

Oviductal Extracellular Vesicles Enhance Porcine In Vitro Embryo Development by Modulating the Embryonic Transcriptome

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
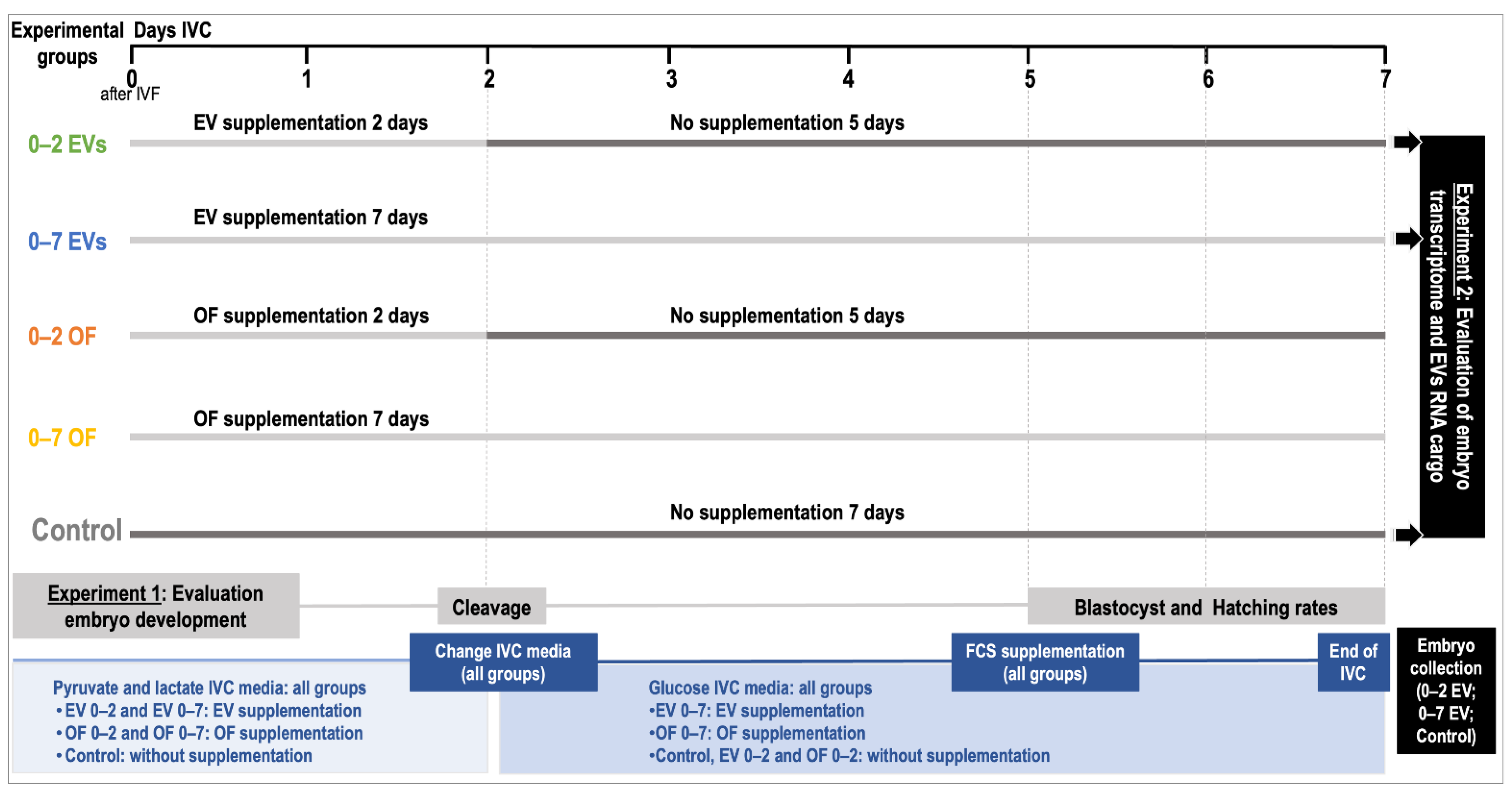
2.1. Experimental Design
2.2. Culture Media
2.3. Oviductal Fluid Collection
2.4. Isolation of Oviductal Extracellular Vesicles
2.5. Transmission Electron Microscopy (TEM)
2.6. Porcine Embryo In Vitro Production
2.6.1. Oocyte Collection and In Vitro Maturation (IVM)
2.6.2. Sperm Preparation and In Vitro Fertilization (IVF)
2.6.3. In Vitro Embryo Culture
2.6.4. Assessment of In Vitro Fertilization Parameters
2.6.5. Assessment of Embryo Development and Embryo Quality
2.7. Statistical Analysis
2.8. Transcriptomic Analysis of poEV Cargo and Embryos by RNA-Sequencing: RNA Isolation, Low-Input Total RNA Library Preparation, Sequencing and Data Analysis
2.9. Quantitative Real-Time Polymerase Chain Reaction (qPCR) Analysis
3. Results
3.1. Transmission Electron Microscopy of Oviductal Extracellular Vesicle Preparations
3.2. Protein Concentration of Oviductal Secretions Used as Supplements in In Vitro Embryo Culture
3.3. In Vitro Embryo Production with Different Oviductal Secretions during In Vitro Culture
3.4. RNA Cargo of poEVs
3.5. Transcriptome Profile of EVs Treated and Control Embryos
3.5.1. Embryonic RNA Profiles
3.5.2. Embryonic miRNA Profiles
3.6. Integration of Embryo and poEVs Datasets
3.6.1. Comparison of RNAs Identified in EVs-Treated Embryos and Contained in poEVs
3.6.2. Comparison of miRNAs Identified in EVs-Treated Embryos and Contained in poEVs
3.6.3. Comparison of Predicted Target Genes of miRNAs Identified in poEVs with RNAs in Embryos
3.6.4. Comparison of Porcine and Bovine oEV RNA Cargo and Their Potential Transcriptomic Alterations in Embryos
3.6.5. Validation of Embryonic Gene Expression Results and EV Transcript Abundance by qPCR of Selected Candidate Genes
4. Discussion
4.1. Effect of poEVs on Embryo Development
4.2. Effect of Sequential System and Oviductal Secretions
4.3. EV RNA Cargo and Their Potential Impact on the Embryo
4.4. Upregulated RNAs in Embryos with Potential poEVs Origin
4.5. Upregulated miRNAs in Embryos with a Potential poEVs Origin
4.6. Downregulated RNAs in Embryo Potentially Affected by miRNAs Derived from poEVs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alminana, C. Snooping on a private conversation between the oviduct and gametes/embryos. Anim. Reprod. 2015, 12, 366–374. [Google Scholar]
- Rizos, D.; Ward, F.; Duffy, P.; Boland, M.P.; Lonergan, P. Consequences of bovine oocyte maturation, fertilization or early embryo development in vitro versus in vivo: Implications for blastocyst yield and blastocyst quality. Mol. Reprod. Dev. 2002, 61, 234–248. [Google Scholar] [CrossRef] [PubMed]
- Romar, R.; Canovas, S.; Matas, C.; Gadea, J.; Coy, P. Pig in vitro fertilization: Where are we and where do we go? Theriogenology 2019, 137, 113–121. [Google Scholar] [CrossRef]
- Coy, P.; Romar, R. In vitro production of pig embryos: A point of view. Reprod. Fertil. Dev. 2002, 14, 275–286. [Google Scholar] [CrossRef]
- Wang, W.H.; Abeydeera, L.R.; Han, Y.M.; Prather, R.S.; Day, B.N. Morphologic evaluation and actin filament distribution in porcine embryos produced in vitro and in vivo. Biol. Reprod. 1999, 60, 1020–1028. [Google Scholar] [CrossRef]
- Perleberg, C.; Kind, A.; Schnieke, A. Genetically engineered pigs as models for human disease. Dis. Model Mech. 2018, 11, dmm030783. [Google Scholar] [CrossRef]
- Ballester, L.; Romero-Aguirregomezcorta, J.; Soriano-Ubeda, C.; Matas, C.; Romar, R.; Coy, P. Timing of oviductal fluid collection, steroid concentrations, and sperm preservation method affect porcine in vitro fertilization efficiency. Fertil. Steril. 2014, 102, 1762–1768.e1. [Google Scholar] [CrossRef]
- Moein-Vaziri, N.; Phillips, I.; Smith, S.; Alminana, C.; Maside, C.; Gil, M.A.; Roca, J.; Martinez, E.A.; Holt, W.V.; Pockley, A.G.; et al. Heat-shock protein A8 restores sperm membrane integrity by increasing plasma membrane fluidity. Reproduction 2014, 147, 719–732. [Google Scholar] [CrossRef]
- Georgiou, A.S.; Gil, M.A.; Alminana, C.; Cuello, C.; Vazquez, J.M.; Roca, J.; Martinez, E.A.; Fazeli, A. Effects of complement component 3 derivatives on pig oocyte maturation, fertilization and early embryo development in vitro. Reprod. Domest. Anim. 2011, 46, 1017–1021. [Google Scholar] [CrossRef]
- Coy, P.; Lloyd, R.; Romar, R.; Satake, N.; Matas, C.; Gadea, J.; Holt, W.V. Effects of porcine pre-ovulatory oviductal fluid on boar sperm function. Theriogenology 2010, 74, 632–642. [Google Scholar] [CrossRef]
- Coy, P.; Canovas, S.; Mondejar, I.; Saavedra, M.D.; Romar, R.; Grullon, L.; Matas, C.; Aviles, M. Oviduct-specific glycoprotein and heparin modulate sperm-zona pellucida interaction during fertilization and contribute to the control of polyspermy. Proc. Natl. Acad. Sci. USA 2008, 105, 15809–15814. [Google Scholar] [CrossRef]
- Alminana, C.; Gil, M.A.; Cuello, C.; Roca, J.; Vazquez, J.M.; Rodriguez-Martinez, H.; Martinez, E.A. Adjustments in IVF system for individual boars: Value of additives and time of sperm-oocyte co-incubation. Theriogenology 2005, 64, 1783–1796. [Google Scholar] [CrossRef]
- Alminana, C.; Gil, M.A.; Cuello, C.; Caballero, I.; Roca, J.; Vazquez, J.M.; Martinez, E.A. In vitro fertilization (IVF) in straws and a short gamete coincubation time improves the efficiency of porcine IVF. Reprod. Domest. Anim. 2008, 43, 747–752. [Google Scholar] [CrossRef]
- Gil, M.A.; Alminana, C.; Cuello, C.; Parrilla, I.; Roca, J.; Vazquez, J.M.; Martinez, E.A. Brief coincubation of gametes in porcine in vitro fertilization: Role of sperm:oocyte ratio and post-coincubation medium. Theriogenology 2007, 67, 620–626. [Google Scholar] [CrossRef]
- Alminana, C.; Gil, M.A.; Cuello, C.; Parrilla, I.; Roca, J.; Vazquez, J.M.; Martinez, E.A. Effects of ultrashort gamete co-incubation time on porcine in vitro fertilization. Anim. Reprod. Sci. 2008, 106, 393–401. [Google Scholar] [CrossRef]
- Cuello, C.; Gomis, J.; Alminana, C.; Maside, C.; Sanchez-Osorio, J.; Gil, M.A.; Sanchez, A.; Parrilla, I.; Vazquez, J.M.; Roca, J.; et al. Effect of MEM vitamins and forskolin on embryo development and vitrification tolerance of in vitro-produced pig embryos. Anim. Reprod. Sci. 2013, 136, 296–302. [Google Scholar] [CrossRef]
- Cheong, S.A.; Kim, E.; Kwak, S.S.; Jeon, Y.; Hyun, S.H. Improvement in the blastocyst quality and efficiency of putative embryonic stem cell line derivation from porcine embryos produced in vitro using a novel culturing system. Mol. Med. Rep. 2015, 12, 2140–2148. [Google Scholar] [CrossRef][Green Version]
- Alminana, C.; Dubuisson, F.; Bauersachs, S.; Royer, E.; Mermillod, P.; Blesbois, E.; Guignot, F. Unveiling how vitrification affects the porcine blastocyst: Clues from a transcriptomic study. J. Anim. Sci. Biotechnol. 2022, 13, 46. [Google Scholar] [CrossRef]
- Van der Weijden, V.A.; Schmidhauser, M.; Kurome, M.; Knubben, J.; Floter, V.L.; Wolf, E.; Ulbrich, S.E. Transcriptome dynamics in early in vivo developing and in vitro produced porcine embryos. BMC Genom. 2021, 22, 139. [Google Scholar] [CrossRef]
- Cambra, J.M.; Martinez, E.A.; Rodriguez-Martinez, H.; Gil, M.A.; Cuello, C. Transcriptional Profiling of Porcine Blastocysts Produced In Vitro in a Chemically Defined Culture Medium. Animals 2021, 11, 1414. [Google Scholar] [CrossRef] [PubMed]
- Alminana, C.; Gil, M.A.; Cuello, C.; Caballero, I.; Roca, J.; Vazquez, J.M.; Gomez, E.; Martinez, E.A. In vitro maturation of porcine oocytes with retinoids improves embryonic development. Reprod. Fertil. Dev. 2008, 20, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Canovas, S.; Ivanova, E.; Romar, R.; Garcia-Martinez, S.; Soriano-Ubeda, C.; Garcia-Vazquez, F.A.; Saadeh, H.; Andrews, S.; Kelsey, G.; Coy, P. DNA methylation and gene expression changes derived from assisted reproductive technologies can be decreased by reproductive fluids. Elife 2017, 6, e23670. [Google Scholar] [CrossRef] [PubMed]
- Alminana, C.; Bauersachs, S. Extracellular Vesicles in the Oviduct: Progress, Challenges and Implications for the Reproductive Success. Bioengineering 2019, 6, 32. [Google Scholar] [CrossRef] [PubMed]
- Alminana, C.; Bauersachs, S. Extracellular vesicles: Multi-signal messengers in the gametes/embryo-oviduct cross-talk. Theriogenology 2020, 150, 59–69. [Google Scholar] [CrossRef]
- Harris, E.A.; Stephens, K.K.; Winuthayanon, W. Extracellular Vesicles and the Oviduct Function. Int. J. Mol. Sci. 2020, 21, 8280. [Google Scholar] [CrossRef]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef]
- Al-Dossary, A.A.; Strehler, E.E.; Martin-Deleon, P.A. Expression and secretion of plasma membrane Ca2+-ATPase 4a (PMCA4a) during murine estrus: Association with oviductal exosomes and uptake in sperm. PLoS ONE 2013, 8, e80181. [Google Scholar] [CrossRef]
- Ferraz, M.; Carothers, A.; Dahal, R.; Noonan, M.J.; Songsasen, N. Oviductal extracellular vesicles interact with the spermatozoon’s head and mid-piece and improves its motility and fertilizing ability in the domestic cat. Sci. Rep. 2019, 9, 9484. [Google Scholar] [CrossRef]
- Alcantara-Neto, A.S.; Fernandez-Rufete, M.; Corbin, E.; Tsikis, G.; Uzbekov, R.; Garanina, A.S.; Coy, P.; Alminana, C.; Mermillod, P. Oviduct fluid extracellular vesicles regulate polyspermy during porcine in vitro fertilisation. Reprod. Fertil. Dev. 2020, 32, 409–418. [Google Scholar] [CrossRef]
- Alcantara-Neto, A.S.; Schmaltz, L.; Caldas, E.; Blache, M.C.; Mermillod, P.; Alminana, C. Porcine oviductal extracellular vesicles interact with gametes and regulate sperm motility and survival. Theriogenology 2020, 155, 240–255. [Google Scholar] [CrossRef]
- Lopera-Vasquez, R.; Hamdi, M.; Maillo, V.; Gutierrez-Adan, A.; Bermejo-Alvarez, P.; Ramirez, M.A.; Yanez-Mo, M.; Rizos, D. Effect of bovine oviductal extracellular vesicles on embryo development and quality in vitro. Reproduction 2017, 153, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Alminana, C.; Corbin, E.; Tsikis, G.; Alcantara-Neto, A.S.; Labas, V.; Reynaud, K.; Galio, L.; Uzbekov, R.; Garanina, A.S.; Druart, X.; et al. Oviduct extracellular vesicles protein content and their role during oviduct-embryo cross-talk. Reproduction 2017, 154, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Bauersachs, S.; Mermillod, P.; Alminana, C. The Oviductal Extracellular Vesicles’ RNA Cargo Regulates the Bovine Embryonic Transcriptome. Int. J. Mol. Sci. 2020, 21, 1303. [Google Scholar] [CrossRef] [PubMed]
- Batista, R.I.; Moro, L.N.; Corbin, E.; Alminana, C.; Souza-Fabjan, J.M.; de Figueiredo Freitas, V.J.; Mermillod, P. Combination of oviduct fluid and heparin to improve monospermic zygotes production during porcine in vitro fertilization. Theriogenology 2016, 86, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Abeydeera, L.R.; Day, B.N. Fertilization and subsequent development in vitro of pig oocytes inseminated in a modified tris-buffered medium with frozen-thawed ejaculated spermatozoa. Biol. Reprod. 1997, 57, 729–734. [Google Scholar] [CrossRef]
- Petters, R.M.; Wells, K.D. Culture of pig embryos. J. Reprod. Fertil. Suppl 1993, 48, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, L.C.; Romar, R.; Aviles, M.; Gadea, J.; Coy, P. Determination of glycosidase activity in porcine oviductal fluid at the different phases of the estrous cycle. Reproduction 2008, 136, 833–842. [Google Scholar] [CrossRef]
- Hafez, E.S.E.; Hafez, B. (Eds.) Folliculogenesis, Egg Maturation, and Ovulation. In Reproduction in Farm Animals; Lippincott, Williams & Wilkins: Baltimore, MD, USA, 2016; pp. 68–81. [Google Scholar]
- Thery, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr. Protoc. Cell Biol. 2006, 30, 3–22. [Google Scholar] [CrossRef]
- Alminana, C.; Tsikis, G.; Labas, V.; Uzbekov, R.; da Silveira, J.C.; Bauersachs, S.; Mermillod, P. Deciphering the oviductal extracellular vesicles content across the estrous cycle: Implications for the gametes-oviduct interactions and the environment of the potential embryo. BMC Genom. 2018, 19, 622. [Google Scholar] [CrossRef]
- Bussiere, J.F.; Bertaud, G.; Guillouet, P. Conservation of boar semen by freezing. Evaluation in vivo and after insemination. Journées Rech. Porc. Fr. 2000, 32, 429–432. Available online: http://www.journees-recherche-porcine.com/texte/2000/00txtGenetique/G0009.pdf (accessed on 19 July 2022).
- Blankenberg, D.; Von Kuster, G.; Coraor, N.; Ananda, G.; Lazarus, R.; Mangan, M.; Nekrutenko, A.; Taylor, J. Galaxy: A web-based genome analysis tool for experimentalists. Curr. Protoc. Mol. Biol. 2010, 89, 19.10.1–19.10.21. [Google Scholar] [CrossRef]
- Sun, J.; Nishiyama, T.; Shimizu, K.; Kadota, K. TCC: An R package for comparing tag count data with robust normalization strategies. BMC Bioinform. 2013, 14, 219. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Licursi, V.; Conte, F.; Fiscon, G.; Paci, P. MIENTURNET: An interactive web tool for microRNA-target enrichment and network-based analysis. BMC Bioinform. 2019, 20, 545. [Google Scholar] [CrossRef]
- Vlachos, I.S.; Zagganas, K.; Paraskevopoulou, M.D.; Georgakilas, G.; Karagkouni, D.; Vergoulis, T.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA-miRPath v3.0: Deciphering microRNA function with experimental support. Nucleic Acids Res. 2015, 43, W460–W466. [Google Scholar] [CrossRef]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef]
- Saeed, A.I.; Sharov, V.; White, J.; Li, J.; Liang, W.; Bhagabati, N.; Braisted, J.; Klapa, M.; Currier, T.; Thiagarajan, M.; et al. TM4: A free, open-source system for microarray data management and analysis. Biotechniques 2003, 34, 374–378. [Google Scholar] [CrossRef]
- Jarrell, V.L.; Day, B.N.; Prather, R.S. The transition from maternal to zygotic control of development occurs during the 4-cell stage in the domestic pig, Sus scrofa: Quantitative and qualitative aspects of protein synthesis. Biol. Reprod. 1991, 44, 62–68. [Google Scholar] [CrossRef]
- Whitten, W.K.; Biggers, J.D. Complete development in vitro of the pre-implantation stages of the mouse in a simple chemically defined medium. J. Reprod. Fertil. 1968, 17, 399–401. [Google Scholar] [CrossRef]
- Petters, R.M. Embryo development in vitro to the blastocyst stage in cattle, pigs and sheep. Anim. Reprod. Sci. 1992, 28, 415–421. [Google Scholar] [CrossRef]
- Braude, P.; Bolton, V.; Moore, S. Human gene expression first occurs between the four- and eight-cell stages of preimplantation development. Nature 1988, 332, 459–461. [Google Scholar] [CrossRef] [PubMed]
- Petters, R.M.; Johnson, B.H.; Reed, M.L.; Archibong, A.E. Glucose, glutamine and inorganic phosphate in early development of the pig embryo in vitro. J. Reprod. Fertil. 1990, 89, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Reed, M.L.; Illera, M.J.; Petters, R.M. Invitro Culture of Pig Embryos. Theriogenology 1992, 37, 95–109. [Google Scholar] [CrossRef]
- Fang, X.; Tanga, B.M.; Bang, S.; Seong, G.; Saadeldin, I.M.; Lee, S.; Cho, J. Oviduct epithelial cells-derived extracellular vesicles improve preimplantation developmental competence of in vitro produced porcine parthenogenetic and cloned embryos. Mol. Reprod. Dev. 2022, 89, 54–65. [Google Scholar] [CrossRef]
- Lopera-Vasquez, R.; Hamdi, M.; Fernandez-Fuertes, B.; Maillo, V.; Beltran-Brena, P.; Calle, A.; Redruello, A.; Lopez-Martin, S.; Gutierrez-Adan, A.; Yanez-Mo, M.; et al. Extracellular Vesicles from BOEC in In Vitro Embryo Development and Quality. PLoS ONE 2016, 11, e0148083. [Google Scholar] [CrossRef]
- Gatien, J.; Mermillod, P.; Tsikis, G.; Bernardi, O.; Idrissi, S.J.; Uzbekov, R.; Le Bourhis, D.; Salvetti, P.; Alminana, C.; Saint-Dizier, M. Metabolomic Profile of Oviductal Extracellular Vesicles across the Estrous Cycle in Cattle. Int. J. Mol. Sci. 2019, 20, 6339. [Google Scholar] [CrossRef]
- Hamdi, M.; Lopera-Vasquez, R.; Maillo, V.; Sanchez-Calabuig, M.J.; Nunez, C.; Gutierrez-Adan, A.; Rizos, D. Bovine oviductal and uterine fluid support in vitro embryo development. Reprod. Fertil. Dev. 2018, 30, 935–945. [Google Scholar] [CrossRef]
- Valinezhad Orang, A.; Safaralizadeh, R.; Kazemzadeh-Bavili, M. Mechanisms of miRNA-Mediated Gene Regulation from Common Downregulation to mRNA-Specific Upregulation. Int. J. Genom. 2014, 2014, 970607. [Google Scholar] [CrossRef]
- Hanaue, M.; Miwa, N.; Uebi, T.; Fukuda, Y.; Katagiri, Y.; Takamatsu, K. Characterization of S100A11, a suppressive factor of fertilization, in the mouse female reproductive tract. Mol. Reprod. Dev. 2011, 78, 91–103. [Google Scholar] [CrossRef]
- Banliat, C.; Tsikis, G.; Labas, V.; Teixeira-Gomes, A.P.; Com, E.; Lavigne, R.; Pineau, C.; Guyonnet, B.; Mermillod, P.; Saint-Dizier, M. Identification of 56 Proteins Involved in Embryo-Maternal Interactions in the Bovine Oviduct. Int. J. Mol. Sci. 2020, 21, 466. [Google Scholar] [CrossRef]
- Teijeiro, J.M.; Ignotz, G.G.; Marini, P.E. Annexin A2 is involved in pig (Sus scrofa)sperm-oviduct interaction. Mol. Reprod. Dev. 2009, 76, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Oh, H.A.; Song, H.; Jun, J.H.; Roh, C.R.; Xie, H.; Dey, S.K.; Lim, H.J. Autophagy regulates embryonic survival during delayed implantation. Endocrinology 2011, 152, 2067–2075. [Google Scholar] [CrossRef] [PubMed]
- Aslund, F.; Beckwith, J. Bridge over troubled waters: Sensing stress by disulfide bond formation. Cell 1999, 96, 751–753. [Google Scholar] [CrossRef]
- De Matos, D.G.; Gasparrini, B.; Pasqualini, S.R.; Thompson, J.G. Effect of glutathione synthesis stimulation during in vitro maturation of ovine oocytes on embryo development and intracellular peroxide content. Theriogenology 2002, 57, 1443–1451. [Google Scholar] [CrossRef]
- Lin, Y.; Zeng, Y.; Zhang, F.; Xue, L.; Huang, Z.; Li, W.; Guo, M. Characterization of microRNA expression profiles and the discovery of novel microRNAs involved in cancer during human embryonic development. PLoS ONE 2013, 8, e69230. [Google Scholar] [CrossRef]
- Kaczmarek, M.M.; Najmula, J.; Guzewska, M.M.; Przygrodzka, E. MiRNAs in the Peri-Implantation Period: Contribution to Embryo-Maternal Communication in Pigs. Int. J. Mol. Sci. 2020, 21, 2229. [Google Scholar] [CrossRef]
- Krawczynski, K.; Bauersachs, S.; Reliszko, Z.P.; Graf, A.; Kaczmarek, M.M. Expression of microRNAs and isomiRs in the porcine endometrium: Implications for gene regulation at the maternal-conceptus interface. BMC Genom. 2015, 16, 906. [Google Scholar] [CrossRef]
- Barranco, I.; Padilla, L.; Martinez, C.A.; Alvarez-Rodriguez, M.; Parrilla, I.; Lucas, X.; Ferreira-Dias, G.; Yeste, M.; Rodriguez-Martinez, H.; Roca, J. Seminal Plasma Modulates miRNA Expression by Sow Genital Tract Lining Explants. Biomolecules 2020, 10, 933. [Google Scholar] [CrossRef]
- Khan, H.L.; Bhatti, S.; Abbas, S.; Kaloglu, C.; Isa, A.M.; Younas, H.; Ziders, R.; Khan, Y.L.; Hassan, Z.; Turhan, B.O.; et al. Extracellular microRNAs: Key players to explore the outcomes of in vitro fertilization. Reprod. Biol. Endocrinol. 2021, 19, 72. [Google Scholar] [CrossRef]
- Kaur, S.; Abu-Shahba, A.G.; Paananen, R.O.; Hongisto, H.; Hiidenmaa, H.; Skottman, H.; Seppanen-Kaijansinkko, R.; Mannerstrom, B. Small non-coding RNA landscape of extracellular vesicles from human stem cells. Sci. Rep. 2018, 8, 15503. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, R.; Palini, S.; Vento, M.E.; La Ferlita, A.; Lo Faro, M.J.; Caroppo, E.; Borzi, P.; Falzone, L.; Barbagallo, D.; Ragusa, M.; et al. Identification of extracellular vesicles and characterization of miRNA expression profiles in human blastocoel fluid. Sci. Rep. 2019, 9, 84. [Google Scholar] [CrossRef]
- Xiao, R.Y.; Zhou, F.Z.; Gui, S. Excessive miR-30a-5p increases the radiosensitivity of hepatoma cells by inhibiting GRP78. Trop. J. Pharm. Res. 2022, 21, 501–506. [Google Scholar] [CrossRef]
- Liu, J.; Huang, Y.Y.; Cai, F.; Dang, Y.; Liu, C.S.; Wang, J.H. MicroRNA-181a regulates endoplasmic reticulum stress in offspring of mice following prenatal microcystin-LR exposure. Chemosphere 2020, 240, 124905. [Google Scholar] [CrossRef]
- Wei, Y.Y.; Tao, X.L.; Xu, H.M.; Chen, Y.; Zhu, L.; Tang, G.Q.; Li, M.Z.; Jiang, A.N.; Shuai, S.R.; Ma, J.D.; et al. Role of miR-181a-5p and endoplasmic reticulum stress in the regulation of myogenic differentiation. Gene 2016, 592, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.Z.; Mao, C.H.; Lee, B.; Lee, A.S. GRP78/BiP is required for cell proliferation and protecting the inner cell mass from apoptosis during early mouse embryonic development. Mol. Cell Biol. 2006, 26, 5688–5697. [Google Scholar] [CrossRef]
- Fradet, S.; Pierredon, S.; Ribaux, P.; Epiney, M.; Shin Ya, K.; Irion, O.; Cohen, M. Involvement of membrane GRP78 in trophoblastic cell fusion. PLoS ONE 2012, 7, e40596. [Google Scholar] [CrossRef]
- Ahn, Y.H.; Park, S.; Choi, J.J.; Park, B.K.; Rhee, K.H.; Kang, E.; Ahn, S.; Lee, C.H.; Lee, J.S.; Inn, K.S.; et al. Secreted tryptophanyl-tRNA synthetase as a primary defence system against infection. Nat. Microbiol. 2016, 2, 16191. [Google Scholar] [CrossRef]
- Nguyen, T.T.T.; Yoon, H.K.; Kim, Y.T.; Choi, Y.H.; Lee, W.K.; Jin, M. Tryptophanyl-tRNA Synthetase 1 Signals Activate TREM-1 via TLR2 and TLR4. Biomolecules 2020, 10, 1283. [Google Scholar] [CrossRef]
- Park, S.R.; Kim, S.R.; Im, J.B.; Lim, S.; Hong, I.S. Tryptophanyl-tRNA Synthetase, a Novel Damage-Induced Cytokine, Significantly Increases the Therapeutic Effects of Endometrial Stem Cells. Mol. Ther. 2020, 28, 2458–2472. [Google Scholar] [CrossRef]
- Hildebrand, D.; Eberle, M.E.; Wolfle, S.M.; Egler, F.; Sahin, D.; Sahr, A.; Bode, K.A.; Heeg, K. Hsa-miR-99b/let-7e/miR-125a Cluster Regulates Pathogen Recognition Receptor-Stimulated Suppressive Antigen-Presenting Cells. Front. Immunol. 2018, 9, 1224. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.E.; Kemper, J.K. Regulation of SIRT1 by microRNAs. Mol. Cells 2013, 36, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Gebremedhn, S.; Ali, A.; Hossain, M.; Hoelker, M.; Salilew-Wondim, D.; Anthony, R.V.; Tesfaye, D. MicroRNA-Mediated Gene Regulatory Mechanisms in Mammalian Female Reproductive Health. Int. J. Mol. Sci. 2021, 22, 938. [Google Scholar] [CrossRef] [PubMed]
- Iljas, J.D.; Wei, Z.; Homer, H.A. Sirt1 sustains female fertility by slowing age-related decline in oocyte quality required for post-fertilization embryo development. Aging Cell 2020, 19, e13204. [Google Scholar] [CrossRef] [PubMed]
- Owczarz, M.; Budzinska, M.; Domaszewska-Szostek, A.; Borkowska, J.; Polosak, J.; Gewartowska, M.; Slusarczyk, P.; Puzianowska-Kuznicka, M. miR-34a and miR-9 are overexpressed and SIRT genes are downregulated in peripheral blood mononuclear cells of aging humans. Exp. Biol. Med. 2017, 242, 1453–1461. [Google Scholar] [CrossRef]
- Saunders, L.R.; Sharma, A.D.; Tawney, J.; Nakagawa, M.; Okita, K.; Yamanaka, S.; Willenbring, H.; Verdin, E. miRNAs regulate SIRT1 expression during mouse embryonic stem cell differentiation and in adult mouse tissues. Aging 2010, 2, 415–431. [Google Scholar] [CrossRef] [PubMed]
- Paredes, F.; Parra, V.; Torrealba, N.; Navarro-Marquez, M.; Gatica, D.; Bravo-Sagua, R.; Troncoso, R.; Pennanen, C.; Quiroga, C.; Chiong, M.; et al. HERPUD1 protects against oxidative stress-induced apoptosis through downregulation of the inositol 1,4,5-trisphosphate receptor. Free Radic. Biol. Med. 2016, 90, 206–218. [Google Scholar] [CrossRef]
- Dong, Y.F.; Chen, Z.Z.; Zhao, Z.; Yang, D.D.; Yan, H.; Ji, J.; Sun, X.L. Potential role of microRNA-7 in the anti-neuroinflammation effects of nicorandil in astrocytes induced by oxygen-glucose deprivation. J. Neuroinflamm. 2016, 13, 60. [Google Scholar] [CrossRef]
- Hao, L.; Vassena, R.; Wu, G.; Han, Z.; Cheng, Y.; Latham, K.E.; Sapienza, C. The unfolded protein response contributes to preimplantation mouse embryo death in the DDK syndrome. Biol. Reprod. 2009, 80, 944–953. [Google Scholar] [CrossRef]
- Cagnone, G.L.; Sirard, M.A. Transcriptomic signature to oxidative stress exposure at the time of embryonic genome activation in bovine blastocysts. Mol. Reprod. Dev. 2013, 80, 297–314. [Google Scholar] [CrossRef]
- Singh, P.K.; Mehla, K.; Hollingsworth, M.A.; Johnson, K.R. Regulation of Aerobic Glycolysis by microRNAs in Cancer. Mol. Cell Pharmacol 2011, 3, 125–134. [Google Scholar] [PubMed]
- Cagnone, G.; Sirard, M.A. The embryonic stress response to in vitro culture: Insight from genomic analysis. Reproduction 2016, 152, R247–R261. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Ma, J.; Li, J.; Wang, D.; Wang, Z.; Wang, S. Mitochondrial phosphatase PGAM5 modulates cellular senescence by regulating mitochondrial dynamics. Nat. Commun. 2020, 11, 2549. [Google Scholar] [CrossRef] [PubMed]
- Lenhausen, A.M.; Wilkinson, A.S.; Lewis, E.M.; Dailey, K.M.; Scott, A.J.; Khan, S.; Wilkinson, J.C. Apoptosis Inducing Factor Binding Protein PGAM5 Triggers Mitophagic Cell Death That Is Inhibited by the Ubiquitin Ligase Activity of X-Linked Inhibitor of Apoptosis. Biochemistry 2016, 55, 3285–3302. [Google Scholar] [CrossRef]
- Li, C.J.; Lin, L.T.; Tsai, H.W.; Wen, Z.H.; Tsui, K.H. Phosphoglycerate mutase family member 5 maintains oocyte quality via mitochondrial dynamic rearrangement during aging. Aging Cell 2022, 21, e13546. [Google Scholar] [CrossRef]
- Liu, G.; Qian, M.; Chen, M.; Chen, T.; Qin, S. miR-21-5p Suppresses Mitophagy to Alleviate Hyperoxia-Induced Acute Lung Injury by Directly Targeting PGAM5. Biomed. Res. Int. 2020, 2020, 4807254. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequence 5′ to 3′ | Product Length, bp | Annealing Temp, °C | Accession No. | Ssc Gene ID |
|---|---|---|---|---|---|
| S100A11 | F: ATGCTGGAAGGGACGGTAAC | 121 | 64 | NM_001004045.1 | 445534 |
| R: ATCATGCGGTCAAGGACACC | |||||
| ANXA2 | F: CGGCAAGTCCCTGTACAACT | 171 | 60 | NM_001005726.1 | 406192 |
| R: TGCAGTTAGTCGCAAGCTGA | |||||
| GABARAPL1 | F: ACACCATCCCTCCCACTAGC | 99 | 59 | NM_001190287.1 | 100462751 |
| R: AGACACTCTCGTCGCTGTAG | |||||
| HSPA5 | F: GAAACCGTGGGAGGTGTCAT | 146 | 64 | XM_001927795.7 | 407060 |
| R: TCTTTTGTCAGGGGTCGCTC | |||||
| WARS | F: TGGACGTGTCCTTCATGTACC | 132 | 59 | XM_003128728.6 | 100515390 |
| R: GCTGTAGCACCTCTATGAGCA | |||||
| SIRT1 | F: GAACCGATGGAGAGTCCAGG | 196 | 64 | NM_001145750.2 | 751859 |
| R: ATACCTCAGCGCCATGGAAA | |||||
| RN18S | F: ATACATGCCGACGGGCGCTG | 84 | 64 | NR_046261.1 | 100861538 |
| R: GGGAGGAGGCTGACCGGGTT | |||||
| UBB | F:GCAGCTGGAAGATGGCCGCA | 88 | 60 | NM_001105309.1 | 100125968 |
| R: GCAGCCACCCCTCAGACGGA | |||||
| H3-3A | F: CTTCCAGCGTCTGGTGCGGG | 200 | 59 | NM_213930.1 | 396970 |
| R: CTCCACGTATGCGGCGTGCT |
| Treatment During IVC | No. Oocytes | Cleavage Rate Day 2 (%) |
|---|---|---|
| Control | 213 | 44.9 ± 0.8 a |
| OF 0–2 | 220 | 46.6 ± 1.3 ab |
| OF 0–7 | 220 | 45.8 ± 1.3 ab |
| EV 0–2 | 221 | 51.8 ± 2.1 b |
| EV 0–7 | 219 | 50.5 ± 2.1 ab |
| Gene | Technology | Embryo Treatment: EV 0–2 vs. CO | Embryo Treatment: EV 0–7 vs. CO | EV Cargo | ||
|---|---|---|---|---|---|---|
| log FC EV 0-2/CO * | PValue RNA-seq PValue qPCR | log FC EV 0–7/CO * | PValue RNA-seq PValue qPCR | CPM RNA-seq Cq qPCR | ||
| S100A11 | RNA-seq | 0.83 | 0.0004 | 0.79 | 0.0009 | 845.40 |
| qPCR | 4.97 | 0.0022 | 5.89 | 0.0018 | 24.24 | |
| ANXA2 | RNA-seq | 1.03 | 0.0007 | 0.78 | 0.0140 | 665.07 |
| qPCR | 1.87 | 0.0290 | 1.37 | 0.3010 | 23.67 | |
| GABARAPL1 | RNA-seq | 1.01 | 0.0025 | 1.07 | 0.0013 | 128.20 |
| qPCR | 9.75 | 0.0050 | 1.04 | 0.0936 | 27.36 | |
| HSPA5 | RNA-seq | 0.04 | 0.9207 | −1.07 | 0.0076 | 120.76 |
| qPCR | −1.61 | 0.1725 | −1.24 | 0.0537 | 27.70 | |
| WARS | RNA-seq | −0.37 | 0.1955 | −1.47 | 0.0000 | 71.56 |
| qPCR | 1.58 | 0.6052 | −13.17 | 0.0310 | 28.71 | |
| SIRT1 | RNA-seq | −0.41 | 0.3391 | −0.60 | 0.2188 | 28.70 |
| qPCR | −3.88 | 0.1772 | 0.24 | 0.9055 | 31.55 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Alcântara-Neto, A.S.; Cuello, C.; Uzbekov, R.; Bauersachs, S.; Mermillod, P.; Almiñana, C. Oviductal Extracellular Vesicles Enhance Porcine In Vitro Embryo Development by Modulating the Embryonic Transcriptome. Biomolecules 2022, 12, 1300. https://doi.org/10.3390/biom12091300
de Alcântara-Neto AS, Cuello C, Uzbekov R, Bauersachs S, Mermillod P, Almiñana C. Oviductal Extracellular Vesicles Enhance Porcine In Vitro Embryo Development by Modulating the Embryonic Transcriptome. Biomolecules. 2022; 12(9):1300. https://doi.org/10.3390/biom12091300
Chicago/Turabian Stylede Alcântara-Neto, Agostinho Soares, Cristina Cuello, Rustem Uzbekov, Stefan Bauersachs, Pascal Mermillod, and Carmen Almiñana. 2022. "Oviductal Extracellular Vesicles Enhance Porcine In Vitro Embryo Development by Modulating the Embryonic Transcriptome" Biomolecules 12, no. 9: 1300. https://doi.org/10.3390/biom12091300
APA Stylede Alcântara-Neto, A. S., Cuello, C., Uzbekov, R., Bauersachs, S., Mermillod, P., & Almiñana, C. (2022). Oviductal Extracellular Vesicles Enhance Porcine In Vitro Embryo Development by Modulating the Embryonic Transcriptome. Biomolecules, 12(9), 1300. https://doi.org/10.3390/biom12091300