
A Runner’s High for New Neurons? Potential Role for Endorphins in Exercise Effects on Adult Neurogenesis


{kind=link}
Abstract
1. Exercise Effects on Brain and Mental Health
2. Hippocampus and Adult Neurogenesis—Potential Involvement in Many of the Exercise-Related Effects
3. Exercise Effects on Adult Neurogenesis
4. Functional Role of Neurogenesis on Exercise-Induced Changes to Mental Functioning
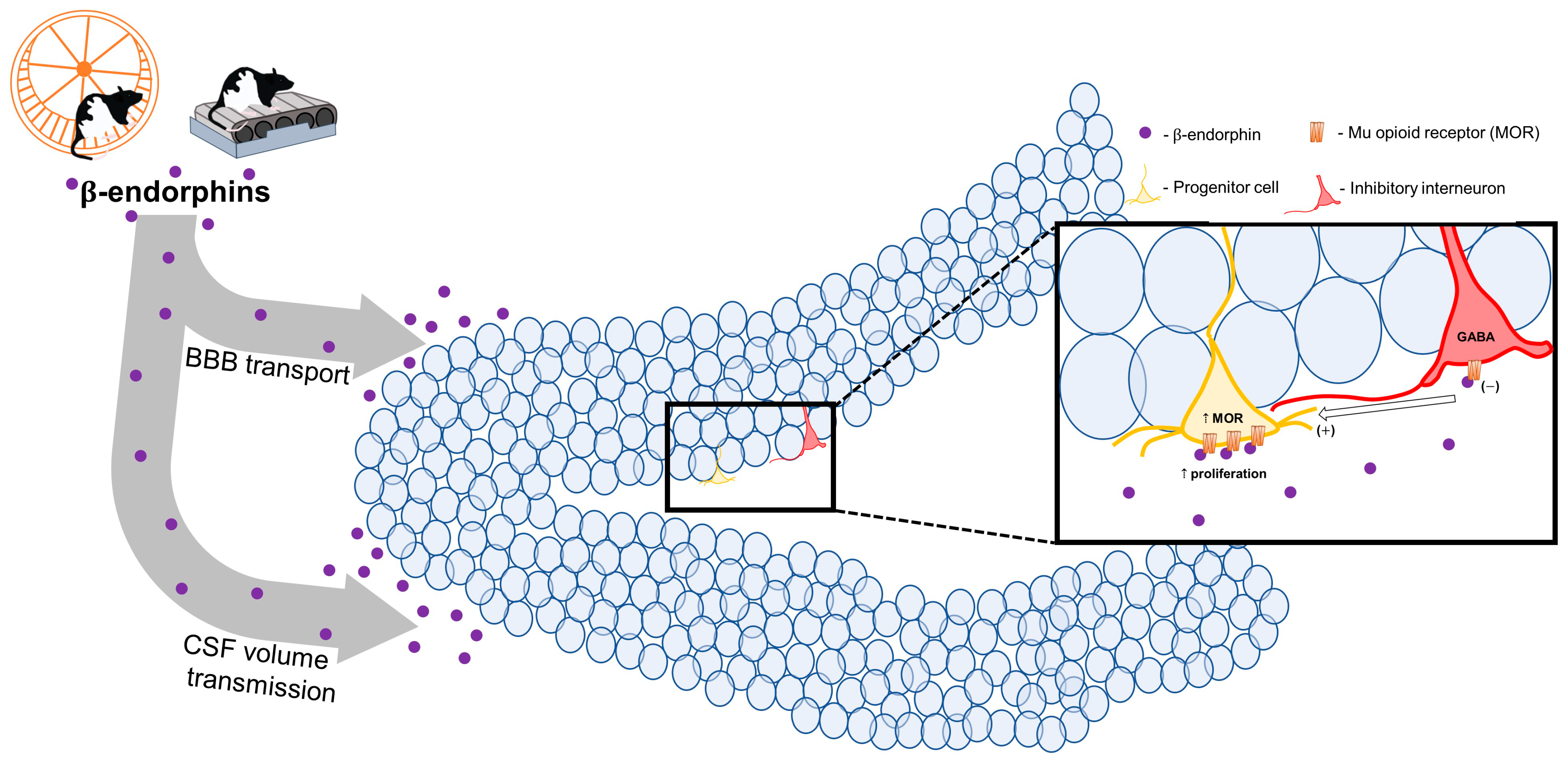
5. Potential Mechanisms Involved in Exercise Effects on Neurogenesis—Endorphins?
6. β-Endorphin: Just for the Periphery?
7. Direct Evidence for Endorphin Influence on Neurogenesis
8. Circumstantial Evidence for β-Endorphin Roles in Neurogenesis—A Need for More Behavioral Studies Utilizing Endogenous Opioids
9. Implications and Conclusions
Funding
Conflicts of Interest
References
- Jesus, I.; Vanhee, V.; Deramaudt, T.B.; Bonay, M. Promising effects of exercise on the cardiovascular, metabolic and immune system during COVID-19 period. J. Hum. Hypertens. 2021, 35, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Gorostegi-Anduaga, I.; Corres, P.; MartinezAguirre-Betolaza, A.; Pérez-Asenjo, J.; Aispuru, G.R.; Fryer, S.M.; Maldonado-Martín, S. Effects of different aerobic exercise programmes with nutritional intervention in sedentary adults with overweight/obesity and hypertension: EXERDIET-HTA study. Eur. J. Prev. Cardiol. 2018, 25, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Al Dahamsheh, Z.; Al Rashdan, K.; Al Hadid, A.; Jaradat, R.; Al Bakheet, M.; Bataineh, Z.S. The Impact of Aerobic Exercise on Female Bone Health Indicators. Med. Arch. 2019, 73, 35–38. [Google Scholar] [CrossRef] [PubMed]
- Howard, V.J.; McDonnell, M.N. Physical Activity in Primary Stroke Prevention: Just Do It! Stroke 2015, 46, 1735–1739. [Google Scholar] [CrossRef] [PubMed]
- Ainslie, P.N.; Cotter, J.D.; George, K.P.; Lucas, S.; Murrell, C.; Shave, R.; Thomas, K.N.; Williams, M.J.A.; Atkinson, G. Elevation in cerebral blood flow velocity with aerobic fitness throughout healthy human ageing. J. Physiol. 2008, 586, 4005–4010. [Google Scholar] [CrossRef]
- Colcombe, S.J.; Erickson, K.I.; Scalf, P.E.; Kim, J.S.; Prakash, R.; McAuley, E.; Elavsky, S.; Marquez, D.X.; Hu, L.; Kramer, A.F. Aerobic exercise training increases brain volume in aging humans. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2006, 61, 1166–1170. [Google Scholar] [CrossRef]
- Ahlskog, J.E. Does vigorous exercise have a neuroprotective effect in Parkinson disease? Neurology 2011, 77, 288–294. [Google Scholar] [CrossRef]
- Paillard, T.; Rolland, Y.; de Souto Barreto, P. Protective Effects of Physical Exercise in Alzheimer’s Disease. J. Clin. Neurol. 2015, 11, 212–219. [Google Scholar] [CrossRef]
- Briken, S.; Gold, S.M.; Patra, S.; Vettorazzi, E.; Harbs, D.; Tallner, A.; Ketels, G.; Schulz, K.H.; Heesen, C. Effects of exercise on fitness and cognition in progressive MS: A randomized, controlled pilot trial. Mult. Scler. J. 2014, 20, 382–390. [Google Scholar] [CrossRef]
- Hillman, C.H.; Erickson, K.I.; Kramer, A.F. Be smart, exercise your heart: Exercise effects on brain and cognition. Nat. Rev. Neurosci. 2008, 9, 58–65. [Google Scholar] [CrossRef]
- Dilorenzo, T.M.; Bargman, E.P.; Stucky-Ropp, R.; Brassington, G.S.; Frensch, P.A.; LaFontaine, T. Long-term effects of aerobic exercise on psychological outcomes. Prev. Med. 1999, 28, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Kandola, A.; Hendrikse, J.; Lucassen, P.J.; Yücel, M. Aerobic Exercise as a Tool to Improve Hippocampal Plasticity and Function in Humans: Practical Implications for Mental Health Treatment. Front. Hum. Neurosci. 2016, 10, 373. [Google Scholar] [CrossRef] [PubMed]
- Way, K.; Kannis-Dymand, L.; Lastella, M.; Lovell, G.P. Mental health practitioners’ reported barriers to prescription of exercise for mental health consumers. Ment. Health Phys. Act. 2018, 14, 52–60. [Google Scholar] [CrossRef]
- Bernstein, E.E.; McNally, R.J. Examining the Effects of Exercise on Pattern Separation and the Moderating Effects of Mood Symptoms. Behav. Ther. 2019, 50, 582–593. [Google Scholar] [CrossRef]
- Erickson, K.I.; Voss, M.W.; Prakash, R.S.; Basak, C.; Szabo, A.; Chaddock, L.; Kim, J.S.; Heo, S.; Alves, H.; White, S.M.; et al. Exercise training increases size of hippocampus and improves memory. Proc. Natl. Acad. Sci. USA 2011, 108, 3017–3022. [Google Scholar] [CrossRef]
- Petruzzello, S.J.; Landers, D.M.; Hatfield, B.D.; Kubitz, K.A.; Salazar, W. A Meta-Analysis on the Anxiety-Reducing Effects of Acute and Chronic Exercise: Outcomes and Mechanisms. Sports Med. 1991, 11, 143–182. [Google Scholar] [CrossRef]
- Craft, L.L.; Landers, D.M. The Effect of Exercise on Clinical Depression and Depression Resulting from Mental Illness: A Meta-Analysis. J. Sport Exerc. Psychol. 1998, 20, 339–357. [Google Scholar] [CrossRef]
- Zschucke, E.; Renneberg, B.; Dimeo, F.; Wüstenberg, T.; Ströhle, A. The stress-buffering effect of acute exercise: Evidence for HPA axis negative feedback. Psychoneuroendocrinology 2015, 51, 414–425. [Google Scholar] [CrossRef]
- Masley, S.; Roetzheim, R.; Gualtieri, T. Aerobic exercise enhances cognitive flexibility. J. Clin. Psychol. Med. Settings 2009, 16, 186–193. [Google Scholar] [CrossRef]
- Cameron, H.A.; Glover, L.R. Adult Neurogenesis: Beyond Learning and Memory. Annu. Rev. Psychol. 2015, 66, 53–81. [Google Scholar] [CrossRef]
- Gould, E. How widespread is adult neurogenesis in mammals? Nat. Rev. Neurosci. 2007, 8, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Jimenez, E.P.; Terreros-Roncal, J.; Flor-Garcia, M.; Rabano, A.; Llorens-Martin, M. Evidences for Adult Hippocampal Neurogenesis in Humans. J. Neurosci. 2021, 41, 2541–2553. [Google Scholar] [CrossRef] [PubMed]
- Garthe, A.; Kempermann, G. An old test for new neurons: Refining the morris water maze to study the functional relevance of adult hippocampal neurogenesis. Front. Neurosci. 2013, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Drew, M.R.; Denny, C.A.; Hen, R. Arrest of adult hippocampal neurogenesis in mice impairs single- but not multiple-trial contextual fear conditioning. Behav. Neurosci. 2010, 124, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Clelland, C.D.; Choi, M.; Romberg, C.; Clemenson, G.D.; Fragniere, A.; Tyers, P.; Jessberger, S.; Saksida, L.M.; Barker, R.A.; Gage, F.H.; et al. A functional role for adult hippocampal neurogenesis in spatial pattern separation. Science 2009, 325, 210–213. [Google Scholar] [CrossRef]
- Weeden, C.S.S.; Mercurio, J.C.; Cameron, H.A. A role for hippocampal adult neurogenesis in shifting attention toward novel stimuli. Behav. Brain Res. 2019, 376. [Google Scholar] [CrossRef] [PubMed]
- Snyder, J.S.; Soumier, A.; Brewer, M.; Pickel, J.; Cameron, H.A. Adult hippocampal neurogenesis buffers stress responses and depressive behaviour. Nature 2011, 476, 458–461. [Google Scholar] [CrossRef]
- Schoenfeld, T.J.; Cameron, H.A. Adult neurogenesis and mental illness. Neuropsychopharmacology 2015, 40. [Google Scholar] [CrossRef] [PubMed]
- Van Praag, H.; Kempermann, G.; Gage, F.H. Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus. Nat. Neurosci. 1999, 2, 266–270. [Google Scholar] [CrossRef]
- Brown, J.; Cooper-Kuhn, C.M.; Kempermann, G.; Van Praag, H.; Winkler, J.; Gage, F.H.; Kuhn, H.G. Enriched environment and physical activity stimulate hippocampal but not olfactory bulb neurogenesis. Eur. J. Neurosci. 2003, 17, 2042–2046. [Google Scholar] [CrossRef]
- Schoenfeld, T.J.; Rada, P.; Pieruzzini, P.R.; Hsueh, B.; Gould, E. Physical exercise prevents stress-induced activation of granule neurons and enhances local inhibitory mechanisms in the dentate gyrus. J. Neurosci. 2013, 33. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.M.; Galea, L.A.M.; Mistlberger, R.E.; Kempermann, G. Adult Hippocampal Neurogenesis and Voluntary Running Activity: Circadian and Dose-Dependent Effects. J. Neurosci. Res. 2004, 76, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, J.S.; Jeffrey, S.; Girard, I.; Mitchell, G.S.; Van Praag, H.; Garland, T.; Gage, F.H. Exercise increases hippocampal neurogenesis to high levels but does not improve spatial learning in mice bred for increased voluntary wheel running. Behav. Neurosci. 2003, 117, 1006–1016. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.C.; Inoue, K.; Okamoto, M.; Liu, Y.F.; Matsui, T.; Yook, J.S.; Soya, H. Voluntary resistance running induces increased hippocampal neurogenesis in rats comparable to load-free running. Neurosci. Lett. 2013, 537, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Nguemeni, C.; McDonald, M.W.; Jeffers, M.S.; Livingston-Thomas, J.; Lagace, D.; Corbett, D. Short- and Long-term Exposure to Low and High Dose Running Produce Differential Effects on Hippocampal Neurogenesis. Neuroscience 2018, 369, 202–211. [Google Scholar] [CrossRef]
- Huang, Y.Q.; Wu, C.; He, X.F.; Wu, D.; He, X.; Liang, F.Y.; Dai, G.Y.; Pei, Z.; Xu, G.Q.; Lan, Y. Effects of voluntary wheel-running types on hippocampal neurogenesis and spatial cognition in middle-aged mice. Front. Cell. Neurosci. 2018, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lan, Y.; Huang, Z.; Jiang, Y.; Zhou, X.; Zhang, J.; Zhang, D.; Wang, B.; Hou, G. Strength exercise weakens aerobic exerciseinduced cognitive improvements in rats. PLoS ONE 2018, 13, e0205562. [Google Scholar] [CrossRef] [PubMed]
- Codina-Martínez, H.; Fernández-García, B.; Díez-Planelles, C.; Fernández, Á.F.; Higarza, S.G.; Fernández-Sanjurjo, M.; Díez-Robles, S.; Iglesias-Gutiérrez, E.; Tomás-Zapico, C. Autophagy is required for performance adaptive response to resistance training and exercise-induced adult neurogenesis. Scand. J. Med. Sci. Sports 2020, 30, 238–253. [Google Scholar] [CrossRef]
- Novaes Gomes, F.G.; Fernandes, J.; Vannucci Campos, D.; Cassilhas, R.C.; Viana, G.M.; D’Almeida, V.; de Moraes Rêgo, M.K.; Buainain, P.I.; Cavalheiro, E.A.; Arida, R.M. The beneficial effects of strength exercise on hippocampal cell proliferation and apoptotic signaling is impaired by anabolic androgenic steroids. Psychoneuroendocrinology 2014, 50, 106–117. [Google Scholar] [CrossRef]
- Nokia, M.S.; Lensu, S.; Ahtiainen, J.P.; Johansson, P.P.; Koch, L.G.; Britton, S.L.; Kainulainen, H. Physical exercise increases adult hippocampal neurogenesis in male rats provided it is aerobic and sustained. J. Physiol. 2016, 594, 1855–1873. [Google Scholar] [CrossRef]
- Gremmelspacher, T.; Gerlach, J.; Hubbe, A.; Haas, C.A.; Häussler, U. Neurogenic processes are induced by very short periods of voluntary wheel-running in male mice. Front. Neurosci. 2017, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Snyder, J.S.; Glover, L.R.; Sanzone, K.M.; Kamhi, J.F.; Cameron, H.A. The effects of exercise and stress on the survival and maturation of adult-generated granule cells. Hippocampus 2009, 19, 898–906. [Google Scholar] [CrossRef] [PubMed]
- Sah, N.; Peterson, B.D.; Lubejko, S.T.; Vivar, C.; Van Praag, H. Running reorganizes the circuitry of one-week-old adult-born hippocampal neurons. Sci. Rep. 2017, 7, 10903. [Google Scholar] [CrossRef]
- Stranahan, A.M.; Lee, K.; Mattson, M.P. Central mechanisms of HPA axis regulation by voluntary exercise. Neuromolecular Med. 2008, 10, 118–127. [Google Scholar] [CrossRef]
- Schoenfeld, T.J.; Gould, E. Stress, stress hormones, and adult neurogenesis. Exp. Neurol. 2012, 233. [Google Scholar] [CrossRef]
- Stranahan, A.M.; Khalil, D.; Gould, E. Social isolation delays the positive effects of running on adult neurogenesis. Nat. Neurosci. 2006, 9, 526–533. [Google Scholar] [CrossRef]
- Leasure, J.L.; Decker, L. Social isolation prevents exercise-induced proliferation of hippocampal progenitor cells in female rats. Hippocampus 2009, 19, 907–912. [Google Scholar] [CrossRef]
- Hauser, T.; Klaus, F.; Lipp, H.P.; Amrein, I. No effect of running and laboratory housing on adult hippocampal neurogenesis in wild caught long-tailed wood mouse. BMC Neurosci. 2009, 10, 43. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liang, A.; Guan, F.; Fan, R.; Chi, L.; Yang, B. Regular treadmill running improves spatial learning and memory performance in young mice through increased hippocampal neurogenesis and decreased stress. Brain Res. 2013, 1531, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Okamoto, M.; Shibato, J.; Lee, M.C.; Matsui, T.; Rakwal, R.; Soya, H. Long-term mild, rather than intense, exercise enhances adult hippocampal neurogenesis and greatly changes the transcriptomic profile of the hippocampus. PLoS ONE 2015, 10, e0128720. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, M.; Yamamura, Y.; Liu, Y.-F.; Min-Chul, L.; Matsui, T.; Shima, T.; Soya, M.; Takahashi, K.; Soya, S.; McEwen, B.S.; et al. Hormetic effects by exercise on hippocampal neurogenesis with glucocorticoid signaling. Brain Plast. 2016, 1, 149–158. [Google Scholar] [CrossRef]
- Clark, P.J.; Brzezinska, W.J.; Thomas, M.W.; Ryzhenko, N.A.; Toshkov, S.A.; Rhodes, J.S. Intact neurogenesis is required for benefits of exercise on spatial memory but not motor performance or contextual fear conditioning in C57BL/6J mice. Neuroscience 2008, 155, 1048–1058. [Google Scholar] [CrossRef] [PubMed]
- Winocur, G.; Becker, S.; Luu, P.; Rosenzweig, S.; Wojtowicz, J.M. Adult hippocampal neurogenesis and memory interference. Behav. Brain Res. 2012, 227, 464–469. [Google Scholar] [CrossRef] [PubMed]
- Wojtowicz, J.M.; Askew, M.L.; Winocur, G. The effects of running and of inhibiting adult neurogenesis on learning and memory in rats. Eur. J. Neurosci. 2008, 27, 1494–1502. [Google Scholar] [CrossRef] [PubMed]
- Wong-Goodrich, S.J.E.; Pfau, M.L.; Flores, C.T.; Fraser, J.A.; Williams, C.L.; Jones, L.W. Voluntary running prevents progressive memory decline and increases adult hippocampal neurogenesis and growth factor expression after whole-brain irradiation. Cancer Res. 2010, 70, 9329–9338. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, G.F.; Majdak, P.; Miller, D.S.; Bucko, P.J.; Merritt, J.R.; Krebs, C.P.; Rhodes, J.S. Evaluation of a C57BL/6J x 129S1/SvImJ Hybrid Nestin-Thymidine Kinase Transgenic Mouse Model for Studying the Functional Significance of Exercise-Induced Adult Hippocampal Neurogenesis. Brain Plast. 2016, 1, 83–95. [Google Scholar] [CrossRef]
- Snyder, J.S.; Cahill, S.P.; Frankland, P.W. Running Promotes Spatial Bias Independently of Adult Neurogenesis. Hippocampus 2017, 27, 871–882. [Google Scholar] [CrossRef] [PubMed]
- Yau, S.Y.; Lau, B.W.M.; Bin Tong, J.; Wong, R.; Ching, Y.P.; Qiu, G.; Tang, S.W.; Lee, T.M.C.; So, K.F. Hippocampal neurogenesis and dendritic plasticity support running-improved spatial learning and depression-like behaviour in stressed rats. PLoS ONE 2011, 6, e24263. [Google Scholar] [CrossRef]
- Schoenfeld, T.J.; McCausland, H.C.; Sonti, A.N.; Cameron, H.A. Anxiolytic Actions of Exercise in Absence of New Neurons. Hippocampus 2016, 26. [Google Scholar] [CrossRef]
- Zheng, J.; Jiang, Y.Y.; Xu, L.C.; Ma, L.Y.; Liu, F.Y.; Cui, S.; Cai, J.; Liao, F.F.; Wan, Y.; Yi, M. Adult hippocampal neurogenesis along the dorsoventral axis contributes differentially to environmental enrichment combined with voluntary exercise in alleviating chronic inflammatory pain in mice. J. Neurosci. 2017, 37, 4145–4157. [Google Scholar] [CrossRef]
- Kim, J.J.; Foy, M.R.; Thompson, R.F. Behavioral stress modifies hippocampal plasticity through N-methyl-D-asparate receptor activation. Proc. Natl. Acad. Sci. USA 1996, 93, 4750–4753. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.J.; Kitamura, T.; Saitoh, Y.; Ohkawa, N.; Kondo, T.; Inokuchi, K. Adult neurogenesis conserves hippocampal memory capacity. J. Neurosci. 2018, 38, 6854–6863. [Google Scholar] [CrossRef]
- Patten, A.R.; Sickmann, H.; Hryciw, B.N.; Kucharsky, T.; Parton, R.; Kernick, A.; Christie, B.R. Long-term exercise is needed to enhance synaptic plasticity in the hippocampus. Learn. Mem. 2013, 20, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Vaynman, S.; Ying, Z.; Gomez-Pinilla, F. Hippocampal BDNF mediates the efficacy of exercise on synaptic plasticity and cognition. Eur. J. Neurosci. 2004, 20, 2580–2590. [Google Scholar] [CrossRef] [PubMed]
- Eadie, B.D.; Redila, V.A.; Christie, B.R. Voluntary exercise alters the cytoarchitecture of the adult dentate gyrus by increasing cellular proliferation, dendritic complexity, and spine density. J. Comp. Neurol. 2005, 486, 39–47. [Google Scholar] [CrossRef]
- Biedermann, S.; Fuss, J.; Zheng, L.; Sartorius, A.; Falfán-Melgoza, C.; Demirakca, T.; Gass, P.; Ende, G.; Weber-Fahr, W. In vivo voxel based morphometry: Detection of increased hippocampal volume and decreased glutamate levels in exercising mice. NeuroImage 2012, 61, 1206–1212. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Holsinger, R.M.D. Exercise-induced brain-derived neurotrophic factor expression: Therapeutic implications for Alzheimer’s dementia. Ageing Res. Rev. 2018, 48, 109–121. [Google Scholar] [CrossRef]
- Jin, K.; Zhu, Y.; Sun, Y.; Mao, X.O.; Xie, L.; Greenberg, D.A. Vascular endothelial growth factor (VEGF) stimulates neurogenesis in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2002, 99, 11946–11950. [Google Scholar] [CrossRef]
- Rich, B.; Scadeng, M.; Yamaguchi, M.; Wagner, P.D.; Breen, E.C. Skeletal myofiber vascular endothelial growth factor is required for the exercise training-induced increase in dentate gyrus neuronal precursor cells. J. Physiol. 2017, 595, 5931–5943. [Google Scholar] [CrossRef]
- Fabel, K.; Fabel, K.; Tam, B.; Kaufer, D.; Baiker, A.; Simmons, N.; Kuo, C.J.; Palmer, T.D. VEGF is necessary for exercise-induced adult hippocampal neurogenesis. Eur. J. Neurosci. 2003, 18, 2803–2812. [Google Scholar] [CrossRef]
- Mukuda, T.; Sugiyama, H. An angiotensin II receptor antagonist suppresses running-enhanced hippocampal neurogenesis in rat. Neurosci. Res. 2007, 58, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Estévez, V.; Defterali, Ç.; Vicario-Abejón, C. IGF-I: A key growth factor that regulates neurogenesis and synaptogenesis from embryonic to adult stages of the brain. Front. Neurosci. 2016, 10, 52. [Google Scholar] [CrossRef] [PubMed]
- Trejo, J.L.; LLorens-Martín, M.V.; Torres-Alemán, I. The effects of exercise on spatial learning and anxiety-like behavior are mediated by an IGF-I-dependent mechanism related to hippocampal neurogenesis. Mol. Cell. Neurosci. 2008, 37, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Hofer, M.; Hofer, M.; Pagliusi, S.R.; Pagliusi, S.R.; Hohn, A.; Hohn, A.; Leibrock, J.; Leibrock, J.; Barde, Y.; Barde, Y. Regional distribution. EMBO J. 1990, 9, 2459–2464. [Google Scholar] [CrossRef] [PubMed]
- Bath, K.G.; Akins, M.R.; Lee, F.S. BDNF control of adult SVZ neurogenesis. Dev. Psychobiol. 2012, 54, 578–589. [Google Scholar] [CrossRef] [PubMed]
- Oliff, H.S.; Berchtold, N.C.; Isackson, P.; Cotman, C.W. Exercise-induced regulation of brain-derived neurotrophic factor (BDNF) transcripts in the rat hippocampus. Mol. Brain Res. 1998, 61, 147–153. [Google Scholar] [CrossRef]
- Adlard, P.A.; Perreau, V.M.; Cotman, C.W. The exercise-induced expression of BDNF within the hippocampus varies across life-span. Neurobiol. Aging 2005, 26, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Duan, W.; Mattson, M.P. Evidence that brain-derived neurotrophic factor is required for basal neurogenesis and mediates, in part, the enhancement of neurogenesis by dietary restriction in the hippocampus of adult mice. J. Neurochem. 2002, 82, 1367–1375. [Google Scholar] [CrossRef]
- Quesseveur, G.; David, D.J.; Gaillard, M.C.; Pla, P.; Wu, M.V.; Nguyen, H.T.; Nicolas, V.; Auregan, G.; David, I.; Dranovsky, A.; et al. BDNF overexpression in mouse hippocampal astrocytes promotes local neurogenesis and elicits anxiolytic-like activities. Transl. Psychiatry 2013, 3. [Google Scholar] [CrossRef]
- Rossi, C.; Angelucci, A.; Costantin, L.; Braschi, C.; Mazzantini, M.; Babbini, F.; Fabbri, M.E.; Tessarollo, L.; Maffei, L.; Berardi, N.; et al. Brain-derived neurotrophic factor (BDNF) is required for the enhancement of hippocampal neurogenesis following environmental enrichment. Eur. J. Neurosci. 2006, 24, 1850–1856. [Google Scholar] [CrossRef]
- Li, Y.; Luikart, B.W.; Birnbaum, S.; Chen, J.; Kwon, C.H.; Kernie, S.G.; Bassel-Duby, R.; Parada, L.F. TrkB Regulates Hippocampal Neurogenesis and Governs Sensitivity to Antidepressive Treatment. Neuron 2008, 59, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Wildmann, J.; Kruger, A.; Schmole, M.; Niemann, J.; Matthaei, H. Increase of circulating beta-endorphin-like immunoreactivity correlates with the change in feeling of pleasantness after running. Life Sci. 1986, 38, 997–1003. [Google Scholar] [CrossRef]
- Fuss, J.; Steinle, J.; Bindila, L.; Auer, M.K.; Kirchherr, H.; Lutz, B.; Gass, P. A runner’s high depends on cannabinoid receptors in mice. Proc. Natl. Acad. Sci. USA 2015, 112, 13105–13108. [Google Scholar] [CrossRef]
- Guillemin, R.; Vargo, T.; Rossier, J.; Minick, S.; Ling, N.; Rivier, C.; Vale, W.; Bloom, F.; Smith, P. β-Endrophin and Adrenocorticotropin Are Secreted Concomitantly by the Pituitary Gland. Science 1977, 197, 1367–1369. [Google Scholar] [CrossRef]
- Parikh, D.; Hamid, A.; Friedman, T.C.; Nguyen, K.; Tseng, A.; Marquez, P.; Lutfy, K. Stress-induced analgesia and endogenous opioid peptides: The importance of stress duration. Eur. J. Pharmacol. 2011, 650, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Apkarian, A.V.; Mutso, A.A.; Centeno, M.V.; Kan, L.; Wu, M.; Levinstein, M.; Banisadr, G.; Gobeske, K.T.; Miller, R.J.; Radulovic, J.; et al. Role of adult hippocampal neurogenesis in persistent pain. Pain 2016, 157, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Sauriyal, D.S.; Jaggi, A.S.; Singh, N. Extending pharmacological spectrum of opioids beyond analgesia: Multifunctional aspects in different pathophysiological states. Neuropeptides 2011, 45, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Bolijn, S.; Lucassen, P.J. How the Body Talks to the Brain; Peripheral Mediators of Physical Activity-Induced Proliferation in the Adult Hippocampus. Brain Plast. 2016, 1, 5–27. [Google Scholar] [CrossRef]
- Millan, M.J.; Przewlock, R.; Jerlicz, M.; Gramsch, C.; Höllt, V.; Herz, A. Stress-induced release of brain and pituitary β-endorphin: Major role of endorphins in generation of hyperthermia, not analgesia. Brain Res. 1981, 208, 325–338. [Google Scholar] [CrossRef]
- Xue, L.; Sun, J.; Zhu, J.; Ding, Y.; Chen, S.; Ding, M.; Pei, H. The patterns of exercise-induced β-endorphin expression in the central nervous system of rats. Neuropeptides 2020, 82. [Google Scholar] [CrossRef]
- Barfield, E.T.; Alexandra Moser, V.; Hand, A.; Grisel, J.E. ß-Endorphin Modulates the Effect of Stress on Novelty-Suppressed Feeding. Front. Behav. Neurosci. 2013, 7, 19. [Google Scholar] [CrossRef] [PubMed]
- Drake, C.T.; Chavkin, C.; Milner, T.A. Opioid systems in the dentate gyrus. Prog. Brain Res. 2007, 163. [Google Scholar] [CrossRef]
- Padilla, S.L.; Reef, D.; Zeltser, L.M. Defining POMC neurons using transgenic reagents: Impact of transient Pomc expression in diverse immature neuronal populations. Endocrinology 2012, 153, 1219–1231. [Google Scholar] [CrossRef]
- Bloom, F.; Battenberg, E.; Rossier, J.; Ling, N.; Guillemin, R. Neurons containing β-endorphin in rat brain exist separately from those containing enkephalin: Immunocytochemical studies. Proc. Natl. Acad. Sci. USA 1978, 75, 1591–1595. [Google Scholar] [CrossRef]
- Wang, D.; He, X.; Zhao, Z.; Feng, Q.; Lin, R.; Sun, Y.; Ding, T.; Xu, F.; Luo, M.; Zhan, C. Whole-brain mapping of the direct inputs and axonal projections of POMC and AgRP neurons. Front. Neuroanat. 2015, 9, 40. [Google Scholar] [CrossRef] [PubMed]
- Porro, C.A.; Cavazzuti, M.; Baraldi, P.; Giuliani, D.; Panerai, A.E.; Corazza, R. CNS pattern of metabolic activity during tonic pain: Evidence for modulation by β-endorphin. Eur. J. Neurosci. 1999, 11, 874–888. [Google Scholar] [CrossRef]
- Wang, J.; Li, X.; Wu, H.; Ke, J.; Zhang, Z.; Wang, Y. Effects of L-655,708 on expression changes of GABA, glutamate, and beta-endorphin induced by propofol anesthesia in rats. Eur. J. Inflamm. 2018, 16. [Google Scholar] [CrossRef]
- Houghten, R.A.; Swann, R.W.; Li, C.H. beta-Endorphin: Stability, clearance behavior, and entry into the central nervous system after intravenous injection of the tritiated peptide in rats and rabbits. Proc. Natl. Acad. Sci. USA 1980, 77, 4588–4591. [Google Scholar] [CrossRef]
- Gao, B.; Hagenbuch, B.; Kullak-Ublick, G.A.; Benke, D.; Aguzzi, A.; Meier-Abt, P.J. Organic anion-transporting polypeptides mediate transport of opioid peptides across blood-brain barrier. J. Pharmacol. Exp. Ther. 2000, 294, 73–79. [Google Scholar]
- Kumagai, A.K.; Eisenberg, J.B.; Pardridge, W.M. Absorptive-mediated endocytosis of cationized albumin and a β-endorphin-cationized albumin chimeric peptide by isolated brain capillaries. Model system of blood-brain barrier transport. J. Biol. Chem. 1987, 262, 15214–15219. [Google Scholar] [CrossRef]
- Rodríguez, E.M.; Blázquez, J.L.; Guerra, M. The design of barriers in the hypothalamus allows the median eminence and the arcuate nucleus to enjoy private milieus: The former opens to the portal blood and the latter to the cerebrospinal fluid. Peptides 2010, 31, 757–776. [Google Scholar] [CrossRef]
- Hoffmann, P.; Terenius, L.; Thoren, P. Cerebrospinal fluid immunoreactive fl-endorphin concentration is increased by voluntary exercise in the spontaneously hypertensive rat. Regul. Pept. 1990, 28, 233–239. [Google Scholar] [CrossRef]
- Veening, J.G.; Gerrits, P.O.; Barendregt, H.P. Volume transmission of beta-endorphin via the cerebrospinal fluid; a review. Fluids Barriers CNS 2012, 9, 16. [Google Scholar] [CrossRef] [PubMed]
- Leak, R.K.; Moore, R.Y. Innervation of ventricular and periventricular brain compartments. Brain Res. 2012, 1463, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Ableitner, A.; Schulz, R. Neuroanatomical sites mediating the central actions of beta-endorphin as mapped by changes in glucose utilization: Involvement of mu opioid receptors. J. Pharmacol. Exp. Ther. 1992, 262, 415–423. [Google Scholar]
- Koehl, M.; Meerlo, P.; Gonzales, D.; Rontal, A.; Turek, F.W.; Abrous, D.N. Exercise-induced promotion of hippocampal cell proliferation requires β-endorphin. FASEB J. 2008, 22, 2253–2262. [Google Scholar] [CrossRef] [PubMed]
- Persson, A.I.; Naylor, A.S.; Jonsdottir, I.H.; Nyberg, F.; Eriksson, P.S.; Thorlin, T. Differential regulation of hippocampal progenitor proliferation by opioid receptor antagonists in running and non-running spontaneously hypertensive rats. Eur. J. Neurosci. 2004, 19, 1847–1855. [Google Scholar] [CrossRef]
- Nieto, S.J.; Quave, C.B.; Kosten, T.A. Naltrexone alters alcohol self-administration behaviors and hypothalamic-pituitary-adrenal axis activity in a sex-dependent manner in rats. Pharmacol. Biochem. Behav. 2018, 167, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Persson, A.I.; Thorlin, T.; Bull, C.; Zarnegar, P.; Ekman, R.; Terenius, L.; Eriksson, P.S. Mu- and delta-opioid receptor antagonists decrease proliferation and increase neurogenesis in cultures of rat adult hippocampal progenitors. Eur. J. Neurosci. 2003, 17, 1159–1172. [Google Scholar] [CrossRef]
- Persson, A.I.; Thorlin, T.; Bull, C.; Eriksson, P.S. Opioid-induced proliferation through the MAPK pathway in cultures of adult hippocampal progenitors. Mol. Cell. Neurosci. 2003, 23, 360–372. [Google Scholar] [CrossRef]
- Zhang, H.; Torregrossa, M.M.; Jutkiewicz, E.M.; Shi, Y.G.; Rice, K.C.; Woods, J.H.; Watson, S.J.; Holden Ko, M.C. Endogenous opioids upregulate brain-derived neurotrophic factor mRNA through δ- and μ-opioid receptors independent of antidepressant-like effects. Eur. J. Neurosci. 2006, 23, 984–994. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, M.S.R.; da Silva Fernandes, M.J.; Scorza, F.A.; Persike, D.S.; Scorza, C.A.; da Ponte, J.B.; de Albuquerque, M.; Cavalheiro, E.A.; Arida, R.M. Acute and chronic exercise modulates the expression of MOR opioid receptors in the hippocampal formation of rats. Brain Res. Bull. 2010, 83, 278–283. [Google Scholar] [CrossRef]
- Neumaier, J.F.; Mailheau, S.; Chavkin, C. Opioid receptor-mediated responses in the dentate gyrus and CA1 region of the rat hippocampus. J. Pharmacol. Exp. Ther. 1988, 244, 564–570. [Google Scholar] [PubMed]
- Drake, C.T.; Milner, T.A. Mu opioid receptors are in somatodendritic and axonal compartments of GABAergic neurons in rat hippocampal formation. Brain Res. 1999, 849, 203–215. [Google Scholar] [CrossRef]
- Svoboda, K.R.; Adams, C.E.; Lupica, C.R. Opioid receptor subtype expression defines morphologically distinct classes of hippocampal interneurons. J. Neurosci. 1999, 19, 85–95. [Google Scholar] [CrossRef]
- Tozuka, Y.; Fukuda, S.; Namba, T.; Seki, T.; Hisatsune, T. GABAergic excitation promotes neuronal differentiation in adult hippocampal progenitor cells. Neuron 2005, 47, 803–815. [Google Scholar] [CrossRef] [PubMed]
- Schoenfeld, T.J.; Gould, E. Differential Effects of Stress and Glucocorticoids on Adult Neurogenesis; Springer: Berlin/Heidelberg, Germany, 2013; Volume 15. [Google Scholar]
- Sahay, A.; Scobie, K.N.; Hill, A.S.; O’carroll, C.M.; Kheirbek, M.A.; Burghardt, N.S.; Fenton, A.A.; Dranovsky, A.; Hen, R. Increasing adult hippocampal neurogenesis is sufficient to improve pattern separation HHS Public Access. Nature 2011, 472, 466–470. [Google Scholar] [CrossRef]
- Hill, A.S.; Sahay, A.; Hen, R. Increasing Adult Hippocampal Neurogenesis is Sufficient to Reduce Anxiety and Depression-Like Behaviors. Neuropsychopharmacology 2015, 40, 2368–2378. [Google Scholar] [CrossRef]
- Culig, L.; Surget, A.; Bourdey, M.; Khemissi, W.; Le Guisquet, A.M.; Vogel, E.; Sahay, A.; Hen, R.; Belzung, C. Increasing adult hippocampal neurogenesis in mice after exposure to unpredictable chronic mild stress may counteract some of the effects of stress. Neuropharmacology 2017, 126, 179–189. [Google Scholar] [CrossRef]
- Izquierdo, I.; Souza, D.O.; Carrasco, M.A.; Dias, R.D.; Perry, M.L.; Eisinger, S.; Elisabetsky, E.; Vendite, D.A. Beta-endorphin causes retrograde amnesia and is released from the rat brain by various forms of training and stimulation. Psychopharmacology 1980, 70, 173–177. [Google Scholar] [CrossRef]
- Heybach, J.P.; Vernikos, J. Naloxone inhibits and morphine potentiates the adrenal steroidogenic response to ACTH. Eur. J. Pharmacol. 1981, 75, 1–6. [Google Scholar] [CrossRef]
- Miller, B.R.; Hen, R. The current state of the neurogenic theory of depression and anxiety. Curr. Opin. Neurobiol. 2015, 30, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Eisch, A.J.; Barrot, M.; Schad, C.A.; Self, D.W.; Nestler, E.J. Opiates inhibit neurogenesis in the adult rat hippocampus. Proc. Natl. Acad. Sci. USA 2000, 97, 7579–7584. [Google Scholar] [CrossRef] [PubMed]
- Molina, V.A.; Heyser, C.J.; Spear, L.P. Chronic variable stress or chronic morphine facilitates immobility in a forced swim test: Reversal by naloxone. Psychopharmacology 1994, 114, 433–440. [Google Scholar] [CrossRef] [PubMed]
- degli Uberti, E.C.; Petraglia, F.; Bondanelli, M.; Guo, A.L.; Valentini, A.; Salvadori, S.; Criscuolo, M.; Nappi, R.E.; Genazzani, A.R. Involvement of μ-opioid receptors in the modulation of pituitary-adrenal axis in normal and stressed rats. J. Endocrinol. Investig. 1995, 18, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ra, S.M.; Kim, H.; Jang, M.H.; Shin, M.C.; Lee, T.H.; Lim, B.V.; Kim, C.J.; Kim, E.H.; Kim, K.M.; Kim, S.S. Treadmill running and swimming increase cell proliferation in the hippocampal dentate gyrus of rats. Neurosci. Lett. 2002, 333, 123–126. [Google Scholar] [CrossRef]
- Hayward, M.D.; Pintar, J.E.; Low, M.J. Selective reward deficit in mice lacking β-endorphin and enkephalin. J. Neurosci. 2002, 22, 8251–8258. [Google Scholar] [CrossRef]
- Karlsson, R.M.; Wang, A.S.; Sonti, A.N.; Cameron, H.A. Adult neurogenesis affects motivation to obtain weak, but not strong, reward in operant tasks. Hippocampus 2018, 28, 512–522. [Google Scholar] [CrossRef]
- McGonigle, C.E.; Nentwig, T.B.; Wilson, D.E.; Rhinehart, E.M.; Grisel, J.E. Β-Endorphin Regulates Alcohol Consumption Induced By Exercise Restriction in Female Mice. Alcohol 2016, 53, 51–60. [Google Scholar] [CrossRef][Green Version]
- He, Y.; Lu, Y.; Shen, Y.; Wu, F.; Xu, X.; Kong, E.; Huang, Z.; Sun, Y.; Yu, W. Transgenic increase in the β-endorphin concentration in cerebrospinal fluid alleviates morphine-primed relapse behavior through the μ opioid receptor in rats. J. Med. Virol. 2019, 91, 1158–1167. [Google Scholar] [CrossRef]
- Goldfarb, A.H.; Jamurtas, A.Z.; Kamimori, G.H.; Hegde, S.; Otterstetter, R.; Brown, D.A. Gender effect on beta-endorphin response to exercise. Med. Sci. Sports Exerc. 1998, 30, 1672–1676. [Google Scholar] [CrossRef] [PubMed]
- Kanarek, R.B.; Gerstein, A.V.; Wildman, R.P.; Mathes, W.F.; D’Anci, K.E. Chronic running-wheel activity decreases sensitivity to morphine-induced analgesia in male and female rats. Pharmacol. Biochem. Behav. 1998, 61, 19–27. [Google Scholar] [CrossRef]
- Hare, B.D.; Beierle, J.A.; Toufexis, D.J.; Hammack, S.E.; Falls, W.A. Exercise-associated changes in the corticosterone response to acute restraint stress: Evidence for increased adrenal sensitivity and reduced corticosterone response duration. Neuropsychopharmacology 2014, 39, 1262–1269. [Google Scholar] [CrossRef] [PubMed]
- White-Welkley, J.E.; Bunnell, B.N.; Mougey, E.H.; Meyerhoff, J.L.; Dishman, R.K. Treadmill exercise training and estradiol differentially modulate hypothalamic-pituitary-adrenal cortical responses to acute running and immobilization. Physiol. Behav. 1995, 57, 533–540. [Google Scholar] [CrossRef]
- Rahimi, S.; Peeri, M.; Azarbayjani, M.A.; Anoosheh, L.; Ghasemzadeh, E.; Khalifeh, N.; Noroozi-Mahyari, S.; Deravi, S.; Saffari-Anaraki, S.; Hemat Zangeneh, F.; et al. Long-term exercise from adolescence to adulthood reduces anxiety- and depression-like behaviors following maternal immune activation in offspring. Physiol. Behav. 2020, 226. [Google Scholar] [CrossRef]
- Duman, C.H.; Schlesinger, L.; Russell, D.S.; Duman, R.S. Voluntary exercise produces antidepressant and anxiolytic behavioral effects in mice. Brain Res. 2008, 1199, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Barha, C.K.; Falck, R.S.; Davis, J.C.; Nagamatsu, L.S.; Liu-Ambrose, T. Sex differences in aerobic exercise efficacy to improve cognition: A systematic review and meta-analysis of studies in older rodents. Front. Neuroendocrinol. 2017, 46, 86–105. [Google Scholar] [CrossRef]
- Ma, X.; Hamadeh, M.J.; Christie, B.R.; Foster, J.A.; Tarnopolsky, M.A. Impact of treadmill running and sex on hippocampal neurogenesis in the mouse model of amyotrophic lateral sclerosis. PLoS ONE 2012, 7, e36048. [Google Scholar] [CrossRef]
- Cahill, S.P.; Cole, J.D.; Yu, R.Q.; Clemans-Gibbon, J.; Snyder, J.S. Differential Effects of Extended Exercise and Memantine Treatment on Adult Neurogenesis in Male and Female Rats. Neuroscience 2018, 390, 241–255. [Google Scholar] [CrossRef]
- Pluchino, N.; Drakopoulos, P.; Casarosa, E.; Freschi, L.; Petignat, P.; Yaron, M.; Genazzani, A.R. Effect of estetrol on beta-endorphin level in female rats. Steroids 2015, 95, 104–110. [Google Scholar] [CrossRef]
- Bernardi, F.; Pluchino, N.; Pieri, M.; Begliuomini, S.; Lenzi, E.; Puccetti, S.; Casarosa, E.; Luisi, S.; Genazzani, A.R. Progesterone and medroxyprogesterone acetate effects on central and peripheral allopregnanolone and beta-endorphin levels. Neuroendocrinology 2006, 83, 348–359. [Google Scholar] [CrossRef] [PubMed]
- Meyer, W.R.; Muoio, D.; Hackney, T.C. Effect of sex steroids on β-endorphin levels at rest and during submaximal treadmill exercise in anovulatory and ovulatory runners. Fertil. Steril. 1999, 71, 1085–1091. [Google Scholar] [CrossRef]
- Schneider, A.M.; Simson, P.E.; Spiller, K.; Adelstein, J.; Vacharat, A.; Short, K.R.; Kirby, L.G. Stress-dependent enhancement and impairment of retention by naloxone: Evidence for an endogenous opioid-based modulatory system protective of memory. Behav. Brain Res. 2009, 205, 290–293. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Holmes, M.M.; Galea, L.A.M. Defensive behavior and hippocampal cell proliferation: Differential modulation by naltrexone during stress. Behav. Neurosci. 2002, 116, 160–168. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Brito, R.G.; Rasmussen, L.A.; Sluka, K.A. Regular physical activity prevents development of chronic muscle pain through modulation of supraspinal opioid and serotonergic mechanisms. Pain Rep. 2017, 2, e618. [Google Scholar] [CrossRef] [PubMed]
- Glasper, E.R.; Schoenfeld, T.J.; Gould, E. Adult neurogenesis: Optimizing hippocampal function to suit the environment. Behav. Brain Res. 2012, 227. [Google Scholar] [CrossRef] [PubMed]
- Cameron, H.A.; Schoenfeld, T.J. Behavioral and structural adaptations to stress. Front. Neuroendocrinol. 2018, 49, 106–113. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schoenfeld, T.J.; Swanson, C. A Runner’s High for New Neurons? Potential Role for Endorphins in Exercise Effects on Adult Neurogenesis. Biomolecules 2021, 11, 1077. https://doi.org/10.3390/biom11081077
Schoenfeld TJ, Swanson C. A Runner’s High for New Neurons? Potential Role for Endorphins in Exercise Effects on Adult Neurogenesis. Biomolecules. 2021; 11(8):1077. https://doi.org/10.3390/biom11081077
Chicago/Turabian StyleSchoenfeld, Timothy J., and Chance Swanson. 2021. "A Runner’s High for New Neurons? Potential Role for Endorphins in Exercise Effects on Adult Neurogenesis" Biomolecules 11, no. 8: 1077. https://doi.org/10.3390/biom11081077
APA StyleSchoenfeld, T. J., & Swanson, C. (2021). A Runner’s High for New Neurons? Potential Role for Endorphins in Exercise Effects on Adult Neurogenesis. Biomolecules, 11(8), 1077. https://doi.org/10.3390/biom11081077