
Overcoming the Limitations of Stem Cell-Derived Beta Cells


Abstract
:1. Introduction
2. A Journey to Produce Functional Beta Cells
3. Addressing the Difficulties
3.1. Efficiency of Differentiation and Polyhormonal Cells
3.2. Functional Maturation
4. Further Steps
4.1. Organoids
4.2. Transplantation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jennings, R.E.; Berry, A.A.; Strutt, J.P.; Gerrard, D.T.; Hanley, N.A. Human pancreas development. Development 2015, 142, 3126–3137. [Google Scholar] [CrossRef] [Green Version]
- Andralojc, K.M.; Mercalli, A.; Nowak, K.W.; Albarello, L.; Calcagno, R.; Luzi, L.; Bonifacio, E.; Doglioni, C.; Piemonti, L. Ghrelin-producing epsilon cells in the developing and adult human pancreas. Diabetologia 2009, 52, 486–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Klughammer, J.; Farlik, M.; Penz, T.; Spittler, A.; Barbieux, C.; Berishvili, E.; Bock, C.; Kubicek, S. Single-cell transcriptomes reveal characteristic features of human pancreatic islet cell types. EMBO Rep. 2016, 17, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Segerstolpe, Å.; Palasantza, A.; Eliasson, P.; Andersson, E.M.; Andréasson, A.C.; Sun, X.; Picelli, S.; Sabirsh, A.; Clausen, M.; Bjursell, M.K.; et al. Single-cell transcriptome profiling of human pancreatic islets in health and type 2 diabetes. Cell Metab. 2016, 24, 593–607. [Google Scholar] [CrossRef] [Green Version]
- Herold, K.C.; Vignali, D.A.; Cooke, A.; Bluestone, J.A. Type 1 diabetes: Translating mechanistic observations into effective clinical outcomes. Nat. Rev. Immunol. 2013, 13, 243–256. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Ding, Y.; Tanaka, Y.; Zhang, W. Risk factors contributing to type 2 diabetes and recent advances in the treatment and prevention. Int. J. Med. Sci. 2014, 11, 1185–1200. [Google Scholar] [CrossRef] [Green Version]
- Heller, S.; White, D.; Lee, E.; Lawton, J.; Pollard, D.; Waugh, N.; Amiel, S.; Barnard, K.; Beckwith, A.; Brennan, A.; et al. A cluster randomised trial, cost-effectiveness analysis and psychosocial evaluation of insulin pump therapy compared with multiple injections during flexible intensive insulin therapy for type 1 diabetes: The REPOSE Trial. Health Technol. Assess. 2017, 21, 1–278. [Google Scholar] [CrossRef]
- Dicembrini, I.; Cosentino, C.; Monami, M.; Mannucci, E.; Pala, L. Effects of real-time continuous glucose monitoring in type 1 diabetes: A meta-analysis of randomized controlled trials. Acta Diabetol. 2021, 58, 401–410. [Google Scholar] [CrossRef]
- Weisman, A.; Bai, J.W.; Cardinez, M.; Kramer, C.K.; Perkins, B.A. Effect of artificial pancreas systems on glycaemic control in patients with type 1 diabetes: A systematic review and meta-analysis of outpatient randomised controlled trials. Lancet Diabetes Endocrinol. 2017, 5, 501–512. [Google Scholar] [CrossRef]
- Marks, B.E.; Williams, K.M.; Sherwood, J.S.; Putman, M.S. Practical aspects of diabetes technology use: Continuous glucose monitors, insulin pumps, and automated insulin delivery systems. J. Clin. Transl. Endocrinol. 2021, 27, 100282. [Google Scholar] [CrossRef]
- Shapiro, A.M.; Lakey, J.R.; Ryan, E.A.; Korbutt, G.S.; Toth, E.; Warnock, G.L.; Kneteman, N.M.; Rajotte, R.V. Islet transplantation in seven patients with type 1 diabetes mellitus using a glucocorticoid-free immunosuppressive regimen. N. Engl. J. Med. 2000, 343, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Markmann, J.F.; Rickels, M.R.; Eggerman, T.L.; Bridges, N.D.; Lafontant, D.E.; Qidwai, J.; Foster, E.; Clarke, W.R.; Kamoun, M.; Alejandro, R.; et al. Phase 3 trial of human islet-after-kidney transplantation in type 1 diabetes. Am. J. Transplant. Off. J. Am. Soc. Transplant. Am. Soc. Transpl. Surg. 2021, 21, 1477–1492. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J.; D’Alessio, D.A.; Fradkin, J.; Kernan, W.N.; Mathieu, C.; Mingrone, G.; Rossing, P.; Tsapas, A.; Wexler, D.J.; Buse, J.B. Management of hyperglycaemia in type 2 diabetes, A consensus report by the American Diabetes Association (ADA) and the European Association for the Study of Diabetes (EASD). Diabetologia 2018, 61, 2461–2498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidu, S.; Gillies, C.; Farooqi, A.; Trivedi, H.; Than, T.; Brady, E.; Davies, M.J.; Khunti, K. A cost comparison of an enhanced primary care diabetes service and standard care. Prim. Care Diabetes 2021, 15, 601–606. [Google Scholar] [CrossRef]
- Zhang, P.; Atkinson, K.M.; Bray, G.A.; Chen, H.; Clark, J.M.; Coday, M.; Dutton, G.R.; Egan, C.; Espeland, M.A.; Evans, M.; et al. Look AHEAD Research Group Within-trial cost-effectiveness of a structured lifestyle intervention in adults with overweight/obesity and type 2 diabetes: Results from the action for health in diabetes (Look AHEAD) study. Diabetes Care 2021, 44, 67–74. [Google Scholar] [CrossRef]
- D’Amour, K.A.; Agulnick, A.D.; Eliazer, S.; Kelly, O.G.; Kroon, E.; Baetge, E.E. Efficient differentiation of human embryonic stem cells to definitive endoderm. Nat. Biotechnol. 2005, 23, 1534–1541. [Google Scholar] [CrossRef]
- Galivo, F.; Benedetti, E.; Wang, Y.; Pelz, C.; Schug, J.; Kaestner, K.H.; Grompe, M. Reprogramming human gallbladder cells into insulin-producing β-like cells. PLoS ONE 2017, 12, e0181812. [Google Scholar] [CrossRef] [Green Version]
- Sapir, T.; Shternhall, K.; Meivar-Levy, I.; Blumenfeld, T.; Cohen, H.; Skutelsky, E.; Eventov-Friedman, S.; Barshack, I.; Goldberg, I.; Pri-Chen, S.; et al. Cell-replacement therapy for diabetes: Generating functional insulin-producing tissue from adult human liver cells. Proc. Natl. Acad. Sci. USA 2005, 102, 7964–7969. [Google Scholar] [CrossRef] [Green Version]
- Berneman-Zeitouni, D.; Molakandov, K.; Elgart, M.; Mor, E.; Fornoni, A.; Domínguez, M.R.; Kerr-Conte, J.; Ott, M.; Meivar-Levy, I.; Ferber, S. The temporal and hierarchical control of transcription factors-induced liver to pancreas transdifferentiation. PLoS ONE 2014, 9, e87812. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.N.; Yi, H.J.; Seo, E.H.; Oh, J.; Lee, S.; Ferber, S.; Okano, T.; Shim, I.K.; Kim, S.C. Improvement of the therapeutic capacity of insulin-producing cells trans-differentiated from human liver cells using engineered cell sheet. Stem Cell Res. Ther. 2021, 12, 3. [Google Scholar] [CrossRef]
- Lemper, M.; Leuckx, G.; Heremans, Y.; German, M.S.; Heimberg, H.; Bouwens, L.; Baeyens, L. Reprogramming of human pancreatic exocrine cells to β-like cells. Cell Death Differ. 2015, 22, 1117–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, M.J.; Muir, K.R.; Docherty, H.M.; McGowan, N.W.; Forbes, S.; Heremans, Y.; Heimberg, H.; Casey, J.; Docherty, K. Generation of functional beta-like cells from human exocrine pancreas. PLoS ONE 2016, 11, e0156204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bramswig, N.C.; Everett, L.J.; Schug, J.; Dorrell, C.; Liu, C.; Luo, Y.; Streeter, P.R.; Naji, A.; Grompe, M.; Kaestner, K.H. Epigenomic plasticity enables human pancreatic α to β cell reprogramming. J. Clin. Investig. 2013, 123, 1275–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Liu, Q.; Zhou, Z.; Ikeda, Y. PDX1, Neurogenin-3, and MAFA: Critical transcription regulators for beta cell development and regeneration. Stem Cell Res. Ther. 2017, 8, 240. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Wang, H.; Wang, T.; Wei, C.; Jiang, H.; Jiang, S.; Yang, J.; Shao, J.; Ma, L. Pax4 synergistically acts with Pdx1, Ngn3 and MafA to induce HuMSCs to differentiate into functional pancreatic β-cells. Exp. Ther. Med. 2019, 18, 2592–2598. [Google Scholar] [CrossRef] [Green Version]
- Swisa, A.; Avrahami, D.; Eden, N.; Zhang, J.; Feleke, E.; Dahan, T.; Cohen-Tayar, Y.; Stolovich-Rain, M.; Kaestner, K.H.; Glaser, B.; et al. PAX6 maintains β cell identity by repressing genes of alternative islet cell types. J. Clin. Investig. 2017, 127, 230–243. [Google Scholar] [CrossRef]
- Murry, C.E.; Keller, G. Differentiation of embryonic stem cells to clinically relevant populations: Lessons from embryonic development. Cell 2008, 132, 661–680. [Google Scholar] [CrossRef] [Green Version]
- Mfopou, J.K.; Chen, B.; Mateizel, I.; Sermon, K.; Bouwens, L. Noggin, retinoids, and fibroblast growth factor regulate hepatic or pancreatic fate of human embryonic stem cells. Gastroenterology 2010, 138, 2233–2245. [Google Scholar] [CrossRef]
- Chen, S.; Borowiak, M.; Fox, J.L.; Maehr, R.; Osafune, K.; Davidow, L.; Lam, K.; Peng, L.F.; Schreiber, S.L.; Rubin, L.L.; et al. A small molecule that directs differentiation of human ESCs into the pancreatic lineage. Nat. Chem. Biol. 2009, 5, 258–265. [Google Scholar] [CrossRef]
- Nostro, M.C.; Sarangi, F.; Ogawa, S.; Holtzinger, A.; Corneo, B.; Li, X.; Micallef, S.J.; Park, I.H.; Basford, C.; Wheeler, M.B.; et al. Stage-specific signaling through TGFβ family members and WNT regulates patterning and pancreatic specification of human pluripotent stem cells. Development 2011, 138, 861–871. [Google Scholar] [CrossRef] [Green Version]
- Shim, J.H.; Kim, J.; Han, J.; An, S.Y.; Jang, Y.J.; Son, J.; Woo, D.H.; Kim, S.K.; Kim, J.H. Pancreatic islet-like three-dimensional aggregates derived from human embryonic stem cells ameliorate hyperglycemia in streptozotocin-induced diabetic mice. Cell Transplant. 2015, 24, 2155–2168. [Google Scholar] [CrossRef] [PubMed]
- Rezania, A.; Bruin, J.E.; Arora, P.; Rubin, A.; Batushansky, I.; Asadi, A.; O’Dwyer, S.; Quiskamp, N.; Mojibian, M.; Albrecht, T.; et al. Reversal of diabetes with insulin-producing cells derived in vitro from human pluripotent stem cells. Nat. Biotechnol. 2014, 32, 1121–1133. [Google Scholar] [CrossRef] [PubMed]
- Pagliuca, F.W.; Millman, J.R.; Gürtler, M.; Segel, M.; Van Dervort, A.; Ryu, J.H.; Peterson, Q.P.; Greiner, D.; Melton, D.A. Generation of functional human pancreatic β cells in vitro. Cell 2014, 159, 428–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russ, H.A.; Parent, A.V.; Ringler, J.J.; Hennings, T.G.; Nair, G.G.; Shveygert, M.; Guo, T.; Puri, S.; Haataja, L.; Cirulli, V.; et al. Controlled induction of human pancreatic progenitors produces functional beta-like cells in vitro. EMBO J. 2015, 34, 1759–1772. [Google Scholar] [CrossRef] [Green Version]
- Nair, G.G.; Liu, J.S.; Russ, H.A.; Tran, S.; Saxton, M.S.; Chen, R.; Juang, C.; Li, M.L.; Nguyen, V.Q.; Giacometti, S.; et al. Recapitulating endocrine cell clustering in culture promotes maturation of human stem-cell-derived β cells. Nat. Cell Biol. 2019, 21, 263–274. [Google Scholar] [CrossRef]
- Liu, H.; Li, R.; Liao, H.K.; Min, Z.; Wang, C.; Yu, Y.; Shi, L.; Dan, J.; Hayek, A.; Martinez Martinez, L.; et al. Chemical combinations potentiate human pluripotent stem cell-derived 3D pancreatic progenitor clusters toward functional β cells. Nat. Commun. 2021, 12, 3330. [Google Scholar] [CrossRef]
- Wang, D.; Wang, J.; Bai, L.; Pan, H.; Feng, H.; Clevers, H.; Zeng, Y.A. Long-term expansion of pancreatic islet organoids from resident Procr+ progenitors. Cell 2020, 180, 1198–1211. [Google Scholar] [CrossRef]
- Gock, H.; Lee, K.F.; Murray-Segal, L.; Mysore, T.B.; d’Apice, A.J.; Salvaris, E.J.; Cowan, P.J. Human endothelial protein C receptor overexpression protects intraportal islet grafts in mice. Transplant. Proc. 2016, 48, 2200–2207. [Google Scholar] [CrossRef]
- Kopp, J.L.; Dubois, C.L.; Schaffer, A.E.; Hao, E.; Shih, H.P.; Seymour, P.A.; Ma, J.; Sander, M. Sox9+ ductal cells are multipotent progenitors throughout development but do not produce new endocrine cells in the normal or injured adult pancreas. Development 2011, 138, 653–665. [Google Scholar] [CrossRef] [Green Version]
- Millman, J.R.; Xie, C.; Van Dervort, A.; Gürtler, M.; Pagliuca, F.W.; Melton, D.A. Generation of stem cell-derived β-cells from patients with type 1 diabetes. Nat. Commun. 2016, 7, 11463. [Google Scholar] [CrossRef] [Green Version]
- Maahs, D.M.; West, N.A.; Lawrence, J.M.; Mayer-Davis, E.J. Epidemiology of type 1 diabetes. Endocrinol. Metab. Clin. N. Am. 2010, 39, 481–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, O.G.; Chan, M.Y.; Martinson, L.A.; Kadoya, K.; Ostertag, T.M.; Ross, K.G.; Richardson, M.; Carpenter, M.K.; D’Amour, K.A.; Kroon, E.; et al. Cell-surface markers for the isolation of pancreatic cell types derived from human embryonic stem cells. Nat. Biotechnol. 2011, 29, 750–756. [Google Scholar] [CrossRef] [PubMed]
- Bocian-Sobkowska, J.; Zabel, M.; Wozniak, W.; Surdyk-Zasada, J. Polyhormonal aspect of the endocrine cells of the human fetal pancreas. Histochem. Cell Biol. 1999, 112, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Veres, A.; Faust, A.L.; Bushnell, H.L.; Engquist, E.N.; Kenty, J.H.; Harb, G.; Poh, Y.C.; Sintov, E.; Gürtler, M.; Pagliuca, F.W.; et al. Charting cellular identity during human in vitro β-cell differentiation. Nature 2019, 569, 368–373. [Google Scholar] [CrossRef]
- Alvarez-Dominguez, J.R.; Melton, D.A. Cell maturation: Hallmarks, triggers, and manipulation. Cell 2022, 185, 235–249. [Google Scholar] [CrossRef]
- Yoshihara, E. Adapting physiology in functional human islet organogenesis. Front. Cell Dev. Biol. 2022, 10, 854604. [Google Scholar] [CrossRef]
- Barsby, T.; Otonkoski, T. Maturation of beta cells: Lessons from in vivo and in vitro models. Diabetologia 2022, 65, 917–930. [Google Scholar] [CrossRef]
- Henquin, J.C. Regulation of insulin secretion: A matter of phase control and amplitude modulation. Diabetologia 2009, 52, 739–751. [Google Scholar] [CrossRef] [Green Version]
- Campbell, J.E.; Newgard, C.B. Mechanisms controlling pancreatic islet cell function in insulin secretion. Nat. Rev. Mol. Cell Biol. 2021, 22, 142–158. [Google Scholar] [CrossRef]
- Gaisano, H.Y. Recent new insights into the role of SNARE and associated proteins in insulin granule exocytosis. Diabetes Obes. Metab. 2017, 19 (Suppl. 1), 115–123. [Google Scholar] [CrossRef] [Green Version]
- Kalwat, M.A.; Cobb, M.H. Mechanisms of the amplifying pathway of insulin secretion in the β cell. Pharmacol. Ther. 2017, 179, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Augsornworawat, P.; Maxwell, K.G.; Velazco-Cruz, L.; Millman, J.R. Single-cell transcriptome profiling reveals β cell maturation in stem cell-derived islets after transplantation. Cell Rep. 2020, 32, 108067. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.L.; Lee, S.; Hoek, P.; van der Meulen, T.; Van, R.; Huising, M.O. Genetic deletion of Urocortin 3 does not prevent functional maturation of beta cells. J. Endocrinol. 2020, 246, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Helman, A.; Cangelosi, A.L.; Davis, J.C.; Pham, Q.; Rothman, A.; Faust, A.L.; Straubhaar, J.R.; Sabatini, D.M.; Melton, D.A. A nutrient-sensing transition at birth triggers glucose-responsive insulin secretion. Cell Metab. 2020, 31, 1004–1016. [Google Scholar] [CrossRef]
- Yoshihara, E.; Wei, Z.; Lin, C.S.; Fang, S.; Ahmadian, M.; Kida, Y.; Tseng, T.; Dai, Y.; Yu, R.T.; Liddle, C.; et al. ERRγ is required for the metabolic maturation of therapeutically functional glucose-responsive β cells. Cell Metab. 2016, 23, 622–634. [Google Scholar] [CrossRef] [Green Version]
- Velazco-Cruz, L.; Song, J.; Maxwell, K.G.; Goedegebuure, M.M.; Augsornworawat, P.; Hogrebe, N.J.; Millman, J.R. Acquisition of dynamic function in human stem cell-derived β cells. Stem Cell Rep. 2019, 12, 351–365. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Liu, J.; Na, L.; Chen, L. Roles of epigenetic modifications in the differentiation and function of pancreatic β-cells. Front. Cell Dev. Biol. 2020, 8, 748. [Google Scholar] [CrossRef]
- Ackermann, A.M.; Wang, Z.; Schug, J.; Naji, A.; Kaestner, K.H. Integration of ATAC-seq and RNA-seq identifies human alpha cell and beta cell signature genes. Mol. Metab. 2016, 5, 233–244. [Google Scholar] [CrossRef]
- Wang, A.; Yue, F.; Li, Y.; Xie, R.; Harper, T.; Patel, N.A.; Muth, K.; Palmer, J.; Qiu, Y.; Wang, J.; et al. Epigenetic priming of enhancers predicts developmental competence of hESC-derived endodermal lineage intermediates. Cell Stem Cell 2015, 16, 386–399. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Li, X. Metabolic and epigenetic regulation of endoderm differentiation. Trends Cell Biol. 2022, 32, 151–164. [Google Scholar] [CrossRef]
- Sabari, B.R.; Tang, Z.; Huang, H.; Yong-Gonzalez, V.; Molina, H.; Kong, H.E.; Dai, L.; Shimada, M.; Cross, J.R.; Zhao, Y.; et al. Intracellular crotonyl-CoA stimulates transcription through p300-catalyzed histone crotonylation. Mol. Cell 2015, 58, 203–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogrebe, N.J.; Augsornworawat, P.; Maxwell, K.G.; Velazco-Cruz, L.; Millman, J.R. Targeting the cytoskeleton to direct pancreatic differentiation of human pluripotent stem cells. Nat. Biotechnol. 2020, 38, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Trogden, K.P.; Lee, J.; Bracey, K.M.; Ho, K.H.; McKinney, H.; Zhu, X.; Arpag, G.; Folland, T.G.; Osipovich, A.B.; Magnuson, M.A.; et al. Microtubules regulate pancreatic β-cell heterogeneity via spatiotemporal control of insulin secretion hot spots. eLife 2021, 10, e59912. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Zhu, X.; Yuan, M.; Ho, K.H.; Kaverina, I.; Gu, G. Microtubules and Gαo-signaling modulate the preferential secretion of young insulin secretory granules in islet β cells via independent pathways. PLoS ONE 2021, 16, e0241939. [Google Scholar] [CrossRef] [PubMed]
- Moede, T.; Leibiger, I.B.; Berggren, P.O. Alpha cell regulation of beta cell function. Diabetologia 2020, 63, 2064–2075. [Google Scholar] [CrossRef]
- Peterson, Q.P.; Veres, A.; Chen, L.; Slama, M.Q.; Kenty, J.; Hassoun, S.; Brown, M.R.; Dou, H.; Duffy, C.D.; Zhou, Q.; et al. A method for the generation of human stem cell-derived alpha cells. Nat. Commun. 2020, 11, 2241. [Google Scholar] [CrossRef]
- Gu, W.; Anker, C.; Christiansen, C.B.; Moede, T.; Berggren, P.O.; Hermansen, K.; Gregersen, S.; Jeppesen, P.B. Pancreatic β cells inhibit glucagon secretion from α cells: An in vitro demonstration of α-β cell interaction. Nutrients 2021, 13, 2281. [Google Scholar] [CrossRef]
- Zhang, Y.; Han, C.; Zhu, W.; Yang, G.; Peng, X.; Mehta, S.; Zhang, J.; Chen, L.; Liu, Y. Glucagon potentiates insulin secretion via β-cell GCGR at physiological concentrations of glucose. Cells 2021, 10, 2495. [Google Scholar] [CrossRef]
- Vivot, K.; Moullé, V.S.; Zarrouki, B.; Tremblay, C.; Mancini, A.D.; Maachi, H.; Ghislain, J.; Poitout, V. The regulator of G-protein signaling RGS16 promotes insulin secretion and β-cell proliferation in rodent and human islets. Mol. Metab. 2016, 5, 988–996. [Google Scholar] [CrossRef]
- Rorsman, P.; Huising, M.O. The somatostatin-secreting pancreatic δ-cell in health and disease. Nat. Rev. Endocrinol. 2018, 14, 404–414. [Google Scholar] [CrossRef]
- Kim, W.; Fiori, J.L.; Shin, Y.K.; Okun, E.; Kim, J.S.; Rapp, P.R.; Egan, J.M. Pancreatic polypeptide inhibits somatostatin secretion. FEBS Lett. 2014, 588, 3233–3239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aragón, F.; Karaca, M.; Novials, A.; Maldonado, R.; Maechler, P.; Rubí, B. Pancreatic polypeptide regulates glucagon release through PPYR1 receptors expressed in mouse and human alpha-cells. Biochim. Et Biophys. Acta 2015, 1850, 343–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dezaki, K.; Kakei, M.; Yada, T. Ghrelin uses Galphai2 and activates voltage-dependent K+ channels to attenuate glucose-induced Ca2+ signaling and insulin release in islet beta-cells: Novel signal transduction of ghrelin. Diabetes 2007, 56, 2319–2327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, J.; Prigeon, R.L.; Davis, H.W.; Bidlingmaier, M.; Kahn, S.E.; Cummings, D.E.; Tschöp, M.H.; D’Alessio, D. Ghrelin suppresses glucose-stimulated insulin secretion and deteriorates glucose tolerance in healthy humans. Diabetes 2010, 59, 2145–2151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorrell, C.; Schug, J.; Canaday, P.S.; Russ, H.A.; Tarlow, B.D.; Grompe, M.T.; Horton, T.; Hebrok, M.; Streeter, P.R.; Kaestner, K.H.; et al. Human islets contain four distinct subtypes of β cells. Nat. Commun. 2016, 7, 11756. [Google Scholar] [CrossRef] [Green Version]
- Nasteska, D.; Hodson, D.J. The role of beta cell heterogeneity in islet function and insulin release. J. Mol. Endocrinol. 2018, 61, R43–R60. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.C.; Jessup, C.F.; Campbell-Thompson, M. The role of accessory cells in islet homeostasis. Curr. Diabetes Rep. 2018, 18, 117. [Google Scholar] [CrossRef]
- Nikolova, G.; Jabs, N.; Konstantinova, I.; Domogatskaya, A.; Tryggvason, K.; Sorokin, L.; Fässler, R.; Gu, G.; Gerber, H.P.; Ferrara, N.; et al. The vascular basement membrane: A niche for insulin gene expression and Beta cell proliferation. Dev. Cell 2006, 10, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Gan, W.J.; Do, O.H.; Cottle, L.; Ma, W.; Kosobrodova, E.; Cooper-White, J.; Bilek, M.; Thorn, P. Local integrin activation in pancreatic β cells targets insulin secretion to the vasculature. Cell Rep. 2018, 24, 2819–2826. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Cottle, L.; Loudovaris, T.; Xiao, D.; Yang, P.; Thomas, H.E.; Kebede, M.A.; Thorn, P. Enhanced structure and function of human pluripotent stem cell-derived beta-cells cultured on extracellular matrix. Stem Cells Transl. Med. 2021, 10, 492–505. [Google Scholar] [CrossRef]
- Almaça, J.; Weitz, J.; Rodriguez-Diaz, R.; Pereira, E.; Caicedo, A. The pericyte of the pancreatic islet regulates capillary diameter and local blood flow. Cell Metab. 2018, 27, 630–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Fu, W. Immune regulation of islet homeostasis and adaptation. J. Mol. Cell Biol. 2020, 12, 764–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brissova, M.; Aamodt, K.; Brahmachary, P.; Prasad, N.; Hong, J.Y.; Dai, C.; Mellati, M.; Shostak, A.; Poffenberger, G.; Aramandla, R.; et al. Islet microenvironment, modulated by vascular endothelial growth factor-A signaling, promotes β cell regeneration. Cell Metab. 2014, 19, 498–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, M.; de Winther, M.; Van den Bossche, J. Epigenetic mechanisms of macrophage activation in type 2 diabetes. Immunobiology 2017, 222, 937–943. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.C.; Baeyens, L.; Shen, C.N.; Peng, S.J.; Chien, H.J.; Scheel, D.W.; Chamberlain, C.E.; German, M.S. Human pancreatic neuro-insular network in health and fatty infiltration. Diabetologia 2018, 61, 168–181. [Google Scholar] [CrossRef] [Green Version]
- Ahrén, B. Autonomic regulation of islet hormone secretion--implications for health and disease. Diabetologia 2000, 43, 393–410. [Google Scholar] [CrossRef]
- Borden, P.; Houtz, J.; Leach, S.D.; Kuruvilla, R. Sympathetic innervation during development is necessary for pancreatic islet architecture and functional maturation. Cell Rep. 2013, 4, 287–301. [Google Scholar] [CrossRef]
- Kato, I.; Suzuki, Y.; Akabane, A.; Yonekura, H.; Tanaka, O.; Kondo, H.; Takasawa, S.; Yoshimoto, T.; Okamoto, H. Transgenic mice overexpressing human vasoactive intestinal peptide (VIP) gene in pancreatic beta cells. Evidence for improved glucose tolerance and enhanced insulin secretion by VIP and PHM-27 in vivo. J. Biol. Chem. 1994, 269, 21223–21228. [Google Scholar] [CrossRef]
- Filipsson, K.; Tornøe, K.; Holst, J.; Ahrén, B. Pituitary adenylate cyclase-activating polypeptide stimulates insulin and glucagon secretion in humans. J. Clin. Endocrinol. Metab. 1997, 82, 3093–3098. [Google Scholar] [CrossRef] [Green Version]
- Sakuma, Y.; Ricordi, C.; Miki, A.; Yamamoto, T.; Mita, A.; Barker, S.; Damaris, R.M.; Pileggi, A.; Yasuda, Y.; Yada, T.; et al. Effect of pituitary adenylate cyclase-activating polypeptide in islet transplantation. Transplant. Proc. 2009, 41, 343–345. [Google Scholar] [CrossRef] [Green Version]
- Bowers, D.T.; Song, W.; Wang, L.H.; Ma, M. Engineering the vasculature for islet transplantation. Acta Biomater. 2019, 95, 131–151. [Google Scholar] [CrossRef] [PubMed]
- Barshes, N.R.; Wyllie, S.; Goss, J.A. Inflammation-mediated dysfunction and apoptosis in pancreatic islet transplantation: Implications for intrahepatic grafts. J. Leukoc. Biol. 2005, 77, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Lebreton, F.; Lavallard, V.; Bellofatto, K.; Bonnet, R.; Wassmer, C.H.; Perez, L.; Kalandadze, V.; Follenzi, A.; Boulvain, M.; Kerr-Conte, J.; et al. Insulin-producing organoids engineered from islet and amniotic epithelial cells to treat diabetes. Nat. Commun. 2019, 10, 4491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, K.S.; Kim, Y.S.; Kim, J.H.; Choi, B.K.; Kim, S.H.; Oh, S.H.; Ahn, Y.R.; Lee, M.S.; Lee, M.K.; Park, J.B.; et al. Influence of human allogenic bone marrow and cord blood-derived mesenchymal stem cell secreting trophic factors on ATP (adenosine-5′-triphosphate)/ADP (adenosine-5′-diphosphate) ratio and insulin secretory function of isolated human islets from cadaveric donor. Transplant. Proc. 2009, 41, 3813–3818. [Google Scholar] [CrossRef]
- Rawal, S.; Williams, S.J.; Ramachandran, K.; Stehno-Bittel, L. Integration of mesenchymal stem cells into islet cell spheroids improves long-term viability, but not islet function. Islets 2017, 9, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Scuteri, A.; Donzelli, E.; Rodriguez-Menendez, V.; Ravasi, M.; Monfrini, M.; Bonandrini, B.; Figliuzzi, M.; Remuzzi, A.; Tredici, G. A double mechanism for the mesenchymal stem cells’ positive effect on pancreatic islets. PLoS ONE 2014, 9, e84309. [Google Scholar] [CrossRef] [Green Version]
- Carlsson, P.O.; Schwarcz, E.; Korsgren, O.; Le Blanc, K. Preserved β-cell function in type 1 diabetes by mesenchymal stromal cells. Diabetes 2015, 64, 587–592. [Google Scholar] [CrossRef] [Green Version]
- Yoshihara, E.; O’Connor, C.; Gasser, E.; Wei, Z.; Oh, T.G.; Tseng, T.W.; Wang, D.; Cayabyab, F.; Dai, Y.; Yu, R.T.; et al. Immune-evasive human islet-like organoids ameliorate diabetes. Nature 2020, 586, 606–611. [Google Scholar] [CrossRef]
- Zhang, Q.; Pan, Y.; Ji, J.; Xu, Y.; Zhang, Q.; Qin, L. Roles and action mechanisms of WNT4 in cell differentiation and human diseases: A review. Cell Death Discov. 2021, 7, 287. [Google Scholar] [CrossRef]
- Zhang, L.; Yu, D. Exosomes in cancer development, metastasis, and immunity. Biochim. Et Biophys. Acta Rev. Cancer 2019, 1871, 455–468. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Mao, Q.; Shen, C.; Wang, C.; Jia, W. Exosomes from β-cells alleviated hyperglycemia and enhanced angiogenesis in islets of streptozotocin-induced diabetic mice. Diabetes Metab. Syndr. Obes. Targets Ther. 2019, 12, 2053–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, Y.; Lin, N.; Ma, Q.; Chen, R.; Zhang, Z.; Wen, W.; Chen, H.; Sun, J. Circulating LncRNAs analysis in patients with type 2 diabetes reveals novel genes influencing glucose metabolism and islet β-cell function. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 46, 335–350. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Du, X.; Xu, J.; Zhang, Y.; Tian, Y.; Liu, G.; Wang, X.; Ma, M.; Du, W.; Liu, Y.; et al. Pancreatic β cell microRNA-26a alleviates type 2 diabetes by improving peripheral insulin sensitivity and preserving β cell function. PLoS Biol. 2020, 18, e3000603. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhou, Y.; Shi, Y.; Zhang, Y.; Liu, K.; Liang, R.; Sun, P.; Chang, X.; Tang, W.; Zhang, Y.; et al. Expression of miRNA-29 in pancreatic β cells promotes inflammation and diabetes via TRAF3. Cell Rep. 2021, 34, 108576. [Google Scholar] [CrossRef]
- Hirshberg, B. Lessons learned from the international trial of the edmonton protocol for islet transplantation. Curr. Diabetes Rep. 2007, 7, 301–303. [Google Scholar] [CrossRef]
- Vegas, A.J.; Veiseh, O.; Gürtler, M.; Millman, J.R.; Pagliuca, F.W.; Bader, A.R.; Doloff, J.C.; Li, J.; Chen, M.; Olejnik, K.; et al. Long-term glycemic control using polymer-encapsulated human stem cell-derived beta cells in immune-competent mice. Nat. Med. 2016, 22, 306–311. [Google Scholar] [CrossRef]
- Ahmad Raus, R.; Wan Nawawi, W.; Nasaruddin, R.R. Alginate and alginate composites for biomedical applications. Asian J. Pharm. Sci. 2021, 16, 280–306. [Google Scholar] [CrossRef]
- Bai, X.; Pei, Q.; Pu, C.; Chen, Y.; He, S.; Wang, B. Multifunctional islet transplantation hydrogel encapsulating A20 high-expressing islets. Drug Des. Dev. Ther. 2020, 14, 4021–4027. [Google Scholar] [CrossRef]
- Su, J.; Hu, B.H.; Lowe, W.L., Jr.; Kaufman, D.B.; Messersmith, P.B. Anti-inflammatory peptide-functionalized hydrogels for insulin-secreting cell encapsulation. Biomaterials 2010, 31, 308–314. [Google Scholar] [CrossRef] [Green Version]
- Osum, K.C.; Burrack, A.L.; Martinov, T.; Sahli, N.L.; Mitchell, J.S.; Tucker, C.G.; Pauken, K.E.; Papas, K.; Appakalai, B.; Spanier, J.A.; et al. Interferon-gamma drives programmed death-ligand 1 expression on islet β cells to limit T cell function during autoimmune diabetes. Sci. Rep. 2018, 8, 8295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-Gutierrez, R.; Alkanani, A.; Mathews, C.E.; Michels, A.; Russ, H.A. Protecting stem cell derived pancreatic beta-like cells from diabetogenic T cell recognition. Front. Endocrinol. 2021, 12, 707881. [Google Scholar] [CrossRef] [PubMed]
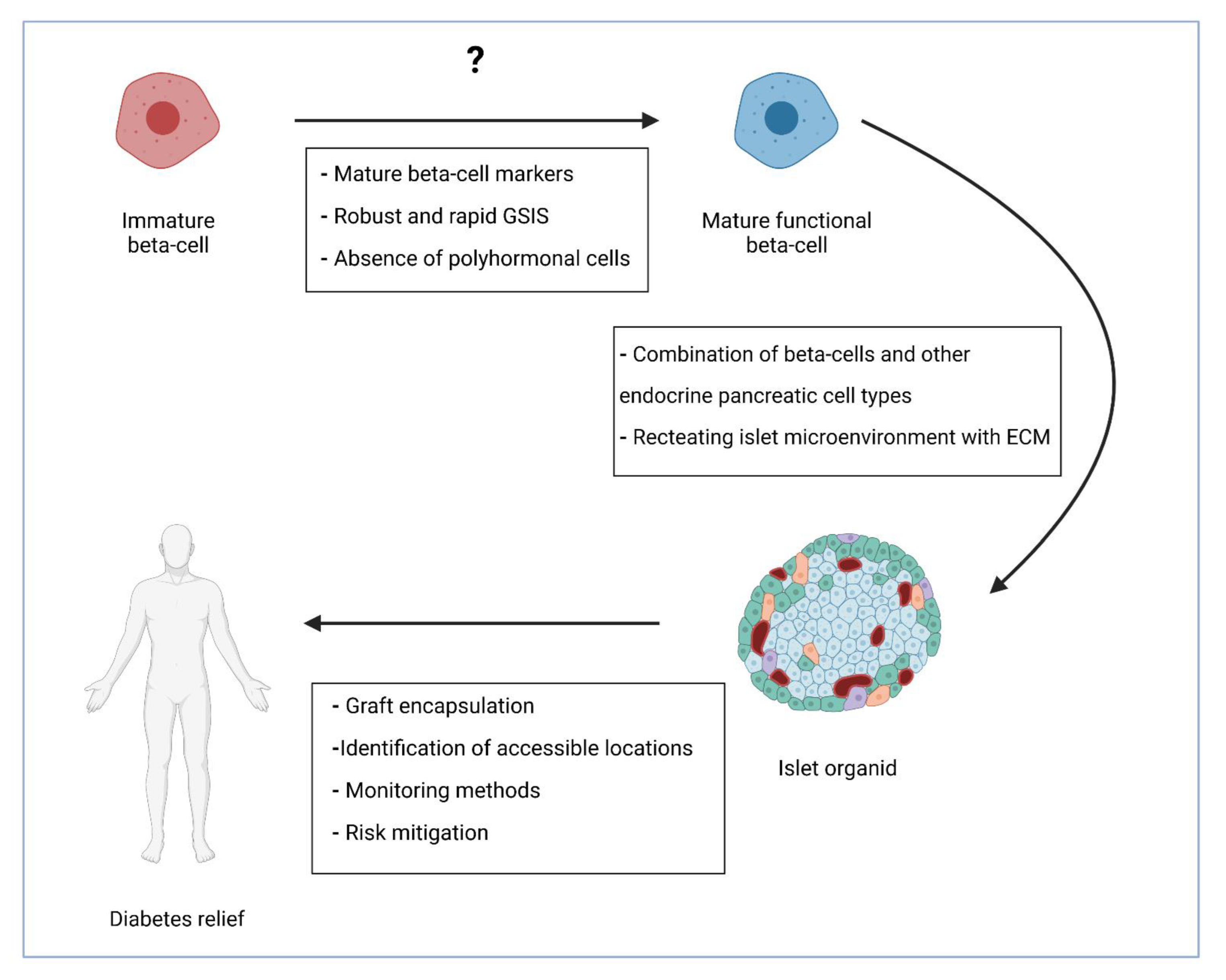

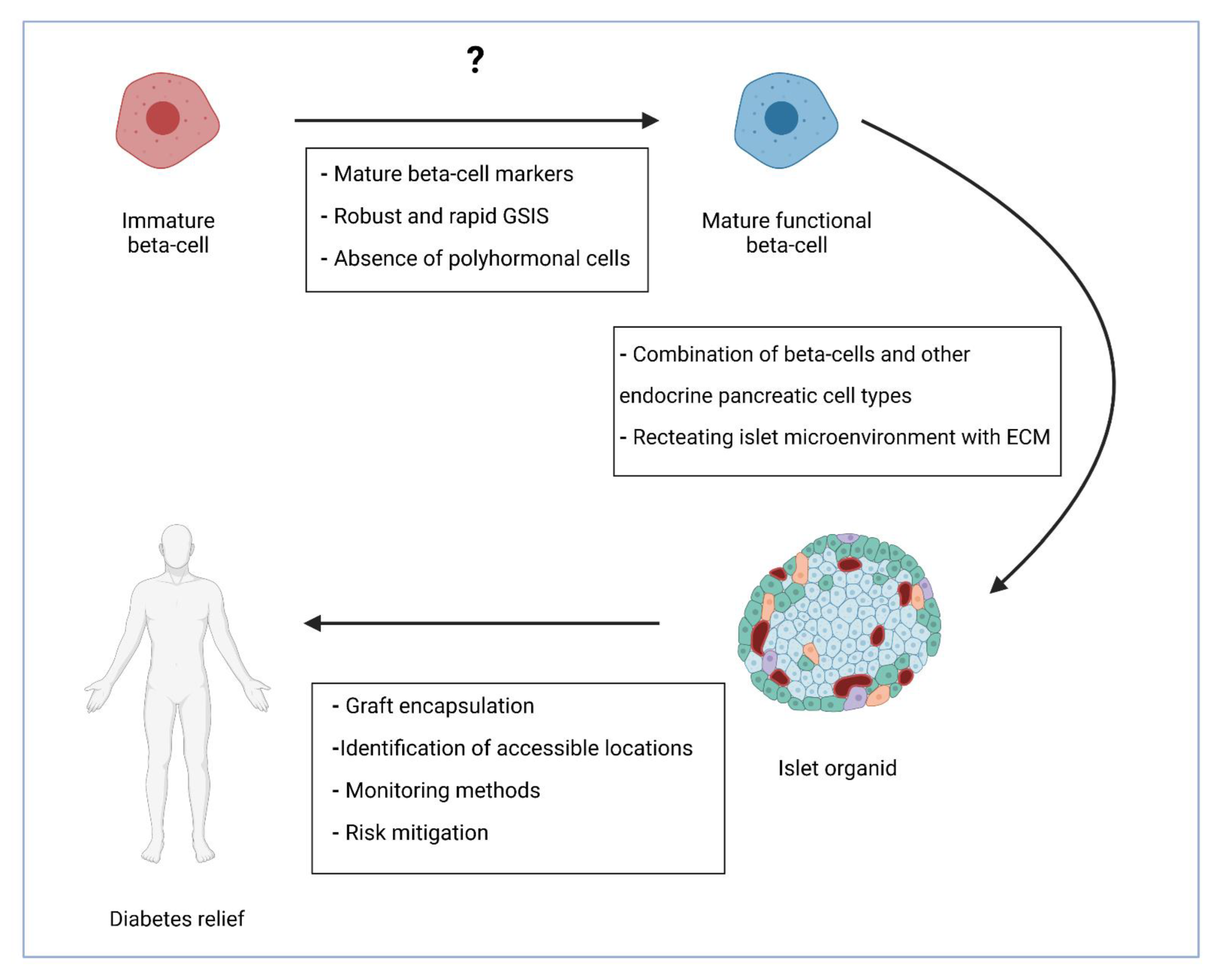{kind=link}
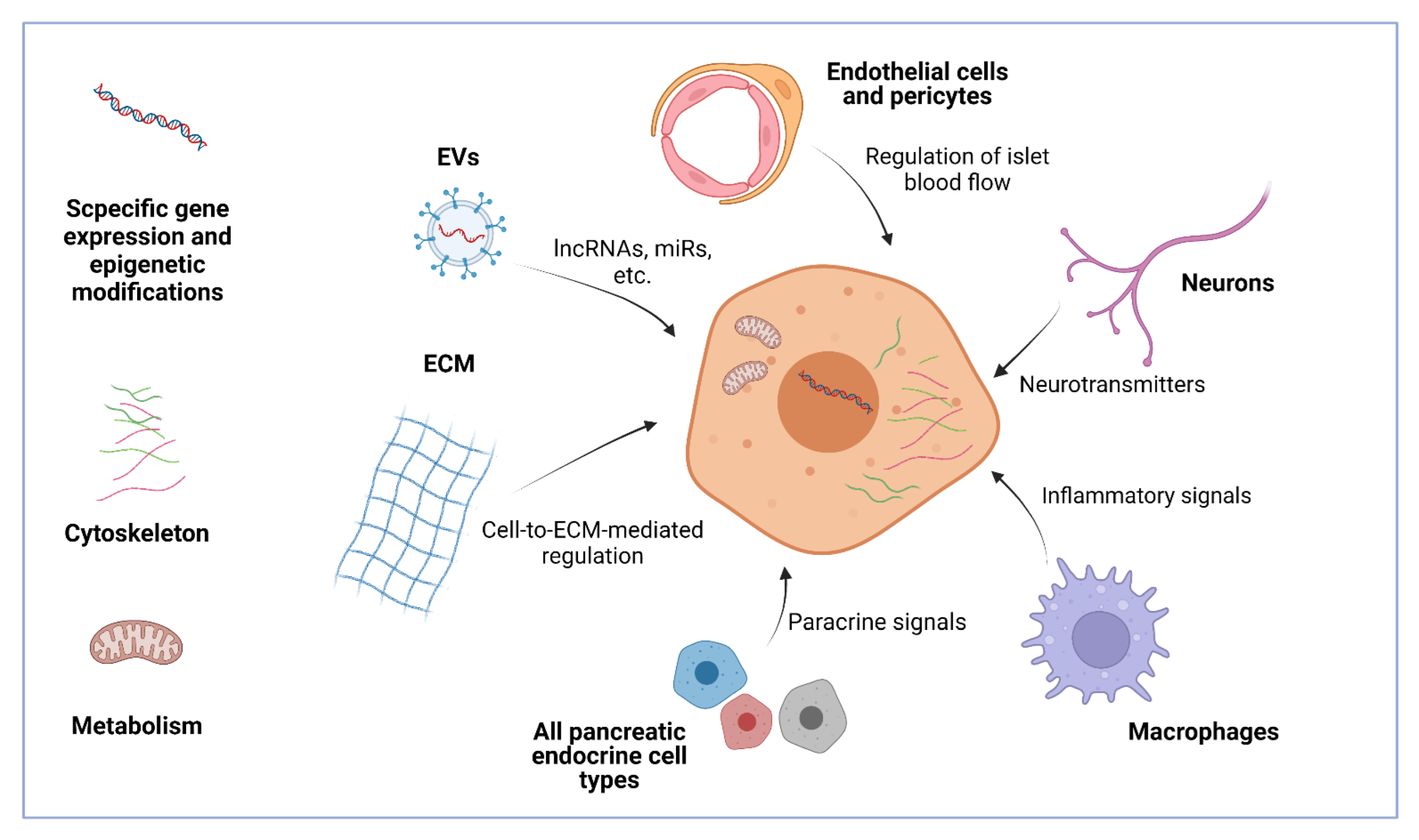
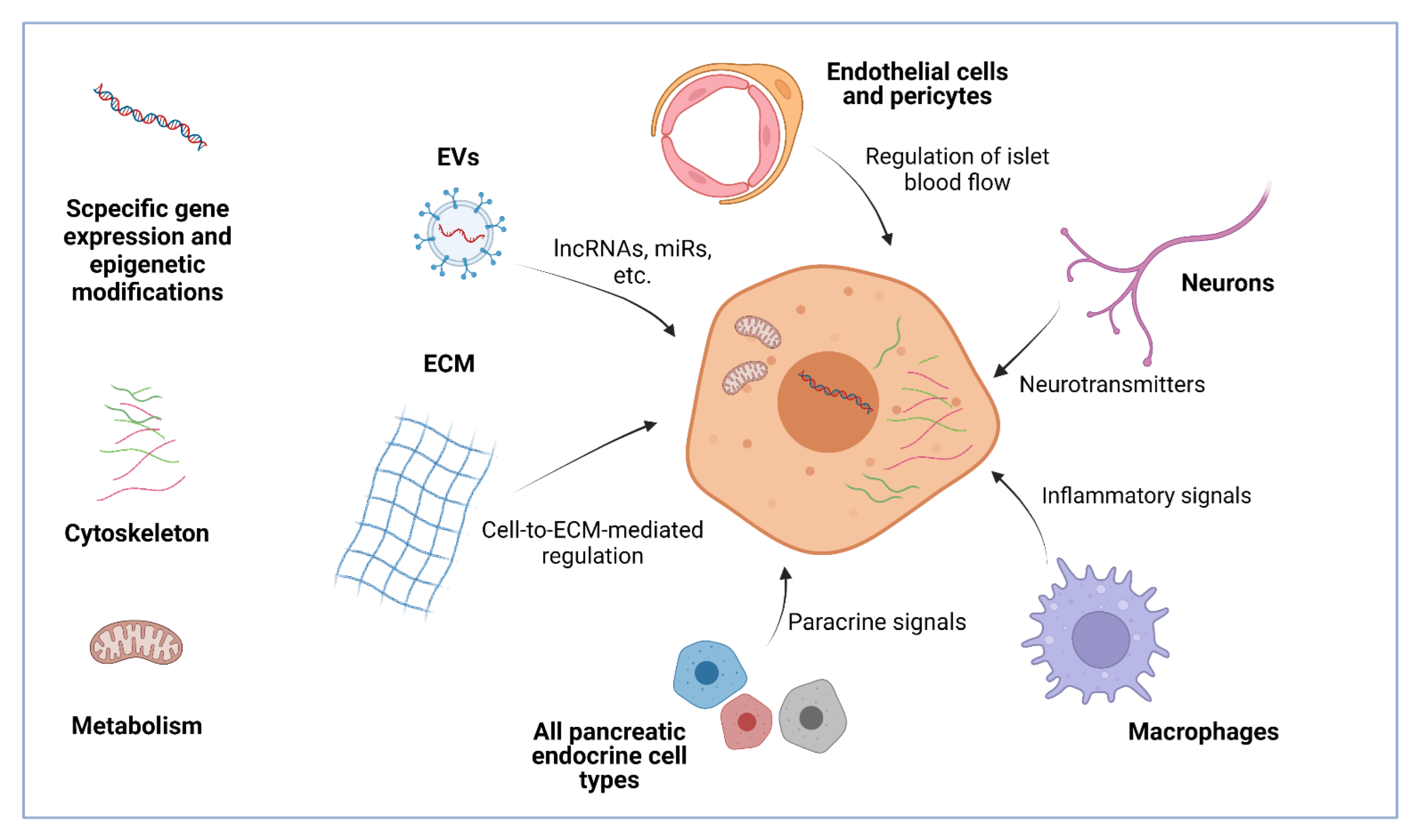{kind=link}
| Method to Generate Beta-Cells | Advantages | Disadvantages | References |
|---|---|---|---|
| Progenitor cells | Pancreatic stem-cell population, self-regeneration of pancreatic islets | No direct evidence of presence in humans | [37,38,39] |
| Transdifferentiation | Cell availability, possibility for personalized cell therapy | Low differentiation efficiency, high cell heterogeneity, functional immaturity, low GSIS, absence of key beta cell markers | [17,18,19,20,21,22,23,24,25,26] |
| Direct differentiation (ESCs, PSCs) | Usage of undifferentiated cells, mimicking embryological development, relatively robust GSIS, low heterogeneity, possibility for personalized cell therapy | Signs of functional immaturity, differences in metabolic pathways | [32,33,34,35,36] |
| Direct differentiation protocol | Advancement | ||
| D’Amour et al., 2005 [16] | Efficient differentiation of hESCs to definitive endoderm | ||
| Chen et al., 2009 [29] | Generation of pancreatic PDX1+-progenitors from hESCs | ||
| Nostro et al., 2011 [30] | Roles of TGF-b, WNT, nodal/activin A, and BMP (bone morphogenetic protein) signaling in the pancreatic lineage cell specification | ||
| Rezania et al., 2014 [32]; Pagliuca et al., 2014 [33] | Generation of stem cell-derived beta cells closely resembling native beta cells | ||
| Russ et al., 2015 [34] | Omission of BMP inhibitors during pancreatic specification results in higher yield of PDX1+/NKX6.1+-cells | ||
| Nair et al., 2019 [35] | Endocrine cell clustering mimics pancreatic organogenesis and promotes beta cell differentiation | ||
| Liu et al., 2021 [36] | New combination of chemicals and 3D pancreatic progenitor clusters promote beta cell functions | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karimova, M.V.; Gvazava, I.G.; Vorotelyak, E.A. Overcoming the Limitations of Stem Cell-Derived Beta Cells. Biomolecules 2022, 12, 810. https://doi.org/10.3390/biom12060810
Karimova MV, Gvazava IG, Vorotelyak EA. Overcoming the Limitations of Stem Cell-Derived Beta Cells. Biomolecules. 2022; 12(6):810. https://doi.org/10.3390/biom12060810
Chicago/Turabian StyleKarimova, Mariana V., Inessa G. Gvazava, and Ekaterina A. Vorotelyak. 2022. "Overcoming the Limitations of Stem Cell-Derived Beta Cells" Biomolecules 12, no. 6: 810. https://doi.org/10.3390/biom12060810
APA StyleKarimova, M. V., Gvazava, I. G., & Vorotelyak, E. A. (2022). Overcoming the Limitations of Stem Cell-Derived Beta Cells. Biomolecules, 12(6), 810. https://doi.org/10.3390/biom12060810