
GPCR-Based Bioactive Peptide Screening Using Phage-Displayed Peptides and an Insect Cell System for Insecticide Discovery



Abstract
1. Introduction
2. Materials and Methods
2.1. Insects, Cloning and Expression of the GPCR, and Binding Assay
2.2. Screening of Small Peptides Using Phage Display and Biopanning
2.3. Preparation of Phage and Ligand Peptides
2.4. Bioassays
2.5. Statistical Analysis
3. Results
3.1. Phage Display Library and Biopanning Using Sf9 Cell Expression
3.2. Analysis of Phage-Displayed PEPTIDES Resulting from the Biopanning
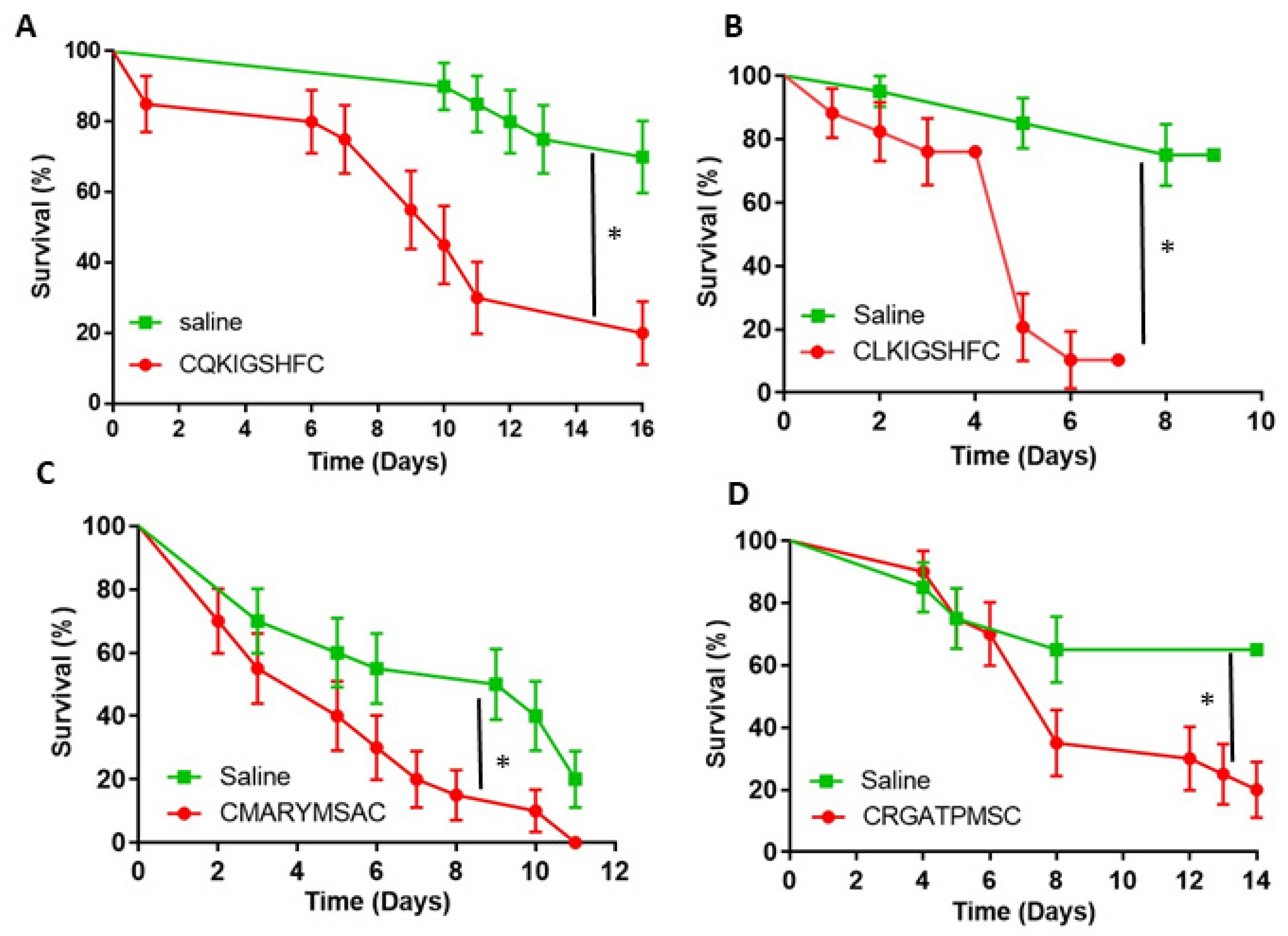
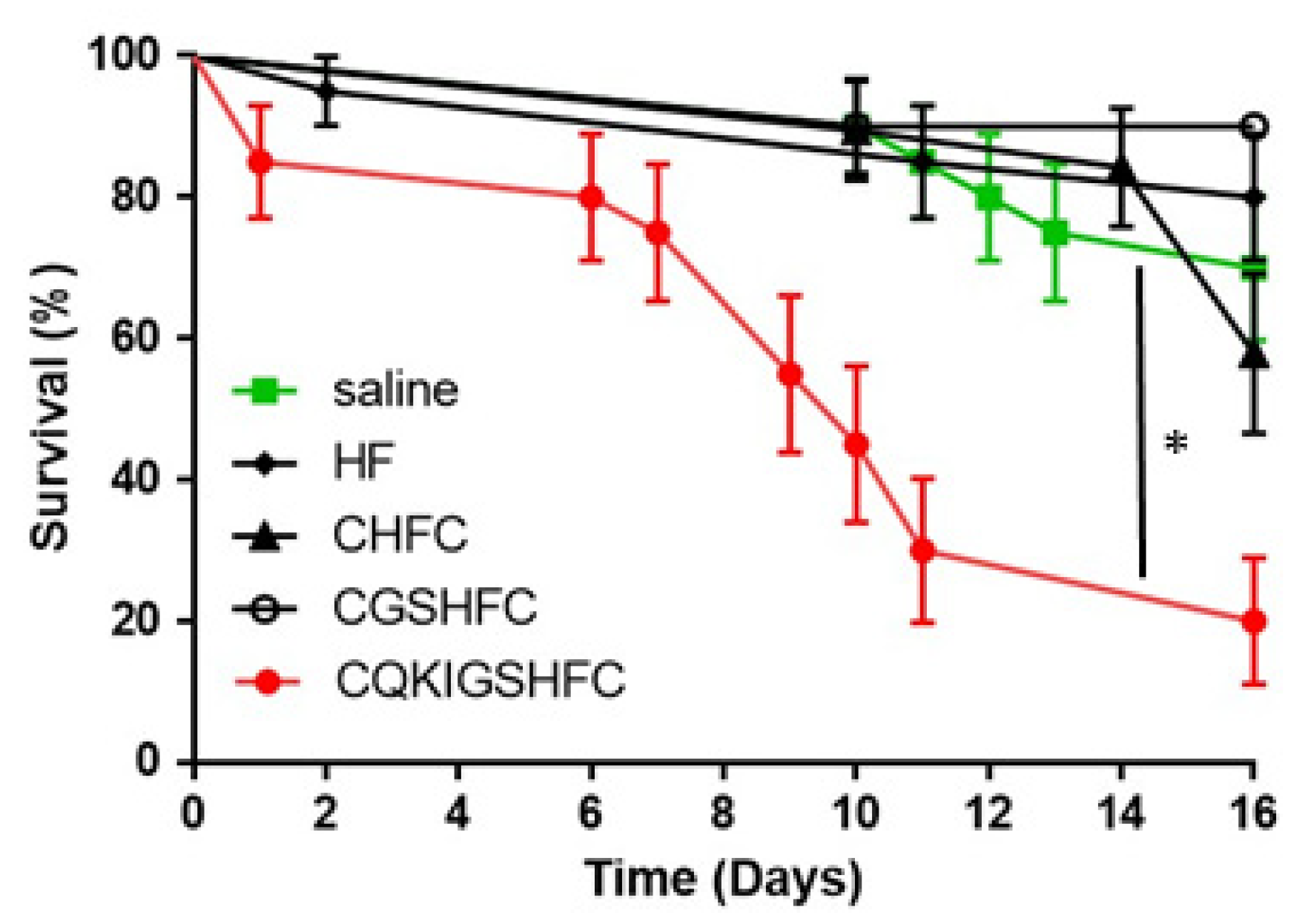
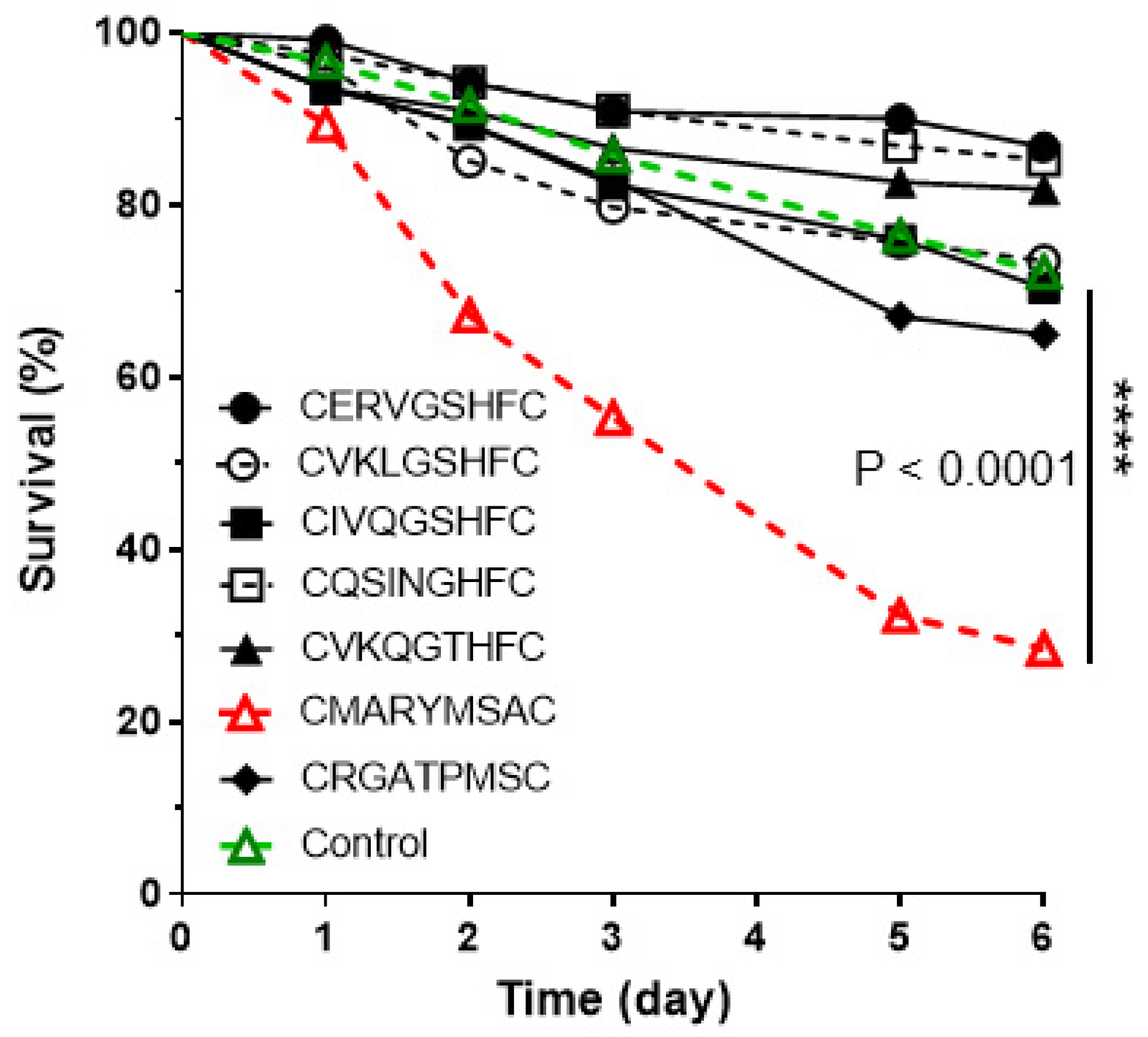
3.3. Phenotypic Impact of the Novel Peptides on Fire Ants
4. Discussion
4.1. GPCR-Based Screening Using a Library of Phage-Displayed Peptides and Biopanning
4.2. Small Peptides and PRX Peptides
4.3. GPCR-Based Insecticide Discovery: A Proof of Concept and Feasibility
5. Conclusions
6. Patents
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Woods, D.; Butler, C.; Williams, T.; Greenwood, K. Receptor-based discovery strategies for insecticides and parasiticides: A review. Adv. Exp. Med. Biol. 2010, 692, 1–9. [Google Scholar]
- Meyer, J.M.; Ejendal, K.F.; Avramova, L.V.; Garland-Kuntz, E.E.; Giraldo-Calderon, G.I.; Brust, T.F.; Watts, V.J.; Hill, C.A. A ‘genome-to-lead’ approach for insecticide discovery: Pharmacological characterization and screening of Aedes aegypti D(1)-like dopamine receptors. PLoS Negl. Trop. Dis. 2012, 6, e1478. [Google Scholar] [CrossRef]
- Grimmelikhuijzen, C.J.; Cazzamali, G.; Williamson, M.; Hauser, F. The promise of insect genomics. Pest Manag. Sci. 2007, 63, 413–416. [Google Scholar] [CrossRef]
- Geary, T.G. Nonpeptide ligands for peptidergic G protein-coupled receptors. Adv. Exp. Med. Biol. 2010, 692, 10–26. [Google Scholar] [CrossRef]
- Nassel, D.R.; Zandawala, M. Recent advances in neuropeptide signaling in Drosophila, from genes to physiology and behavior. Prog. Neurobiol. 2019, 179, 101607. [Google Scholar] [CrossRef] [PubMed]
- Caers, J.; Verlinden, H.; Zels, S.; Vandersmissen, H.P.; Vuerinckx, K.; Schoofs, L. More than two decades of research on insect neuropeptide GPCRs: An overview. Front. Endocrinol. 2012, 3, 151. [Google Scholar] [CrossRef] [PubMed]
- Yeoh, J.G.C.; Pandit, A.A.; Zandawala, M.; Nassel, D.R.; Davies, S.A.; Dow, J.A.T. DINeR: Database for insect neuropeptide research. Insect Biochem. Mol. Biol. 2017, 86, 9–19. [Google Scholar] [CrossRef]
- Audsley, N.; Down, R.E. G protein coupled receptors as targets for next generation pesticides. Insect Biochem. Mol. Biol. 2015, 67, 27–37. [Google Scholar] [CrossRef]
- Altstein, M.; Nassel, D.R. Neuropeptide signaling in insects. Adv. Exp. Med. Biol. 2010, 692, 155–165. [Google Scholar] [CrossRef]
- Jurenka, R. The PRXamide neuropeptide signalling system: Conserved in animals. Adv. Insect Physiol. 2015, 49, 123–170. [Google Scholar] [CrossRef]
- Hewes, R.S.; Taghert, P.H. Neuropeptides and neuropeptide receptors in the Drosophila melanogaster genome. Genome Res. 2001, 11, 1126–1142. [Google Scholar] [CrossRef]
- Vanden Broeck, J. Insect G protein-coupled receptors and signal transduction. Arch. Insect Biochem. Physiol. 2001, 48, 1–12. [Google Scholar] [CrossRef]
- Park, Y.; Kim, Y.J.; Adams, M.E. Identification of G protein-coupled receptors for Drosophila PRXamide peptides, CCAP, corazonin, and AKH supports a theory of ligand-receptor coevolution. Proc. Natl. Acad. Sci. USA 2002, 99, 11423–11428. [Google Scholar] [CrossRef] [PubMed]
- Howard, A.D.; Wang, R.; Pong, S.S.; Mellin, T.N.; Strack, A.; Guan, X.M.; Zeng, Z.; Williams, D.L., Jr.; Feighner, S.D.; Nunes, C.N.; et al. Identification of receptors for neuromedin U and its role in feeding. Nature 2000, 406, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.Y.; Vander Meer, R.K.; Coy, M.; Scharf, M.E. Phenotypic impacts of PBAN RNA interference in an ant, Solenopsis invicta, and a moth, Helicoverpa zea. J. Insect Physiol. 2012, 58, 1159–1165. [Google Scholar] [CrossRef]
- Choi, M.Y.; Vander Meer, R.K. Phenotypic effects of PBAN RNAi using oral delivery of dsRNA to corn earworm (Lepidoptera: Noctuidae) and tobacco budworm larvae. J. Econ. Entomol. 2019, 112, 434–439. [Google Scholar] [CrossRef]
- Lu, Q.; Huang, L.Y.; Chen, P.; Yu, J.F.; Xu, J.; Deng, J.Y.; Ye, H. Identification and RNA Interference of the pheromone biosynthesis activating neuropeptide (PBAN) in the common cutworm moth Spodoptera litura (Lepidoptera: Noctuidae). J. Econ. Entomol. 2015, 108, 1344–1353. [Google Scholar] [CrossRef]
- Choi, M.Y.; Vander Meer, R.K. Ant trail pheromone biosynthesis is triggered by a neuropeptide hormone. PLoS ONE 2012, 7, e50400. [Google Scholar] [CrossRef]
- Banks, W.A.; Lofgren, C.S.; Jouvenaz, D.P.; Stringer, C.E.; Bishop, P.M.; Williams, D.F.; Wojcik, D.P.; Glancey, B.M. Techniques for Collecting, Rearing, and Handling Imported Fire Ants; AAT-S-21; USDA Science and Education Administration: New Orleans, LA, USA, 1981; 9p.
- Choi, M.Y.; Fuerst, E.J.; Rafaeli, A.; Jurenka, R. Identification of a G protein-coupled receptor for pheromone biosynthesis activating neuropeptide from pheromone glands of the moth Helicoverpa zea. Proc. Natl. Acad. Sci. USA 2003, 100, 9721–9726. [Google Scholar] [CrossRef]
- Choi, M.Y.; Vander Meer, R.K. Bioactive Peptides Having Insecticide Activity. U.S. Patent 9771393B2, 26 September 2017. [Google Scholar]
- Choi, M.Y.; Vander Meer, R.K. Bioactive Peptides Having Insecticides Activity. U.S. Patent 10017358B2, 10 July 2018. [Google Scholar]
- Parmley, S.F.; Smith, G.P. Antibody-selectable filamentous fd phage vectors: Affinity purification of target genes. Gene 1988, 73, 305–318. [Google Scholar] [CrossRef]
- Christianson, D.R.; Ozawa, M.G.; Pasqualini, R.; Arap, W. Techniques to decipher molecular diversity by phage display. Methods Mol. Biol. 2007, 357, 385–406. [Google Scholar] [CrossRef]
- Scott, J.K.; Smith, G.P. Searching for peptide ligands with an epitope library. Science 1990, 249, 386–390. [Google Scholar] [CrossRef]
- Azzazy, H.M.; Highsmith, W.E., Jr. Phage display technology: Clinical applications and recent innovations. Clin. Biochem. 2002, 35, 425–445. [Google Scholar] [CrossRef]
- Sidhu, S.S.; Fairbrother, W.J.; Deshayes, K. Exploring protein-protein interactions with phage display. ChemBioChem 2003, 4, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Molek, P.; Strukelj, B.; Bratkovic, T. Peptide phage display as a tool for drug discovery: Targeting membrane receptors. Molecules 2011, 16, 857–887. [Google Scholar] [CrossRef]
- Giordano, R.J.; Cardo-Vila, M.; Lahdenranta, J.; Pasqualini, R.; Arap, W. Biopanning and rapid analysis of selective interactive ligands. Nat. Med. 2001, 7, 1249–1253. [Google Scholar] [CrossRef]
- Sergeeva, A.; Kolonin, M.G.; Molldrem, J.J.; Pasqualini, R.; Arap, W. Display technologies: Application for the discovery of drug and gene delivery agents. Adv. Drug Deliv. Rev. 2006, 58, 1622–1654. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Ribolla, P.E.; Jacobs-Lorena, M. Targeting Plasmodium ligands on mosquito salivary glands and midgut with a phage display peptide library. Proc. Natl. Acad. Sci. USA 2001, 98, 13278–13281. [Google Scholar] [CrossRef]
- Wess, J. Molecular basis of receptor/G-protein-coupling selectivity. Pharmacol. Ther. 1998, 80, 231–264. [Google Scholar] [CrossRef]
- Choi, M.Y.; Vander Meer, R.K. Identification of a new member of the PBAN family of neuropeptides from the fire ant, Solenopsis invicta. Insect Mol. Biol. 2009, 18, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Rafaeli, A. Pheromone biosynthesis activating neuropeptide (PBAN): Regulatory role and mode of action. Gen. Comp. Endocrinol. 2009, 162, 69–78. [Google Scholar] [CrossRef]
- Reagan, J.D. Expression cloning of an insect diuretic hormone receptor. A member of the calcitonin/secretin receptor family. J. Biol. Chem. 1994, 269, 9–12. [Google Scholar] [CrossRef]
- Brody, T.; Cravchik, A. Drosophila melanogaster G protein-coupled receptors. J. Cell Biol. 2000, 150, F83–F88. [Google Scholar] [CrossRef]
- Adams, M.D.; Celniker, S.E.; Holt, R.A.; Evans, C.A.; Gocayne, J.D.; Amanatides, P.G.; Scherer, S.E.; Li, P.W.; Hoskins, R.A.; Galle, R.F.; et al. The genome sequence of Drosophila melanogaster. Science 2000, 287, 2185–2195. [Google Scholar] [CrossRef]
- FlyBase. D. Melanogaster GPCR. Available online: http://flybase.org/ (accessed on 2 February 2020).
- Lee, J.M.; Hull, J.J.; Kawai, T.; Goto, C.; Kurihara, M.; Tanokura, M.; Nagata, K.; Nagasawa, H.; Matsumoto, S. Re-evaluation of the PBAN receptor molecule: Characterization of PBANR variants expressed in the pheromone glands of moths. Front. Endocrinol. 2012, 3, 6. [Google Scholar] [CrossRef] [PubMed]
- Nusawardani, T.; Kroemer, J.A.; Choi, M.Y.; Jurenka, R.A. Identification and characterization of the pyrokinin/pheromone biosynthesis activating neuropeptide family of G protein-coupled receptors from Ostrinia nubilalis. Insect Mol. Biol. 2013, 22, 331–340. [Google Scholar] [CrossRef]
- Jiang, L.; Zhang, F.; Hou, Y.; Thakur, K.; Hu, F.; Zhang, J.G.; Jiang, X.F.; Liu, Y.Q.; Wei, Z.J. Isolation and functional characterization of the pheromone biosynthesis activating neuropeptide receptor of Chinese oak silkworm, Antheraea pernyi. Int. J. Biol. Macromol. 2018, 117, 42–50. [Google Scholar] [CrossRef]
- Luo, M.; Zhou, X.C.; Wang, Z.; Chen, J.X.; Chung, H.; Wei, H.Y. Identification and gene expression analysis of the pheromone biosynthesis activating neuropeptide receptor (PBANR) from the Ostrinia furnacalis (Lepidoptera: Pyralidae). J. Insect Sci. 2019, 19, 25. [Google Scholar] [CrossRef]
- Ahn, S.J.; Corcoran, J.A.; Vander Meer, R.K.; Choi, M.Y. Identification and characterization of GPCRs for pyrokinin and capa peptides in the brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae). Front. Physiol. 2020, 11, 559. [Google Scholar] [CrossRef] [PubMed]
- Raina, A.K.; Jaffe, H.; Kempe, T.G.; Keim, P.; Blacher, R.W.; Fales, H.M.; Riley, C.T.; Klun, J.A.; Ridgway, R.L.; Hayes, D.K. Identification of a neuropeptide hormone that regulates sex pheromone production in female moths. Science 1989, 244, 796–798. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, S.; Kitamura, A.; Nagasawa, H.; Kataoka, H.; Orikasa, C.; Mitsui, T.; Suzuki, A. Functional diversity of a neurohormone produced by the suboesophageal ganglion: Molecular identity of melanization and reddish colouration hormone and pheromone biosynthesis activating neuropeptide. J. Insect Physiol. 1990, 36, 427–432. [Google Scholar] [CrossRef]
- Suwan, S.; Isobe, M.; Yamashita, O.; Minakata, H.; Imai, K. Silkworm diapause hormone, structure-activity relationships indispensable role of C-terminal amide. Insect Biochem. Mol. Biol. 1994, 24, 1001–1007. [Google Scholar] [CrossRef]
- Xu, W.H.; Denlinger, D.L. Molecular characterization of prothoracicotropic hormone and diapause hormone in Heliothis virescens during diapause, and a new role for diapause hormone. Insect Mol. Biol. 2003, 12, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Predel, R.; Nachman, R.J. Efficacy of native FXPRLamides (pyrokinins) and synthetic analogs on visceral muscles of the American cockroach. J. Insect Physiol. 2001, 47, 287–293. [Google Scholar] [CrossRef]
- Zdarek, J.; Nachman, R.J.; Hayes, T.K. Insect neuropeptides of the pyrokinin/PBAN family accelerate pupariation in the fleshfly (Sarcophaga bullata) larvae. Ann. N. Y. Acad. Sci. 1997, 814, 67–72. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phage Sequence (CXXXXXXXC) | Abundance of Phage Clones | Phage Sequence (CXXXXXXXC) | Abundance of Phage Clones |
|---|---|---|---|
| QKIGSHF | 11 | ERVGTHY | 1 |
| LKIGSHF | 7 | IKVGPHY | 1 |
| IQQGSHF | 3 | IKIGSHY | 1 |
| ERVGSHF | 3 | QRIGLHY | 1 |
| VKLGSHF | 2 | VSRTSHL | 1 |
| IVQGSHF | 2 | LPWQIHN | 1 |
| TXVGSHF | 1 | LPMTKHV | 1 |
| IHIGSHF | 1 | TNANHYF | 1 |
| VKQGTHF | 2 | QQTKNYY | 1 |
| TQIGTHF | 1 | SQLPWYS | 1 |
| IQIGTHF | 1 | MARYMSA | 2 |
| QSIGTHF | 1 | RGATPMS | 2 |
| ERVGTHF | 1 | NTGGSMA | 1 |
| IHXGTHF | 1 | NTGSPYE | 1 |
| QSINGHF | 2 | HSRVSGT | 1 |
| YSSPSHF | 1 | TNGDSAR | 1 |
| Characteristics | RECEPTORi | RNAi |
|---|---|---|
| Target site | GPCR drive | mRNA drive |
| Mode of action | Interfere receptor | Interfere mRNA |
| Potential target genes Feasible target genes Average molecular weight | ~400–600 genes ~20–30 genes ~1–8 amino acids (MW < 900) | ~20,000–40,000 genes ~200–400 genes ~21–500 nucleotides (MW > 13K) |
| Screening period for targets | Rapid | Slow |
| Delivery method | ||
| - Topical | Available (after formulation) | Limited due to molecule size |
| - transgenic (plant-incorporated) | Available | Available |
| - Ingestion (oral) | Available | Available |
| - Spraying plant | Available | Available |
| Bt alternative | Possible | Limited (synergistic) |
| Synthetic mimic (chemical) | Possible | Limited |
| Target insect | Unlimited | Unlimited |
| Biodegradable Resistance development | Possible (rapid) Slow | Possible Moderate |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, M.-Y.; Vander Meer, R.K. GPCR-Based Bioactive Peptide Screening Using Phage-Displayed Peptides and an Insect Cell System for Insecticide Discovery. Biomolecules 2021, 11, 583. https://doi.org/10.3390/biom11040583
Choi M-Y, Vander Meer RK. GPCR-Based Bioactive Peptide Screening Using Phage-Displayed Peptides and an Insect Cell System for Insecticide Discovery. Biomolecules. 2021; 11(4):583. https://doi.org/10.3390/biom11040583
Chicago/Turabian StyleChoi, Man-Yeon, and Robert K. Vander Meer. 2021. "GPCR-Based Bioactive Peptide Screening Using Phage-Displayed Peptides and an Insect Cell System for Insecticide Discovery" Biomolecules 11, no. 4: 583. https://doi.org/10.3390/biom11040583
APA StyleChoi, M.-Y., & Vander Meer, R. K. (2021). GPCR-Based Bioactive Peptide Screening Using Phage-Displayed Peptides and an Insect Cell System for Insecticide Discovery. Biomolecules, 11(4), 583. https://doi.org/10.3390/biom11040583