
Transient Receptor Potential (TRP) Channels in Haematological Malignancies: An Update

 ,
,  ,
,  ,
,  ,
, 
Abstract
1. Introduction
2. Myeloid Acute and Chronic Leukaemia
2.1. TRP Channels in AML
2.2. TRP Channels in CML
3. Acute Lymphoblastic (ALL) and Chronic Lymphocytic (CLL) Leukaemia
3.1. TRP Channels in ALL
3.2. TRP Channels in CLLs
4. Hodgkin and Non-Hodgkin Lymphomas
TRP Channels in HLs
5. TRPs as Promising Diagnostic, Prognostic, and Therapeutic Markers in the Clinical Management of Haematological Malignancies
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nilius, B. TRP channels in disease. Biochim. Biophys. Acta Mol. Basis Dis. 2007, 1772, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, I.S.; Delling, M.; Clapham, D.E. An introduction to TRP channels. Annu. Rev. Physiol. 2006, 68, 619–647. [Google Scholar] [CrossRef] [PubMed]
- Montell, C.; Rubin, G.M. Molecular characterization of the drosophila trp locus: A putative integral membrane protein required for phototransduction. Neuron 1989, 2, 1313–1323. [Google Scholar] [CrossRef]
- Zheng, J. Molecular Mechanism of TRP Channels. In Comprehensive Physiology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; Volume 3, pp. 221–242. [Google Scholar]
- Duan, J.; Li, Z.; Li, J.; Hulse, R.E.; Santa-Cruz, A.; Valinsky, W.C.; Abiria, S.A.; Krapivinsky, G.; Zhang, J.; Clapham, D.E. Structure of the mammalian TRPM7, a magnesium channel required during embryonic development. Proc. Natl. Acad. Sci. USA 2018, 115, E8201–E8210. [Google Scholar] [CrossRef]
- Ferrandiz-Huertas, C.; Mathivanan, S.; Wolf, C.J.; Devesa, I.; Ferrer-Montiel, A. Trafficking of thermo TRP channels. Membranes 2014, 4, 525–564. [Google Scholar] [CrossRef]
- Méndez-Reséndiz, K.A.; Enciso-Pablo, Ó.; González-Ramírez, R.; Juárez-Contreras, R.; Rosenbaum, T.; Morales-Lázaro, S.L. Steroids and TRP Channels: A close relationship. Int. J. Mol. Sci. 2020, 21, 3819. [Google Scholar] [CrossRef]
- Nagatomo, K.; Kubo, Y. Caffeine activates mouse TRPA1 channels but suppresses human TRPA1 channels. Proc. Natl. Acad. Sci. USA 2008, 105, 17373–17378. [Google Scholar] [CrossRef]
- Talavera, K.; Gees, M.; Karashima, Y.; Meseguer, V.M.; Vanoirbeek, J.A.J.; Damann, N.; Everaerts, W.; Benoit, M.; Janssens, A.; Vennekens, R.; et al. Nicotine activates the chemosensory cation channel TRPA1. Nat. Neurosci. 2009, 12, 1293–1299. [Google Scholar] [CrossRef]
- Bandell, M.; Story, G.M.; Hwang, S.W.; Viswanath, V.; Eid, S.R.; Petrus, M.J.; Earley, T.J.; Patapoutian, A. Noxious Cold Ion Channel TRPA1 Is Activated by Pungent Compounds and Bradykinin. Neuron 2004, 41, 849–857. [Google Scholar] [CrossRef]
- Rubaiy, H.N. Treasure troves of pharmacological tools to study transient receptor potential canonical 1/4/5 channels. Br. J. Pharmacol. 2019, 176, 832–846. [Google Scholar] [CrossRef]
- Venkatachalam, K.; Zheng, F.; Gill, D.L. Regulation of Canonical Transient Receptor Potential (TRPC) Channel Function by Diacylglycerol and Protein Kinase, C.J. Biol. Chem. 2003, 278, 29031–29040. [Google Scholar] [CrossRef] [PubMed]
- Leuner, K.; Kazanski, V.; Muller, M.; Essin, K.; Henke, B.; Gollasch, M.; Harteneck, C.; Müller, W.E. Hyperforin—A key constituent of St. John’s wort specifically activates TRPC6 channels. FASEB J. 2007, 21, 4101–4111. [Google Scholar] [CrossRef] [PubMed]
- Leuner, K.; Heiser, J.H.; Derksen, S.; Mladenov, M.I.; Fehske, C.J.; Schubert, R.; Gollasch, M.; Schneider, G.; Harteneck, C.; Chatterjee, S.S.; et al. Simple 2,4-Diacylphloroglucinols as classic transient receptor Potential-6 activators—Identification of a novel pharmacophore. Mol. Pharmacol. 2010, 77, 368–377. [Google Scholar] [CrossRef]
- García-Ávila, M.; Islas, L.D. What is new about mild temperature sensing? A review of recent findings. Temperature 2019, 6, 132–141. [Google Scholar] [CrossRef]
- Kraft, R.; Grimm, C.; Frenzel, H.; Harteneck, C. Inhibition of TRPM2 cation channels by N-(p-amylcinnamoyl)anthranilic acid. Br. J. Pharmacol. 2006, 148, 264–273. [Google Scholar] [CrossRef]
- Hill, K.; McNulty, S.; Randall, A.D. Inhibition of TRPM2 channels by the antifungal agents clotrimazole and econazole. Naunyn. Schmiedebergs. Arch. Pharmacol. 2004, 370, 227–237. [Google Scholar] [CrossRef]
- Hill, K.; Benham, C.; McNulty, S.; Randall, A. Flufenamic acid is a pH-dependent antagonist of TRPM2 channels. Neuropharmacology 2004, 47, 450–460. [Google Scholar] [CrossRef]
- Malysz, J.; Maxwell, S.; Yarotskyy, V.; Petkov, G.V. Compound-dependent effects of TRPM4 Channel Modulators on Guinea Pig detrusor smooth muscle excitability and contractility. FASEB J. 2019, 33, 837.4. [Google Scholar] [CrossRef]
- Takezawa, R.; Cheng, H.; Beck, A.; Ishikawa, J.; Launay, P.; Kubota, H.; Kinet, J.-P.; Fleig, A.; Yamada, T.; Penner, R. A Pyrazole derivative potently inhibits lymphocyte Ca2+ Influx and cytokine production by facilitating transient Receptor Potential Melastatin 4 Channel Activity. Mol. Pharmacol. 2006, 69, 1413–1420. [Google Scholar] [CrossRef]
- Wong, R.; Turlova, E.; Feng, Z.-P.; Rutka, J.T.; Sun, H.-S. Activation of TRPM7 by naltriben enhances migration and invasion of glioblastoma cells. Oncotarget 2017, 8, 11239–11248. [Google Scholar] [CrossRef]
- Saldanha, S.; Grimm, C.; Mercer, B.; Choi, J.; Allais, C.; Roush, W.; Heller, S.; Hodder, P. Agonists of Transient Receptor Potential Channels 3 and 2 (TRPML3 & TRPML2). Probe Rep. NIH Mol. Libr. Progr. Internet 2009, 2, 1–21. [Google Scholar]
- Plesch, E.; Chen, C.-C.; Butz, E.; Scotto Rosato, A.; Krogsaeter, E.K.; Yinan, H.; Bartel, K.; Keller, M.; Robaa, D.; Teupser, D.; et al. Selective agonist of TRPML2 reveals direct role in chemokine release from innate immune cells. Elife 2018, 7, e39720. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, Y.; Szallasi, A. Transient receptor potential (TRP) channels: A clinical perspective. Br. J. Pharmacol. 2014, 171, 2474–2507. [Google Scholar] [CrossRef] [PubMed]
- Chow, J.; Norng, M.; Zhang, J.; Chai, J. TRPV6 mediates capsaicin-induced apoptosis in gastric cancer cells--Mechanisms behind a possible new “hot” cancer treatment. Biochim. Biophys. Acta 2007, 1773, 565–576. [Google Scholar] [CrossRef]
- Gees, M.; Colsoul, B.; Nilius, B. The Role of transient receptor potential cation channels in Ca2+ signaling. Cold Spring Harb. Perspect. Biol. 2010, 2, a003962. [Google Scholar] [CrossRef]
- Lev, S.; Zeevi, D.A.; Frumkin, A.; Offen-Glasner, V.; Bach, G.; Minke, B. Constitutive activity of the human TRPML2 Channel induces cell degeneration. J. Biol. Chem. 2010, 285, 2771–2782. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.P.; Wang, X.; Xu, H. TRP channels of intracellular membranes. J. Neurochem. 2010, 113, 313–328. [Google Scholar] [CrossRef]
- Caterina, M.J.; Pang, Z. TRP channels in skin biology and pathophysiology. Pharmaceuticals 2016, 9, 77. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Merritt, R.; Fu, L.; Pan, Z. Targeting calcium signaling in cancer therapy. Acta Pharm. Sin. B 2017, 7, 3–17. [Google Scholar] [CrossRef]
- Hempel, N.; Trebak, M. Crosstalk between calcium and reactive oxygen species signaling in cancer. Cell Calcium 2017, 63, 70–96. [Google Scholar] [CrossRef]
- Arber, D.A.; Orazi, A.; Hasserjian, R.; Thiele, J.; Borowitz, M.J.; Le Beau, M.M.; Bloomfield, C.D.; Cazzola, M.; Vardiman, J.W. The 2016 revision to the World Health Organization classification of myeloid neoplasms and acute leukemia. Blood 2016, 127, 2391–2405. [Google Scholar] [CrossRef]
- Taylor, J.; Xiao, W.; Abdel-Wahab, O. Diagnosis and classification of hematologic malignancies on the basis of genetics. Blood 2017, 130, 410–423. [Google Scholar] [CrossRef]
- Morelli, M.B.; Liberati, S.; Amantini, C.; Nabiss, M.; Santoni, M.; Farfariello, V.; Santoni, G. Expression and function of the transient receptor potential ion channel family in the hematologic malignancies. Curr. Mol. Pharmacol. 2013, 6, 137–148. [Google Scholar] [CrossRef]
- Tomilin, V.N.; Cherezova, A.L.; Negulyaev, Y.A.; Semenova, S.B. TRPV5/V6 Channels Mediate Ca 2+ Influx in Jurkat T Cells under the control of extracellular pH. J. Cell. Biochem. 2016, 117, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Vassilieva, I.O.; Tomilin, V.N.; Marakhova, I.I.; Shatrova, A.N.; Negulyaev, Y.A.; Semenova, S.B. Expression of transient receptor potential vanilloid channels TRPV5 and TRPV6 in human blood lymphocytes and Jurkat leukemia T cells. J. Membr. Biol. 2013, 246, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Punzo, F.; Manzo, I.; Tortora, C.; Pota, E.; D’Angelo, V.; Bellini, G.; Di Paola, A.; Verace, F.; Casale, F.; Rossi, F. Effects of CB2 and TRPV1 receptors’ stimulation in pediatric acute T-lymphoblastic leukemia. Oncotarget 2018, 9, 21244–21258. [Google Scholar] [CrossRef][Green Version]
- Chatterton, Z.; Morenos, L.; Mechinaud, F.; Ashley, D.M.; Craig, J.M.; Sexton-Oates, A.; Halemba, M.S.; Parkinson-Bates, M.; Ng, J.; Morrison, D.; et al. Epigenetic deregulation in pediatric acute lymphoblastic leukemia. Epigenetics 2014, 9, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Almamun, M.; Levinson, B.T.; van Swaay, A.C.; Johnson, N.T.; McKay, S.D.; Arthur, G.L.; Davis, J.W.; Taylor, K.H. Integrated methylome and transcriptome analysis reveals novel regulatory elements in pediatric acute lymphoblastic leukemia. Epigenetics 2015, 10, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Pottosin, I.; Delgado-Enciso, I.; Bonales-Alatorre, E.; Nieto-Pescador, M.G.; Moreno-Galindo, E.G.; Dobrovinskaya, O. Mechanosensitive Ca(2)(+)-permeable channels in human leukemic cells: Pharmacological and molecular evidence for TRPV2. Biochim. Biophys. Acta 2015, 1848, 51–59. [Google Scholar] [CrossRef]
- Wenning, A.S.; Neblung, K.; Strauß, B.; Wolfs, M.J.; Sappok, A.; Hoth, M.; Schwarz, E.C. TRP expression pattern and the functional importance of TRPC3 in primary human T-cells. Biochim. Biophys. Acta Mol. Cell Res. 2011, 1813, 412–423. [Google Scholar] [CrossRef]
- Klumpp, D.; Misovic, M.; Szteyn, K.; Shumilina, E.; Rudner, J.; Huber, S.M. Targeting TRPM2 channels impairs radiation-induced cell cycle arrest and fosters cell death of T cell leukemia cells in a Bcl-2-dependent manner. Oxid. Med. Cell. Longev. 2016, 2016. [Google Scholar] [CrossRef]
- Bobkov, D.; Yudintceva, N.; Lomert, E.; Shatrova, A.; Kever, L.; Semenova, S. Lipid raft integrity is required for human leukemia Jurkat T-cell migratory activity. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2021, 1866, 2–11. [Google Scholar] [CrossRef]
- Abdoul-Azize, S.; Buquet, C.; Vannier, J.P.; Dubus, I. Pyr3, a TRPC3 channel blocker, potentiates dexamethasone sensitivity and apoptosis in acute lymphoblastic leukemia cells by disturbing Ca 2+ signaling, mitochondrial membrane potential changes and reactive oxygen species production. Eur. J. Pharmacol. 2016, 784, 90–98. [Google Scholar] [CrossRef]
- Siveen, K.S.; Prabhu, K.S.; Parray, A.S.; Merhi, M.; Arredouani, A.; Chikri, M.; Uddin, S.; Dermime, S.; Mohammad, R.M.; Steinhoff, M.; et al. Evaluation of cationic channel TRPV2 as a novel biomarker and therapeutic target in Leukemia-Implications concerning the resolution of pulmonary inflammation. Sci. Rep. 2019, 9, 1554. [Google Scholar] [CrossRef]
- Tian, C.; Huang, R.; Tang, F.; Lin, Z.; Cheng, N.; Han, X.; Li, S.; Zhou, P.; Deng, S.; Huang, H.; et al. Transient Receptor Potential Ankyrin 1 Contributes to Lysophosphatidylcholine-Induced Intracellular Calcium Regulation and THP-1-Derived Macrophage Activation. J. Membr. Biol. 2020, 253, 43–55. [Google Scholar] [CrossRef]
- Tian, C.; Han, X.; He, L.; Tang, F.; Huang, R.; Lin, Z.; Li, S.; Deng, S.; Xu, J.; Huang, H.; et al. Transient receptor potential ankyrin 1 contributes to the ATP-elicited oxidative stress and inflammation in THP-1-derived macrophage. Mol. Cell. Biochem. 2020, 473, 179–192. [Google Scholar] [CrossRef]
- Miller, B.A. TRPM2 in Cancer. Cell Calcium 2019, 80, 8–17. [Google Scholar] [CrossRef]
- Chen, S.; Bao, L.; Keefer, K.; Shanmughapriya, S.; Chen, L.; Lee, J.; Wang, J.; Zhang, X.-Q.; Hirschler-Laszkiewicz, I.; Merali, S.; et al. Transient receptor potential ion channel TRPM2 promotes AML proliferation and survival through modulation of mitochondrial function, ROS, and autophagy. Cell Death Dis. 2020, 11, 247. [Google Scholar] [CrossRef]
- Garaud, S.; Taher, T.E.; Debant, M.; Burgos, M.; Melayah, S.; Berthou, C.; Parikh, K.; Pers, J.O.; Luque-Paz, D.; Chiocchia, G.; et al. CD5 expression promotes IL-10 production through activation of the MAPK/Erk pathway and upregulation of TRPC1 channels in B lymphocytes. Cell. Mol. Immunol. 2018, 15, 158–170. [Google Scholar] [CrossRef] [PubMed]
- Cabanas, H.; Harnois, T.; Magaud, C.; Cousin, L.; Constantin, B.; Bourmeyster, N.; Déliot, N. Deregulation of calcium homeostasis in Bcr-Abl-dependent chronic myeloid leukemia. Oncotarget 2018, 9, 26309–26327. [Google Scholar] [CrossRef]
- Takahashi, K.; Umebayashi, C.; Numata, T.; Honda, A.; Ichikawa, J.; Hu, Y.; Yamaura, K.; Inoue, R. TRPM7-mediated spontaneous Ca2+ entry regulates the proliferation and differentiation of human leukemia cell line K562. Physiol. Rep. 2018, 6, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Liu, B.; Lu, X.; Yang, L.; Zhai, Y.; Eaton, A.F.; Thai, T.L.; Eaton, D.C.; Ma, H.; Shen, B. Biochimica et Biophysica Acta Lovastatin inhibits human B lymphoma cell proliferation by reducing intracellular ROS and TRPC6 expression. BBA Mol. Cell Res. 2014, 1843, 894–901. [Google Scholar] [CrossRef]
- Carter, B.Z.; Mak, D.H.; Cortes, J.; Andreeff, M. The elusive chronic myeloid leukemia stem cell: Does it matter and how do we eliminate it? Semin. Hematol. 2010, 47, 362–370. [Google Scholar] [CrossRef][Green Version]
- Hochhaus, A.; Reiter, A.; Skladny, H.; Melo, J.V.; Sick, C.; Berger, U.; Guo, J.Q.; Arlinghaus, R.B.; Hehlmann, R.; Goldman, J.M.; et al. A novel BCR-ABL fusion gene (e6a2) in a patient with Philadelphia chromosome-negative chronic myelogenous leukemia. Blood 1996, 88, 2236–2240. [Google Scholar] [CrossRef]
- Vago, L.; Gojo, I. Immune escape and immunotherapy of acute myeloid leukemia. J. Clin. Investig. 2020, 130, 1552–1564. [Google Scholar] [CrossRef]
- Si, J.; Collins, S.J. Activated Ca 2+/Calmodulin-Dependent protein kinase IIγ is a critical regulator of myeloid leukemia cell proliferation. Cancer Res. 2008, 68, 3733–3742. [Google Scholar] [CrossRef]
- Heise, N.; Palme, D.; Misovic, M. cation channel-mediated Ca2+-entry and activation of Ca2+/calmodulin-dependent kinase II contribute to G2/M cell cycle arrest and survival of irradiated leukemia. Cell. Physiol. 2010, 597–608. [Google Scholar]
- De Kouchkovsky, I.; Abdul-Hay, M. Acute myeloid leukemia: A comprehensive review and 2016 update. Blood Cancer J. 2016, 6, e441. [Google Scholar] [CrossRef]
- Ferrara, F.; Lessi, F.; Vitagliano, O.; Birkenghi, E.; Rossi, G. Current therapeutic results and treatment options for older patients with relapsed acute myeloid leukemia. Cancers 2019, 11, 224. [Google Scholar] [CrossRef]
- Omari, S.A.; Adams, M.J.; Geraghty, D.P. TRPV1 Channels in Immune Cells and Hematological Malignancies, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2017; Volume 79. [Google Scholar]
- Bujak, J.K.; Kosmala, D.; Szopa, I.M.; Majchrzak, K.; Bednarczyk, P. Inflammation, cancer and immunity—implication of TRPV1 Channel. Front. Oncol. 2019, 9. [Google Scholar] [CrossRef]
- Omari, S.A.; Adams, M.J.; Kunde, D.A.; Geraghty, D.P. Capsaicin-Induced Death of Human Haematological Malignant Cell Lines is Independent of TRPV1 Activation. Pharmacology 2016, 98, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Kunde, D.A.; Yingchoncharoen, J.; Jurković, S.; Geraghty, D.P. TRPV1 mediates capsaicin-stimulated metabolic activity but not cell death or inhibition of interleukin-1β release in human THP-1 monocytes. Toxicol. Appl. Pharmacol. 2018, 360, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Siveen, K.S.; Nizamuddin, P.B.; Uddin, S.; Al-Thani, M.; Frenneaux, M.P.; Janahi, I.A.; Steinhoff, M.; Azizi, F. TRPV2: A Cancer Biomarker and Potential Therapeutic Target. Dis. Markers 2020, 2020, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Leveque, M.; Penna, A.; Le Trionnaire, S.; Belleguic, C.; Desrues, B.; Brinchault, G.; Jouneau, S.; Lagadic-Gossmann, D.; Martin-Chouly, C. Phagocytosis depends on TRPV2-mediated calcium influx and requires TRPV2 in lipids rafts: Alteration in macrophages from patients with cystic fibrosis. Sci. Rep. 2018, 8, 4310. [Google Scholar] [CrossRef]
- Link, T.M.; Park, U.; Vonakis, B.M.; Raben, D.M.; Soloski, M.J.; Caterina, M.J. TRPV2 has a pivotal role in macrophage particle binding and phagocytosis. Nat. Immunol. 2010, 11, 232–239. [Google Scholar] [CrossRef]
- Yamashiro, K.; Sasano, T.; Tojo, K.; Namekata, I.; Kurokawa, J.; Sawada, N.; Suganami, T.; Kamei, Y.; Tanaka, H.; Tajima, N.; et al. Role of transient receptor potential vanilloid 2 in LPS-induced cytokine production in macrophages. Biochem. Biophys. Res. Commun. 2010, 398, 284–289. [Google Scholar] [CrossRef]
- Issa, C.M.; Hambly, B.D.; Wang, Y.; Maleki, S.; Wang, W.; Fei, J.; Bao, S. TRPV2 in the development of experimental colitis. Scand. J. Immunol. 2014, 80, 307–312. [Google Scholar] [CrossRef]
- Haladyna, J.N.; Pastuer, T.; Riedel, S.S.; Perraud, A.; Bernt, K.M. Transient potential receptor melastatin-2 (Trpm2) does not influence murine MLL-AF9-driven AML leukemogenesis or in vitro response to chemotherapy. Exp. Hematol. 2016, 44, 596–602. [Google Scholar] [CrossRef]
- Canales, J.; Morales, D.; Blanco, C.; Rivas, J.; Díaz, N.; Angelopoulos, I.; Cerda, O. A TR(i)P to cell migration: New roles of TRP channels in mechanotransduction and cancer. Front. Physiol. 2019, 10. [Google Scholar] [CrossRef]
- Diez-Bello, R.; Jardin, I.; Salido, G.M.; Rosado, J.A. Orai1 and Orai2 mediate store-operated calcium entry that regulates HL60 cell migration and FAK phosphorylation. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 1064–1070. [Google Scholar] [CrossRef]
- Guntur, A.R.; Gu, P.; Takle, K.; Chen, J.; Xiang, Y.; Yang, C.-H. Drosophila TRPA1 isoforms detect UV light via photochemical production of H2O2. Proc. Natl. Acad. Sci. USA 2015, 112, E5753–E5761. [Google Scholar] [CrossRef]
- Hajna, Z.; Sághy, É.; Payrits, M.; Aubdool, A.A.; Szőke, É.; Pozsgai, G.; Bátai, I.Z.; Nagy, L.; Filotás, D.; Helyes, Z.; et al. Capsaicin-sensitive sensory nerves mediate the cellular and microvascular effects of H2S via TRPA1 receptor activation and neuropeptide release. J. Mol. Neurosci. 2016, 60, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Pereira, I.; Mendes, S.J.F.; Pereira, D.M.S.; Muniz, T.F.; Colares, V.L.P.; Monteiro, C.R.A.V.; Martins, M.M.R.d.S.; Grisotto, M.A.G.; Monteiro-Neto, V.; Monteiro, S.G.; et al. Transient receptor potential ankyrin 1 channel expression on peripheral blood leukocytes from rheumatoid arthritic patients and correlation with pain and disability. Front. Pharmacol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, V.; Swain, S.; Murray, K.; Reardon, C. Neural Immune Communication in the Control of Host-Bacterial Pathogen Interactions in the Gastrointestinal Tract. Infect. Immun. 2020, 88. [Google Scholar] [CrossRef]
- Kun, J.; Perkecz, A.; Knie, L.; Sétáló, G.; Tornóczki, T.; Pintér, E.; Bán, Á. TRPA1 receptor is upregulated in human oral lichen planus. Oral Dis. 2017, 23, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Silverman, H.A.; Chen, A.; Kravatz, N.L.; Chavan, S.S.; Chang, E.H. Involvement of neural transient receptor potential channels in peripheral inflammation. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, S.S.; Majhi, R.K.; Tiwari, A.; Acharya, T.; Kumar, P.S.; Saha, S.; Kumar, A.; Goswami, C.; Chattopadhyay, S. Transient receptor potential ankyrin1 channel is endogenously expressed in T cells and is involved in immune functions. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef]
- Zhao, J.-F.; Shyue, S.-K.; Kou, Y.R.; Lu, T.-M.; Lee, T.-S. Transient Receptor potential ankyrin 1 channel involved in atherosclerosis and macrophage-foam cell formation. Int. J. Biol. Sci. 2016, 12, 812–823. [Google Scholar] [CrossRef]
- Bertin, S.; Aoki-Nonaka, Y.; Lee, J.; de Jong, P.R.; Kim, P.; Han, T.; Yu, T.; To, K.; Takahashi, N.; Boland, B.S.; et al. The TRPA1 ion channel is expressed in CD4+ T cells and restrains T-cell-mediated colitis through inhibition of TRPV1. Gut 2017, 66, 1584–1596. [Google Scholar] [CrossRef] [PubMed]
- Tefferi, A. The history of myeloproliferative disorders: Before and after Dameshek. Leukemia 2008, 22, 3–13. [Google Scholar] [CrossRef]
- Westerweel, P.E.; Te Boekhorst, P.A.W.; Levin, M.-D.; Cornelissen, J.J. New approaches and treatment combinations for the management of chronic myeloid leukemia. Front. Oncol. 2019, 9, 665. [Google Scholar] [CrossRef] [PubMed]
- Yee, N.S. Role of TRPM7 in cancer: Potential as molecular biomarker and therapeutic target. Pharmaceuticals 2017, 10, 39. [Google Scholar] [CrossRef] [PubMed]
- Ong, H.L.; de Souza, L.B.; Ambudkar, I.S. Role of TRPC Channels in store-operated calcium entry. In Advances in Experimental Medicine and Biology; Springer: Berlin/Heidelberg, Germany, 2016; pp. 87–109. [Google Scholar]
- Shim, A.H.-R.; Tirado-Lee, L.; Prakriya, M. Structural and functional mechanisms of CRAC Channel Regulation. J. Mol. Biol. 2015, 427, 77–93. [Google Scholar] [CrossRef] [PubMed]
- Ambudkar, I.S.; de Souza, L.B.; Ong, H.L. TRPC1, Orai1, and STIM1 in SOCE: Friends in tight spaces. Cell Calcium 2017, 63, 33–39. [Google Scholar] [CrossRef]
- Kuang, C.; Yu, Y.; Wang, K.; Qian, D.; Den, M.; Huang, L. Knockdown of transient receptor potential canonical-1 reduces the proliferation and migration of endothelial progenitor cells. Stem Cells Dev. 2012, 21, 487–496. [Google Scholar] [CrossRef]
- Bosch, F.; Dalla-Favera, R. Chronic lymphocytic leukaemia: From genetics to treatment. Nat. Rev. Clin. Oncol. 2019, 16, 684–701. [Google Scholar] [CrossRef]
- Iacobucci, I.; Mullighan, C.G. Genetic basis of acute lymphoblastic leukemia. J. Clin. Oncol. 2017, 35, 975–983. [Google Scholar] [CrossRef]
- Olivas-Aguirre, M.; Pottosin, I.; Dobrovinskaya, O. Mitochondria as emerging targets for therapies against T cell acute lymphoblastic leukemia. J. Leukoc. Biol. 2019, 105, 935–946. [Google Scholar] [CrossRef]
- Bozok Cetintas, V.; Tezcanli Kaymaz, B.; Aktug, H.; Oltulu, F.; Taskiran, D. Capsaicin induced apoptosis and gene expression dysregulation of human acute lymphoblastic leukemia CCRF-CEM cells. J. BUON 2014, 19, 183–190. [Google Scholar]
- Erra Díaz, F.; Dantas, E.; Geffner, J. Unravelling the interplay between extracellular acidosis and immune cells. Mediators Inflamm. 2018, 2018, 1–11. [Google Scholar] [CrossRef]
- Marum, L. Cancer cell line encyclopedia launched by novartis and broad institute. Future Med. Chem. 2012, 4, 947. [Google Scholar]
- Pedersen, S.F.; Stock, C. Ion channels and transporters in cancer: Pathophysiology, regulation, and clinical potential. Cancer Res. 2013, 73, 1658–1661. [Google Scholar] [CrossRef] [PubMed]
- Santoni, G.; Santoni, M.; Maggi, F.; Marinelli, O.; Morelli, M.B. Emerging Role of Mucolipins TRPML Channels in Cancer. Front. Oncol. 2020, 10, 659. [Google Scholar] [CrossRef] [PubMed]
- Hallek, M.; Shanafelt, T.D.; Eichhorst, B. Chronic lymphocytic leukaemia. Lancet 2018, 391, 1524–1537. [Google Scholar] [CrossRef]
- Takizawa, J. Chronic lymphocytic leukemia: Pathophysiology and current therapy. Rinsho. Ketsueki. 2017, 58, 471–479. [Google Scholar] [CrossRef]
- Cuneo, A.; Foà, R. Management of CLL in the elderly. Ann. Oncol. 2017, 28, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Hemon, P.; Renaudineau, Y.; Debant, M.; Le Goux, N.; Mukherjee, S.; Brooks, W.; Mignen, O. Calcium signaling: From Normal B Cell development to tolerance breakdown and autoimmunity. Clin. Rev. Allergy Immunol. 2017, 53, 141–165. [Google Scholar] [CrossRef] [PubMed]
- Debant, M.; Hemon, P.; Brigaudeau, C.; Renaudineau, Y.; Mignen, O. Calcium signaling and cell fate: How can Ca2+ signals contribute to wrong decisions for Chronic Lymphocytic Leukemic B lymphocyte outcome? Int. J. Dev. Biol. 2015, 59, 379–389. [Google Scholar] [CrossRef]
- Burdin, N.; Rousset, F.; Banchereau, J. B-Cell-Derived IL-10: Production and function. Methods 1997, 11, 98–111. [Google Scholar] [CrossRef]
- Gary-Gouy, H.; Harriague, J.; Bismuth, G.; Platzer, C.; Schmitt, C.; Dalloul, A.H. Human CD5 promotes B-cell survival through stimulation of autocrine IL-10 production. Blood 2002, 100, 4537–4543. [Google Scholar] [CrossRef]
- Inada, H.; Iida, T.; Tominaga, M. Different expression patterns of TRP genes in murine B and T lymphocytes. Biochem. Biophys. Res. Commun. 2006, 350, 762–767. [Google Scholar] [CrossRef]
- Vaeth, M.; Feske, S. NFAT control of immune function: New frontiers for an abiding trooper. F1000Research 2018, 7, 260. [Google Scholar] [CrossRef]
- Jaffe, E.S. Diagnosis and classification of lymphoma: Impact of technical advances. Semin. Hematol. 2019, 56, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Bennani, N.N.; Feldman, A.L. Lymphoma classification update: T-cell lymphomas, Hodgkin lymphomas, and histiocytic/dendritic cell neoplasms. Expert Rev. Hematol. 2017, 10, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Campo, E.; Swerdlow, S.H.; Harris, N.L.; Pileri, S.; Stein, H.; Jaffe, E.S. The 2008 WHO classification of lymphoid neoplasms and beyond: Evolving concepts and practical applications. Blood 2011, 117, 5019–5032. [Google Scholar] [CrossRef]
- Borgström, A.; Peinelt, C.; Stokłosa, P.; Uversky, N.; Morelli, M.B. TRPM4 in cancer-a new potential drug target. Biomolecules 2021, 11, 229. [Google Scholar] [CrossRef]
- Oliver, M.F. Might treatment of hypercholesterolaemia increase non-cardiac mortality? Lancet 1991, 337, 1529–1531. [Google Scholar] [CrossRef]
- Graaf, M.R.; Beiderbeck, A.B.; Egberts, A.C.G.; Richel, D.J.; Guchelaar, H.-J. The risk of cancer in users of statins. J. Clin. Oncol. 2004, 22, 2388–2394. [Google Scholar] [CrossRef] [PubMed]
- Demierre, M.-F.; Higgins, P.D.R.; Gruber, S.B.; Hawk, E.; Lippman, S.M. Statins and cancer prevention. Nat. Rev. Cancer 2005, 5, 930–942. [Google Scholar] [CrossRef]
- Jacobs, E.J.; Newton, C.C.; Thun, M.J.; Gapstur, S.M. Long-term use of cholesterol-lowering drugs and cancer incidence in a large United States Cohort. Cancer Res. 2011, 71, 1763–1771. [Google Scholar] [CrossRef] [PubMed]
- Tonomura, N.; Elvers, I.; Thomas, R.; Megquier, K.; Turner-Maier, J.; Howald, C.; Sarver, A.L.; Swofford, R.; Frantz, A.M.; Ito, D.; et al. Genome-wide association study identifies shared risk loci common to two malignancies in Golden Retrievers. PLoS Genet. 2015, 11, e1004922. [Google Scholar] [CrossRef]
- Shapovalov, G.; Ritaine, A.; Skryma, R.; Prevarskaya, N. Role of TRP ion channels in cancer and tumorigenesis. Semin. Immunopathol. 2016, 38, 357–369. [Google Scholar] [CrossRef]
- Santoni, G.; Farfariello, V. TRP channels and cancer: New targets for diagnosis and chemotherapy. Endocr. Metab. Immune Disord. Drug Targets 2011, 11, 54–67. [Google Scholar] [CrossRef] [PubMed]
- Liberati, S.; Morelli, M.B.; Nabissi, M.; Santoni, M.; Santoni, G. Oncogenic and anti-oncogenic effects of transient receptor potential channels. Curr. Top. Med. Chem. 2013, 13, 344–366. [Google Scholar] [CrossRef] [PubMed]
- Smani, T.; Shapovalov, G.; Skryma, R.; Prevarskaya, N.; Rosado, J.A. Functional and physiopathological implications of TRP channels. Biochim. Biophys. Acta Mol. Cell Res. 2015, 1853, 1772–1782. [Google Scholar] [CrossRef]
- Santoni, G.; Morelli, M.B.; Marinelli, O.; Nabissi, M.; Santoni, M.; Amantini, C. Calcium signaling and the regulation of chemosensitivity in cancer cells: Role of the transient receptor potential channels. Adv. Exp. Med. Biol. 2020, 1131, 505–517. [Google Scholar] [PubMed]
- Gil-Kulik, P.; Dudzińska, E.; Radzikowska-Büchner, E.; Wawer, J.; Jojczuk, M.; Nogalski, A.; Wawer, G.A.; Feldo, M.; Kocki, W.; Cioch, M.; et al. Different regulation of PARP1, PARP2, PARP3 and TRPM2 genes expression in acute myeloid leukemia cells. BMC Cancer 2020, 20, 1–9. [Google Scholar] [CrossRef]
- Yang, S.; Lu, W.; Zhao, C.; Zhai, Y.; Wei, Y.; Liu, J.; Yu, Y.; Li, Z.; Shi, J. Leukemia cells remodel marrow adipocytes via TRPV4-dependent lipolysis. Haematologica 2019. [Google Scholar] [CrossRef]
- Debant, M.; Burgos, M.; Hemon, P.; Buscaglia, P.; Fali, T.; Melayah, S.; Le Goux, N.; Vandier, C.; Potier-Cartereau, M.; Pers, J.O.; et al. STIM1 at the plasma membrane as a new target in progressive chronic lymphocytic leukemia. J. Immunother. Cancer 2019, 7, 1–13. [Google Scholar] [CrossRef]
- Hirai, A.; Aung, N.Y.; Ohe, R.; Nishida, A.; Kato, T.; Meng, H.; Ishizawa, K.; Fujii, J.; Yamakawa, M. Expression of TRPM8 in human reactive lymphoid tissues and mature B-cell neoplasms. Oncol. Lett. 2018, 16, 5930–5938. [Google Scholar] [CrossRef]
- Loo, S.K.; Ch’ng, E.S.; Md Salleh, M.S.; Banham, A.H.; Pedersen, L.M.; Moller, M.B.; Green, T.M.; Wong, K.K. TRPM4 expression is associated with activated B cell subtype and poor survival in diffuse large B cell lymphoma. Histopathology 2017, 71, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Bowen, C.V.; DeBay, D.; Ewart, H.S.; Gallant, P.; Gormley, S.; Ilenchuk, T.T.; Iqbal, U.; Lutes, T.; Martina, M.; Mealing, G.; et al. In vivo detection of human TRPV6-rich tumors with anti-cancer peptides derived from soricidin. PLoS ONE 2013, 8, e58866. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Gao, Y.; Wei, S.; Low, S.W.; Ng, G.; Yu, D.; Tu, T.M.; Soong, T.W.; Nilius, B.; Liao, P. TRPM4-specific blocking antibody attenuates reperfusion injury in a rat model of stroke. Pflügers Arch. Eur. J. Physiol. 2019, 471, 1455–1466. [Google Scholar] [CrossRef] [PubMed]



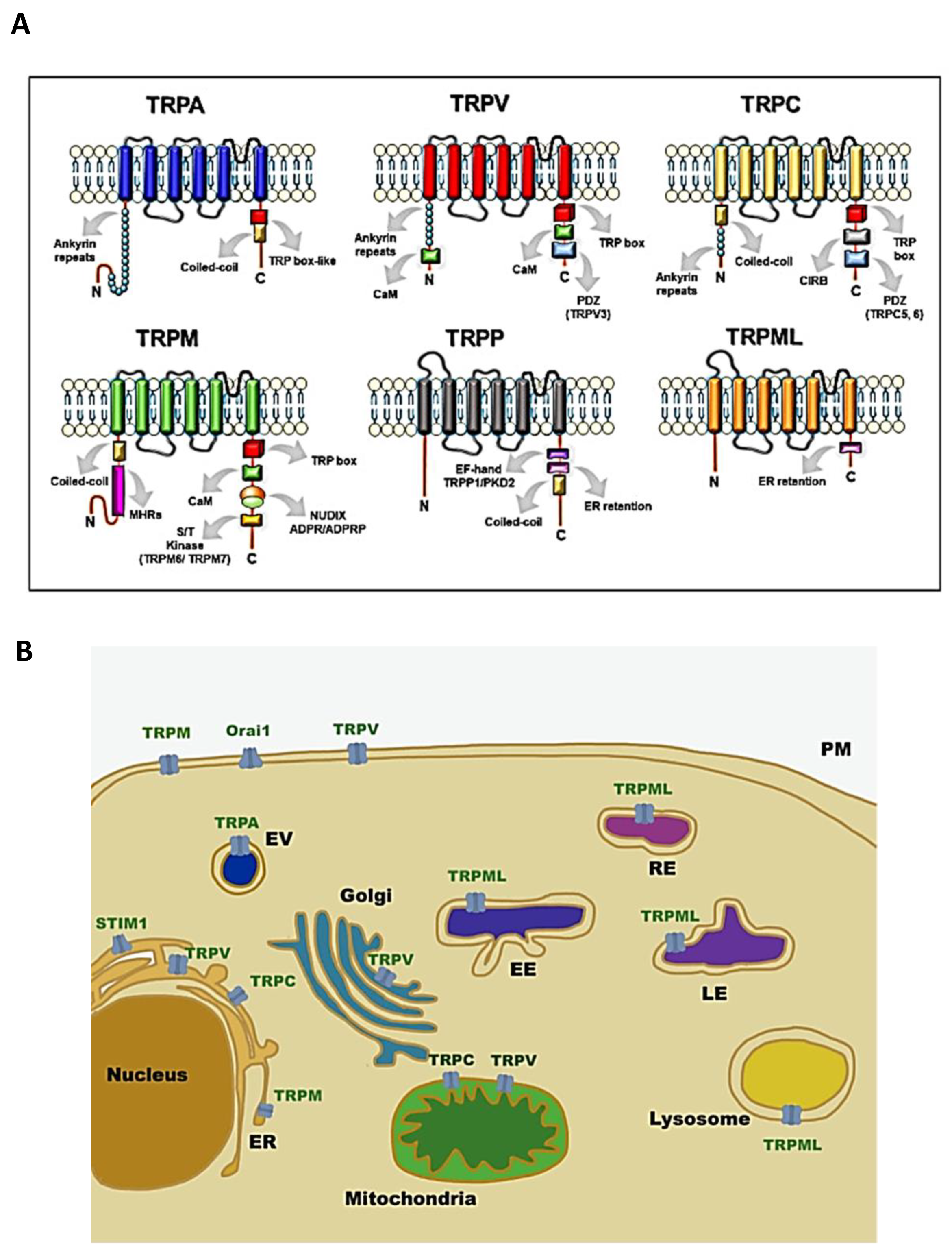{kind=link}
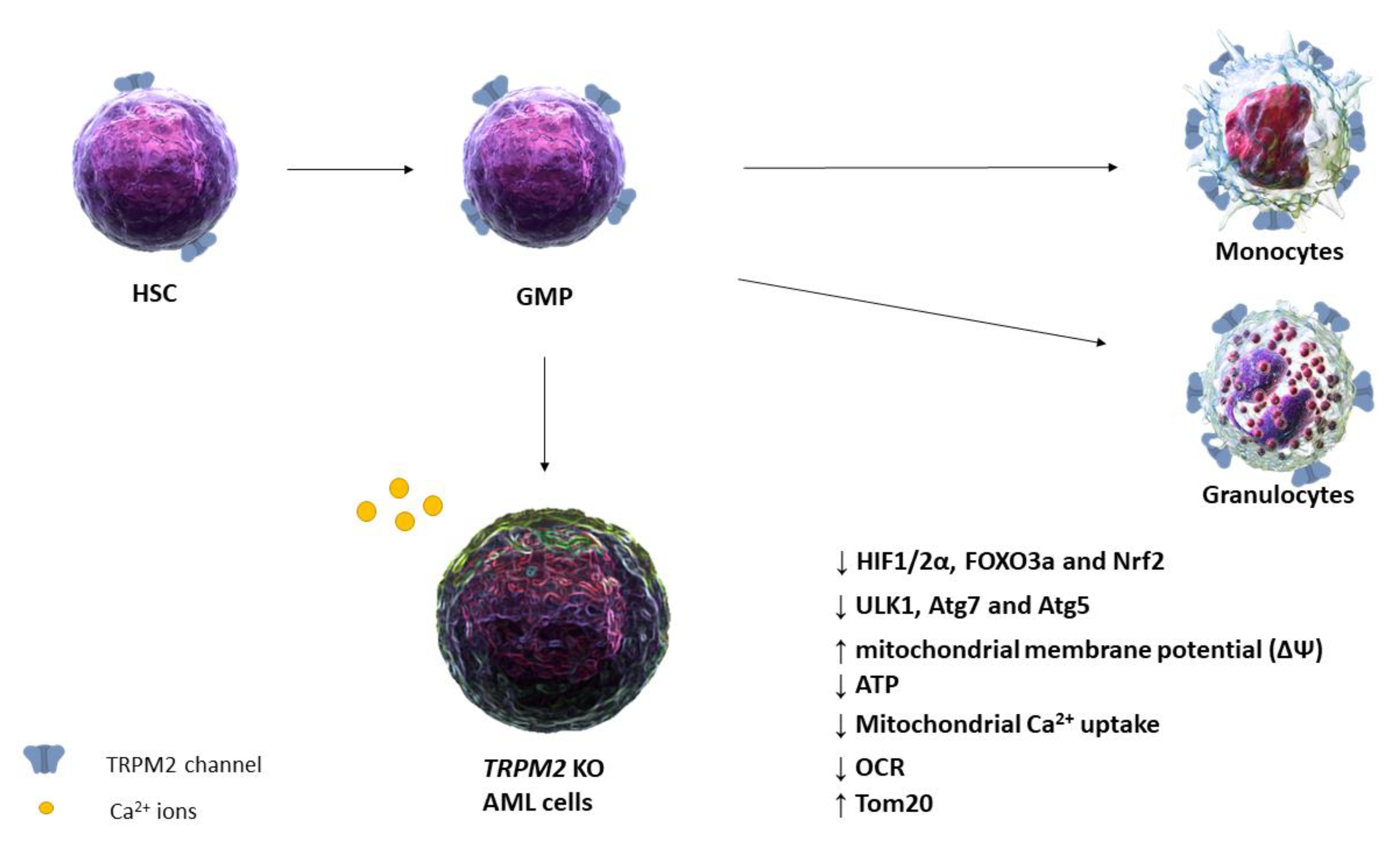{kind=link}
| TRP | Ca2+ Selectivity | Activation Temperature | Exogenous Activators | References |
|---|---|---|---|---|
| TRPA1 | Medium/Low | <17 °C | Caffeine, Cinnamaldehyde, Nicotine | [8,9,10] |
| TRPC1 | Medium | Lanthanide ions (La3+, Gd3+), carbachol | [11] | |
| TRPC3 | Medium | OAG | [12] | |
| TRPC6 | Medium | OAG, Hyperforin, 2,4-Diacylphloroglucinol | [12,13,14] | |
| TRPM2 | Medium/Low | >38 °C | N-(p-amylcinnamoyl)anthranilic acid, Clotrimazole, Flufenamic acid | [15,16,17,18] |
| TRPM4 | Non selective | 15–35 °C | BTP2 | [15,19,20] |
| TRPM7 | Medium/Low | Naltriben | [21] | |
| TRPM8 | Medium | 17–25 °C | Menthol, eucalyptol, geraniol | [6] |
| TRPML2 | Non selective | SID24801657, SID24787221, ML2SA1 | [22,23] | |
| TRPV1 | Medium/High | >42 °C | CPS, Piperine, Gingerol | [6,24] |
| TRPV2 | Medium | (≈52 °C) | 2-aminoethoxydiphenyl borate, diphenylyboronic anhydride, and Cannabis sativa derivatives | [6] |
| TRPV4 | Medium | 24–27 °C | Bisandrographolide A, Apigenin, 4-alpha-phorbol12,13-didecanoate | [6] |
| TRPV5 | High | - | ||
| TRPV6 | High | CPS | [25] |
| Cancer Type | Cell Line/Model | TRP | Methods | Functions | References |
|---|---|---|---|---|---|
| ALL | Jurkat | TRPV5/V6 | RT-PCR, WB | Early endosome formation | [35,36] |
| T-ALL patient | TRPV1 | RT-PCR | Proliferation Cell death | [37] | |
| B-ALL patient | TRPC | FISH, RT-PCR | (?) | [38] | |
| B-ALL patient | TRPML2 | RT-PCR | (?) | [39] | |
| Jurkat | TRPV2 | PCR | Deformation stretch-activated current | [40,41] | |
| Jurkat | TRPM2 | qRT-PCR, WB | ROS production | [42] | |
| Jurkat | TRPV1 | RT-PCR | Cell proliferation | [37] | |
| Jurkat | TRPV6 | RT-PCR, WB | Cell migration | [36,43] | |
| Nalm-6 Reh | TRPC3 | Functional studies | ROS production, Cell death | [44] | |
| AML | U937 THP1 | TRPV2 | RT-PCR, WB RT-PCR, WB | Cell growth, apoptosis, and cell cycle (?) | [45] |
| THP1 | TRPA1 | Immunofluorescence | Macrophage cytotoxicity | [46,47] | |
| U937 | TRPM2 | RT-PCR, WB | Autophagy | [48,49] | |
| CLL | Jok-1/E1A-E1B | TRPC1 | Flow cytometry | Cytokine production | [50] |
| CML | 32d and 32d-p210 | TRPC1 | WB | Cell proliferation | [51] |
| K562 | TRPM7 | RT-PCR, IF | Erythroid differentiation | [52] | |
| K562 | TRPV2 | RT-PCR, WB | Cell death | [45] | |
| Lymphoma | Daudi B-cell | TRPC6 | WB | Cell proliferation | [53] |
| Cancer Type | TRP | Effects | References |
|---|---|---|---|
| ALL | TRPC3 | ↓Glucocorticoid resistance | [44] |
| AML | TRPM2 | ↓OS ↓Doxo-sensitivity | [120] |
| TRPV4 | ↑Chemotherapy efficacy | [121] | |
| CLL | TRPC1 | OS | [122] |
| Lymphoma | TRPM8 | Prognostic | [123] |
| TRPM4 | ↓OS ↓PFS | [124] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maggi, F.; Morelli, M.B.; Nabissi, M.; Marinelli, O.; Zeppa, L.; Aguzzi, C.; Santoni, G.; Amantini, C. Transient Receptor Potential (TRP) Channels in Haematological Malignancies: An Update. Biomolecules 2021, 11, 765. https://doi.org/10.3390/biom11050765
Maggi F, Morelli MB, Nabissi M, Marinelli O, Zeppa L, Aguzzi C, Santoni G, Amantini C. Transient Receptor Potential (TRP) Channels in Haematological Malignancies: An Update. Biomolecules. 2021; 11(5):765. https://doi.org/10.3390/biom11050765
Chicago/Turabian StyleMaggi, Federica, Maria Beatrice Morelli, Massimo Nabissi, Oliviero Marinelli, Laura Zeppa, Cristina Aguzzi, Giorgio Santoni, and Consuelo Amantini. 2021. "Transient Receptor Potential (TRP) Channels in Haematological Malignancies: An Update" Biomolecules 11, no. 5: 765. https://doi.org/10.3390/biom11050765
APA StyleMaggi, F., Morelli, M. B., Nabissi, M., Marinelli, O., Zeppa, L., Aguzzi, C., Santoni, G., & Amantini, C. (2021). Transient Receptor Potential (TRP) Channels in Haematological Malignancies: An Update. Biomolecules, 11(5), 765. https://doi.org/10.3390/biom11050765