarity in overexpression between female and male cell lines to summarize the results in a simplified way. Therefore, we defined the expressions of hMMP1, hMMP3, hMMP10 and hMMP12 for the human system and hMmp1a, hMmp3 and hMmp10 for the murine system as appropriate markers. hMMP3 and hMMP12 protein expression was determined using an ELISA or, respectively, immunofluorescent detection.
The comparison of the hMMP1, hMMP3, hMMP10 and hMMP12 mRNA expressions in the human cell lines all revealed a significant induction in PXEFs. It was increased by 5.1-fold (±0.9) for hMMP1, 10.5-fold (±2.4) for hMMP3, 3.9-fold (±0.6) for hMMP10 and 6.6-fold (±1.5) for hMMP12 (Figure 2A).
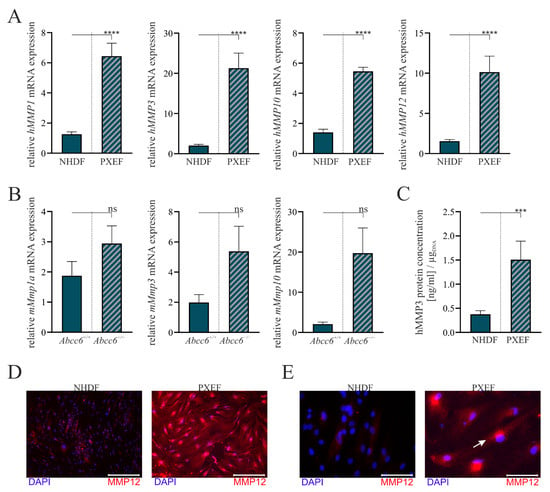
Figure 2.
The MMP expression in human and murine fibroblasts with or without ABCC6 deficiency. Fibroblasts were cultured for 72 h in delipidated medium prior to the harvest. (A) mRNA expression of hMMP1, hMMP3, hMMP10 and hMMP12 in NHDFs (n = 4) and PXEFs (n = 4) and (B) mRNA expression of mMmp1a, mMmp3 and mMmp10 in fibroblasts derived from Abcc6+/+ or Abcc6−/− mice were measured via RT-qPCR. (C) hMMP3 protein concentration in the cell culture supernatant of NHDFs (n = 4) and PXEFs (n = 4) was evaluated utilizing an ELISA and normalizing it to the DNA content in the cell layer. (D,E) Immunofluorescent staining of hMMP12 (red) and nuclear counterstaining with DAPI (blue) of NHDF and PXEF monolayers. Representative images are shown in 10× magnification ((D), scale bar 400 µm) and 40× magnification ((E), scale bar 100 µm). Data are shown as mean ± SEM. Mann–Whitney U test significance levels: not significant (ns), p < 0.001 (***), p < 0.0001 (****).
Differences in MMP mRNA expression patterns were not statistically significant in the murine cell lines but showed a similar tendency to those in human cells. mMmp1a mRNA expression was induced by 1.6-fold (±0.5), mMmp3a mRNA expression was elevated by 2.7-fold (±1.1) and mMmp10 mRNA expression was enhanced by 9.5-fold (±3.7) in Abcc6−/− fibroblasts towards Abcc6+/+ fibroblasts (Figure 2B).
An hMMP3-specific ELISA was utilized to confirm the overexpression on the transcriptional level in the human cell culture model. The hMMP3 protein concentration in cell culture supernatants normalized to the DNA content in the cell layer was found to be 0.4 ng/mL per µgDNA (±0.1) for the NHDFs and 1.5 ng/mL per µgDNA (±0.4) for the PXEFs mirroring a 4.0-fold (±1.2) increase (Figure 2C).
The immunofluorescent labeling of hMMP12 demonstrates an overall higher load of hMMP12 in PXEFs compared to NHDFs, correlating similarly with hMMP12 mRNA expression (Figure 2D). A localization in the endoplasmic reticulum is assumed due to the detection near the nucleus (white arrow) at a higher magnification (Figure 2E).
3.2. Correlation of the Calcification Phenotype and Matrix Metalloproteinases
Our results up to here confirm MMPs, a major regulator in ECM remodeling, as dysregulated upon ABCC6 deficiency. However, it remains unclear how this factor fits into the overall construction of PXE pathobiochemistry, namely, the ectopic calcification. Consequently, we applied a broad range MMP inhibitor, Marimastat, at a concentration of 10 µM alone or in combination with calcification culture conditions (21 days with 8 mM CaCl2) on human dermal fibroblasts.
Major changes in the cellular shape after 21 days were observed comparing the morphology of fibroblasts under control conditions, cultivation with 10 µM Marimastat or 8 mM CaCl2 or both in combination. NHDFs possessed a spindle-shaped cell body and grew to confluence under the control conditions. Partially overlapping cells made it difficult to recognize individual cells (blue arrow). Cultured in the control medium, PXEFs had flat cell bodies resulting in distinguishable cells and a lower cell density (red circle). These typical cell-shape phenotypes were maintained under cultivation with 10 µM Marimastat. When cultured under calcification-favorable circumstances, the morphologies of NHDFs and PXEFs became more similar. The cellular shape of NHDFs was marginally flattened, as seen in the cell in the blue circle exemplarily. Meanwhile, the PXEF morphology became more spindle-shaped and the cell density appeared to be higher (red arrow). An additional application of 10 µM Marimastat had no further influence on the cellular appearance in either NHDFs or PXEFs. Cultivation with 8 mM CaCl2 resulted in the deposits pointed out by white arrows (Figure 3).
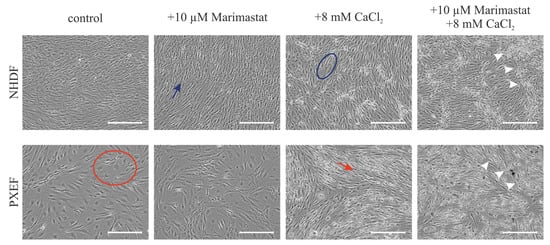
Figure 3.
Representative bright-field microscopy pictures of NHDFs and PXEFs cultured for 21 days. Cultivation conditions included control treatment with DMSO, 10 µM Marimastat, 8 mM CaCl2 or a combination of 10 µM Marimastat and 8 mM CaCl2. Exemplary calcium deposits are highlighted with white arrows. The blue arrow points to an exemplary healthy fibroblast spindle-shape while the blue circle marks an exemplary healthy fibroblast with extended cytoplasm. The red circle surrounds an exemplary PXEF with a flat cell body whereas the red arrow points to a PXEF with an acquired spindle-shape. Scale bars: 400 µm.
In the following section, we study the fibroblasts treated in detail, beginning with the evaluation of MMP expression. The mRNA expression of hMMP3, hMMP1 and hMMP12 was measured and supported by the quantification of hMMP3 in the cell culture supernatant.
A comparison between the mRNA expressions of hMMP3 in NHDFs and PXEFs revealed a 6.7-fold (±1.4) induction between the controls in a similar manner as described above and an 11.4-fold (±2.9) increase under calcifying conditions. While there was no significant change in the hMMP3 mRNA expression upon the application of 10 µM Marimastat, the induction of calcification by 8 mM CaCl2 resulted in a 29.1-fold (±6.4) higher expression in NHDFs. A combined application of Marimastat and CaCl2 did not have any significant impact on this effect. Regarding PXEFs, there was similarly no alteration in the hMMP3 mRNA expression under Marimastat treatment but an increase by 50.7-fold (±14.7) under calcifying conditions. In contrast to the NHDFs, the simultaneous administration of Marimastat and CaCl2 lowered the hMMP3 mRNA expression significantly compared to a sole administration of calcium in PXEFs to 0.8-fold (±0.3) (Figure 4A).
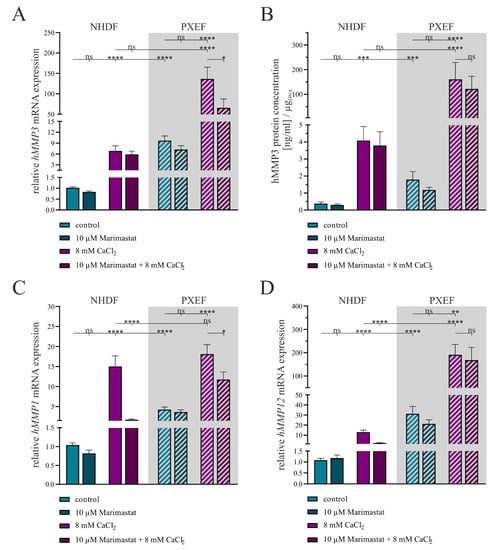
Figure 4.
The expression of MMPs in human dermal fibroblasts under separate or combined treatment with 10 µM Marimastat and 8 mM CaCl2 for 21 days. The NHDFs (n = 3) and PXEFs (n = 3, grey highlighted) were kept under the conditions specified for 21 days. (A) mRNA expressions of hMMP3, (C) hMMP1 and (D) hMMP12 were measured via RT-qPCR. (B) hMMP3 protein concentration in the cell culture supernatant of NHDFs (n = 3) and PXEFs (n = 3) was evaluated utilizing an ELISA and normalizing it to the DNA content in the cell layer. Data are shown as mean ± SEM. Mann–Whitney U test significance levels: not significant (ns), p < 0.05 (*), p < 0.01 (**), p < 0.001 (***), p < 0.0001 (****).
The quantification of hMMP3 in the cell culture supernatants illustrates once more a significantly higher expression in PXEFs relative to that in NHDFs under the control (4.7-fold ± 1.7) and calcifying (39.3-fold ± 18.5) conditions. The amount of hMMP3 was not significantly altered under the application of 10 µM Marimastat in either NHDFs or PXEFs but a higher load of hMMP3 was recognized for both when culturing the fibroblasts with 8 mM CaCl2, namely 10.8-fold (±3.5) in NHDFs and 89.6-fold (±45.0) in PXEFs. A combined application of Marimastat and calcium exhibited no significant reduced hMMP3 expression in either NHDFs or PXEFs (Figure 4B).
The gene expression analysis of hMMP1 disclosed once again a significant elevation by 4.1-fold (±0.6) in PXEFs towards NHDF that was not mirrored under calcification. A gain in hMMP1 gene expression was determined under treatment with 8 mM CaCl2 in NHDFs (14.4-fold ± 2.7) and PXEFs (4.3-fold ± 0.8). By contrast, the application of Marimastat under non-calcifying conditions resulted in a modified hMMP1 expression neither in NHDF nor in PXEF. Under calcifying conditions, supplementation with Marimastat significantly reduced the hMMP1 expression in NHDFs to 0.1-fold (±0.1) and in PXEFs to 0.7-fold (±0.1) (Figure 4C).
As a third marker, hMMP12 gene expression was considered. The NHDFs and PXEFs differed in their hMMP12 expression, reflected by a 28.8-fold (±7.0) and 14.7-fold (±4.1) induction under calcifying conditions, respectively. While a treatment with 10 µM Marimastat had no influence on this factor in either of the cell types, the application of 8 mM CaCl2 resulted in NHDF and in PXEFs in a 12.0-fold (±2.1) and a 6.1-fold (±2.0) higher hMMP12 mRNA level, respectively. The PXEFs simultaneously treated with Marimastat and calcium did not feature a modified hMMP12 gene expression, but the same conditions lead to a significant decline of hMMP12 RNA in NHDFs (0.18-fold ± 0.04) (Figure 4D).
Having evaluated the course of MMP expression in our experimental set-up, our next step was to assess the influence of the MMP inhibitor Marimastat administration on calcification. Therefore, we performed a quantitative ARS, determined the gene expression of hSPP1 and hENPP1 and confirmed the results with an hOPN ELISA and an ENPP1 activity assay.
Alizarin red, a dye to color calcium deposits, was used to evaluate the grade of calcification in the fibroblast monolayers. Exemplary pictures demonstrate the successful staining and a stronger redness of cultures treated with CaCl2 (Figure 5A).
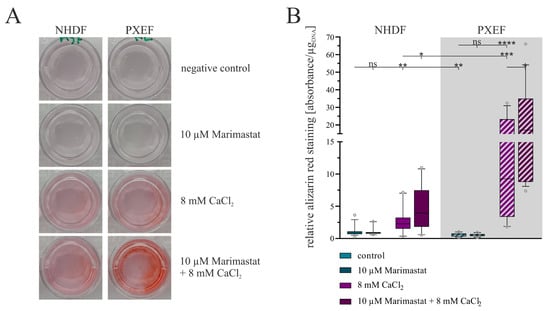
Figure 5.
Evaluation of the calcification status in human fibroblasts concerning separate or combined treatment with 10 µM Marimastat and 8 mM CaCl2 for 21 days. The NHDFs (n = 3) and PXEFs (n = 3, grey highlighted) were kept under the conditions specified for 21 days. (A) Exemplary pictures of fibroblasts’ cell culture wells after ARS and (B) its quantification normalized to the DNA content in the cell layer and relative to the staining in control NHDFs. Data are shown as mean and boxes marking the 10–90 percentile. Mann–Whitney U test significance levels: not significant (ns), p < 0.05 (*), p < 0.001 (**), p < 0.001 (***), p < 0.0001 (****).
A subsequent quantification of the staining attested this impression. Both NHDFs and PXEFs exhibited a higher grade of staining after cultivation with 8 mM CaCl2 compared to cultivation under control conditions: the staining was 2.4-fold (±0.6) stronger in NHDFs and 20.8-fold (±5.2) stronger in PXEFs. The basal calcification mapped by the degree of dyeing was found to be significantly lower in PXEF than in NHDF (0.5-fold ± 0.1) but higher under calcification conditions (4.6-fold ± 1.3). The additional administration of 10 µM Marimastat had no significant influence on the staining in non-calcifying circumstances. Marimastat and calcium in combination triggered a gain in the dyeing in comparison to cultivation with CaCl2 alone. For NHDF, the factor for this variance amounted to 1.8-fold (±0.4), and for PXEF, 1.9-fold (±0.5) (Figure 5B).
The relative hSPP1 gene expression, a marker for the mineralization of tissues, did not differ significantly under control conditions between NHDFs and PXEFs. The additional application of 10 µM Marimastat had no influence on the control status. By contrast, the hSPP1 mRNA expression was significantly enhanced when cultivation was carried out in the presence of 8 mM CaCl2 in both NHDFs (6.4-fold ± 1.1) and PXEFs (1.8-fold ± 0.4). By direct comparison, PXEFs exhibited a 0.5-fold (±0.1) reduced hSPP1 gene expression within this condition. Beyond this observation, a further addition of Marimastat enlarged the hSPP1 gene expression increase by another 2.3-fold (±0.8) in NHDFs and 1.7-fold (±0.3) in PXEFs compared to CaCl2 administration only (Figure 6A).
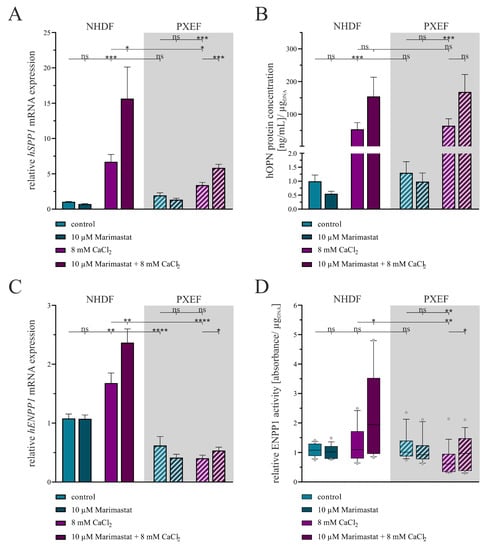
Figure 6.
Evaluation of the calcification status in human fibroblasts concerning separate or combined treatment with 10 µM Marimastat and 8 mM CaCl2 for 21 days. The NHDFs (n = 3) and PXEFs (n = 3, grey highlighted) were kept under the conditions specified for 21 days. (A) Relative hSPP1 mRNA gene expression measured by RT-qPCR and (B) hOPN concentration in cell culture supernatant determined by ELISA and normalized to the DNA content in the cell layer. (C) Relative hENPP1 mRNA expression assessed by RT-qPCR and (D) ENPP1 activity in the cell culture supernatant normalized to the DNA content in the cell layer. (A–C) Data are shown as mean ± SEM or (D) as mean and boxes marking the 10–90 percentile. Mann–Whitney U test significance levels: not significant (ns), p < 0.05 (*), p < 0.001 (**), p < 0.001 (***), p < 0.0001 (****).
An hOPN ELISA was conducted with cell culture supernatant samples to substantiate the gene expression results. A significant difference in the hOPN concentration in the supernatants between NHDFs and PXEFs was not ascertainable in direct comparison under control and calcifying conditions or in the presence of 10 µM Marimastat. Nevertheless, culturing the fibroblasts with 8 mM CaCl2 provoked a rise in the hOPN concentration by 53.7-fold (±23.7) in NHDFs and 49.7-fold (±22.3) in PXEFs. A combined treatment with Marimastat and calcium further enhanced, although not significantly, the accumulation of hOPN in the supernatants of NHDFs and PXEFs by 2.9-fold (±1.6) and 2.6-fold (±1.2), respectively (Figure 6B).
Regarding the hENPP1 gene expression, a representative marker for calcification inhibition and regulation, a significantly diminished value was determined in PXEFs compared to NHDFs under the control (0.6-fold ± 0.1) and calcifying conditions (0.2-fold ± 0.1). The cultivation only with 10 µM Marimastat had no additional effect at that point in either of the fibroblast types. While the administration of 8 mM CaCl2 did not influence the hENPP1 mRNA level in PXEFs, the calcifying condition resulted in its 1.6-fold (±0.2) increase in NHDFs. Furthermore, Marimastat and CaCl2 in combination evoked an induction of hENPP1 mRNA expression by 1.4-fold (±0.2) in NHDFs and 1.3-fold (±0.2) in PXEFs (Figure 6C).
An ENPP1 activity assay, based on the implementation of its substrate thymidine 5′-monophosphate p-nitrophenyl ester, was performed to strengthen the gene expression results. Both NHDFs and PXEFs were found to have an equal ENPP1 activity in control cultures that was not influenced by 10 µM Marimastat. NHDF exhibited a slight, yet not significant, rise in ENPP1 activity by 1.2-fold (±0.1) in a calcifying surrounding brought about by 8 mM CaCl2, while this parameter was reduced significantly by 0.6-fold (±0.1) in PXEFs under the same condition. Supplementation with 10 µM Marimastat had no impact on the ENPP1 activity in PXEFs treated with CaCl2 but led to an even higher ENPP1 activity in NHDFs (1.8-fold ± 0.3) compared to only CaCl2 cultivation (Figure 6D).
4. Discussion
The biggest challenge in adequately managing PXE still remains the understanding of its basic pathomechanisms. As the disease is characterized by a complex multisystemic pathobiochemistry influenced by a variety of genetic modifiers [46] not only the identification of the ABCC6 substrate, assumed to be predominantly transported into the bloodstream by hepatocytes [47], is of the utmost interest. Instead, the research on and connection of all resulting related pathomechanisms will enable us to ensure good care for the patients. These are as diverse as the manifestation of the disease itself and include the ectopic calcification in the dermis, retinal and small vessels [48,49]; the abnormal remodeling of the ECM [24,25,50]; altered lipid metabolisms [35,51,52]; premature aging and senescence [53,54]; and a proinflammatory phenotype [36]. The present study focused on a better understanding between the ectopic calcification and the abnormal dermal ECM remodeling.
Our first approach was to identify predominantly altered gene ontologies in human and murine dermal fibroblasts with ABCC6 deficiency by RNA sequencing to assess reliable markers for a comparison. As expected, due to the phenotype of these exhibited fibroblasts described previously [29,50], modifications in ECM-related ontologies were prevailing. A profound examination of the genes affected revealed striking changes in the MMP expression. The involvement of MMPs in PXE pathobiochemistry was unsurprising. MMP2 has been outlined to raise the degradative potential of human PXEFs [29]. Patients feature elevated MMP2 and MMP9 serum levels, and an MMP2 promoter polymorphism is suspected as a genetic modifier for the disease [28,55].
However, to date, no extensive study on the expression of the whole MMP family has been carried out. RNA sequencing allowed us to close this gap. Our data confirmed a slight mRNA overexpression of MMP2 in humans, but not in murine fibroblasts. Instead, we found the gene expression of hMMP8, hMMP12, hMMP10, hMMP3, hMMP27 and hMMP1 to be increased. The fact that these genes clustered on chromosome 11q21-23 was remarkable [56]. An RNA sequencing analysis of fibroblasts isolated from the murine Abcc6−/− mouse model further strengthened this observation. Here, the expressions of mMmp10, mMmp1a, mMmp13, mMmp1b and mMmp3 were elevated compared to fibroblasts isolated from wild-type mice. Together with mMmp15 and mMmp27, these genes clustered on chromosome 9. Here, the close orthologous relation between the human and the murine gene clusters as well as the corresponding MMPs had to be distinguished. The cellular function of the individual MMPs in these groups was negligible due to their large substrate diversity [26]. Furthermore, minor differences in the expression patterns within these clusters might be due to species-specific regulation and display a point for further investigation. Affirmative representative measurements of the mMmp1a, mMmp3 and mMmp10 mRNA gene expression in murine fibroblasts via RT-qPCR went along with the results from RNA sequencing, although they were not statistically significant. This fact might be due to the high variances between the murine fibroblast cultures. The results gained from the murine cell cultures should be considered as additional supportive data in the present study as we were limited in the number of murine cell lines. As this was a limitation of our study, further studies focusing on the role of MMPs in the murine model are of high interest.
Therefore, we focused on the human fibroblast cultures in the remaining parts of the study. For these, we successfully demonstrated a significant increase in mRNA expression levels of hMMP1, hMMP3, hMMP10 and hMMP12 by a multiple. The findings could be transferred to the protein level, demonstrated by the quantification of MMP3 in the cell culture supernatant and immunofluorescent staining of MMP12. Consequently, a strong correlation between MMP mRNA expression and translation into protein was indicated. Moreover, the nuclear near-detection of MMP12 might correspond to localization in the endoplasmic reticulum and subsequent secretion to the extracellular space. This fits with the characteristic that MMPs are predominantly secreted except from individual membrane-bound family members (such as MMP14 and MMP15) [57]. Although MMPs are regulated at several stages, including post-translational secretion and activation, the transcriptional regulation is of the greatest importance, as our results regarding PXEFs confirm [58].
It is worthwhile to take a common expression regulation into account for the finding that a cluster of MMPs is overexpressed due to ABCC6 deficiency. The 5′ promotors’ regions of the MMP genes involved share a TATA Box and, except for hMMP8, binding sites for activator protein 1 (AP-1) and polyoma enhancer A-binding protein 3 (PEA3) [58]. The binding of the two corresponding transcription factors is a necessary factor in MMP gene transcription modulated by exogenous signals, such as cytokines, growth factors or cell matrix interactions [59,60]. The factor decisive for the overexpression in the present case can only be assumed and requires further studies. A potential mediator might be interleukin-1β, a proinflammatory cytokine, which is known to induce MMP expression and is associated with a severe course of PXE [61,62]. Interestingly, this induction process was observed in calcific aortic valve stenosis, an age-related simultaneous occurrence of calcification and ECM remodeling [63]. However, the binding of activator elements in the promotor region might not be the only factor contributing to the gene expression regulation of MMPs. The hypomethylation of the promotor region resulted in the overexpression of several MMPs. This was notably proven for human osteoarthritic chondrocytes: another disease featuring abnormal ECM dynamics and calcification [64]. Taking these considerations into account, a calcification-dependent induction of cluster Chr11q21-23 MMPs’ expression by a calcification response element in the promotors is conceivable.
Against this background, the fact that we observed a potent induction of exemplarily measured hMMP1, hMMP3 and hMMP12 mRNA expression and hMMP3 content in cell culture supernatants for fibroblasts cultured with CaCl2 fits well. The induction was detectable for NHDFs and PXEFs, although the final amount was higher in PXEFs due to the increased basal expression. A general association of, especially, vascular calcification and MMPs has been described primarily for rodents. Subsequently to the induction of aortic calcification in rats, the administration of the MMP inhibitor GM6001 resulted in the reduced calcification of arteries [65]. Another rat model, in this case displaying vascular calcification in chronic kidney diseases, exhibited a raised MMP-2 and MMP-9 expression and activity upon arterial calcification [66]. The MMP knock-out mice did not respond to the administration of CaCl2 with arterial calcification [67]. The hypothesis which emerged from these studies was, therefore, that the MMPs made elastic fibers of arteries accessible to calcium deposits, thereby linking these two hallmarks of PXE pathobiochemistry. Moreover, the linkage was underlined by an approach to intervene in the DNA damage response of PXEFs, resulting in the reduced hMMP2 expression and diminished calcification simultaneously [68].
We created an in vitro cultivation set-up to evaluate the status of human fibroblasts treated with Marimastat under standard and calcifying conditions to apply the assumption of a correlation between calcification and MMP-based elastic fiber degradation to our model. Marimastat or BB2516 is a broad range MMP inhibitor initially developed to treat migration-dependent metastasis in cancer [69]. It should be noted that the targeted inhibition of individual MMPs could be a focus for further studies to specify their functional role in the biochemistry of PXE.
Here, the application of Marimastat led to marginal changes but no full repression in MMP expression, which goes along with the fact that Marimastat inhibits the MMP activity by mimicking the substrate and competing with it rather than intervening in its expression [70]. No differences in morphology in either NHDFs or PXEFs were observed upon Marimastat treatment. The administration of the calcification medium had two effects on the appearance properties of the cultures. Firstly, massive deposits, later identified as calcium-containing by ARS, were observed in the NDHF and PXEF cultures. Secondly, PXEFs showed a decline in their senescence-associated morphology represented by the spread shape turning into a spindle shape. The opposite was observed for NHDFs. A possible conclusion might be that PXEFs are adapted to calcification conditions, which is reflected by their higher responsiveness to calcification stimuli [13]. Future studies considering the proliferation and migration rates of NHDF and PXEF under calcifying conditions may provide further insight into the general adaptation of fibroblasts to these circumstances.
We assessed the calcification status of the fibroblasts by the quantification of alizarin red-stained deposits; the expression of the apatite crystal formation-inhibitor OPN; and the mRNA expression and activity of ENPP1, which are relevant in the PPi household and, thereby, prevent calcification. As expected, calcifying conditions induced calcium deposition, as well as regulating mechanisms of calcification displayed by OPN expression and, in the case of NHDF, ENPP1 expression and activity. Upon cultivation with calcium, the higher potential for PXEFs to calcify was displayed by a stronger ARS, a lower mRNA expression of hSPP1 and hENPP1 and a reduced ENPP1 activity compared to NHDFs. These data resemble the observations of failing calcification restriction by inhibitors in PXEFs made previously [13,14].
The ambivalent results gained under combined treatment with Marimastat were of greater interest.
The application of the MMP inhibitor provoked an extended deposition of calcium in both NHDFs and PXEFs. This observation was rather surprising as it was stated that MMP inhibitors can prevent calcification in vivo [65,66]. In addition, it must be considered that MMPs do not only affect elastic fibers, even though these are essentially affected by ECM calcification in PXE. The overexpression of MMPs reduces cell–matrix contact. This has been shown to induce the calcification of vessel walls by vascular smooth muscle cells [71,72]. In further experiments, this relationship could be elucidated by culturing on culture surfaces coated with different ECM components.
Furthermore, the Marimastat treatment caused an induction of OPN expression in NHDFs and PXEFs, and thereby contributed to the prevention of apatite crystal growth. Moreover, it led to a gain in ENPP1 gene expression and activity in NHDFs, similarly supporting an anti-calcifying effect. It should be noted that PXEFs did not respond to Marimastat or the CaCl2 stimuli by adjusting their PPi household through ENPP1. The latter is compliant with the declining PPi availability not only in fibroblasts’ cultures but also in the serum of PXE patients [11,13,14].
The fact that hSPP1 has been identified as a modifier gene in PXE reflects that OPN is critical for limiting the calcification process in this disease [73]. A hypothetical explanation for our surprising findings that MMP inhibition enhances calcification, and MMPs might as well have a regulatory effect on ectopic calcification, might therefore be found in the association of OPN and elastic fibers. A fine analysis of the calcified and fragmented elastic fibers performed by Contri et al. revealed an association of OPN and other calcium-binding proteins to the mineral deposits [22]. In the case of a calcium overflow, the fibroblasts may react with the secretion of MMPs and, thus, make elastic fibers accessible for calcium association with proteins, such as OPN. This hypothesis is strengthened by a study using aluminum to pre-treat elastic fibers and, thereby, prevent the degradation by MMPs and calcium deposition in the fiber core [67]. Apart from that, the consequence might be that no excess calcium is intercepted by incorporation into the elastic fibers, where OPN restricts further crystal growth. In this way, the calcification of elastic fibers would represent a mechanism to handle calcium overload by fibroblasts.
The inhibition of elastic fiber-based calcium interception in our cell culture model may have led to the strengthening of other anti-calcifying processes, such as ENPP1 expression. Nonetheless, since calcification is based on an enormous regulatory network [74,75], it cannot be ruled out that other mechanisms not considered in this work will come into play in this case.
The fact that an in vitro model is limited when mirroring the in vivo situation should be considered. Future research may focus on the ultrastructure of elastic fibers under MMP inhibitor treatment and the systemic consequences.
In conclusion, our study gives detailed insight into the contribution of MMPs to calcification in the pathomechanism of PXE. Not only was a whole spectrum of MMP genes belonging to the human cluster Chr11q21-23 overexpressed in PXEFs, but those MMPs were also strongly associated with the induction of calcification. The results were strengthened by the overexpression of a related MMP gene cluster on chromosome 9 in the Abcc6−/− mouse model. The treatment with Marimastat confirmed the synergy between matrix degradation and calcification. Contrary to expectations, Marimastat did not abolish excessive calcification but reinforced it, giving a first hint regarding a defanging of excess calcium by the incorporation and binding to OPN inside elastic fibers.
Author Contributions
Conceptualization, C.K. and D.H.; methodology, R.P. and C.L.; validation, R.P., C.L. and M.R.O.; formal analysis, R.P.; investigation, R.P., C.L. and M.R.O.; resources, C.K. and D.H.; data curation, R.P.; writing—original draft preparation, R.P.; writing—review and editing, C.L., M.R.O., C.K. and D.H.; visualization, R.P.; supervision, C.K.; project administration, D.H.; funding acquisition, D.H. All authors have read and agreed to the published version of the manuscript.
Funding
We acknowledge support by the Open Access Publication Funds of the Ruhr-Universität Bochum.
Institutional Review Board Statement
The isolation and study of human fibroblasts were conducted in accordance with the Declaration of Helsinki, and approved by the Ethics Committee of the HDZ NRW, Department of Medicine, Ruhr-University of Bochum (registry no. 32/2008, approval date is 3rd November 2008). As the animals were killed solely for tissue dissection without prior burden, the animal preparation is in accordance with the law on animal welfare for scientific purpose in Germany. Therefore, ethical approval of this part of the study was not required.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The original raw data and materials presented in the study will be made available upon request. Further inquiries can be directed to the corresponding author.
Acknowledgments
We are grateful to all PXE patients and their relatives and to the Selbsthilfegruppe für PXE-Erkrankte Deutschlands e.V. We also thank Christoph Lichtenberg for his excellent technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Neldner, K.H. Pseudoxanthoma elasticum. Int. J. Dermatol. 1988, 27, 98–100. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Plomp, A.S.; van Soest, S.; Wijnholds, J.; de Jong, P.T.V.M.; Bergen, A.A.B. Pseudoxanthoma elasticum: A clinical, histopathological, and molecular update. Surv. Ophthalmol. 2003, 48, 424–438. [Google Scholar] [CrossRef] [PubMed]
- Jensen, O.A. Bruch’s membrane in pseudoxanthoma elasticum. Albrecht von Graefes Arch. Klin. Ophthalmol. 1977, 203, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Connor, P.J.; Juergens, J.L.; Perry, H.O.; Hollenhorst, R.W.; Edwards, J.E. Pseudoxanthoma elasticum and angioid streaks: A review of 106 cases. Am. J. Med. 1961, 30, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Laube, S.; Moss, C. Pseudoxanthoma elasticum. Arch. Dis. Child. 2005, 90, 754–756. [Google Scholar] [CrossRef]
- Campens, L.; Vanakker, O.M.; Trachet, B.; Segers, P.; Leroy, B.P.; De Zaeytijd, J.; Voet, D.; De Paepe, A.; De Backer, T.; De Backer, J. Characterization of cardiovascular involvement in pseudoxanthoma elasticum families. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 2646–2652. [Google Scholar] [CrossRef]
- Terkeltaub, R.A. Inorganic pyrophosphate generation and disposition in pathophysiology. Am. J. Physiol. Cell. Physiol. 2001, 281, C1–C11. [Google Scholar] [CrossRef]
- Fleisch, H.; Bisaz, S. Mechanism of calcification: Inhibitory role of pyrophosphate. Nature 1962, 195, 911. [Google Scholar] [CrossRef]
- Hessle, L.; Johnson, K.A.; Anderson, H.C.; Narisawa, S.; Sali, A.; Goding, J.W.; Terkeltaub, R.; Millán, J.L. Tissue-nonspecific alkaline phosphatase and plasma cell membrane glycoprotein-1 are central antagonistic regulators of bone mineralization. Proc. Natl. Acad. Sci. USA 2002, 99, 9445–9449. [Google Scholar] [CrossRef]
- Jansen, R.S.; Küçükosmanoglu, A.; de Haas, M.; Sapthu, S.; Otero, J.A.; Hegman, I.E.M.; Bergen, A.A.B.; Gorgels, T.G.M.F.; Borst, P.; van de Wetering, K. ABCC6 prevents ectopic mineralization seen in pseudoxanthoma elasticum by inducing cellular nucleotide release. Proc. Natl. Acad. Sci. USA 2013, 110, 20206–20211. [Google Scholar] [CrossRef]
- Sánchez-Tévar, A.M.; García-Fernández, M.; Murcia-Casas, B.; Rioja-Villodres, J.; Carrillo, J.L.; Camacho, M.; Van Gils, M.; Sánchez-Chaparro, M.A.; Vanakker, O.; Valdivielso, P. Plasma inorganic pyrophosphate and alkaline phosphatase in patients with pseudoxanthoma elasticum. Ann. Transl. Med. 2019, 7, 798. [Google Scholar] [CrossRef]
- Pomozi, V.; Julian, C.B.; Zoll, J.; Pham, K.; Kuo, S.; Tőkési, N.; Martin, L.; Váradi, A.; Le Saux, O. Dietary pyrophosphate modulates calcification in a mouse model of pseudoxanthoma elasticum: Implication for treatment of patients. J. Investig. Dermatol. 2019, 139, 1082–1088. [Google Scholar] [CrossRef] [PubMed]
- Boraldi, F.; Annovi, G.; Bartolomeo, A.; Quaglino, D. Fibroblasts from patients affected by pseudoxanthoma elasticum exhibit an altered PPi metabolism and are more r to pro-calcifying stimuli. J. Dermatol. Sci. 2014, 74, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Dabisch-Ruthe, M.; Kuzaj, P.; Götting, C.; Knabbe, C.; Hendig, D. Pyrophosphates as a major inhibitor of matrix calcification in pseudoxanthoma elasticum. J. Dermatol. Sci. 2014, 75, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Sundberg, J.P.; Levine, M.A.; Terry, S.F.; Uitto, J. The effects of bisphosphonates on ectopic soft tissue mineralization caused by mutations in the ABCC6 gene. Cell Cycle 2015, 14, 1082–1089. [Google Scholar] [CrossRef]
- Shimada, B.K.; Pomozi, V.; Zoll, J.; Kuo, S.; Martin, L.; Le Saux, O. ABCC6, Pyrophosphate and ectopic calcification: Therapeutic Solutions. Int. J. Mol. Sci. 2021, 22, 4555. [Google Scholar] [CrossRef] [PubMed]
- Addison, W.N.; Azari, F.; Sørensen, E.S.; Kaartinen, M.T.; McKee, M.D. Pyrophosphate inhibits mineralization of osteoblast cultures by binding to mineral, up-regulating osteopontin, and inhibiting alkaline phosphatase activity. J. Biol. Chem. 2007, 282, 15872–15883. [Google Scholar] [CrossRef]
- Harmey, D.; Hessle, L.; Narisawa, S.; Johnson, K.A.; Terkeltaub, R.; Millán, J.L. Concerted regulation of inorganic pyrophosphate and osteopontin by Akp2, Enpp1, and Ank: An integrated model of the pathogenesis of mineralization disorders. Am. J. Pathol. 2004, 164, 1199–1209. [Google Scholar] [CrossRef]
- Denhardt, D.T.; Guo, X. Osteopontin: A protein with diverse functions. FASEB J. 1993, 7, 1475–1482. [Google Scholar] [CrossRef]
- Butler, W.T. The nature and significance of osteopontin. Connect. Tissue Res. 1989, 23, 123–136. [Google Scholar] [CrossRef]
- Walker, E.R.; Frederickson, R.G.; Mayes, M.D. The mineralization of elastic fibers and alterations of extracellular matrix in pseudoxanthoma elasticum: Ultrastructure, immunocytochemistry, and X-ray analysis. Arch. Dermatol. 1989, 125, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Contri, M.B.; Boraldi, F.; Taparelli, F.; De Paepe, A.; Ronchetti, I.P. Matrix proteins with high affinity for calcium ions are associated with mineralization within the elastic fibers of pseudoxanthoma elasticum dermis. Am. J. Pathol. 1996, 148, 569–577. [Google Scholar] [PubMed]
- Otkjær-Nielsen, A.; Johnson, E.; Hentzer, B.; Danielsen, L.; Carlsen, F. Apatite crystals in pseudoxanthoma elasticum: A combined study using electron microscopy and selected area diffraction analysis. J. Investig. Dermatol. 1977, 69, 376–378. [Google Scholar] [CrossRef] [PubMed]
- Tiozzo Costa, R.; Contri, M.B.; Cingi, M.R.; Pasquali Ronchetti, I.; Salvini, R.; Rindi, S.; De Luca, G. Pseudoxanthoma elasticum (PXE): Ultrastructural and biochemical study on proteoglycan and proteoglycan-associated material produced by skin fibroblasts in vitro. Coll. Relat. Res. 1988, 8, 49–64. [Google Scholar] [CrossRef]
- Gheduzzi, D.; Sammarco, R.; Quaglino, D.; Bercovitch, L.; Terry, S.; Taylor, W.; Ronchetti, I.P. Extracutaneous ultrastructural alterations in pseudoxanthoma elasticum. Ultrastruct. Pathol. 2003, 27, 375–384. [Google Scholar] [CrossRef]
- Jackson, B.C.; Nebert, D.W.; Vasiliou, V. Update of human and mouse matrix metalloproteinase families. Hum. Genom. 2010, 4, 194. [Google Scholar] [CrossRef]
- Nagase, H.; Visse, R.; Murphy, G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovasc. Res. 2006, 69, 562–573. [Google Scholar] [CrossRef]
- Diekmann, U.; Zarbock, R.; Hendig, D.; Szliska, C.; Kleesiek, K.; Götting, C. Elevated circulating levels of matrix metalloproteinases MMP-2 and MMP-9 in pseudoxanthoma elasticum patients. J. Mol. Med. 2009, 87, 965–970. [Google Scholar] [CrossRef]
- Quaglino, D.; Sartor, L.; Garbisa, S.; Boraldi, F.; Croce, A.; Passi, A.; De Luca, G.; Tiozzo, R.; Pasquali-Ronchetti, I. Dermal fibroblasts from pseudoxanthoma elasticum patients have raised MMP-2 degradative potential. Biochim. Biophys. Acta 2005, 1741, 42–47. [Google Scholar] [CrossRef]
- Ringpfeil, F.; McGuigan, K.; Fuchsel, L.; Kozic, H.; Larralde, M.; Lebwohl, M.; Uitto, J. Pseudoxanthoma elasticum is a recessive disease characterized by compound heterozygosity. J. Investig. Dermatol. 2006, 126, 782–786. [Google Scholar] [CrossRef]
- Bergen, A.A.B.; Plomp, A.S.; Schuurman, E.J.; Terry, S.; Breuning, M.; Dauwerse, H.; Swart, J.; Kool, M.; van Soest, S.; Baas, F.; et al. Mutations in ABCC6 cause pseudoxanthoma elasticum. Nat. Genet. 2000, 25, 228–231. [Google Scholar] [CrossRef] [PubMed]
- Pseudoxanthoma Elasticum and ABCC6. ClinVar. NCBI. Available online: https://www.ncbi.nlm.nih.gov/clinvar/?term=pseudoxanthoma+elasticum+abcc6 (accessed on 9 December 2022).
- Beck, K.; Hayashi, K.; Dang, K.; Hayashi, M.; Boyd, C.D. Analysis of ABCC6 (MRP6) in normal human tissues. Histochem. Cell. Biol. 2005, 123, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Le Saux, O.; Bunda, S.; VanWart, C.M.; Douet, V.; Got, L.; Martin, L.; Hinek, A. Serum factors from pseudoxanthoma elasticum patients alter elastic fiber formation in vitro. J. Investig. Dermatol. 2006, 126, 1497–1505. [Google Scholar] [CrossRef]
- Kuzaj, P.; Kuhn, J.; Michalek, R.D.; Karoly, E.D.; Faust, I.; Dabisch-Ruthe, M.; Knabbe, C.; Hendig, D. Large-scaled metabolic profiling of human dermal fibroblasts derived from pseudoxanthoma elasticum patients and healthy controls. PLoS ONE 2014, 9, e108336. [Google Scholar] [CrossRef] [PubMed]
- Tiemann, J.; Wagner, T.; Lindenkamp, C.; Plümers, R.; Faust, I.; Knabbe, C.; Hendig, D. Linking ABCC6 deficiency in primary human dermal fibroblasts of PXE patients to P21-mediated premature cellular senescence and the development of a proinflammatory secretory phenotype. Int. J. Mol. Sci. 2020, 21, 9665. [Google Scholar] [CrossRef] [PubMed]
- Gorgels, T.G.M.F.; Hu, X.; Scheffer, G.L.; van der Wal, A.C.; Toonstra, J.; de Jong, P.T.V.M.; van Kuppevelt, T.H.; Levelt, C.N.; de Wolf, A.; Loves, W.J.P.; et al. Disruption of Abcc6 in the mouse: Novel insight in the pathogenesis of pseudoxanthoma elasticum. Hum. Mol. Genet. 2005, 14, 1763–1773. [Google Scholar] [CrossRef]
- Hendig, D.; Langmann, T.; Kocken, S.; Zarbock, R.; Szliska, C.; Schmitz, G.; Kleesiek, K.; Götting, C. Gene expression profiling of ABC transporters in dermal fibroblasts of pseudoxanthoma elasticum patients identifies new candidates involved in PXE pathogenesis. Lab. Investig. 2008, 88, 1303–1315. [Google Scholar] [CrossRef]
- Scott, K.A.; Wood, E.J.; Karran, E.H. A matrix metalloproteinase inhibitor which prevents fibroblast-mediated collagen lattice contraction. FEBS Lett. 1998, 441, 137–140. [Google Scholar] [CrossRef]
- Lugo-Cintrón, K.M.; Gong, M.M.; Ayuso, J.M.; Tomko, L.A.; Beebe, D.J.; Virumbrales-Muñoz, M.; Ponik, S.M. Breast fibroblasts and ECM components modulate breast cancer cell migration through the secretion of MMPs in a 3D microfluidic co-culture model. Cancers 2020, 12, 1173. [Google Scholar] [CrossRef]
- Neidhart, M.; Seemayer, C.A.; Hummel, K.M.; Michel, B.A.; Gay, R.E.; Gay, S. Functional characterization of adherent synovial fluid cells in rheumatoid arthritis: Destructive potential in vitro and in vivo. Arthritis Rheum. 2003, 48, 1873–1880. [Google Scholar] [CrossRef]
- Dabisch-Ruthe, M. Charakterisierung des Pyrophosphatmetabolismus bei pseudoxanthoma elasticum; Universität Bielefeld: Bielefeld, Germany, 2014. [Google Scholar]
- Takashima, A. Establishment of fibroblast cultures. Curr. Protoc. Cell. Biol. 1998, 2.1, 2.1.1–2.1.12. [Google Scholar] [CrossRef] [PubMed]
- Gregory, C.A.; Grady Gunn, W.; Peister, A.; Prockop, D.J. An alizarin red-based assay of mineralization by adherent cells in culture: Comparison with Cetylpyridinium chloride extraction. Anal. Biochem. 2004, 329, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Lau, W.M.; Doucet, M.; Stadel, R.; Huang, D.; Weber, K.L.; Kominsky, S.L. Enpp1: A potential facilitator of breast cancer bone metastasis. PLoS ONE 2013, 8, e66752. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Faghankhani, M.; Cao, Y.; Uitto, J.; Li, Q. Molecular genetics and modifier genes in pseudoxanthoma elasticum, a heritable multisystem ectopic mineralization disorder. J. Investig. Dermatol. 2021, 141, 1148–1156. [Google Scholar] [CrossRef]
- Pomozi, V.; Le Saux, O.; Brampton, C.; Apana, A.; Iliás, A.; Szeri, F.; Martin, L.; Monostory, K.; Paku, S.; Sarkadi, B.; et al. ABCC6 is a basolateral plasma membrane protein. Circ. Res. 2013, 112, e148–e151. [Google Scholar] [CrossRef]
- Klement, J.F.; Matsuzaki, Y.; Jiang, Q.-J.; Terlizzi, J.; Choi, H.Y.; Fujimoto, N.; Li, K.; Pulkkinen, L.; Birk, D.E.; Sundberg, J.P.; et al. Targeted ablation of the Abcc6 gene results in ectopic mineralization of connective tissues. Mol. Cell. Biol. 2005, 25, 8299–8310. [Google Scholar] [CrossRef]
- Li, Q.; van de Wetering, K.; Uitto, J. Pseudoxanthoma elasticum as a paradigm of heritable ectopic mineralization disorders: Pathomechanisms and treatment development. Am. J. Pathol. 2019, 189, 216–225. [Google Scholar] [CrossRef]
- Quaglino, D.; Boraldi, F.; Barbieri, D.; Croce, A.; Tiozzo, R.; Pasquali Ronchetti, I. Abnormal phenotype of in vitro dermal fibroblasts from patients with pseudoxanthoma elasticum (PXE). Biochim. Biophys. Acta Mol. Basis Dis. 2000, 1501, 51–62. [Google Scholar] [CrossRef]
- Brampton, C.; Pomozi, V.; Chen, L.-H.; Apana, A.; McCurdy, S.; Zoll, J.; Boisvert, W.A.; Lambert, G.; Henrion, D.; Blanchard, S.; et al. ABCC6 deficiency promotes dyslipidemia and atherosclerosis. Sci. Rep. 2021, 11, 3881. [Google Scholar] [CrossRef]
- Plümers, R.; Osterhage, M.R.; Lindenkamp, C.; Knabbe, C.; Hendig, D. Targeting ABCC6 in mesenchymal stem cells: Impairment of mature adipocyte lipid homeostasis. Int. J. Mol. Sci. 2022, 23, 9218. [Google Scholar] [CrossRef]
- Tiemann, J.; Wagner, T.; Vanakker, O.M.; van Gils, M.; Cabrera, J.-L.B.; Ibold, B.; Faust, I.; Knabbe, C.; Hendig, D. Cellular and molecular biomarkers indicate premature aging in pseudoxanthoma elasticum patients. Aging Dis. 2019, 11, 536–546. [Google Scholar] [CrossRef] [PubMed]
- Miglionico, R.; Ostuni, A.; Armentano, M.F.; Milella, L.; Crescenzi, E.; Carmosino, M.; Bisaccia, F. ABCC6 knockdown in HepG2 cells induces a senescent-like cell phenotype. Cell. Mol. Biol. Lett. 2017, 22, 7. [Google Scholar] [CrossRef]
- Zarbock, R.; Hendig, D.; Szliska, C.; Kleesiek, K.; Götting, C. Analysis of MMP2 promoter polymorphisms in patients with pseudoxanthoma elasticum. Clin. Chim. Acta 2010, 411, 1487–1490. [Google Scholar] [CrossRef] [PubMed]
- Pendás, A.M.; Santamaría, I.; Alvarez, M.V.; Pritchard, M.; López-Otín, C. Fine physical mapping of the human matrix metalloproteinase genes clustered on chromosome 11q22.3. Genomics 1996, 37, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Butler, G.S.; Overall, C.M. Updated Biological Roles for Matrix Metalloproteinases and New “Intracellular” Substrates Revealed by Degradomics. Biochemistry 2009, 48, 10830–10845. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Boyd, D.D. Regulation of matrix metalloproteinase gene expression. J. Cell. Physiol. 2007, 211, 19–26. [Google Scholar] [CrossRef]
- Angel, P.; Szabowski, A.; Schorpp-Kistner, M. Function and regulation of AP-1 subunits in skin physiology and pathology. Oncogene 2001, 20, 2413–2423. [Google Scholar] [CrossRef]
- Benbow, U.; Brinckerhoff, C.E. The AP-1 site and MMP gene regulation: What is all the fuss about? Matrix Biol. 1997, 15, 519–526. [Google Scholar] [CrossRef]
- Liacini, A.; Sylvester, J.; Li, W.Q.; Zafarullah, M. Inhibition of interleukin-1-stimulated MAP kinases, activating protein-1 (AP-1) and nuclear factor kappa B (NF-ΚB) transcription factors down-regulates matrix metalloproteinase gene expression in articular chondrocytes. Matrix Biol. 2002, 21, 251–262. [Google Scholar] [CrossRef]
- De Vilder, E.Y.G.; Martin, L.; Lefthériotis, G.; Coucke, P.; Van Nieuwerburgh, F.; Vanakker, O.M. Rare modifier variants alter the severity of cardiovascular disease in pseudoxanthoma elasticum: Identification of novel candidate modifier genes and disease pathways through mixture of effects analysis. Front. Cell. Dev. Biol. 2021, 9, 612581. [Google Scholar] [CrossRef]
- Kaden, J.J.; Dempfle, C.-E.; Grobholz, R.; Tran, H.-T.; Kılıç, R.; Sarıkoç, A.; Brueckmann, M.; Vahl, C.; Hagl, S.; Haase, K.K.; et al. Interleukin-1 beta promotes matrix metalloproteinase expression and cell proliferation in calcific aortic valve stenosis. Atherosclerosis 2003, 170, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Roach, H.I.; Yamada, N.; Cheung, K.S.C.; Tilley, S.; Clarke, N.M.P.; Oreffo, R.O.C.; Kokubun, S.; Bronner, F. Association between the abnormal expression of matrix-degrading enzymes by human osteoarthritic chondrocytes and demethylation of specific CpG sites in the promoter regions. Arthritis Rheum. 2005, 52, 3110–3124. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Corriere, M.A.; Matrisian, L.M.; Guzman, R.J. Matrix metalloproteinase inhibition attenuates aortic calcification. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 1510–1516. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.X.; O’Neill, K.D.; Chen, X.; Kiattisunthorn, K.; Gattone, V.H.; Moe, S.M. Activation of arterial matrix metalloproteinases leads to vascular calcification in chronic kidney disease. Am. J. Nephrol. 2011, 34, 211–219. [Google Scholar] [CrossRef]
- Basalyga, D.M.; Simionescu, D.T.; Xiong, W.; Baxter, B.T.; Starcher, B.C.; Vyavahare, N.R. Elastin degradation and calcification in an abdominal aorta injury model. Circulation 2004, 110, 3480–3487. [Google Scholar] [CrossRef]
- Nollet, L.; Van Gils, M.; Willaert, A.; Coucke, P.J.; Vanakker, O.M. Minocycline attenuates excessive DNA damage response and reduces ectopic calcification in pseudoxanthoma elasticum. J. Investig. Dermatol. 2022, 142, 1629–1638.e6. [Google Scholar] [CrossRef]
- Sparano, J.A.; Bernardo, P.; Stephenson, P.; Gradishar, W.J.; Ingle, J.N.; Zucker, S.; Davidson, N.E. Randomized phase III trial of Marimastat versus placebo in patients with metastatic breast cancer who have responding or stable disease after first-line chemotherapy: Eastern Cooperative Oncology Group Trial E2196. J. Clin. Oncol. 2004, 22, 4683–4690. [Google Scholar] [CrossRef]
- Rasmussen, H.S.; McCann, P.P. Matrix metalloproteinase inhibition as a novel anticancer strategy: A review with special focus on Batimastat and Marimastat. Pharmacol. Ther. 1997, 75, 69–75. [Google Scholar] [CrossRef]
- Durham, A.L.; Speer, M.Y.; Scatena, M.; Giachelli, C.M.; Shanahan, C.M. Role of Smooth Muscle Cells in Vascular Calcification: Implications in Atherosclerosis and Arterial Stiffness. Cardiovascular Research 2018, 114, 590–600. [Google Scholar] [CrossRef]
- Johnson, J.L.; Dwivedi, A.; Somerville, M.; George, S.J.; Newby, A.C. Matrix Metalloproteinase (MMP)-3 Activates MMP-9 Mediated Vascular Smooth Muscle Cell Migration and Neointima Formation in Mice. Arterioscler. Thromb. Vasc. Biol. 2011, 31, e35–e44. [Google Scholar] [CrossRef]
- Hendig, D.; Arndt, M.; Szliska, C.; Kleesiek, K.; Götting, C. SPP1 Promoter polymorphisms: Identification of the first modifier gene for pseudoxanthoma elasticum. Clin. Chem. 2007, 53, 829–836. [Google Scholar] [CrossRef] [PubMed]
- Ronchetti, I.; Boraldi, F.; Annovi, G.; Cianciulli, P.; Quaglino, D. Fibroblast involvement in soft connective tissue calcification. Front. Genet. 2013, 4, 22. [Google Scholar] [CrossRef] [PubMed]
- Sage, A.P.; Tintut, Y.; Demer, L.L. Regulatory mechanisms in vascular calcification. Nat. Rev. Cardiol. 2010, 7, 528–536. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).