
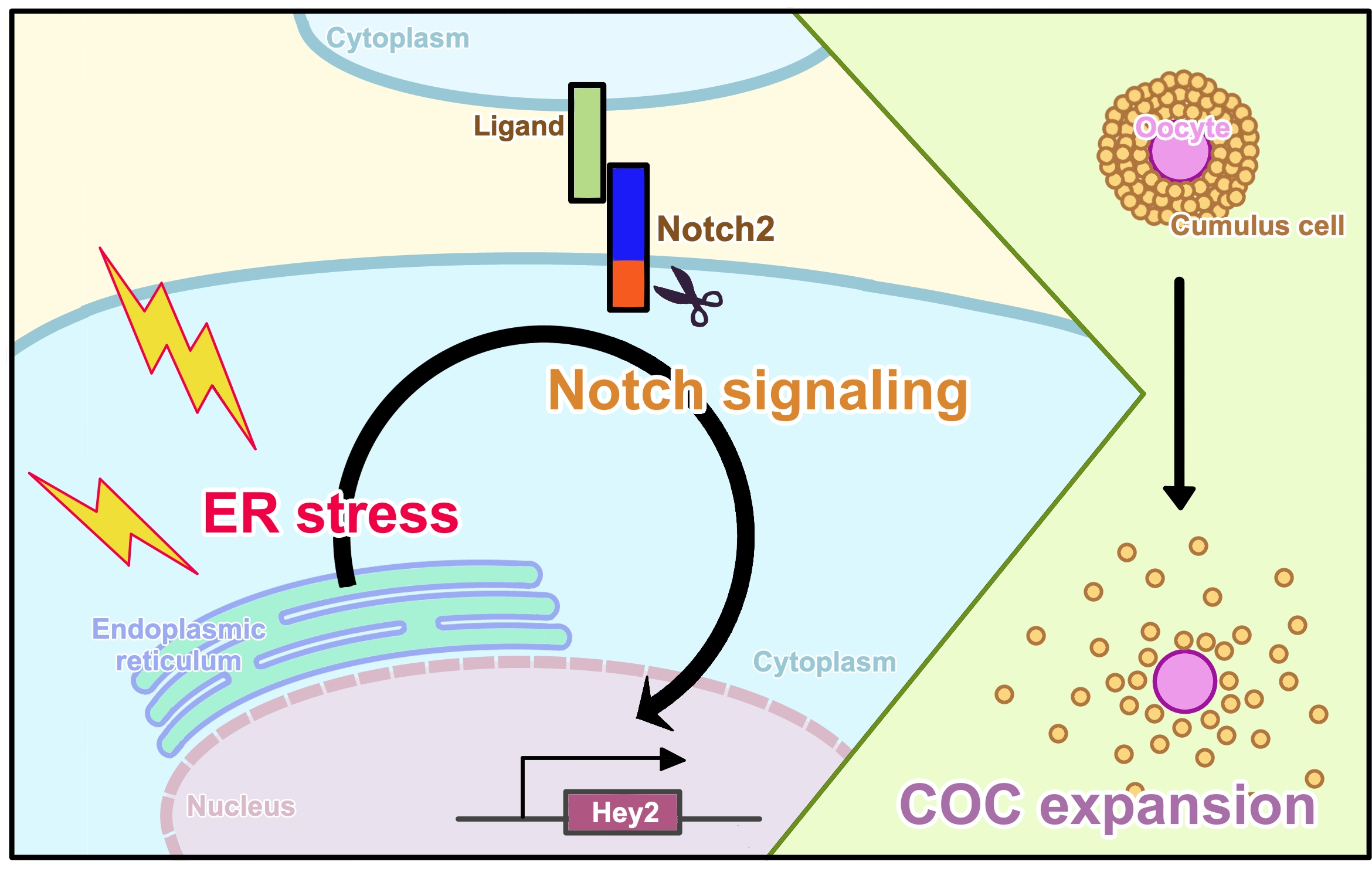
Notch Signaling Induced by Endoplasmic Reticulum Stress Regulates Cumulus-Oocyte Complex Expansion in Polycystic Ovary Syndrome

 ,
,  ,
,  ,
,
Abstract

1. Introduction
2. Materials and Methods
2.1. Human Specimens
2.2. PCOS Animal Model
2.3. Follicle Isolation and COC Culture
2.4. Isolation and Culture of Human GLCs
2.5. Treatment of Human GLCs
2.6. RNA Extraction, Reverse Transcription, and Real-Time Quantitative PCR
2.7. Western Blotting
2.8. Histology and Immunohistochemistry
2.9. Statistical Analysis
3. Results
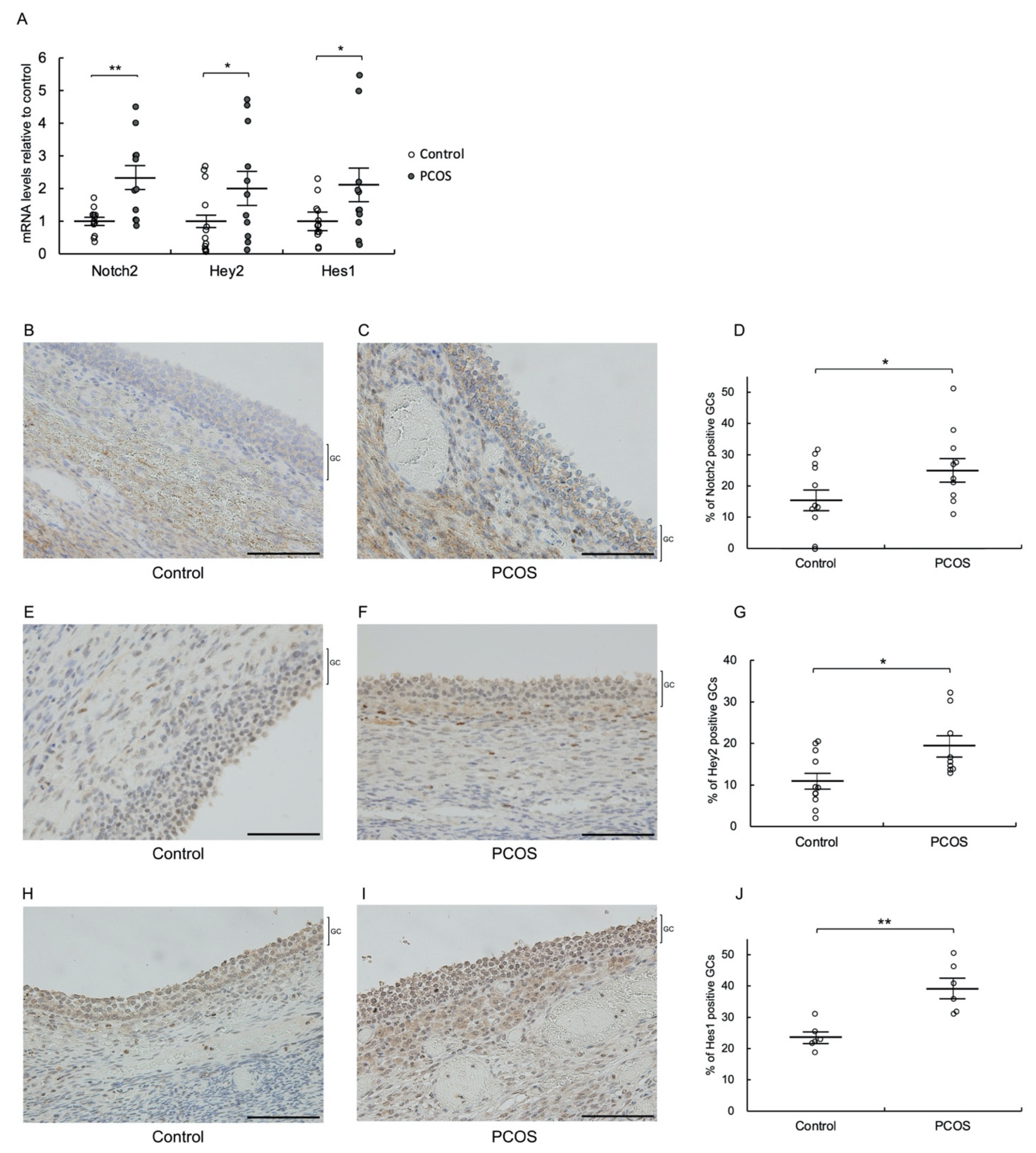
3.1. Notch2, Hey2, and Hes1 Are Upregulated in Granulosa Cells of PCOS Patients
3.2. Notch2, Hey2, and Hes1 Are Upregulated in Granulosa Cells of PCOS Model Mice
3.3. ER Stress Increases Notch2 and Hey2 Expression in Cultured Human GLCs
3.4. ER Stress Induces Notch2 Signaling via the ATF4 Pathway in Cultured Human GLCs
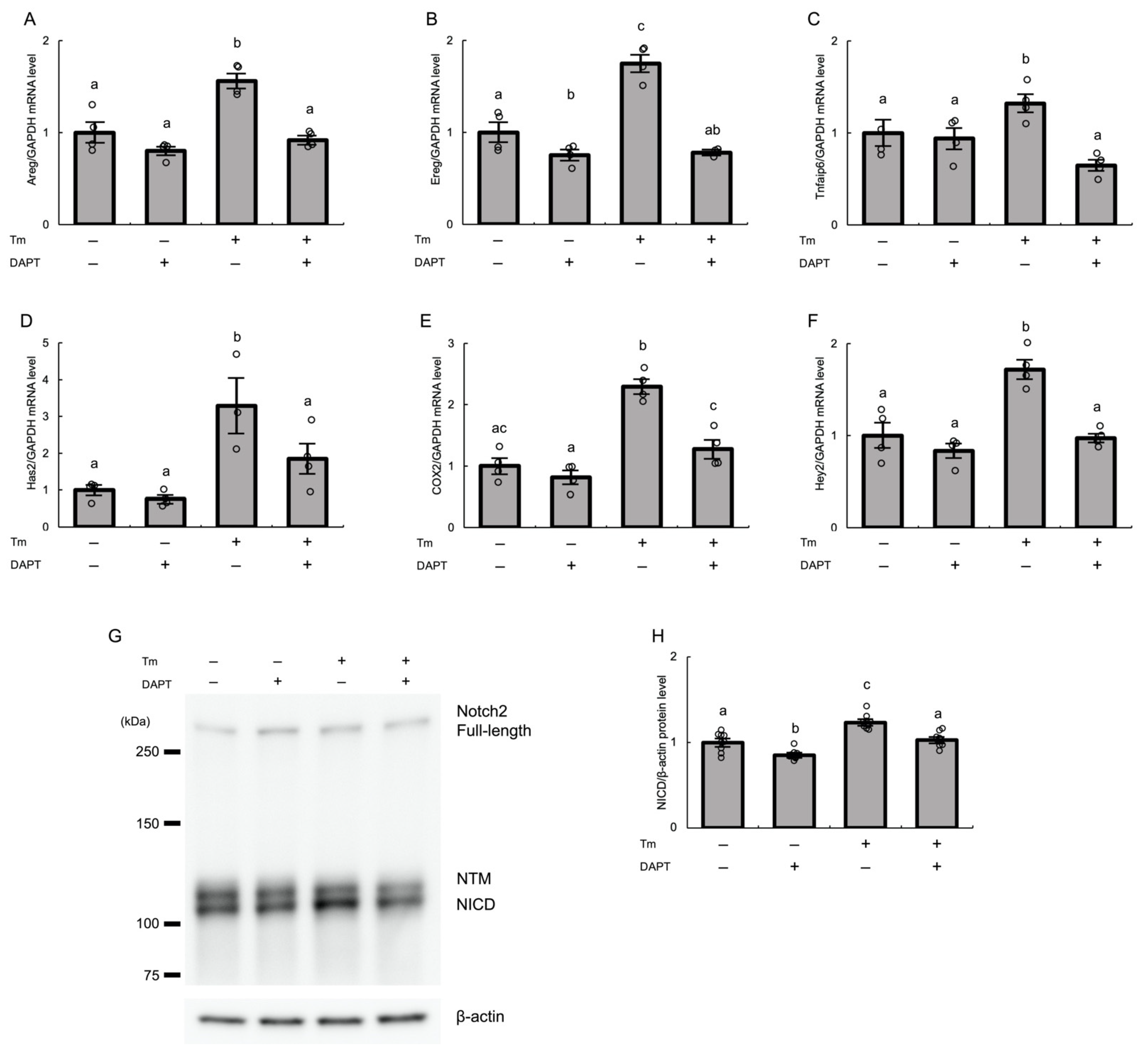
3.5. Notch2 Signaling Mediates ER Stress-Induced Expression of Various Genes Associated with COC Expansion in Cultured Human GLCs
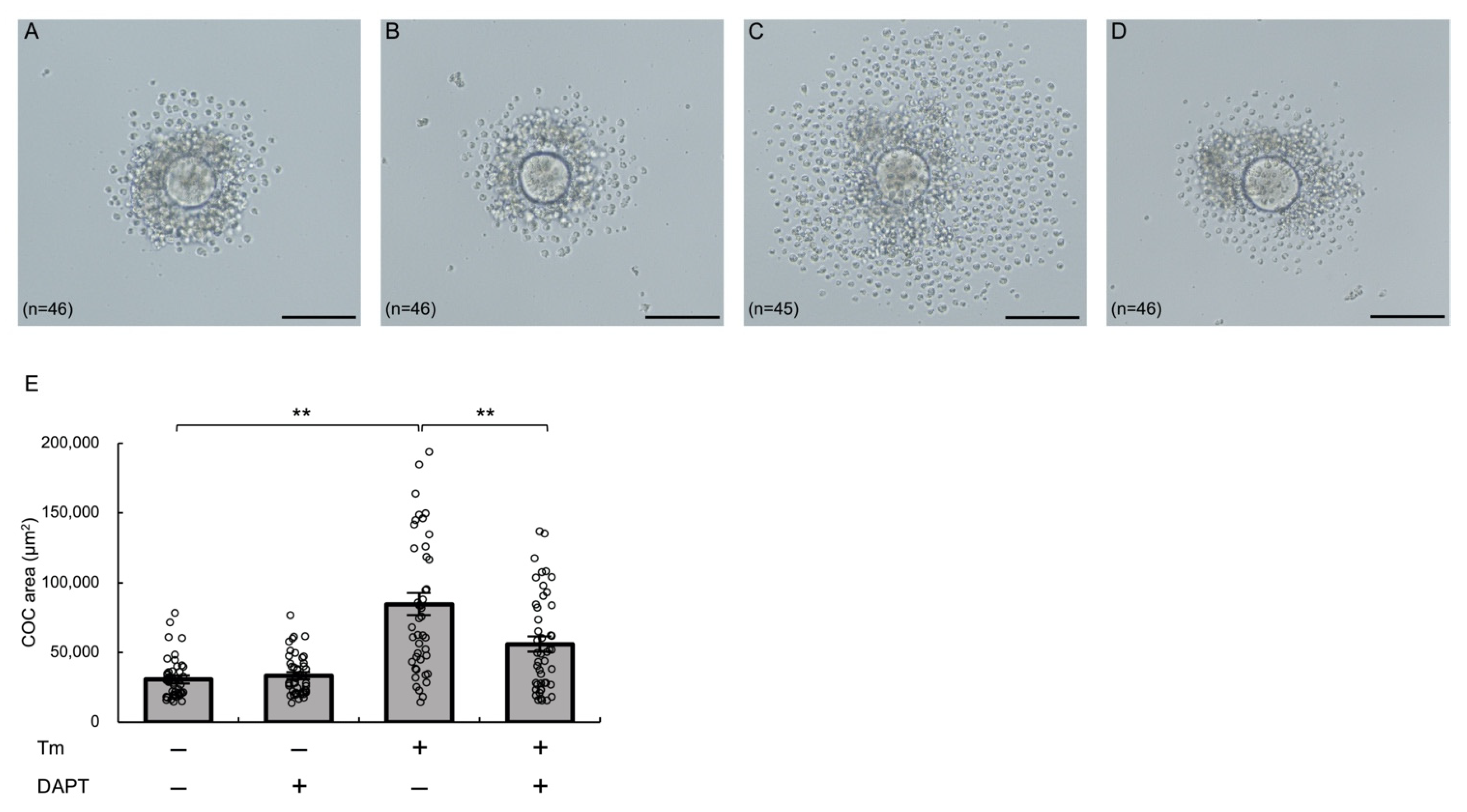
3.6. Notch Signaling Mediates ER Stress-Enhanced Expansion of Cultured Murine COCs
3.7. Notch Signaling Mediates COC Expansion in PCOS Model Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Escobar-Morreale, H.F. Polycystic ovary syndrome: Definition, aetiology, diagnosis and treatment. Nat. Rev. Endocrinol. 2018, 14, 270–284. [Google Scholar] [CrossRef] [PubMed]
- Harada, M.; Takahashi, N.; Azhary, J.M.; Kunitomi, C.; Fujii, T.; Osuga, Y. Endoplasmic reticulum stress: A key regulator of the follicular microenvironment in the ovary. Mol. Hum. Reprod. 2021, 27, gaaa088. [Google Scholar] [CrossRef] [PubMed]
- Rutkowski, D.T.; Kaufman, R.J. That which does not kill me makes me stronger: Adapting to chronic ER stress. Trends Biochem. Sci. 2007, 32, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Hetz, C.; Axten, J.M.; Patterson, J.B. Pharmacological targeting of the unfolded protein response for disease intervention. Nat. Chem. Biol. 2019, 15, 764–775. [Google Scholar] [CrossRef] [PubMed]
- Walter, P.; Ron, D. The unfolded protein response: From stress pathway to homeostatic regulation. Science 2011, 334, 1081–1086. [Google Scholar] [CrossRef] [PubMed]
- Hetz, C.; Zhang, K.; Kaufman, R.J. Mechanisms, regulation and functions of the unfolded protein response. Nat. Rev. Mol. Cell Biol. 2020, 21, 421–438. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Harada, M.; Hirota, Y.; Nose, E.; Azhary, J.M.; Koike, H.; Kunitomi, C.; Yoshino, O.; Izumi, G.; Hirata, T.; et al. Activation of Endoplasmic Reticulum Stress in Granulosa Cells from Patients with Polycystic Ovary Syndrome Contributes to Ovarian Fibrosis. Sci. Rep. 2017, 7, 10824. [Google Scholar] [CrossRef] [PubMed]
- Azhary, J.M.K.; Harada, M.; Takahashi, N.; Nose, E.; Kunitomi, C.; Koike, H.; Hirata, T.; Hirota, Y.; Koga, K.; Wada-Hiraike, O.; et al. 2019 Endoplasmic Reticulum Stress Activated by Androgen Enhances Apoptosis of Granulosa Cells via Induction of Death Receptor 5 in PCOS. Endocrinology 2019, 160, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Azhary, J.M.K.; Harada, M.; Kunitomi, C.; Kusamoto, A.; Takahashi, N.; Nose, E.; Oi, N.; Wada-Hiraike, O.; Urata, Y.; Hirata, T.; et al. 2020 Androgens increase accumulation of advanced glycation end products in granulosa cells by activating ER stress in PCOS. Endocrinology 2020, 161, bqaa015. [Google Scholar] [CrossRef]
- Kunitomi, C.; Harada, M.; Kusamoto, A.; Azhary, J.M.; Nose, E.; Koike, H.; Xu, Z.; Urata, Y.; Takahashi, N.; Wada-Hiraike, O.; et al. Induction of aryl hydrocarbon receptor in granulosa cells by endoplasmic reticulum stress contributes to pathology of polycystic ovary syndrome. Mol. Hum. Reprod. 2021, 27, gaab003. [Google Scholar] [CrossRef] [PubMed]
- Vanorny, D.A.; Mayo, K.E. The role of Notch signaling in the mammalian ovary. Reproduction 2017, 153, R187–R204. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Quan, S.; Zou, S. Roles of the Notch Signaling Pathway in Ovarian Functioning. Reprod. Sci. 2021, 28, 2770–2778. [Google Scholar] [CrossRef]
- Wood, J.R.; Dumesic, D.A.; Abbott, D.H.; Strauss, J.F., 3rd. Molecular Abnormalities in Oocytes from Women with Polycystic Ovary Syndrome Revealed by Microarray Analysis. J. Clin. Endocrinol. Metab. 2007, 92, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Skrtic, A.; Sokolic, L.; Borovecki, A.; Rosa, J.; Fenzl, V. Immunohistochemical localization of CD31, NOTCH1 and JAGGED1 proteins in experimentally induced polycystic ovaries of immature rats. Acta Histochem. 2011, 113, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Archer, K.J.; Devi, M.G.; Kriplani, A.; Strauss, J.F., 3rd; Singh, R. Differential gene expression in granulosa cells from polycystic ovary syndrome patients with and without insulin resistance: Identification of susceptibility gene sets through network analysis. J. Clin. Endocrinol. Metab. 2012, 97, E2016–E2021. [Google Scholar] [CrossRef] [PubMed]
- Haouzi, D.; Assou, S.; Monzo, C.; Vincens, C.; Dechaud, H.; Hamamah, S. Altered gene expression profile in cumulus cells of mature MII oocytes from patients with polycystic ovary syndrome. Hum. Reprod. 2012, 27, 3523–3530. [Google Scholar] [CrossRef]
- Xu, B.; Zhang, Y.W.; Tong, X.H.; Liu, Y.S. Characterization of microRNA profile in human cumulus granulosa cells: Identification of microRNAs that regulate Notch signaling and are associated with PCOS. Mol. Cell. Endocrinol. 2015, 404, 26–36. [Google Scholar] [CrossRef]
- Liu, Q.; Li, Y.; Feng, Y.; Liu, C.; Ma, J.; Li, Y.; Xiang, H.; Ji, Y.; Cao, Y.; Tong, X.; et al. Single-cell analysis of differences in transcriptomic profiles of oocytes and cumulus cells at GV, MI, MII stages from PCOS patients. Sci. Rep. 2016, 6, 39638. [Google Scholar] [CrossRef]
- Masoudi, M.; Yamini, N.; Salehi, F.; Aflatoonian, R.; Kutenaee, M.A.; Esfandiyari, S.; Hosseini, E.; Asadi, E.; Najarkolaei, A.H.; Mortezaee, K.; et al. 2021 Notch signaling pathway in cumulus cells reflecting zygote and embryo quality in polycystic ovary syndrome. Arch. Gynecol. Obstet. 2021, 304, 1097–1105. [Google Scholar] [CrossRef]
- Yang, D.; Li, N.; Ma, A.; Dai, F.; Zheng, Y.; Hu, X.; Wang, Y.; Xian, S.; Zhang, L.; Yuan, M.; et al. Identification of Potential Biomarkers of Polycystic Ovary Syndrome via Integrated Bioinformatics Analysis. Reprod. Sci. 2021, 28, 1353–1361. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yan, Z.; Qin, Q.; Nisenblat, V.; Chang, H.M.; Yu, Y.; Wang, T.; Lu, C.; Yang, M.; Yang, S.; et al. Transcriptome Landscape of Human Folliculogenesis Reveals Oocyte and Granulosa Cell Interactions. Mol. Cell. 2018, 72, 1021–1034.e4. [Google Scholar] [CrossRef] [PubMed]
- Rotterdam ESHRE/ASRM-Sponsored PCOS Consensus Workshop Group. Revised 2003 consensus on diagnostic criteria and long-term health risks related to polycystic ovary syndrome (PCOS). Hum. Reprod. 2004, 19, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Elia, E.; Sander, V.; Luchetti, C.G.; Solano, M.E.; Di Girolamo, G.; Gonzalez, C.; Motta, A.B. The mechanisms involved in the action of metformin in regulating ovarian function in hyperandrogenized mice. Mol. Hum. Reprod. 2006, 12, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Zhu, J.D.; Zhang, X.H.; Long, T.T.; Ge, G.; Yu, Y. Neuroprotective effect of Notch pathway inhibitor DAPT against focal cerebral ischemia/reperfusion 3 hours before model establishment. Neural Regen. Res. 2019, 14, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Körbel, C.; Gerstner, M.D.; Menger, M.D.; Laschke, M.W. Notch signaling controls sprouting angiogenesis of endometriotic lesions. Angiogenesis 2018, 21, 37–46. [Google Scholar] [CrossRef]
- Bertoldo, M.J.; Caldwell, A.S.L.; Riepsamen, A.H.; Lin, D.; Gonzalez, M.B.; Robker, R.L.; Ledger, W.L.; Gilchrist, R.B.; Handelsman, D.J.; Walters, K.A. A Hyperandrogenic Environment Causes Intrinsic Defects That Are Detrimental to Follicular Dynamics in a PCOS Mouse Model. Endocrinology 2019, 160, 699–715. [Google Scholar] [CrossRef]
- Takahashi, N.; Harada, M.; Azhary, J.M.K.; Kunitomi, C.; Nose, E.; Terao, H.; Koike, H.; Wada-Hiraike, O.; Hirata, T.; Hirota, Y.; et al. Accumulation of advanced glycation end products in follicles is associated with poor oocyte developmental competence. Mol. Hum. Reprod. 2019, 25, 684–694. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Ohta, K.; Mizuno, A.; Li, S.; Itoh, M.; Ueda, M.; Ohta, E.; Hida, Y.; Wang, M.X.; Furoi, M.; Tsuzuki, Y.; et al. Endoplasmic reticulum stress enhances gamma-secretase activity. Biochem. Biophys. Res. Commun. 2011, 416, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Y.; Fu, H.; Huang, H.; Lu, Q.; Qin, H.; Wu, Y.; Huang, H.; Mao, G.; Wei, Z.; et al. Hes1 Knockdown Exacerbates Ischemic Stroke Following tMCAO by Increasing ER Stress-Dependent Apoptosis via the PERK/eIF2alpha/ATF4/CHOP Signaling Pathway. Neurosci. Bull. 2020, 36, 134–142. [Google Scholar] [CrossRef]
- Zhao, L.; Cai, H.; Tang, Z.; Cui, Q.; Liu, Z.; Lu, S. Matrine suppresses advanced glycation end products-induced human coronary smooth muscle cells phenotype conversion by regulating endoplasmic reticulum stress-dependent Notch signaling. Eur. J. Pharmacol. 2020, 882, 173257. [Google Scholar] [CrossRef] [PubMed]
- Szabat, M.; Kalynyak, T.B.; Lim, G.E.; Chu, K.Y.; Yang, Y.H.; Asadi, A.; Gage, B.K.; Ao, Z.; Warnock, G.L.; Piret, J.M.; et al. Musashi expression in β-cells coordinates insulin expression, apoptosis and proliferation in response to endoplasmic reticulum stress in diabetes. Cell Death Dis. 2011, 2, e232. [Google Scholar] [CrossRef]
- Dadey, D.Y.; Kapoor, V.; Khudanyan, A.; Urano, F.; Kim, A.H.; Thotala, D.; Hallahan, D.E. The ATF6 pathway of the ER stress response contributes to enhanced viability in glioblastoma. Oncotarget 2016, 7, 2080–2092. [Google Scholar] [CrossRef]
- Lee, J.E.; Morrison, W.; Hollien, J. Hairy and enhancer of split 1 (HES1) protects cells from endoplasmic reticulum stress-induced apoptosis through repression of GADD34. J. Biol. Chem. 2018, 293, 5947–5955. [Google Scholar] [CrossRef] [PubMed]
- Jaud, M.; Philippe, C.; Van Den Berghe, L.; Ségura, C.; Mazzolini, L.; Pyronnet, S.; Laurell, H.; Touriol, C. The PERK Branch of the Unfolded Protein Response Promotes DLL4 Expression by Activating an Alternative Translation Mechanism. Cancers 2019, 11, 142. [Google Scholar] [CrossRef]
- Ge, D.; Jing, Q.; Zhao, W.; Yue, H.; Su, L.; Zhang, S.; Zhao, J. Finding ATF4/p75NTR/IL-8 signal pathway in endothelial-mesenchymal transition by safrole oxide. PLoS ONE 2014, 9, e99378. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Li, H.; Liu, J.; Xu, L.; Yang, L.; Sun, Z.; Zhou, B. Tunicamycin aggravates endoplasmic reticulum stress and airway inflammation via PERKATF4-CHOP signaling in a murine model of neutrophilic asthma. J. Asthma 2017, 54, 125–133. [Google Scholar] [CrossRef]
- Yu, J.; Zhu, C.; Wang, X.; Kim, K.J.; Bartolome, A.; Dongiovanni, P.; Yates, K.P.; Valenti, L.; Carrer, M.; Sadowski, T.; et al. Hepatocyte TLR4 triggers inter-hepatocyte Jagged1/Notch signaling to determine NASH-induced fibrosis. Sci. Transl. Med. 2021, 13, eabe1692. [Google Scholar] [CrossRef]
- Monteagudo, S.; Lories, R.J.A. Notch in the joint that exacerbates osteoarthritis. Nat. Rev. Rheumatol. 2018, 14, 563–564. [Google Scholar] [CrossRef]
- Zhou, B.; Lin, W.; Long, Y.; Yang, Y.; Zhang, H.; Wu, K.; Chu, Q. Notch signaling pathway: Architecture, disease, and therapeutics. Signal Transduct. Target. Ther. 2022, 7, 95. [Google Scholar] [CrossRef]
- Dumesic, D.A.; Oberfield, S.E.; Stener-Victorin, E.; Marshall, J.C.; Laven, J.S.; Legro, R.S. Scientific Statement on the Diagnostic Criteria, Epidemiology, Pathophysiology, and Molecular Genetics of Polycystic Ovary Syndrome. Endocr. Rev. 2015, 36, 487–525. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.; Liu, Z.; Ren, Y.A.; Wun, W.S.; Zhou, W.; Kenigsberg, S.; Librach, C.; Valdes, C.; Gibbons, W.; Richards, J. Enhanced Inflammatory Transcriptome in the Granulosa Cells of Women with Polycystic Ovarian Syndrome. J. Clin. Endocrinol. Metab. 2016, 101, 3459–3468. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Ma, Y.; Tong, X.; Yang, W.; Dai, Y.; Pan, Y.; Ren, P.; Liu, L.; Fan, H.Y.; Zhang, Y.; et al. Metformin inhibits testosterone-induced endoplasmic reticulum stress in ovarian granulosa cells via inactivation of p38 MAPK. Hum. Reprod. 2020, 35, 1145–1158. [Google Scholar] [CrossRef] [PubMed]
- Di, F.; Liu, J.; Li, S.; Yao, G.; Hong, Y.; Chen, Z.J.; Li, W.; Du, Y. ATF4 Contributes to Ovulation via Regulating COX2/PGE2 Expression: A Potential Role of ATF4 in PCOS. Front. Endocrinol. 2018, 9, 669. [Google Scholar] [CrossRef] [PubMed]
- Patil, K.; Shinde, G.; Hinduja, I.; Mukherjee, S. Compromised Cumulus-Oocyte Complex Matrix Organization and Expansion in Women with PCOS. Reprod. Sci. 2022, 29, 836–848. [Google Scholar] [CrossRef]
- Robker, R.L.; Hennebold, J.D.; Russell, D.L. Coordination of Ovulation and Oocyte Maturation: A Good Egg at the Right Time. Endocrinology 2018, 159, 3209–3218. [Google Scholar] [CrossRef] [PubMed]
- Teramoto, S.; Osada, H.; Shozu, M. Prematurely ruptured dominant follicles often retain competent oocytes in infertile women. Sci. Rep. 2019, 9, 15041. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (n = 12) | PCOS (n = 11) | p-Value | |
|---|---|---|---|
| Age (years) | 34.0 (26–40) | 31.0 (24–37) | 0.1236 |
| BMI (kg/m2) | 21.9 (19.7–25.4) | 21.8 (18.1–26.4) | 0.8916 |
| LH (mIU/mL) | 4.8 (2.1–5.6) | 7.9 (5.9–15.4) | 0.0002 |
| FSH (mIU/mL) | 7.2 (5.3–8.3) | 6.5 (4.9–8.4) | 0.1057 |
| LH/FSH | 0.64 (0.32–0.91) | 1.28 (0.87–2.23) | <0.0001 |
| AMH (ng/mL) | 2.56 (1.51–6.73) | 8.33 (5.44–24.63) | 0.0010 |
| Gene | Forward 5′–3′ | Reverse 5′–3′ |
|---|---|---|
| Notch2 | CAACCGCAATGGAGGCTATG | GCGAAGGCACAATCATCAATGTT |
| Hey2 | GCCCGCCCTTGTCAGTATC | CCAGGGTCGGTAAGGTTTATTG |
| Hes1 | CCCAACGCAGTGTCACCTTC | TACAAAGGCGGCAATCCAATATG |
| ATF4 | GGCTGGCTGTGGATGGGTTG | CTCCTGGACTAGGGGGGCAA |
| Areg | GTGGTGCTGTCGCTCTTGATA | CCCCAGAAAATGGTTCACGCT |
| Ereg | GTGATTCCATCATGTATCCCAGG | GCCATTCATGTCAGAGCTACACT |
| Tnfaip6 | TTTCTCTTGCTATGGGAAGACAC | GAGCTTGTATTTGCCAGACCG |
| Has2 | CTCTTTTGGACTGTATGGTGCC | AGGGTAGGTTAGCCTTTTCACA |
| COX2 | CTGGCGCTCAGCCATACAG | CGCACTTATACTGGTCAAATCCC |
| GAPDH | TGGACCTGACCTGCCGTCTA | CTGCTTCACCACCTTCTTGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koike, H.; Harada, M.; Kusamoto, A.; Kunitomi, C.; Xu, Z.; Tanaka, T.; Urata, Y.; Nose, E.; Takahashi, N.; Wada-Hiraike, O.; et al. Notch Signaling Induced by Endoplasmic Reticulum Stress Regulates Cumulus-Oocyte Complex Expansion in Polycystic Ovary Syndrome. Biomolecules 2022, 12, 1037. https://doi.org/10.3390/biom12081037
Koike H, Harada M, Kusamoto A, Kunitomi C, Xu Z, Tanaka T, Urata Y, Nose E, Takahashi N, Wada-Hiraike O, et al. Notch Signaling Induced by Endoplasmic Reticulum Stress Regulates Cumulus-Oocyte Complex Expansion in Polycystic Ovary Syndrome. Biomolecules. 2022; 12(8):1037. https://doi.org/10.3390/biom12081037
Chicago/Turabian StyleKoike, Hiroshi, Miyuki Harada, Akari Kusamoto, Chisato Kunitomi, Zixin Xu, Tsurugi Tanaka, Yoko Urata, Emi Nose, Nozomi Takahashi, Osamu Wada-Hiraike, and et al. 2022. "Notch Signaling Induced by Endoplasmic Reticulum Stress Regulates Cumulus-Oocyte Complex Expansion in Polycystic Ovary Syndrome" Biomolecules 12, no. 8: 1037. https://doi.org/10.3390/biom12081037
APA StyleKoike, H., Harada, M., Kusamoto, A., Kunitomi, C., Xu, Z., Tanaka, T., Urata, Y., Nose, E., Takahashi, N., Wada-Hiraike, O., Hirota, Y., Koga, K., & Osuga, Y. (2022). Notch Signaling Induced by Endoplasmic Reticulum Stress Regulates Cumulus-Oocyte Complex Expansion in Polycystic Ovary Syndrome. Biomolecules, 12(8), 1037. https://doi.org/10.3390/biom12081037