

Distinguishing Between Embryonic Provisioning Strategies in Teleost Fishes Using a Threshold Value for Parentotrophy


Abstract
1. Introduction
2. Teleost Parentotrophy Indices: Methodology and Provisioning Strategy Classification
2.1. Methodological Sources of Variation in Parentotrophy Indices
2.1.1. Raw vs. Regression Estimated Means
2.1.2. Embryonic Staging
2.1.3. Variation in Maternal Size
2.1.4. Source and Preservation of Samples

{kind=link}
| Species | MI/PI | Real v Estimated | Parentotrophy Threshold | Statistical Test for Significant Difference | Classification of Nutrient Provisioning Strategy | Resource |
|---|---|---|---|---|---|---|
| Viviparous | ||||||
| Alfaro huberi | 0.64 | EEN | >1 | NA | Lecithotrophy | [72] P |
| Belonesox belizanus | 0.70 | EEN | >1 | NA | Lecithotrophy | * [72] P |
| Brachyrhaphis episcopi | 0.78 | R | Not stated | NA | Not specified | [73] PF |
| Brachyrhaphis holdridgei | 0.66 | EEN | >1 | NA | Lecithotrophy | [72] P |
| Brachyrhaphis rhabdophora | 0.77 | EEN | >1 | NA | Lecithotrophy | * [72] P |
| Dermogenys burmanica | 0.67 | EEN | >0.7 | 0.7 | Lecithotrophy | [49] PE |
| Dermogenys siamensis | 0.64 | EEN | >0.7 | 0.7 | Lecithotrophy | [49] PE |
| Gambusia affinis | 0.62 | EEN | >1 | NA | Lecithotrophy | * [72] P |
| Gambusia aurata | 0.82 | EEN | ≥0.8 | NA | Matrotrophy | [70] PE |
| Gambusia holbrooki | 0.70 | R | Not stated | NA | Not specified | ** [42] |
| 0.64 | EEN | >1 | NA | Lecithotrophy | * [72] P | |
| Gambusia hubbsi | 0.86 | EN | >0.7 | 0.7 | Both | ** [74] PE |
| Gambusia punctata | 0.78 | EEN | >0.7 | 0.7 | Lecithotrophy | [75] P |
| Gambusia sexradiata | 0.73 | EEN | >0.7 | 0.7 | Lecithotrophy | [75] P |
| Gambusia vittata | 0.77 | EEN | >0.7 | 0.7 | Lecithotrophy | ** [75] P |
| 0.74 | EEN | >1 | NA | Lecithotrophy | [72] P | |
| 1.29 | EEN | ≥1 | NA | Both | ** [41] PE | |
| Gambusia wrayi | 0.70 | EEN | >0.7 | 0.7 | Lecithotrophy | [75] P |
| Hemirhamphodon kapuasensis | 0.61 | EEN | >0.7 | 0.7 | Lecithotrophy | [49] PE |
| Hemirhamphodon pogonognathus | 0.64 | EEN | >0.7 | 0.7 | Lecithotrophy | [49] PE |
| Heterophallus milleri | 0.74 | EEN | >0.75 | NA | Lecithotrophy | [48] PF |
| Hippocampus abdominalisp | 1 | R | >0.7 | Stage | Patrotrophy | [3] F |
| Hippocampus fuscusp | 0.75 | R | Not stated | NA | Not specified | [54] F |
| Limia dominicensis | 0.65 | EEN | ≥1 | NA | Lecithotrophy | [76] P |
| 0.51 | EEN | ≥1 | NA | Lecithotrophy | [77] N | |
| Limia heterandria | 0.67 | EEN | ≥1 | NA | Lecithotrophy | [77] N |
| Limia melanogaster | 0.71 | EEN | ≥1 | NA | Lecithotrophy | ** [77] N |
| 0.67 | EEN | ≥1 | NA | Lecithotrophy | [76] P | |
| Limia melanonotata | 0.67 | EEN | ≥1 | NA | Lecithotrophy | [77] N |
| Limia nigrofasciata | 0.64 | EEN | ≥1 | NA | Lecithotrophy | ** [77] N |
| Limia pauciradiata | 0.66 | EEN | ≥1 | NA | Lecithotrophy | [77] N |
| Limia perugiae | 0.90 | EEN | ≥1 | NA | Lecithotrophy | [77] N |
| Limia tridens | 0.90 | EEN | ≥1 | NA | Lecithotrophy | [77] N |
| Limia versicolor | 0.74 | EEN | ≥1 | NA | Lecithotrophy | [77] N |
| Limia vittata | 0.76 | EEN | ≥1 | NA | Lecithotrophy | ** [77] N |
| Limia zonata | 0.91 | EEN | ≥1 | NA | Lecithotrophy | [77] N |
| Micropoecilia picta | 0.78 | EEN | >0.7 | 0.7 | Lecithotrophy | ** [78] PF |
| Nomorhamphus kolonodalensis | 0.66 | EEN | >0.7 | 0.7 | Lecithotrophy | [49] PE |
| Nomorhamphus megarrhamphus | 0.84 | EEN | >0.7 | 0.7 | Lecithotrophy | [49] PE |
| Nomorhamphus weberi | 0.77 | EEN | >0.7 | 0.7 | Lecithotrophy | [49] PE |
| Phallichthys fairweatheri | 0.65 | EEN | ≥0.7 | 0.7 | Lecithotrophy | [25] P |
| Phallichthys quadripunctatus | 0.75 | EEN | ≥0.7 | 0.7 | Lecithotrophy | ** [25] N |
| Poecilia caucana | 0.77 | EEN | ≥1 | NA | Lecithotrophy | [76] P |
| Poecilia latipinna | 0.92 | EEN | Not stated | NA | Both | ** [79] PF |
| Poecilia latipunctata | 0.85 | EEN | ≥1 | NA | Lecithotrophy | [80] PF |
| Poecilia mexicana | 0.63 | EN | >1 | NA | Lecithotrophy | [72] P |
| 0.57 | EEN | >0.7 | NA | Lecithotrophy | ** [81] PF | |
| 0.68 | EN | >0.65 | NA | Lecithotrophy | ** [34] PF | |
| Poecilia reticulata | 0.70 | EEN | >0.7 | 0.7 | Lecithotrophy | ** [78] PF |
| Poecilia wingei | 0.84 | EEN | >0.7 | 0.7 | Lecithotrophy | [78] PF |
| Poeciliopsis baenschi | 0.98 | EEN | ≥0.8 | NA | Matrotrophy | [70] PE |
| Poeciliopsis balsas | 1.05 | EN | >0.6 | 0.7 | Lecithotrophy | [23] P |
| Poeciliopsis catemaco | 0.68 | EN | >0.6 | 0.7 | Lecithotrophy | [23] P |
| Poeciliopsis fasciata | 0.81 | EN | >0.6 | 0.7 | Lecithotrophy | [23] P |
| Poeciliopsis gracilis | 0.69 | EN | >0.6 | 0.7 | Lecithotrophy | [23] P |
| 0.84 | EEN | ≥0.8 | NA | Matrotrophy | [70] PE | |
| 0.80 | EEN | ≥1 | NA | Both | ** [82] PE | |
| 0.72 | R | Not stated | NA | Lecithotrophy | [51] PE | |
| Poeciliopsis hnilickai | 0.86 | EN | >0.6 | 0.7 | Lecithotrophy | [23] P |
| Poeciliopsis infans | 0.86 | EN | >0.6 | 0.7 | Lecithotrophy | [23] P |
| 1.05 | EEN | ≥0.8 | NA | Matrotrophy | [70] PE | |
| Poeciliopsis latidens | 0.86 | EN | >0.6 | 0.7 | Matrotrophy | [23] P |
| Poeciliopsis monacha | 0.61 | R | Not stated | NA | Not specified | [43] PE |
| Poeciliopsis scarlli | 0.87 | EN | >0.6 | 0.7 | Lecithotrophy | [23] P |
| Poeciliopsis turrubarensis | 0.66 | EN | >0.6 | 0.7 | Lecithotrophy | [23] P |
| 1.05 | R | Not stated | NA | Matrotrophy | ** [52] PE | |
| Poeciliopsis viriosa | 0.93 | EN | >0.6 | 0.7 | Matrotrophy | [23] P |
| Priapella chamulae | 0.71 | EEN | >0.75 | Stage | Lecithotrophy | [34] PF |
| Priapella intermedia | 1.03 | R | Not stated | NA | Matrotrophy | [51] PE |
| Priapella olmecae | 0.76 | EEN | ≥0.8 | NA | Lecithotrophy | [70] |
| Priapichthys festae | 0.60 | R | >0.65 | NA | Lecithotrophy | [83] PF |
| Pseudoxhiphophorus jonesii | 0.65 | EEN | ≥0.8 | NA | Lecithotrophy | [70] PE |
| Syngnathus schlegelip | 0.71 | R | Not stated | Stage | Patrotrophy | [33] PE |
| Xiphophorus hellerii | 0.61 | EEN | >1 | NA | Lecithotrophy | [72] P |
| Oviparous | ||||||
| Clupea harengus | 0.73 | R | Not stated | NA | Not specified | [84] F |
| Danio rerio | 0.77 | R | Not stated | NA | Not specified | [62] FL |
| Salmo fario | 0.63 | R | Not stated | NA | Not specified | [50] F |
| Salmo salar | 0.70 | R | Not stated | NA | Not specified | [85] F |
| Salmo irideus | 0.62 | R | Not stated | NA | Not specified | [86] F |
| Salvelinus fontinalis | 0.75 | R | Not stated | NA | Not specified | [87] F |
2.2. Methodological Sources of Variation in Provisioning Strategy Classification
2.2.1. Discrepant Use of Threshold Value for Parentotrophy
2.2.2. Variation in Use of Statistical Tests
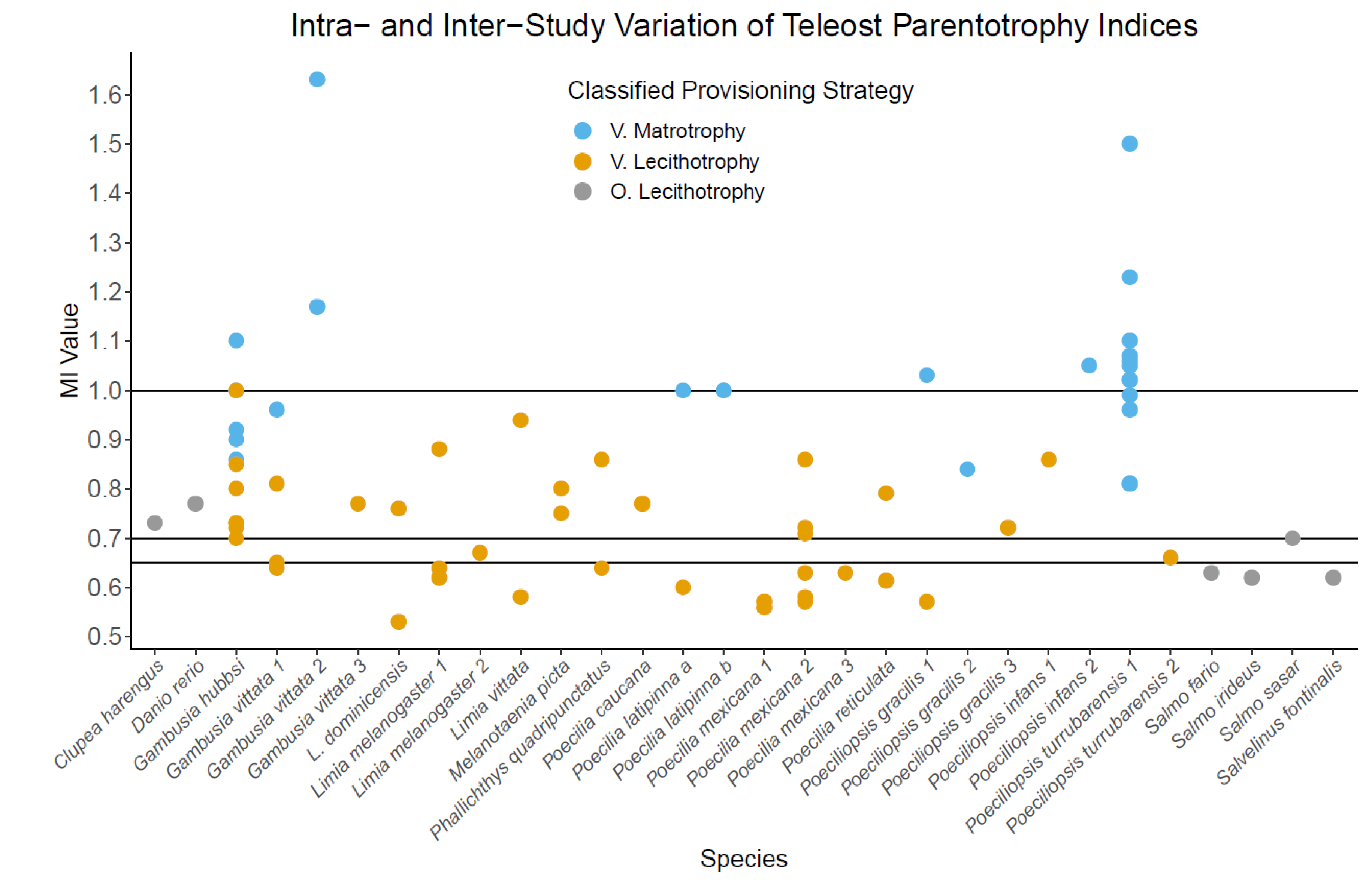
2.2.3. Intra-Specific Variation in Parentotrophy Indices
Intra-Specific Variation in MI/PI Suggests Parentotrophy
Temporal Intra-Specific Variation in Parentotrophy
3. Alternative Approaches to Measuring Parentotrophy
4. Conclusions and Future Directions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blackburn, D.G. Evolution of vertebrate viviparity and specializations for fetal nutrition: A quantitative and qualitative analysis. J. Morphol. 2014, 276, 961–990. [Google Scholar] [CrossRef] [PubMed]
- Wourms, J.P.; Grove, B.D.; Lombardi, J. The Maternal-Embryonic Relationship in Viviparous FishesHoar, W.S., Randall, D.J., Eds.; Fish Physiology1988; Volume 11, pp. 1–34.
- Skalkos, Z.M.G.; Van Dyke, J.U.; Whittington, C.M. Paternal nutrient provisioning during male pregnancy in the seahorse Hippocampus abdominalis. J. Comp. Physiol. B 2020, 190, 547–556. [Google Scholar] [CrossRef]
- Morrison, K.R.; Ngo, V.; Cardullo, A.R.; Reznick, D.N. How fish eggs are preadapted for the evolution of matrotrophy. Proc. R. Soc. 2017, 284, 20171342. [Google Scholar] [CrossRef]
- Stewart, J.R. Fetal nutrition in lecithotrophic squamate reptiles: Toward a comprehensive model for evolution of viviparity and placentation. J. Morphol. 2013, 274, 824–843. [Google Scholar] [CrossRef]
- Stewart, J.R.; Thompson, M. Evolution of placentation among squamate reptiles: Recent research and future directions. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2000, 127, 411–431. [Google Scholar] [CrossRef]
- Thompson, M.B.; Speake, B.K. A review of the evolution of viviparity in lizards: Structure, function and physiology of the placenta. J. Comp. Physiol. B 2005, 176, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Silverthorn, D.U. Human Physiology: An Integrated Approach, 8th ed.Pearson Education Limited: Harlow, UK, 2018. [Google Scholar]
- Blackburn, D.G. Convergent evolution of viviparity, matrotrophy, and specializations for fetal nutrition in reptiles and other vertebrates. Am. Zool. 1992, 32, 313–321. [Google Scholar] [CrossRef]
- Wourms, J.P. Viviparity: The maternal-fetal relationship in fishes. Am. Zool. 1981, 21, 473–515. [Google Scholar] [CrossRef]
- Whittington, C.M.; Friesen, C.R. The evolution and physiology of male pregnancy in syngnathid fishes. Biol. Rev. 2020, 95, 1252–1272. [Google Scholar] [CrossRef]
- Ostrovsky, A.N.; Lidgard, S.; Gordon, D.P.; Schwaha, T.; Genikhovich, G.; Ereskovsky, A.V. Matrotrophy and placentation in invertebrates: A new paradigm. Biol. Rev. 2015, 91, 673–711. [Google Scholar] [CrossRef]
- Sutton, F.B.; Wilson, A.B. Parental behavior in fish. In Encyclopedia of Reproduction, 2nd ed.; Skinner, M.K., Ed.; Elsevier Science & Technology: Amsterdam, The Netherlands, 2018; pp. 106–114. [Google Scholar]
- Blackburn, D.G. Classification of vertebrate reproductive modes. Am. Zool. 2000, 22, 371–377. [Google Scholar]
- Stölting, K.N.; Wilson, A.B. Male pregnancy in seahorses and pipefish: Beyond the mammalian model. Bioessays 2007, 29, 884–896. [Google Scholar] [CrossRef] [PubMed]
- Kvarnemo, C.; Mobley, K.B.; Partridge, C.; Jones, A.; Ahnesjö, I. Evidence of paternal nutrient provisioning to embryos in broad-nosed pipefish Syngnathus typhle. J. Fish Biol. 2011, 78, 1725–1737. [Google Scholar] [CrossRef]
- Turner, C.L. Viviparity in teleost fishes. Sci. Mon. 1947, 65, 508–518. [Google Scholar] [PubMed]
- Uribe, M.C.; Cruz, G.D.L.R.; Alarcón, A.G.; Caballero, J.C.C.; Bárcenas, M.G.G. Structures Associated with Oogenesis and Embryonic Development during Intraovarian Gestation in Viviparous teleosts (Poeciliidae). Fishes 2019, 4, 35. [Google Scholar] [CrossRef]
- Uribe, M.C.; Grier, H.J.; De la Rosa Cruz, G.; García Alarcón, A. Modifications in ovarian and testicular morphology associated with viviparity in teleosts. In Reproductive Biology and Phylogeny of Fishes (Agnathans and Bony Fishes); Jamieson, B.G.M., Ed.; CRC Press: Boca Raton, FL, USA, 2009; pp. 99–132. [Google Scholar]
- Blackburn, D.G. The evolution of viviparity and matrotrophy in vertebrates, with special reference to reptiles. Ph.D. Dissertation, Cornell University, New York, NY, USA, 1985. [Google Scholar]
- Knight, F.M.; Lombardi, J.; Wourms, J.P.; Burns, J.R. Follicular placenta and embryonic growth of the viviparous four-eyed fish (Anableps). J. Morphol. 1985, 185, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Hollenberg, F.; Wourms, J.P. Ultrastructure and protein uptake of the embryonic trophotaeniae of four species of goodeid fishes (Teleostei: Atheriniformes). J. Morphol. 1994, 219, 105–129. [Google Scholar] [CrossRef]
- Reznick, D.N.; Mateos, M.; Springer, M.S. Independent origins and rapid evolution of the placenta in the fish genus Poeciliopsis. Science 2002, 298, 1018–1020. [Google Scholar] [CrossRef]
- Pollux, B.; Pires, M.; Banet, A.; Reznick, D. Evolution of placentas in the fish family Poeciliidae: An empirical study of macroevolution. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 271–289. [Google Scholar] [CrossRef]
- Regus, J.U.; Johnson, J.B.; Webb, S.A.; Reznick, D.N. Comparative life histories of fishes in the genus Phallichthys (Pisces: Poeciliidae). J. Fish Biol. 2013, 83, 144–155. [Google Scholar] [CrossRef]
- Scrimshaw, N.S. Embryonic growth in the viviparous Poeciliid, Heterandria formosa. Biol. Bull. 1944, 87, 37–51. [Google Scholar] [CrossRef]
- Balon, E.K. Reproductive guilds of fishes: A proposal and definition. J. Fish. Res. Board Can. 1975, 32, 821–864. [Google Scholar] [CrossRef]
- Lima, A.R.A.; Barletta, M.; Dantas, D.V.; Ramos, J.A.A.; Costa, M.F. Early development of marine catfishes (Ariidae): From mouth brooding to the release of juveniles in nursery habitats. J. Fish Biol. 2013, 82, 1990–2014. [Google Scholar] [CrossRef]
- Whittington, C.M.; Buddle, A.L.; Griffith, O.W.; Carter, A.M. Embryonic specializations for vertebrate placentation. Philosoophical Trans. R. Soc. B Biol. Sci. 2022, 377, 20210261. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.R.; Castillo, R.E. Nutritional provision of the yolk of two species of viviparous reptiles. Physiol. Zool. 1984, 57, 377–383. [Google Scholar] [CrossRef]
- Stewart, J.R.; Thompson, M. A novel pattern of embryonic nutrition in a viviparous reptile. J. Exp. Biol. 1993, 174, 97–108. [Google Scholar] [CrossRef]
- Paiva, R.B.; Neves, A.; Sequeira, V.; Nunes, M.L.; Gordo, L.S.; Bandarra, N. Reproductive strategy of the female deep-water shark birdbeak dogfish, Deania calcea: Lecithotrophy or matrotrophy? J. Mar. Biol. Assoc. U. K. 2011, 92, 387–394. [Google Scholar] [CrossRef]
- Watanabe, S.; Watanabe, Y. Relationship between male size and newborn size in the Seaweed Pipefish, Syngnathus schlegeli. J. Appl. Phycol. 2002, 65, 319–325. [Google Scholar] [CrossRef]
- Riesch, R.; Plath, M.; Schlupp, I. Toxic hydrogen sulfide and dark caves: Life-history adaptations in a livebearing fish (Poecilia mexicana, Poeciliidae). Ecology 2010, 91, 1494–1505. [Google Scholar] [CrossRef]
- Huveneers, C.; Otway, N.M.; Harcourt, R.G.; Ellis, M. Quantification of the maternal-embryonal nutritional relationship of elasmobranchs: Case study of wobbegong sharks (genus Orectolobus). J. Fish Biol. 2011, 78, 1375–1389. [Google Scholar] [CrossRef]
- Boehlert, G.W.; Yamada, J. Rockfishes of the Genus Sebastes: Their Reproduction and Early Life History; Springer: Berlin/Heidelberg, Germany, 1991. [Google Scholar] [CrossRef]
- Edwards, T.M.; Miller, H.D.; Guillette, L.J. Water quality influences reproduction in female Mosquitofish (Gambusia holbrooki) from eight Florida springs. Environ. Health Perspect. 2006, 114, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Frazer, H.A.; Ellis, M.; Huveneers, C. Can a threshold value be used to classify Chondrichthyan reproductive modes: Systematic review and validation using an oviparous species. PLoS ONE 2012, 7, e50196. [Google Scholar] [CrossRef] [PubMed]
- Constantz, G.D. Energetics of viviparity in the Gila topminnow (Pisces: Poeciliidae). Copeia 1980, 1980, 876. [Google Scholar] [CrossRef]
- Schrader, M.; Travis, J. Do embryos influence maternal investment? Evaluating maternal-fetal coadaptation and the potential for parent-offspring conflict in a placental fish. Evolution 2009, 63, 2805–2815. [Google Scholar] [CrossRef]
- Weldele, M.L.; Zúñiga-Vega, J.J.; Johnson, J.B. Life history of Gambusia vittate (Pisces: Poeciliidae). Southwest. Nat. 2014, 59, 449–460. [Google Scholar] [CrossRef]
- Fernandez-Delgado, C.; Rossomanno, S. Reproductive biology of the mosquitofish in a permanent natural lagoon in south-west Spain: Two tactics for one species. J. Fish Biol. 1997, 51, 80–92. [Google Scholar] [CrossRef]
- Thibault, R.E.; Schultz, R.J. Reproductive adaptations among viviparous fishes (Cyprinodontiformes: Poeciliidae). Evolution 1978, 32, 320–333. [Google Scholar] [CrossRef]
- Haynes, J.L. Standardized classification of Poeciliid development for life-history studies. Copeia 1995, 1995, 147. [Google Scholar] [CrossRef]
- Reznick, D. “Grandfather effects”: The genetics of interpopulation differences in offspring size in the Mosquito fish. Evolution 1981, 35, 941–953. [Google Scholar] [CrossRef]
- Gillooly, J.F.; Charnov, E.L.; West, G.B.; Savage, V.M.; Brown, J.H. Effects of size and temperature on developmental time. Nature 2002, 417, 70–73. [Google Scholar] [CrossRef]
- Mason, M.W.; Bertucci, E.M.; Leri, F.M.; Parrott, B.B. Transient copper exposure during embryogenesis and temperature affect developmental rate, survival, and fin regeneration in Japanese Medaka (Oryzias latipes). Environ. Toxicol. Chem. 2022, 41, 748–757. [Google Scholar] [CrossRef] [PubMed]
- Riesch, R.; Colston, T.J.; Joachim, B.L.; Schlupp, I. Natural history and life history of the Grijalva gambusia Heterophallus milleri Radda, 1987 (Teleostei: Poeciliidae). Aqua Int. J. Ichthyol. 2011, 17, 95–102. [Google Scholar]
- Reznick, D.; Hrbek, T.; Caura, S.; De Greef, J.; Roff, D. Life history of Xenodexia ctenolepis: Implications for life history evolution in the family Poeciliidae. Biol. J. Linn. Soc. 2007, 92, 77–85. [Google Scholar] [CrossRef]
- Gray, J. The growth of fish: I. The relationship between embryo and yolk in Salmo fario. J. Exp. Biol. 1926, 4, 215–226. [Google Scholar] [CrossRef]
- Saleh-Subaie, N.; Ramírez-Cruz, G.A.; Zúñiga-Vega, J.J. Examination of the Trexler-DeAngelis Model of maternal provisioning reveals that matrotrophy is costly. Front. Ecol. Evol. 2021, 9, 635. [Google Scholar] [CrossRef]
- Zúñiga-Vega, J.J.; Reznick, D.N.; Johnson, J.B. Habitat predicts reproductive superfetation and body shape in the livebearing fish Poeciliopsis turrubarensis. Oikos 2007, 116, 995–1005. [Google Scholar] [CrossRef]
- Gorini-Pacheco, B.; Zandonà, E.; Mazzoni, R. Predation effects on matrotrophy, superfetation and other life history traits in Phalloceros harpagos. Ecol. Freshw. Fish 2017, 27, 442–452. [Google Scholar] [CrossRef]
- Vincent, A.C.J. Reproductive Ecology of Seahorses. Ph.D. Dissertation, Cambridge University, Cambridge, UK, 1990. [Google Scholar]
- Kindsvater, H.K.; Bonsall, M.B.; Alonzo, S.H. Survival costs of reproduction predict age-dependent variation in maternal investment. J. Evol. Biol. 2011, 24, 2230–2240. [Google Scholar] [CrossRef]
- Kindsvater, H.K.; Rosenthal, G.G.; Alonzo, S.H. Correction: Maternal size and age shape offspring size in a live-bearing fish, Xiphophorus birchmanni. PLoS ONE 2012, 8, e48473. [Google Scholar] [CrossRef]
- Berkeley, S.A.; Chapman, C.; Sogard, S.M. Maternal age as a determinant of larval growth and survival in a marine fish, Sebastes melanops. Ecology 2004, 85, 1258–1264. [Google Scholar] [CrossRef]
- Cheong, R.T.; Henrich, S.; Farr, J.A.; Travis, J. Variation in fecundity and its relationship to body size in a population of the Least Killifish, Heterandria formosa (Pisces: Poeciliidae). Copeia 1984, 1984, 720. [Google Scholar] [CrossRef]
- Hagmayer, A.; Furness, A.I.; Reznick, D.N.; Pollux, B.J.A. Maternal size and body condition predict the amount of post-fertilization maternal provisioning in matrotrophic fish. Ecol. Evol. 2018, 8, 12386–12396. [Google Scholar] [CrossRef]
- Marshall, D.J.; Heppell, S.S.; Munch, S.B.; Warner, R.R. The relationship between maternal phenotype and offspring quality: Do older mothers really produce the best offspring? Ecology 2010, 91, 2862–2873. [Google Scholar] [CrossRef] [PubMed]
- Saleh-Subaie, N.; Johnson, J.B.; Zúñiga-Vega, J.J. Small sizes, big strategies: The relationship between female size, matrotrophy and superfetation throughout the reproductive lives of poeciliid fishes. J. Zool. 2021, 315, 261–275. [Google Scholar] [CrossRef]
- Hachicho, N.; Reithel, S.; Miltner, A.; Heipieper, H.J.; Küster, E.; Luckenbach, T. Body mass parameters, lipid profiles and protein contents of zebrafish embryos and effects of 2,4-Dinitrophenol exposure. PLoS ONE 2015, 10, e0134755. [Google Scholar] [CrossRef] [PubMed]
- Hislop, J.R.G.; Bell, M.A. Observations on the size, dry weight and energy content of the eggs of some demersal fish species from British marine waters. J. Fish Biol. 1987, 31, 1–20. [Google Scholar] [CrossRef]
- Frimpong, E.A.; Henebry, M.L. Short-term effects of Formalin and Ethanol fixation and preservation techniques on weight and size of fish eggs. Trans. Am. Fish. Soc. 2012, 141, 1472–1479. [Google Scholar] [CrossRef]
- Johnston, T.A.; Mathias, J.A. Length reduction and dry weight loss in frozen and formalin-preserved larval walleye, Stizostedion vitreum (Mitchill). Aquac. Res. 1993, 24, 365–371. [Google Scholar] [CrossRef]
- Leist, D.P.; Nettleton, G.S.; Feldhoff, R.C. Determination of lipid loss during aqueous and phase partition fixation using formalin and glutaraldehyde. J. Histochem. Cytochem. 1986, 34, 437–441. [Google Scholar] [CrossRef]
- Yang, F.; Xiang, W.; Sun, X.; Wu, H.; Li, T.; Long, L. A novel lipid extraction method from wet Microalga Picochlorum sp. at room temperature. Mar. Drugs 2014, 12, 1258–1270. [Google Scholar] [CrossRef]
- Arias, A.-L.; Reznick, D. Life History of Phalloceros caudiomaculatus: A novel variation on the theme of livebearing in the family Poeciliidae. Copeia 2000, 2000, 792–798. [Google Scholar] [CrossRef]
- Wetzel, M.A.; Leuchs, H.; Koop, J.H.E. Preservation effects on wet weight, dry weight, and ash-free dry weight biomass estimates of four common estuarine macro-invertebrates: No difference between ethanol and formalin. Helgol. Mar. Res. 2005, 59, 206–213. [Google Scholar] [CrossRef]
- Olivera-Tlahuel, C.; Ossip-Klein, A.G.; Espinosa-Pérez, H.S.; Zúñiga-Vega, J.J. Have superfetation and matrotrophy facilitated the evolution of larger offspring in poeciliid fishes? Biol. J. Linn. Soc. 2015, 116, 787–804. [Google Scholar] [CrossRef]
- Almeida-Silva, P.H.; Mazzoni, R. Life history aspects of Phalloceros anisophallos Lucinda, 2008 (Osteichthyes, Poeciliidae) from Córrego Andorinha, Ilha Grande (RJ, Brazil). Stud. Neotrop. Fauna Environ. 2014, 49, 191–198. [Google Scholar] [CrossRef]
- Pollux, B.J.A.; Meredith, R.W.; Springer, M.S.; Garland, T.; Reznick, D.N. The evolution of the placenta drives a shift in sexual selection in livebearing fish. Nature 2014, 513, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Jennions, M.D.; Telford, S. Life-history phenotypes in populations of Brachyrhaphis episcopi (Poeciliidae) with different predator communities. Oecologia 2002, 132, 44–50. [Google Scholar] [CrossRef]
- Riesch, R.; Martin, R.A.; Langerhans, R.B. Predation’s role in life-history evolution of a livebearing fish and a test of the Trexler-DeAngelis Model of maternal provisioning. Am. Nat. 2013, 181, 78–93. [Google Scholar] [CrossRef]
- Torres-Mejia, R.M. Ecomorphology of Body Shape and Life History in Females of the Genus Gambusia (Poeciliidae) and in Guppies (Poecilia reticulata, Poeciliidae). Ph.D. Thesis, University of California, Berkeley, CA, USA, 2012. [Google Scholar]
- Pires, M.N.; Reznick, D.N. Life-history evolution in the fish genus Poecilia (Poeciliidae: Cyprinodontiformes: Subgenus Pamphorichthys): An evolutionary origin of extensive matrotrophy decoupled from superfetation. Biol. J. Linn. Soc. 2018, 125, 547–560. [Google Scholar] [CrossRef]
- Cohen, S.N.; Regus, J.U.; Reynoso, Y.; Mastro, T.; Reznick, D.N. Comparative life histories of fishes in the subgenus Limia (Pisces: Poeciliidae). J. Fish Biol. 2015, 87, 100–114. [Google Scholar] [CrossRef]
- Pires, M.N.; Arendt, J.; Reznick, D.N. The evolution of placentas and superfetation in the fish genus Poecilia (Cyprinodontiformes: Poeciliidae: Subgenera Micropoecilia and Acanthophacelus). Biol. J. Linn. Soc. 2010, 99, 784–796. [Google Scholar] [CrossRef]
- Trexler, J.C. Variation in the degree of viviparity in the Sailfin Molly, Poecilia latipinna. Copeia 1985, 1985, 999. [Google Scholar] [CrossRef]
- Pires, M.N. The evolution of placentas in Poeciliid fishes. Ph.D. Dissertation, University of California, Berkeley, CA, USA, 2007. [Google Scholar]
- Riesch, R.; Plath, M.; Schlupp, I.; Marsh-Matthews, E. Matrotrophy in the cave molly: An unexpected provisioning strategy in an extreme environment. Evol. Ecol. 2009, 24, 789–801. [Google Scholar] [CrossRef]
- Molina-Moctezuma, A.; Hernández-Rosas, A.L.; Zúñiga-Vega, J.J. Resource availability and its effects on mother to embryo nutrient transfer in two viviparous fish species. J. Exp. Zool. Part A Ecol. Integr. Physiol. 2020, 333, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Reznick, D.; Callahan, H.; Llauredo, R. Maternal effects on offspring quality in Poeciliid fishes. Amer. Zool. 1996, 36, 147–156. [Google Scholar] [CrossRef]
- Paffenhöfer, G.A.; Rosenthal, H. Trockengewicht und Kaloriengehalt sich entwickelnder Heringseier. Helgol. Mar. Res. 1968, 18, 45–52. [Google Scholar] [CrossRef]
- Hayes, F.R.; Armstrong, F.H. Physical changes in the constituent parts of developing salmon eggs. Can. J. Res. 1942, 20d, 99–114. [Google Scholar] [CrossRef]
- Smith, S. Studies in the development of the rainbow trout (Salmo irideus) I. The heat production and nitrogenous excretion. J. Exp. Biol. 1947, 23, 357–378. [Google Scholar] [CrossRef]
- Gortner, R.A. Studies on the chemistry of embryonic growth I. certain changes in the nitrogen ratios of developing trout eggs. J. Am. Chem. Soc. 1913, 35, 632–645. [Google Scholar] [CrossRef]
- Veith, W. Reproduction in the live-bearing teleost Clinus superciliosus. S. Afr. J. Zool. 1979, 14, 208–211. [Google Scholar] [CrossRef]
- López-Rodríguez, N.C.; de Barros, C.M.; Petry, A.C. A macroscopic classification of the embryonic development of the one-sided livebearer Jenynsia multidentata (Teleostei: Anablepidae). Neotrop. Ichthyol. 2017, 15, e160170. [Google Scholar] [CrossRef]
- Zandonà, E.; Kajin, M.; Buckup, P.A.; Amaral, J.R.; Souto-Santos, I.C.A.; Reznick, D.N. Mode of maternal provisioning in the fish genus Phalloceros: A variation on the theme of matrotrophy. Biol. J. Linn. Soc. 2021, 134, 867–878. [Google Scholar] [CrossRef]
- Zúñiga-Vega, J.J.; Suárez-Rodríguez, M.; Espinosa-Pérez, H.; Johnson, J.B. Morphological and reproductive variation among populations of the Pacific molly Poecilia butleri. J. Fish Biol. 2011, 79, 1029–1046. [Google Scholar] [CrossRef]
- Turcotte, M.M.; Pires, M.N.; Vrijenhoek, R.C.; Reznick, D.N. Pre- and post-fertilization maternal provisioning in livebearing fish species and their hybrids (Poeciliidae: Poeciliopsis). Funct. Ecol. 2008, 22, 1118–1124. [Google Scholar] [CrossRef]
- Hagmayer, A.; Furness, A.I.; Reznick, D.N.; Dekker, M.L.; Pollux, B.J.A. Predation risk shapes the degree of placentation in natural populations of live-bearing fish. Ecol. Lett. 2020, 23, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Marsh-Matthews, E.; Deaton, R. Resources and offspring provisioning: A test of the Trexler-DeAngelis model for matrotrophy evolution. Ecology 2006, 87, 3014–3020. [Google Scholar] [CrossRef]
- Trexler, J.C. Resource availability and plasticity in offspring provisioning: Embryo nourishment in Sailfin mollies. Ecology 1997, 78, 1370–1381. [Google Scholar] [CrossRef]
- Stewart, J.R. Facultative Placentotrophy and the Evolution of Squamate Placentation: Quality of Eggs and Neonates in Virginia striatula. Am. Nat. 1989, 133, 111–137. [Google Scholar] [CrossRef]
- Van Dyke, J.U.; Griffith, O.W.; Thompson, M.B. High food abundance permits the evolution of placentotrophy: Evidence from a placental lizard, Pseudemoia entrecasteauxii. Am. Nat. 2014, 184, 198–210. [Google Scholar] [CrossRef]
- Ramírez-Pinilla, M.P. Placental transfer of nutrients during gestation in an Andean population of the highly matrotrophic lizard genus Mabuya (Squamata: Scincidae). Herpetol. Monogr. 2006, 20, 194–204. [Google Scholar] [CrossRef]
- Swain, R.; Jones, S.M. Maternal-fetal transfer of 3H-labelled leucine in the viviparous lizard Niveoscincus metallicus (scincidae: Lygosominae). J. Exp. Zool. 1997, 277, 139–145. [Google Scholar] [CrossRef]
- Van Dyke, J.U.; Beaupre, S.J. Stable isotope tracer reveals that viviparous snakes transport amino acids to offspring during gestation. J. Exp. Biol. 2012, 215, 760–765. [Google Scholar] [CrossRef] [PubMed]
- Haresign, T.W.; Shumway, S. Permeability of the marsupium of the pipefish Syngnathus fuscus to [14C]-alpha amino isobutyric acid. Comp. Biochem. Physiol. Part A Physiol. 1981, 69, 603–604. [Google Scholar] [CrossRef]
- Marsh-Matthews, E. Matrotrophy. In Ecology and Evolution of Fishes; Evans, J., Pilastro, A., Eds.; Chicago Press: Chicago, IL, USA, 2011; pp. 18–22. [Google Scholar]
- Marsh-Matthews, E.; Brooks, M.; Deaton, R.; Tan, H. Effects of maternal and embryo characteristics on post-fertilization provisioning in fishes of the genus Gambusia. Oecologia 2005, 144, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Ripley, J.L.; Foran, C.M. Direct evidence for embryonic uptake of paternally-derived nutrients in two pipefishes (Syngnathidae: Syngnathus spp.). J. Comp. Physiol. B 2008, 179, 325–333. [Google Scholar] [CrossRef]
- Kim, J.; Vallet, J. Secreted and Placental Membrane Forms of Folate-Binding Protein Occur Sequentially During Pregnancy in Swine1. Biol. Reprod. 2004, 71, 1214–1219. [Google Scholar] [CrossRef]

| Species | Matrotrophy Index | Classification of Nutrient Provisioning Strategy | Resource |
|---|---|---|---|
| Ameca splendens | 150.00 | Matrotrophy | [2] |
| Clinus superciliosus | 35.60 | Matrotrophy | [88] |
| Dermogenys bispina | 152.00 | Matrotrophy | [49] |
| Dermogenys orientalis | 18.35 | Matrotrophy | [49] |
| Dermogenys sumatrana | 198.50 | Matrotrophy | [49] |
| Gambusia rhizophorae | 1.24 | Matrotrophy | [75] |
| Gambusia speciosa | 0.45 | Lecithotrophy | [70] |
| Gambusia yucatana | 0.53 | Lecithotrophy | [70] |
| Hemirhamphodon keukenthali | 0.58 | Lecithotrophy | [49] |
| Heterandria formosa | 14.18 | Matrotrophy | [40] |
| Jenysia multidentata | 606.14 | Matrotrophy | [89] |
| Limia caymanensis | 0.57 | Lecithotrophy | [77] |
| Micropoecilia bifurca | 55.06 | Matrotrophy | [78] |
| Micropoecilia branneri | 86.84 | Matrotrophy | [78] |
| Micropoecilia parae | 6.33 | Matrotrophy | [78] |
| Nomorhamphus bakeri | 3.44 | Matrotrophy | [49] |
| Nomorhamphus brembachi | 11.40 | Matrotrophy | [49] |
| Nomorhamphus manifesta | 15.80 | Matrotrophy | [49] |
| Nomorhamphus rossi | 22.00 | Matrotrophy | [49] |
| Phallichthys amates | 0.52 | Lecithotrophy | [25] |
| Phallichthys tico | 0.43 | Lecithotrophy | [25] |
| Phalloceros anisophallos | 2.80 | Matrotrophy | [71] |
| 2.82 | Matrotrophy | [90] | |
| Phalloceros aspilos | 2.50 | Matrotrophy | [90] |
| Phalloceros enneaktinos | 2.43 | Matrotrophy | [90] |
| Phalloceros harpagos | 3.33 | Matrotrophy | [53] |
| 2.52 | Matrotrophy | [90] | |
| Phalloceros leptokeras | 1.52 | Matrotrophy | [90] |
| Phalloceros tupinamba | 2.17 | Matrotrophy | [90] |
| Phalloceros wai | 2.64 | Matrotrophy | [90] |
| Pamphorichthys araguaiensis | 9.62 | Matrotrophy | [76] |
| Pamphorichthys hasemani | 36.37 | Matrotrophy | [76] |
| Pamphorichthys hollandi | 21.29 | Matrotrophy | [76] |
| Pamphorichthys minor | 1.63 | Matrotrophy | [76] |
| Pamphorichthys scalpridens | 16.58 | Matrotrophy | [76] |
| Poecilia butleri | 2.30 | Matrotrophy | [91] |
| Poeciliopsis elongata | 68.90 | Matrotrophy | [23] |
| Poeciliopsis lucida | 1.34 | Matrotrophy | [43] |
| 1.79 | Matrotrophy | [92] | |
| Poeciliopsis occidentalis | 1.12 | Matrotrophy | [23] |
| 1.50 | Matrotrophy | [39] | |
| Poeciliopsis pleurospilus | 0.50 | Lecithotrophy | [70] |
| Poeciliopsis presidionis | 21.5 | Matrotrophy | [23] |
| Poeciliopsis prolifica | 5.40 | Matrotrophy | [23] |
| 7.01 | Matrotrophy | [80] | |
| Poeciliopsis retropinna | 22.39 | Matrotrophy | [93] |
| Poeciliopsis turneri | 41.4 | Matrotrophy | [23] |
| Xenodexia ctenolepis | 3.86 | Matrotrophy | [49] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skalkos, Z.M.G.; Van Dyke, J.U.; Whittington, C.M. Distinguishing Between Embryonic Provisioning Strategies in Teleost Fishes Using a Threshold Value for Parentotrophy. Biomolecules 2023, 13, 166. https://doi.org/10.3390/biom13010166
Skalkos ZMG, Van Dyke JU, Whittington CM. Distinguishing Between Embryonic Provisioning Strategies in Teleost Fishes Using a Threshold Value for Parentotrophy. Biomolecules. 2023; 13(1):166. https://doi.org/10.3390/biom13010166
Chicago/Turabian StyleSkalkos, Zoe M. G., James U. Van Dyke, and Camilla M. Whittington. 2023. "Distinguishing Between Embryonic Provisioning Strategies in Teleost Fishes Using a Threshold Value for Parentotrophy" Biomolecules 13, no. 1: 166. https://doi.org/10.3390/biom13010166
APA StyleSkalkos, Z. M. G., Van Dyke, J. U., & Whittington, C. M. (2023). Distinguishing Between Embryonic Provisioning Strategies in Teleost Fishes Using a Threshold Value for Parentotrophy. Biomolecules, 13(1), 166. https://doi.org/10.3390/biom13010166