
Anti-Planktonic and Anti-Biofilm Properties of Pentacyclic Triterpenes—Asiatic Acid and Ursolic Acid as Promising Antibacterial Future Pharmaceuticals


Abstract
1. Introduction
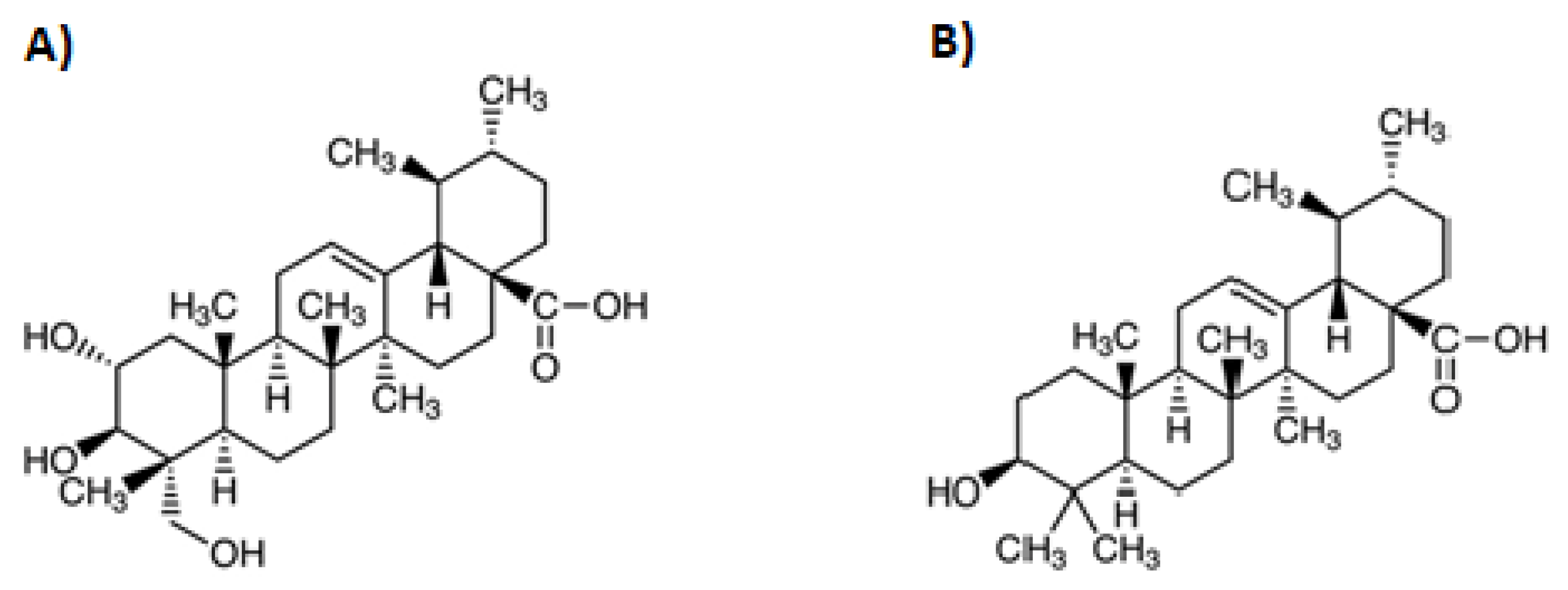
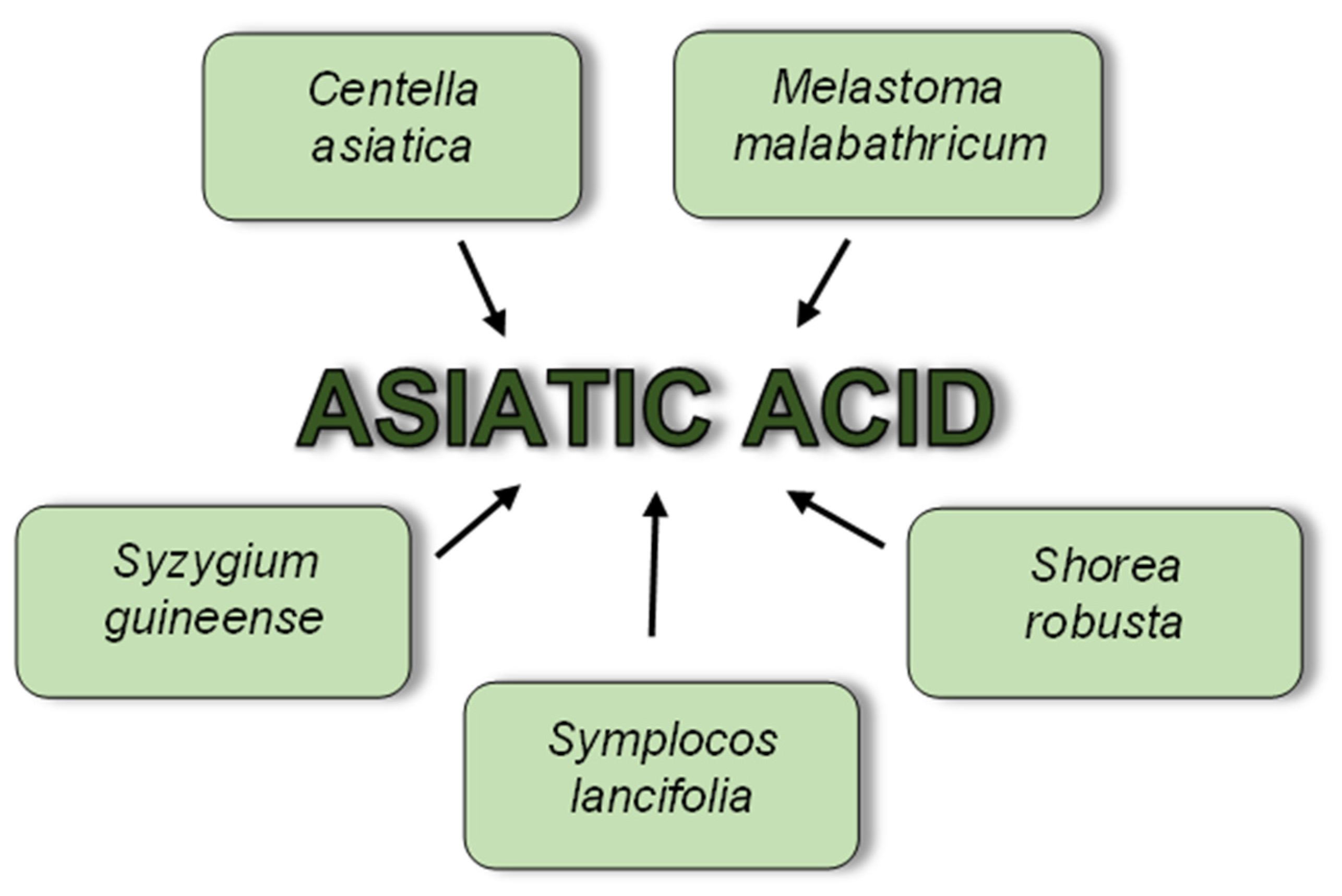
2. Asiatic Acid
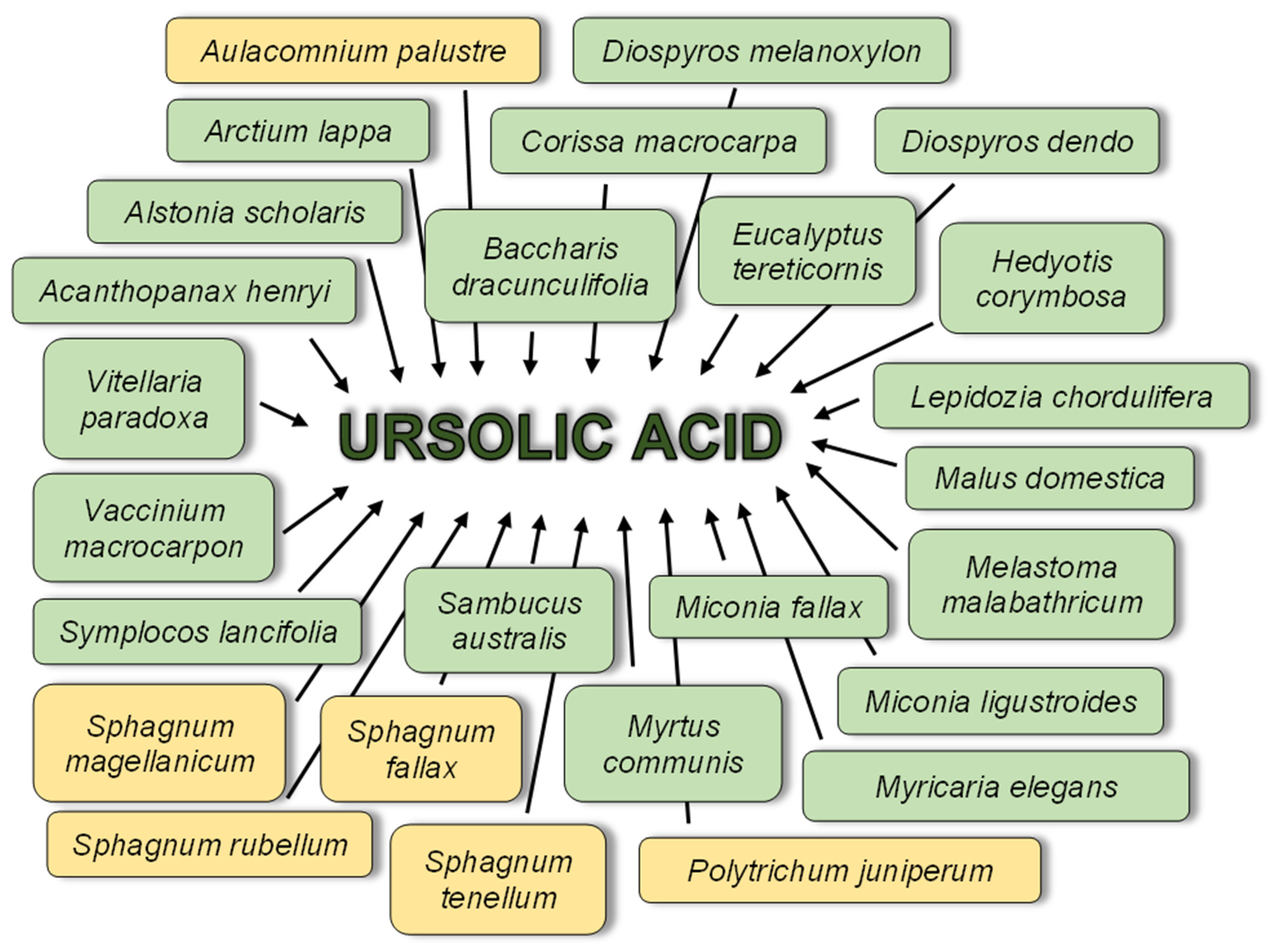
3. Ursolic Acid
4. Pharmacological Activities of AA and UA
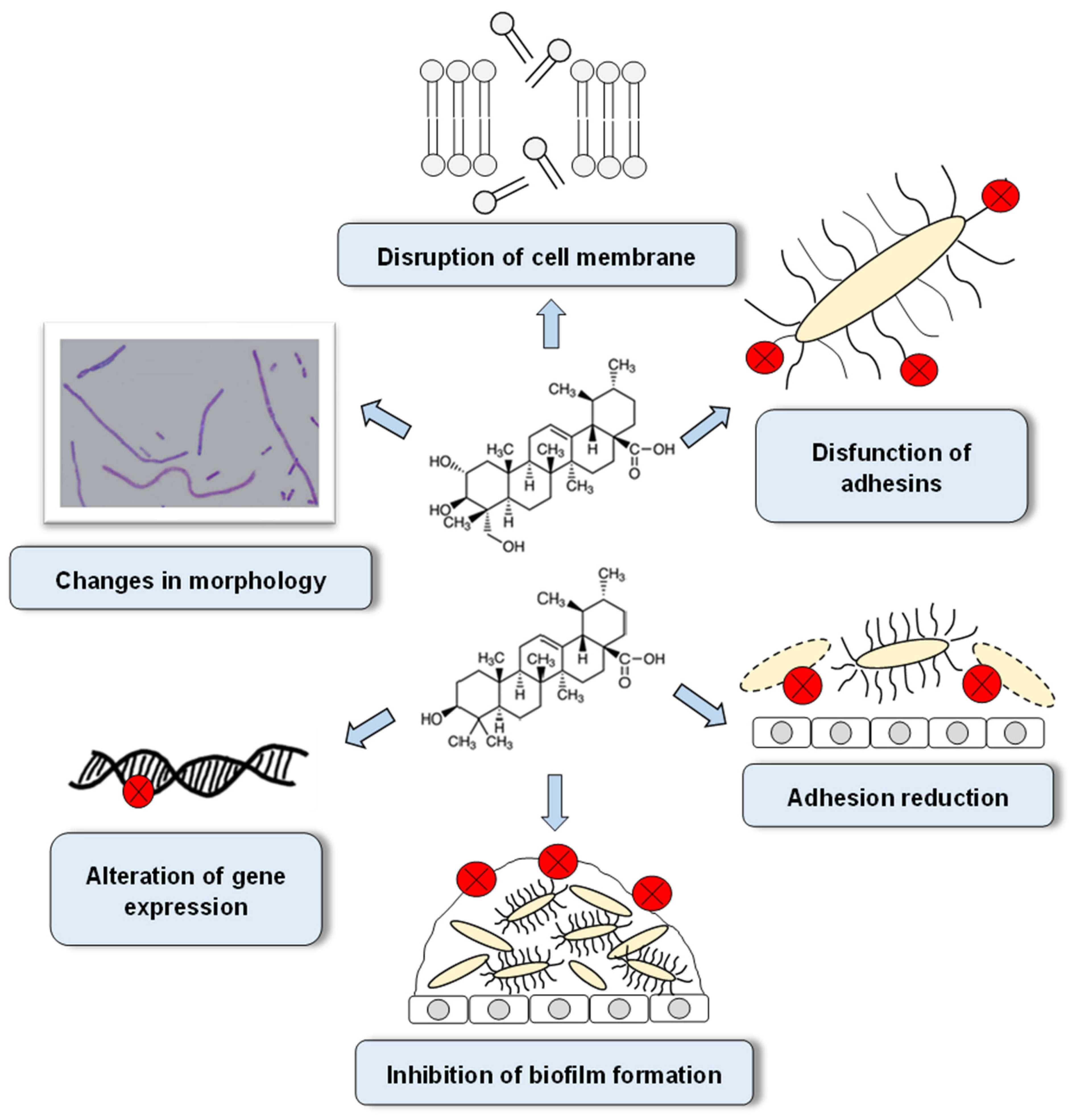
5. The Mechanisms of Antibacterial Activity of AA and UA
6. Antibacterial Properties of Pentacyclic Triterpenes
6.1. Anti-Planktonic Activity of Asiatic Acid

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Group | Species | Asiatic Acid | Ref. | ||
|---|---|---|---|---|---|
| MIC [mg/L] | Source | Antibacterial Activity | |||
| Gram-positive | Bacillus cereus | 36–44 | purchased *B | good | [50] |
| Bacillus subtilis | 0.75 | S. guineense | very good | [60] | |
| Clostridium difficile | 10–20 | purchased *A | good | [11] | |
| Enterococcus faecalis | 18–22 | purchased *B | good | [50] | |
| 64–128 | purchased *A | good–weak | [46] | ||
| 128 | S. lancifolia | weak | [63] | ||
| Listeria monocytogenes | 32–40 | purchased *B | good | [50] | |
| Staphylococcus aureus | 20–160 | C. asiatica | good–weak | [61] | |
| 26–30 | purchased *B | good | [50] | ||
| >128 | S. lancifolia | weak | [63] | ||
| Gram-negative | Enterobacter cloacae | 1024 | purchased *A | weak | [9] |
| Escherichia coli | 5 | S. guineense | very good | [60] | |
| 20–28 | purchased *B | good | [50] | ||
| >128 | S. lancifolia | weak | [63] | ||
| 512–>1024 | purchased *A | weak | [10] | ||
| 512–>1024 | purchased *A | weak | [47] | ||
| >1024 | purchased *A | weak | [9] | ||
| Pseudomonas aeruginosa | 32–40 | purchased *B | good | [50] | |
| 64 | S. robusta | good | [62] | ||
| >128 | purchased **A | weak | [64] | ||
| >128 | S. lancifolia | weak | [63] | ||
| >1024 | purchased *A | weak | [9] | ||
| Salmonella typhimurium | 30–34 | purchased *B | good | [50] | |
| Shigella sonnei | 30 | S. guineense | good | [60] | |
| Bacterial Group | Species | Inhibition Zone [mm] | Asiatic Acid | Ref. | ||
|---|---|---|---|---|---|---|
| Concentration [mg/L] | Source | Antibacterial Activity | ||||
| Gram-positive | Bacillus cereus | 10.0 | 250 | M. malabathricum | active | [66] |
| 11.5 | 500 | active | ||||
| 12.0 | 1000 | active | ||||
| 13.5 | 2000 | very active | ||||
| Bacillus subtilis | 7.0 | 25 | C. asiatica | active | [67] | |
| 9.0 | 50 | active | ||||
| 15.0 | 75 | very active | ||||
| 17.0 | 100 | very active | ||||
| Staphylococcus aureus | 8.0 | 1000 | purchased *C | active | [68] | |
| 9.0 | 250 | M. malabathricum | active | [66] | ||
| 10.0 | 500 | active | ||||
| 10.5 | 1000 | active | ||||
| 11.0 | 2000 | active | ||||
| 7.0 | 25 | C. asiatica | active | [67] | ||
| 8.0 | 50 | active | ||||
| 12.0 | 75 | active | ||||
| 13.0 | 100 | very active | ||||
| Streptococcus pneumoniae | 7.0 | 1000 | purchased *C | active | [68] | |
| Gram-negative | Escherichia coli | 7.0 | 1000 | purchased *C | active | [68] [62] |
| 7.0 | 64 | S. robusta | active | |||
| Helicobacter pylori | 12.0 | 1000 | purchased *C | active | [68] | |
| 8.0 | 500 | active | ||||
| Klebsiella pneumoniae | 8.0 | 1000 | M. malabathricum | active | [66] | |
| 9.0 | 2000 | active | ||||
| 13.0 | 25 | C. asiatica | very active | [67] | ||
| 23.0 | 50 | very active | ||||
| 26.0 | 75 | very active | ||||
| 28.0 | 100 | very active | ||||
| Pseudomonas aeruginosa | 8.0 | 64 | S. robusta | active | [62] | |
| 6.0 | 75 | C. asiatica | inactive | [67] | ||
| 6.0 | 100 | |||||
6.2. Anti-Planktonic Activity of Ursolic Acid
| Bacterial Group | Species | Ursolic Acid | Ref. | ||
|---|---|---|---|---|---|
| MIC [mg/L] | Source | Antibacterial Activity | |||
| Gram-positive | Actinomyces naeslundii | 16 | purchased *A | good | [89] |
| Actinomyces viscosus | 32 | purchased *A | good | [89] | |
| Bacillus cereus | 8 | A. scholaris | very good | [69] | |
| 20 | M. ligustroides | good | [80] | ||
| ≥1024 | S. australis | weak | [82] | ||
| Bacillus sphaericus | 50 | D. melanoxylon | good | [79] | |
| Bacillus subtilis | 25 | D. melanoxylon | good | [79] | |
| Enterococcus faecalis | 1 | A. scholaris | very good | [69] | |
| 4 | S. officinalis | very good | [70] | ||
| 4–256 | purchased *A | very good–weak | [72] | ||
| 16 | S. lancifolia | good | [63] | ||
| 32–512 | purchased *A | good–weak | [46] | ||
| 50 | M. fallax | good | [87] | ||
| 250 | M. ligustroides | weak | [80] | ||
| Enterococcus faecium | 4 | S. officinalis | very good | [70] | |
| 256 | purchased *A | weak | [72] | ||
| 500 | C. macrocarpa | weak | [88] | ||
| Listeria monocytogenes | 2 | A. scholaris | very good | [69] | |
| 6.5 | purchased *A | very good | [76] | ||
| 8 | purchased *A | very good | [75] | ||
| 8 | purchased *A | very good | [77] | ||
| ≥1024 | S. australis | weak | [82] | ||
| Staphylococcus aureus | 4–8 | purchased *A | very good | [74] | |
| 7.8 and 15.6 | purchased *A | very good, good | [73] | ||
| 8 | S. officinalis | very good | [70] | ||
| 8–16 | V. paradoxa | very good–good | [78] | ||
| 8–>256 | purchased *A | very good–weak | [72] | ||
| 10 | B. dracunculifolia | good | [81] | ||
| 10 | purchased *A | good | [75] | ||
| 16 | A. scholaris | good | [69] | ||
| 32 and ≥1024 | S. australis | good, weak | [82] | ||
| 37 | natural products | good | [85] | ||
| 50 | D. melanoxylon | good | [79] | ||
| 60 | natural products | good | [86] | ||
| 64 | A. scholaris | good | [69] | ||
| 64 | purchased *A | good | [83] | ||
| 64 | purchased **A | good | [84] | ||
| 64 | purchased *A | good | [54] | ||
| 128 | S. lancifolia | weak | [63] | ||
| 250 | C. macrocarpa | weak | [88] | ||
| 250 | purchased *A | weak | [90] | ||
| Staphylococcus epidermidis | 7.5 | purchased *A | very good | [75] | |
| Staphylococcus saprophyticus | 250 | C. macrocarpa | weak | [88] | |
| Streptococcus mitis | 50 | M. fallax | good | [87] | |
| Streptococcus gordonii | 7.8 | purchased ***A | very good | [14] | |
| 64 | purchased *A | good | [89] | ||
| Streptococcus mutans | 2–4 | purchased *A | very good | [71] | |
| 7.8 | purchased ***A | very good | [14] | ||
| 80 | M. fallax | good | [87] | ||
| 128–256 | purchased *A | weak | [91] | ||
| 250 | purchased ****A | weak | [13] | ||
| 256 | purchased *A | weak | [89] | ||
| 1024 | purchased *A | weak | [94] | ||
| Streptococcus pneumoniae | 8 | S. officinalis | very good | [70] | |
| 1000 | M. ligustroides | weak | [80] | ||
| Streptococcus salivarius | 50 | M. fallax | good | [87] | |
| Streptococcus sanguinis | 7.8 | purchased ***A | very good | [14] | |
| 50 | M. fallax | good | [87] | ||
| 128 | purchased *A | weak | [89] | ||
| Streptococcus sobrinus | 2–4 | purchased *A | very good | [71] | |
| 50 | M. fallax | good | [87] | ||
| 64 | purchased *A | good | [89] | ||
| 128 | purchased *A | weak | [91] | ||
| Gram-negative | Aeromonas caveae | ≥1024 | S. australis | weak | [82] |
| Escherichia coli | 50 | D. melanoxylon | good | [79] | |
| 64 and 512 | S. australis | good, weak | [82] | ||
| >128 | S. officinalis | weak | [70] | ||
| >128 | S. lancifolia | weak | [63] | ||
| >128 | A. scholaris | weak | [69] | ||
| 250 | C. macrocarpa | weak | [88] | ||
| 256 | purchased **A | weak | [84] | ||
| 256 | purchased *A | weak | [93] | ||
| >256 | purchased *A | weak | [72] | ||
| >256 | V. macrocarpon | weak | [92] | ||
| 512–>1024 | purchased *A | weak | [10] | ||
| 512–>1024 | purchased *A | weak | [47] | ||
| 1000 | E. tereticornis | weak | [95] | ||
| (1024 | S. australis | weak | [82] | ||
| Klebsiella pneumoniae | 64 | S. australis | good | [82] | |
| 500 | C. macrocarpa | weak | [88] | ||
| 512 | purchased *A | weak | [93] | ||
| 1000 | M. ligustroides | weak | [80] | ||
| Pseudomonas aeruginosa | 22.5 | purchased *A | good | [75] | |
| >128 | S. officinalis | weak | [70] | ||
| >128 | S. lancifolia | weak | [63] | ||
| >128 | A. scholaris | weak | [69] | ||
| 256 | purchased **A | weak | [84] | ||
| ≥256 | purchased *A | weak | [72] | ||
| 500 | C. macrocarpa | weak | [88] | ||
| 512 | S. australis | weak | [82] | ||
| Pseudomonas syringae | 25 | D. melanoxylon | good | [79] | |
| Salmonella choleraesuis | 1000 | M. ligustroides | weak | [80] | |
| Salmonella enterica | >128 | A. scholaris | weak | [69] | |
| Salmonella typhi | 50 | D. melanoxylon | good | [79] | |
| Serratia marcescens | >128 | S. officinalis | weak | [70] | |
| Shigella flexneri | 64 | S. australis | good | [82] | |
| Vibrio cholerae | >1000 | M. ligustroides | weak | [80] | |
| ≥1024 | S. australis | weak | [82] | ||
| Bacterial Group | Species | Inhibition Zone [mm] | Ursolic acid | Ref | ||
|---|---|---|---|---|---|---|
| Concentration [mg/L] | Source | Antibacterial Activity | ||||
| Gram-positive | Bacillus cereus | 7.0 | 250 | M. malabathricum | active | [66] |
| 7.0 | 500 | active | ||||
| 7.0 | 1000 | active | ||||
| 7.0 | 2000 | active | ||||
| Bacillus subtilis | 8.0 | 250 | M. malabathricum | active | [66] | |
| 9.0 | 500 | active | ||||
| 9.5 | 1000 | active | ||||
| 10.5 | 2000 | active | ||||
| Staphylococcus aureus | 21.0 | 100 | H. corymbosa | very active | [96] | |
| 7.5 | 250 | M. malabathricum | active | [66] | ||
| 8.5 | 500 | active | ||||
| 9.5 | 1000 | active | ||||
| 10.5 | 2000 | active | ||||
| Gram-negative | Pseudomonas aeruginosa | 10.0 | 100 | H. corymbosa | active | [96] |
| Salmonella typhi | 9.5 | 250 | M. malabathricum | active | [66] | |
| 10.0 | 500 | active | ||||
| 10.0 | 1000 | active | ||||
| 11.0 | 2000 | active | ||||
| Shigella boydii | 19.0 | 100 | H. corymbosa | very active | [96] | |
7. Anti-Biofilm Properties of Pentacyclic Triterpenes
7.1. Activity of Asiatic Acid against Bacterial Biofilms
7.2. Activity of Ursolic Acid against Bacterial Biofilms
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bjarnsholt, T. Introduction to biofilms. In Biofilm Infections; Bjarnsholt, T., Jensen, P., Moser, C., Høiby, N., Eds.; Springer: New York, NY, USA, 2011; pp. 1–9. [Google Scholar]
- Elias, S.; Banin, E. Multi-species biofilms: Living with friendly neighbors. FEMS Microbiol. Rev. 2012, 36, 990–1004. [Google Scholar] [CrossRef]
- Rendueles, O.; Ghigo, J.M. Multi-species biofilms: How to avoid unfriendly neighbors. FEMS Microbiol. Rev. 2012, 36, 972–989. [Google Scholar] [CrossRef]
- Gebreyohannes, G.; Nyerere, A.; Bii, C.; Sbhatu, D.B. Challenges of intervention, treatment, and antibiotic resistance of biofilm-forming microorganisms. Heliyon 2019, 5, e02192. [Google Scholar] [CrossRef]
- Olsen, I. Biofilm-specific antibiotic tolerance and resistance. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 877–886. [Google Scholar] [CrossRef]
- Ramos-Vivas, J.; Chapartegui-González, I.; Fernández-Martínez, M.; González-Rico, C.; Fortún, J.; Escudero, R.; Marco, F.; Linares, L.; Montejo, M.; Aranzamendi, M.; et al. Biofilm formation by multidrug resistant Enterobacteriaceae strains isolated from solid organ transplant recipients. Sci. Rep. 2019, 9, 8928. [Google Scholar] [CrossRef]
- Gollan, B.; Grabe, G.; Michaux, C.; Helaine, S. Bacterial persisters and infection: Past, present, and progressing. Annu. Rev. Microbiol. 2019, 73, 359–385. [Google Scholar] [CrossRef]
- Brady, A.J.; Laverty, G.; Gilpin, D.F.; Kearney, P.; Tunney, M. Antibiotic susceptibility of planktonic- and biofilm-grown staphylococci isolated from implant-associated infections: Should MBEC and nature of biofilm formation replace MIC? J. Med. Microbiol. 2017, 66, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Sycz, Z.; Tichaczek-Goska, D.; Jezierska-Domaradzka, A.; Wojnicz, D. Are uropathogenic bacteria living in multispecies biofilm susceptible to active plant ingredient—Asiatic acid? Biomolecules 2021, 11, 1754. [Google Scholar] [CrossRef]
- Wojnicz, D.; Kicia, M.; Tichaczek-Goska, D. Effect of asiatic and ursolic acids on morphology, hydrophobicity and adhesion of UPECs to uroepithelial cells. Folia Microbiol. 2013, 58, 245–252. [Google Scholar]
- Harnvoravongchai, P.; Chankhamhaengdecha, S.; Ounjai, P.; Singhakaew, S.; Boonthaworn, K.; Janvilisri, T. Antimicrobial effect of asiatic acid against Clostridium difficile is associated with disruption of membrane permeability. Front. Microbiol. 2018, 9, 2125. [Google Scholar] [CrossRef] [PubMed]
- Chi, J.; Sun, L.; Cai, L.; Fan, L.; Shao, C.; Shang, L.; Zhao, Y. Chinese herb microneedle patch for wound healing. Bioact. Mater. 2021, 6, 3507–3514. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, Y.; Fan, C.; Chi, Z.; Bai, M.; Sun, L.; Yang, L.; Yu, C.; Song, Z.; Yang, X.; et al. Ursolic acid targets glucosyltransferase and inhibits its activity to prevent Streptococcus mutans biofilm formation. Front. Microbiol. 2021, 12, 743305. [Google Scholar] [CrossRef] [PubMed]
- Lyu, X.; Wang, L.; Shui, Y.; Jiang, Q.; Chen, L.; Yang, W.; He, X.; Zeng, J.; Li, Y. Ursolic acid inhibits multi-species biofilms developed by Streptococcus mutans, Streptococcus sanguinis, and Streptococcus gordonii. Arch. Oral Biol. 2021, 125, 105107. [Google Scholar] [CrossRef] [PubMed]
- James, J.T.; Dubery, I.A. Pentacyclic triterpenoids from the medicinal herb, Centella asiatica (L.) Urban. Molecules 2009, 14, 3922–3941. [Google Scholar] [CrossRef]
- Jäger, S.; Trojan, H.; Kopp, T.; Laszczyk, M.N.; Scheffler, A. Pentacyclic triterpene distribution in various plants—Rich sources for a new group of multi-potent plant extracts. Molecules 2009, 14, 2016–2031. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Wu, L.; Wu, Y.; Zhang, C.; Qin, L.; Hayashi, M.; Kudo, M.; Gao, M.; Liu, T. Therapeutic potential of Centella asiatica and its triterpenes: A review. Front. Pharm. 2020, 11, 568032. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Sharma, A.; Zhang, T.; Wu, Y.; Ding, X. Pharmacological review on asiatic acid and its derivatives: A potential compound. SLAS Technol. 2018, 23, 111–127. [Google Scholar] [CrossRef]
- Nagoor Meeran, M.F.; Goyal, S.N.; Suchal, K.; Sharma, C.; Patil, C.R.; Ojha, S.K. Pharmacological properties, molecular mechanisms, and pharmaceutical development of asiatic acid: A pentacyclic triterpenoid of therapeutic promise. Front. Pharm. 2018, 9, 892. [Google Scholar] [CrossRef]
- González-Coloma, A.; López-Balboa, C.; Santana, O.; Reina, M.; Fraga, B. Triterpene-based plant defenses. Phytochem. Rev. 2011, 10, 245–260. [Google Scholar] [CrossRef]
- Bylka, W.; Znajdek-Awizeń, P.; Studzińska-Sroka, E.; Dańczak-Pazdrowska, A.; Brzezińska, M. Centella asiatica in dermatology: An overview. Phytother. Res. 2014, 28, 1117–1124. [Google Scholar] [CrossRef]
- Chandrika, U.G.; Prasad Kumarab, P.A. Gotu Kola (Centella asiatica): Nutritional properties and plausible health benefits. Adv. Food Nutr. Res. 2015, 76, 125–157. [Google Scholar]
- Kashyap, D.; Tuli, H.S.; Sharma, A.K. Ursolic acid (UA): A metabolite with promising therapeutic potential. Life Sci. 2016, 146, 201–213. [Google Scholar] [CrossRef] [PubMed]
- López-Hortas, L.; Pérez-Larrán, P.; González-Muñoz, M.J.; Falquéb, E.; Domíngueza, H. Recent developments on the extraction and application of ursolic acid. A review. Food Res. Int. 2018, 103, 130–149. [Google Scholar] [CrossRef]
- Pironi, A.M.; de Araújo, P.R.; Fernandes, M.A.; Regina, H.; Salgado, N.; Chorilli, M. Characteristics, biological properties and analytical methods of ursolic acid: A review. Crit. Rev. Anal. Chem. 2018, 48, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Mlala, S.; Oyedeji, A.O.; Gondwe, M.; Oyedeji, O.O. Ursolic acid and its derivatives as bioactive agents. Molecules 2019, 24, 2751. [Google Scholar] [CrossRef]
- Klavina, L.; Springe, G.; Nikolajeva, V.; Martsinkevich, I.; Nakurte, I.; Dzabijeva, D.; Steinberga, I. Chemical composition analysis, antimicrobial activity and cytotoxicity screening of moss extracts (moss phytochemistry). Molecules 2015, 20, 17221–17243. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Liu, P.; Shen, Y.M.; Dai, C. Studies on the chemical constituents from herb of Rhodobryum roseum [Chinese]. Zhongguo Zhong Yao Za Zhi 2005, 30, 895–897. [Google Scholar]
- Yang, S.; Liu, M.; Liang, N.; Zhao, Q.; Zhang, Y.; Xue, W.; Yang, S. Discovery and antitumor activities of constituents from Cyrtomium fortumei (J.) Smith rhizomes. Chem. Cent. J. 2013, 7, 24. [Google Scholar] [CrossRef][Green Version]
- Lee, J.S.; Miyashiro, H.; Nakamura, N.; Hattori, M. Two new triterpenes from the rhizome of Dryopteris crassirhizoma, and inhibitory activities of its constituents on human immunodeficiency virus-1 protease. Chem. Pharm. Bull. 2008, 56, 711–714. [Google Scholar] [CrossRef]
- Jaroniewski, W. Medicinal plants of Polish forests. Bearberry Arctostaphylos uva-ursi (L.) Sprengl. Wszechświat 1986, 87, 135–136. (In Polish) [Google Scholar]
- Mohd Azman, N.A.; Gallego, M.G.; Segovia, F.; Abdullah, S.; Shaarani, S.M.; Almajano Pablos, M.P. Study of the properties of bearberry leaf extract as a natural antioxidant in model foods. Antioxidants 2016, 5, 11. [Google Scholar] [CrossRef]
- Allen, D.; Bilz, M.; Leaman, D.J.; Miller, R.M.; Timoshyna, A.; Window, J. European Red List of Medicinal Plants; Publications Office of the European Union: Luxembourg, 2014. [Google Scholar]
- Chaika, N.; Koshovyi, O.; Raal, A.; Kireyev, I.; Zupanets, A.; Odyntsova, V. Phytochemical profile and pharmacological activity of the dry extract from Arctostaphylos uva-ursi leaves modified with phenylalanine. Sci. Pharm. Sci. 2020, 6, 77–84. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, C.; Zhao, X.; Wang, D.; Liu, Y.; Sun, S. Antifungal activity and potential mechanism of asiatic acid alone and in combination with fluconazole against Candida albicans. Biomed. Pharm. 2021, 139, 111568. [Google Scholar] [CrossRef]
- Zhu, Z.; Cui, L.; Yang, J.; Vong, C.T.; Hu, Y.; Xiao, J.; Chan, G.; He, Z.; Zhong, Z. Anticancer effects of asiatic acid against doxorubicin-resistant breast cancer cells via an AMPK-dependent pathway in vitro. Phytomedicine 2021, 92, 153737. [Google Scholar] [CrossRef] [PubMed]
- Thanusha, A.V.; Dinda, A.K.; Koul, V. Evaluation of nano hydrogel composite based on gelatin/HA/CS suffused with asiatic acid/ZnO and CuO nanoparticles for second degree burns. Mater. Sci. Eng. C Mater. Biol. Appl. 2018, 89, 378–386. [Google Scholar]
- Yuyun, X.; Xi, C.; Qing, Y.; Lin, X.; Ke, R.; Bingwei, S. Asiatic acid attenuates lipopolysaccharide-induced injury by suppressing activation of the Notch signaling pathway. Oncotarget 2018, 9, 15036–15046. [Google Scholar] [CrossRef]
- Han, Y.; Jiang, Y.; Li, Y.; Wang, M.; Fan, T.; Liu, M.; Ke, Q.; Xu, H.; Yi, Z. An aligned porous electrospun fibrous scaffold with embedded asiatic acid for accelerating diabetic wound healing. J. Mater. Chem. B 2019, 7, 6125–6138. [Google Scholar] [CrossRef]
- Liew, K.Y.; Hafiz, M.F.; Chong, Y.J.; Harith, H.H.; Israf, D.A.; Tham, C.L. A review of malaysian herbal plants and their active constituents with potential therapeutic applications in sepsis. Evid. Based Complement. Altern. Med. 2020, 2020, 8257817. [Google Scholar] [CrossRef]
- Wolska, K.I.; Grudniak, A.M.; Fiecek, B.; Kraczkiewicz-Dowjat, A.; Kurek, A. Antibacterial activity of oleanolic and ursolic acids and their derivatives. Cent. Eur. J. Biol. 2010, 5, 543–553. [Google Scholar] [CrossRef]
- Jesus, J.A.; Lago, J.H.; Laurenti, M.D.; Yamamoto, E.S.; Passero, L.F.D. Antimicrobial activity of oleanolic and ursolic acids: An update. Evid. Based Complement. Altern. Med. 2015, 2015, 620472. [Google Scholar] [CrossRef]
- Woźniak, Ł.; Skąpska, S.; Marszałek, K. Ursolic acid—A pentacyclic triterpenoid with a wide spectrum of pharmacological activities. Molecules 2015, 20, 20614–20641. [Google Scholar] [CrossRef] [PubMed]
- Navina, R.; Lee, Y.G.; Kim, S.M. Molecular biological roles of ursolic acid in the treatment of human diseases. Curr. Bioact. Compd. 2017, 13, 177–185. [Google Scholar] [CrossRef]
- Huang, L.; Luo, H.; Li, Q.; Wang, D.; Zhang, J.; Hao, X.; Yang, X. Pentacyclic triterpene derivatives possessing polyhydroxyl ring A inhibit Gram-positive bacteria growth by regulating metabolism and virulence genes expression. Eur. J. Med. Chem. 2015, 95, 64–75. [Google Scholar] [CrossRef]
- Wojnicz, D.; Tichaczek-Goska, D.; Korzekwa, K.; Kicia, M.; Hendrich, A. Anti-enterococcal activities of pentacyclic triterpenes. Adv. Clin. Exp. Med. 2017, 26, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Wojnicz, D.; Tichaczek-Goska, D.; Kicia, M. Effect of asiatic and ursolic acids on growth and virulence factors of uropathogenic Escherichia coli strains. Turk. J. Biol. 2013, 37, 556–564. [Google Scholar] [CrossRef]
- Ford, M. Medical Microbiology; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Wojnicz, D.; Tichaczek-Goska, D.; Kicia, M. Pentacyclic triterpenes combined with ciprofloxacin help to eradicate the biofilm formed in vitro by Escherichia coli. Indian J. Med. Res. 2015, 141, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.H.; Liu, T.C.; Mong, M.C. Antibacterial effects and action modes of asiatic acid. Biomedicine 2015, 5, 22–29. [Google Scholar] [CrossRef]
- Broniatowski, M.; Mastalerz, P.; Flasiński, M. Studies of the interactions of ursane-type bioactive terpenes with the model of Escherichia coli inner membrane-Langmuir monolayer approach. Biochim. Biophys. Acta 2015, 1848, 469–476. [Google Scholar] [CrossRef]
- Gu, W.; Hao, Y.; Zhang, G.; Wang, S.F.; Miao, T.T.; Zhang, K.P. Synthesis, in vitro antimicrobial and cytotoxic activities of new carbazole derivatives of ursolic acid. Bioorg. Med. Chem. Lett. 2015, 25, 554–557. [Google Scholar] [CrossRef]
- Usmani, Y.; Ahmed, A.; Faizi, S.; Versiani, M.A.; Shamshad, S.; Khan, S.; Simjee, S.U. Antimicrobial and biofilm inhibiting potential of an amide derivative [N-(2′, 4′-dinitrophenyl)-3β-hydroxyurs-12-en-28-carbonamide] of ursolic acid by modulating membrane potential and quorum sensing against colistin resistant Acinetobacter Baumannii. Microb. Pathog. 2021, 157, 104997. [Google Scholar] [CrossRef]
- Ghasemzadeh, F.; Najafpour, G.D.; Mohammadi, M. Antiinfective properties of ursolic acid-loaded chitosan nanoparticles against Staphylococcus aureus. Turk. J. Chem. 2021, 45, 1454–1462. [Google Scholar] [CrossRef] [PubMed]
- Oprean, C.; Zambori, C.; Borcan, F.; Soica, C.; Zupko, I.; Minorics, R.; Bojin, F.; Ambrus, R.; Muntean, D.; Danciu, C.; et al. Anti-proliferative and antibacterial in vitro evaluation of the polyurethane nanostructures incorporating pentacyclic triterpenes. Pharm. Biol. 2016, 54, 2714–2722. [Google Scholar] [CrossRef]
- Dincer, S.; Uslu, F.M.; Delik, A. Antibiotic resistance in biofilm. In Bacterial Biofilms; Dincer, S., Ed.; IntechOpen: London, UK, 2020; pp. 1–8. [Google Scholar]
- Sánchez, M.C.; Romero-Lastra, P.; Ribeiro-Vidal, H.; Llama-Palacios, A.; Figuero, E.; Herrera, D.; Sanz, M. Comparative gene expression analysis of planktonic Porphyromonas gingivalis ATCC 33277 in the presence of a growing biofilm versus planktonic cells. BMC Microbiol. 2019, 19, 58. [Google Scholar] [CrossRef]
- Chen, H.; Wubbolts, R.W.; Haagsman, H.P.; Veldhuizen, E.J.A. Inhibition and eradication of Pseudomonas aeruginosa biofilms by host defence peptides. Sci. Rep. 2018, 8, 10446. [Google Scholar] [CrossRef] [PubMed]
- Bitchagno, G.T.M.; Nchiozem-Ngnitedem, V.A.; Wandji, N.T.; Noulala, G.C.T.; Fobofou, S.A.T.; Lenta, B.N. Plant-Derived Compounds Against Microbial Infections and Cancers. In Bioactive Compounds in Nutraceutical and Functional Food for Good Human Health; Sharma, K., Mishra, K., Senapati, K.K., Danciu, C., Eds.; IntechOpen Limited: London, UK, 2020; pp. 53–77. [Google Scholar]
- Djoukeng, J.D.; Abou-Mansour, E.; Tabacchi, R.; Tapondjou, A.L.; Bouda, H.; Lontsi, D. Antibacterial triterpenes from Syzygium guineense (Myrtaceae). J. Ethnopharmacol. 2005, 101, 283–286. [Google Scholar] [CrossRef] [PubMed]
- Taemchuay, D.; Rukkwamsuk, T.; Sakpuaram, T.; Ruangwises, N. Antibacterial activity of crude extracts of Centella asiatica against Staphylococcus aureus in bovine mastitis. Kasetsart Vet. 2009, 19, 119–128. [Google Scholar]
- Bharitkar, Y.P.; Banerjee, M.; Kumar, S.; Paira, R.; Meda, R.; Kuotsu, K.; Mondal, N.B. Search for a potent microbicidal spermicide from the isolates of Shorea robusta resin. Contraception 2013, 88, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Acebey-Castellon, I.L.; Voutquenne-Nazabadioko, L.; Doan Thi Mai, H.; Roseau, N.; Bouthagane, N.; Muhammad, D.; Le Magrex Debar, E.; Gangloff, S.C.; Litaudon, M.; Sevenet, T.; et al. Triterpenoid saponins from Symplocos lancifolia. J. Nat. Prod. 2011, 74, 163–168. [Google Scholar] [CrossRef]
- Garo, E.; Eldridge, G.R.; Goering, M.G.; DeLancey Pulcini, E.; Hamilton, M.A.; Costerton, J.W.; James, G.A. Asiatic acid and corosolic acid enhance the susceptibility of Pseudomonas aeruginosa biofilms to tobramycin. Antimicrob. Agents Chemother. 2007, 51, 1813–1817. [Google Scholar] [CrossRef]
- Mutai, C.; Bii, C.; Rukunga, G.; Ondicho, J.; Mwitari, P.; Abatis, D.; Vagias, C.; Roussis, V.; Kirui, J. Antimicrobial activity of pentacyclic triterpenes isolated from Acacia mellifera. Afr. J. Tradit. Complement. Altern. Med. 2009, 6, 42–48. [Google Scholar] [CrossRef][Green Version]
- Wong, K.C.; Hag Ali, D.M.; Boey, P.L. Chemical constituents and antibacterial activity of Melastoma malabathricum L. Nat. Prod. Res. 2012, 26, 609–618. [Google Scholar] [CrossRef]
- Ashella, S.; Fleming, A.T. Antimicrobial activity of asiatic acid against bacteria and fungi. IJSR Online 2016, 5, 920–921. [Google Scholar]
- Norzaharaini, M.G.; Wan Norshazwani, W.S.; Hasmah, A.; Nor Izani, N.J.; Rapeah, S. Preliminary study on antimicrobial activities of asiaticoside and asiatic acid against selected gram-positive and gram-negative bacteria. Health Environ. J. 2011, 2, 23–26. [Google Scholar]
- Wang, C.M.; Chen, H.T.; Wu, Z.Y.; Jhan, Y.L.; Shyu, C.L.; Chou, C.H. Antibacterial and synergistic activity of pentacyclic triterpenoids isolated from Alstonia scholaris. Molecules 2016, 21, 139. [Google Scholar] [CrossRef]
- Horiuchi, K.; Shiota, S.; Hatano, T.; Yoshida, T.; Kuroda, T.; Tsuchiya, T. Antimicrobial activity of oleanolic acid from Salvia officinalis and related compounds on vancomycin-resistant enterococci (VRE). Biol. Pharm. Bull. 2007, 30, 1147–1149. [Google Scholar] [CrossRef]
- Kim, M.J.; Kim, C.S.; Park, J.Y.; Lim, Y.K.; Park, S.N.; Ahn, S.J.; Jin, D.C.; Kim, T.H.; Kook, J.K. Antimicrobial effects of ursolic acid against mutans Streptococci isolated from Koreans. Int. J. Oral Biol. 2011, 36, 7–11. [Google Scholar]
- Fontanay, S.; Grare, M.; Mayer, J.; Finance, C.; Duval, R.E. Ursolic, oleanolic and betulinic acids: Antibacterial spectra and selectivity indexes. J. Ethnopharmacol. 2008, 120, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Filocamo, A.; Bisignano, C.; D’Arrigo, M.; Ginestra, G.; Mandalari, G.; Galati, E.M. Norfloxacin and ursolic acid: In vitro association and postantibiotic effect against Staphylococcus aureus. Lett. Appl. Microbiol. 2011, 53, 193–197. [Google Scholar] [CrossRef]
- Kim, S.G.; Kim, M.J.; Jin, D.; Park, S.N.; Cho, E.; Oliveira Freire, M.; Jang, S.J.; Park, Y.J.; Kook, J. K Antimicrobial effect of ursolic acid and oleanolic acid against methicillin-resistant Staphylococcus aureus. Korean J. Microbiol. 2012, 48, 212–215. [Google Scholar] [CrossRef][Green Version]
- Kurek, A.; Nadkowska, P.; Pliszka, S.; Wolska, K.I. Modulation of antibiotic resistance in bacterial pathogens by oleanolic acid and ursolic acid. Phytomedicine 2012, 19, 515–519. [Google Scholar] [CrossRef]
- Kurek, A.; Grudniak, A.M.; Szwed, M.; Klicka, A.; Samluk, Ł.; Wolska, K.I.; Janiszowska, W.; Popowska, M. Oleanolic acid and ursolic acid affect peptidoglycan metabolism in Listeria monocytogenes. Antonie Van Leeuwenhoek 2010, 97, 61–68. [Google Scholar] [CrossRef]
- Kurek, A.; Markowska, K.; Grudniak, A.M.; Janiszowska, W.; Wolska, K.I. The effect of oleanolic and ursolic acids on the hemolytic properties and biofilm formation of List Monocytogenes. Pol. J. Microbiol. 2014, 63, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Catteau, L.; Reichmann, N.T.; Olson, J.; Pinho, M.G.; Nizet, V.; Van Bambeke, F.; Quetin-Leclercq, J. Synergy between ursolic and oleanolic acids from Vitellaria paradoxa leaf extract and β-lactams against methicillin-resistant Staphylococcus aureus: In vitro and in vivo activity and underlying mechanisms. Molecules 2017, 22, 2245. [Google Scholar] [CrossRef]
- Mallavadhani, U.V.; Mahapatra, A.; Jamil, K.; Reddy, P.S. Antimicrobial activity of some pentacyclic triterpenes and their synthesized 3-O-lipophilic chains. Biol. Pharm. Bull. 2004, 27, 1576–1579. [Google Scholar] [CrossRef]
- Cunha, W.R.; de Matos, G.X.; Souza, M.G.; Tozatti, M.G.; Andrade e Silva, M.L.; Martins, C.H.G.; da Silva, R.; da Silva Filho, A.A. Evaluation of the antibacterial activity of the methylene chloride extract of Miconia ligustroides, isolated triterpene acids, and ursolic acid derivatives. Pharm. Biol. 2010, 48, 166–169. [Google Scholar] [CrossRef] [PubMed]
- da Silva Filho, A.A.; de Sousa, J.P.; Soares, S.; Furtado, N.A.; e Silva, M.L.; Cunha, W.R.; Gregório, L.E.; Nanayakkara, N.D.; Bastos, J.K. Antimicrobial activity of the extract and isolated compounds from Baccharis dracunculifolia D. C. (Asteraceae). Z. Nat. C. J. Bio. Sci. 2008, 63, 40–46. [Google Scholar]
- do Nascimento, P.G.G.; Lemos, T.L.G.; Bizerra, A.M.C.; Arriaga, A.M.C.; Ferreira, D.A.; Santiago, G.M.P.; Braz-Filho, R.; Costa, J.G.M. Antibacterial and antioxidant activities of ursolic acid and derivatives. Molecules 2014, 19, 1317–1327. [Google Scholar] [CrossRef]
- Wang, C.M.; Jhan, Y.L.; Tsai, S.J.; Chou, C.H. The pleiotropic antibacterial mechanisms of ursolic acid against methicillin-resistant Staphylococcus aureus (MRSA). Molecules 2016, 21, 884. [Google Scholar] [CrossRef] [PubMed]
- Oloyede, H.O.B.; Ajiboye, H.O.; Salawu, M.O.; Ajiboye, T.O. Influence of oxidative stress on the antibacterial activity of betulin, betulinic acid and ursolic acid. Microb. Pathog. 2017, 111, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Qin, N.; Tan, X.; Jiao, Y.; Liu, L.; Zhao, W.; Yang, S.; Jia, A. RNA-Seq-based transcriptome analysis of methicillin-resistant Staphylococcus aureus biofilm inhibition by ursolic acid and resveratrol. Sci. Rep. 2014, 4, 5467. [Google Scholar] [CrossRef]
- Tan, X.; Qin, N.; Wu, C.; Sheng, J.; Yang, R.; Zheng, B.; Ma, Z.; Liu, L.; Peng, X.; Jia, A. Transcriptome analysis of the biofilm formed by methicillin-susceptible Staphylococcus aureus. Sci. Rep. 2015, 5, 11997. [Google Scholar] [CrossRef]
- Scalon Cunha, L.C.; Andrade e Silva, M.L.; Cardoso Furtado, N.A.; Vinhólis, A.H.; Martins, C.H.; da Silva Filho, A.A.; Cunha, W.R. Antibacterial activity of triterpene acids and semi-synthetic derivatives against oral pathogens. Z. Nat. C J. Biosci. 2007, 62, 668–672. [Google Scholar]
- Moodley, R.; Chenia, H.; Jonnalagadda, S.B.; Koorbanally, N. Antibacterial and anti-adhesion activity of the pentacyclic triterpenoids isolated from the leaves and edible fruits of Carissa macrocarpa Antibacterial and anti-adhesion activity of the pentacyclic triterpenoids isolated from the leaves and edible fruits of Carissa macrocarpa. J. Med. Plant. Res. 2011, 5, 4851–4858. [Google Scholar]
- Zhou, L.; Ding, Y.; Chen, W.; Zhang, P.; Chen, Y.; Lv, X. The in vitro study of ursolic acid and oleanolic acid inhibiting cariogenic microorganisms as well as biofilm. Oral Dis. 2013, 19, 494–500. [Google Scholar] [CrossRef]
- Micota, B.; Sadowska, B.; Podsędek, A.; Redzynia, M.; Różalska, B. Leonurus cardiaca L. herb—A derived extract and an ursolic acid as the factors affecting the adhesion capacity of Staphylococcus aureus in the context of infective endocarditis. Acta Biochim. Pol. 2014, 61, 385–388. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Lee, Y.; Huh, J.; Park, J.W. Synergistic effect of xylitol and ursolic acid combination on oral biofilms. Restor. Dent. Endod. 2014, 39, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Nikolic, D.; Pendland, S.; Doyle, B.J.; Locklear, T.D.; Mahady, G.B. Effects of cranberry extracts and ursolic acid derivatives on P-fimbriated Escherichia coli, COX-2 activity, pro-inflammatory cytokine release and the NF-kappabeta transcriptional response in vitro. Pharm. Biol. 2009, 47, 18–25. [Google Scholar] [CrossRef]
- Sundaramoorthy, N.S.; Mohan, H.M.; Subramaniam, S.; Raman, T.; Ganesan, S.S.; Sivasubamanian, A.; Nagarajan, S. Ursolic acid inhibits colistin efflux and curtails colistin resistant Enterobacteriaceae. AMB Express 2019, 9, 27. [Google Scholar] [CrossRef]
- Chung, S.H.; Cho, S.; Kim, K.; Lim, B.S.; Ahn, S.J. Antimicrobial and physical characteristics of orthodontic primers containing antimicrobial agents. Angle Orthod. 2017, 87, 307–312. [Google Scholar] [CrossRef]
- Dwivedi, G.R.; Maurya, A.; Yadav, D.K.; Khan, F.; Darokar, M.P.; Srivastava, S.K. Drug resistance reversal potential of ursolic acid derivatives against nalidixic acid- and multidrug-resistant Escherichia coli. Chem. Biol. Drug Des. 2015, 86, 272–283. [Google Scholar] [CrossRef]
- Sultana, T.; Rashid, M.A.; Ali, M.A.; Mahmood, S.F. Hepatoprotective and antibacterial activity of ursolic acid extracted from Hedyotis corymbosa L. Bangladesh J. Sci. Ind. Res. 2010, 45, 27–34. [Google Scholar] [CrossRef]
- Ahmad, M.; Ahmad, W.; Khan, S.; Zeeshan, M.; Obaidullah Nisar, M.; Shaheen, F.; Ahmad, M. New antibacterial pentacyclic triterpenes from Myricaria elegans Royle (tamariscineae). J. Enzym. Inhib. Med. Chem. 2008, 23, 1023–1027. [Google Scholar] [CrossRef]
- Park, S.N.; Ahn, S.J.; Kook, J.K. Oleanolic acid and ursolic acid inhibit peptidoglycan biosynthesis in Streptococcus mutans UA159. Braz. J. Microbiol. 2015, 46, 613–617. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhou, T.; Li, Z.; Kang, O.H.; Mun, S.H.; Seo, Y.S.; Kong, R.; Shin, D.W.; Liu, X.Q.; Kwon, D.Y. Antimicrobial activity and synergism of ursolic acid 3-O-α-L-arabinopyranoside with oxacillin against methicillin-resistant Staphylococcus aureus. Int. J. Mol. Med. 2017, 40, 1285–1293. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jabeen, M.; Ahmad, S.; Shahid, K.; Sadiq, A.; Rashid, U. Ursolic acid hydrazide based organometallic complexes: Synthesis, characterization, antibacterial, antioxidant, and docking studies. Front. Chem. 2018, 6, 55. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Zuo, R.; Gonzalez Barrios, A.F.; Bedzyk, L.A.; Eldridge, G.R.; Pasmore, M.E.; Wood, T.K. Differential gene expression for investigation of Escherichia coli biofilm inhibition by plant extract ursolic acid. Appl. Environ. Microbiol. 2005, 71, 4022–4034. [Google Scholar] [CrossRef]
- Kim, S.; Song, M.; Roh, B.D.; Park, S.H.; Park, J.W. Inhibition of Streptococcus mutans biofilm formation on composite resins containing ursolic acid. Restor. Endod. 2013, 38, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Gilabert, M.; Marcinkevicius, K.; Andujar, S.; Schiavone, M.; Arena, M.E.; Bardón, A. Sesqui- and triterpenoids from the liverwort Lepidozia chordulifera inhibitors of bacterial biofilm and elastase activity of human pathogenic bacteria. Phytomedicine 2015, 22, 77–85. [Google Scholar] [CrossRef]
- Tielen, P.; Rosenau, F.; Wilhelm, S.; Jaeger, K.E.; Flemming, H.C.; Wingender, J. Extracellular enzymes affect biofilm formation of mucoid Pseudomonas aeruginosa. Microbiology 2010, 156, 2239–2252. [Google Scholar] [CrossRef]
- Yu, H.; He, X.; Xie, W.; Xiong, J.; Sheng, H.; Guo, S.; Huang, C.; Zhang, D.; Zhang, K. Elastase LasB of Pseudomonas aeruginosa promotes biofilm formation partly through rhamnolipid mediated regulation. Can. J. Microbiol. 2014, 60, 227–235. [Google Scholar] [CrossRef]
- Lou, Z.; Tang, Y.; Song, X.; Wang, H. Metabolomics-based screening of biofilm-inhibitory compounds against Pseudomonas aeruginosa from burdock leaf. Molecules 2015, 20, 16266–16277. [Google Scholar] [CrossRef]
- Feuillolay, C.; Pecastaings, S.; Le Gac, C.; Fiorini-Puybaret, C.; Luc, J.; Joulia, P.; Roques, C. A Myrtus communis extract enriched in myrtucummulones and ursolic acid reduces resistance of Propionibacterium acnes biofilms to antibiotics used in acne vulgaris. Phytomedicine 2016, 23, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Ray, C.; Shenoy, A.T.; Orihuela, C.J.; González-Juarbe, N. Killing of Serratia marcescens biofilms with chloramphenicol. Ann. Microbiol. Antimicrob. 2017, 16, 19. [Google Scholar] [CrossRef] [PubMed]
- Jyothi, J.S.; Putty, K.; Reddy, Y.N.; Dhanalakshmi, K.; Umair, M.A.H. Antagonistic effect of ursolic acid on Staphylococcal biofilms. Vet. World 2018, 11, 1440–1444. [Google Scholar] [CrossRef]
- Silva, G.N.S.; Primon-Barros, M.; Macedo, A.J.; Gnoatto, S.C.B. Triterpene derivatives as relevant scaffold for new antibiofilm drugs. Biomolecules 2019, 9, 58. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sycz, Z.; Tichaczek-Goska, D.; Wojnicz, D. Anti-Planktonic and Anti-Biofilm Properties of Pentacyclic Triterpenes—Asiatic Acid and Ursolic Acid as Promising Antibacterial Future Pharmaceuticals. Biomolecules 2022, 12, 98. https://doi.org/10.3390/biom12010098
Sycz Z, Tichaczek-Goska D, Wojnicz D. Anti-Planktonic and Anti-Biofilm Properties of Pentacyclic Triterpenes—Asiatic Acid and Ursolic Acid as Promising Antibacterial Future Pharmaceuticals. Biomolecules. 2022; 12(1):98. https://doi.org/10.3390/biom12010098
Chicago/Turabian StyleSycz, Zuzanna, Dorota Tichaczek-Goska, and Dorota Wojnicz. 2022. "Anti-Planktonic and Anti-Biofilm Properties of Pentacyclic Triterpenes—Asiatic Acid and Ursolic Acid as Promising Antibacterial Future Pharmaceuticals" Biomolecules 12, no. 1: 98. https://doi.org/10.3390/biom12010098
APA StyleSycz, Z., Tichaczek-Goska, D., & Wojnicz, D. (2022). Anti-Planktonic and Anti-Biofilm Properties of Pentacyclic Triterpenes—Asiatic Acid and Ursolic Acid as Promising Antibacterial Future Pharmaceuticals. Biomolecules, 12(1), 98. https://doi.org/10.3390/biom12010098