
Insights into the Activation of a Crustacean G Protein-Coupled Receptor: Evaluation of the Red Pigment-Concentrating Hormone Receptor of the Water Flea Daphnia pulex (Dappu-RPCH R)


Abstract
1. Introduction
2. Materials and Methods
2.1. Residue Scanning
2.2. Molecular Dynamics
3. Results and Discussion
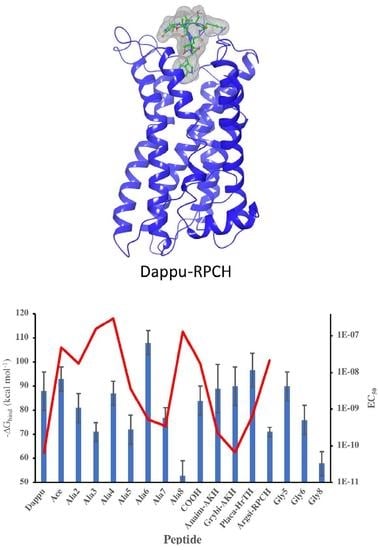
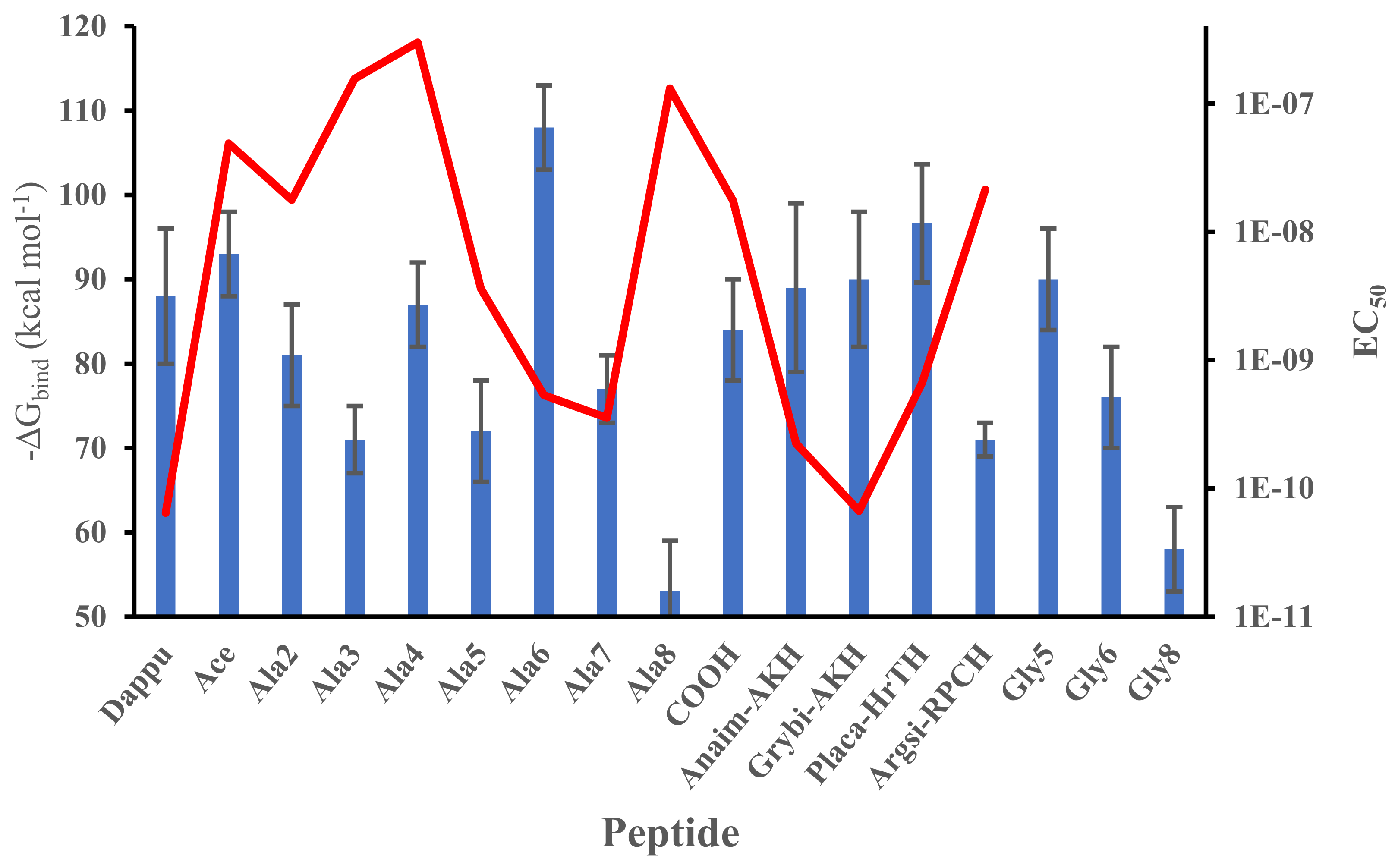
3.1. Residue Scanning
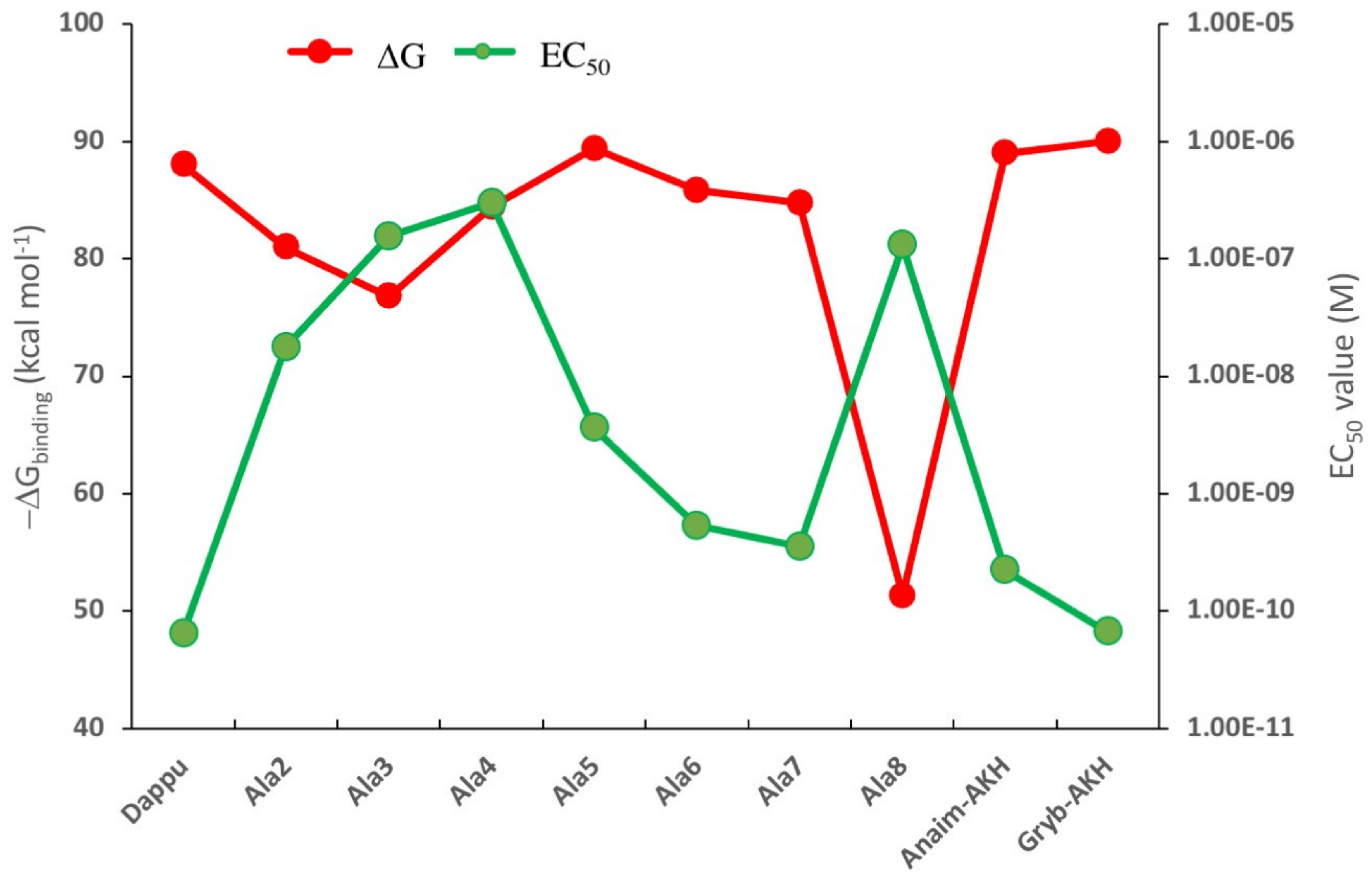
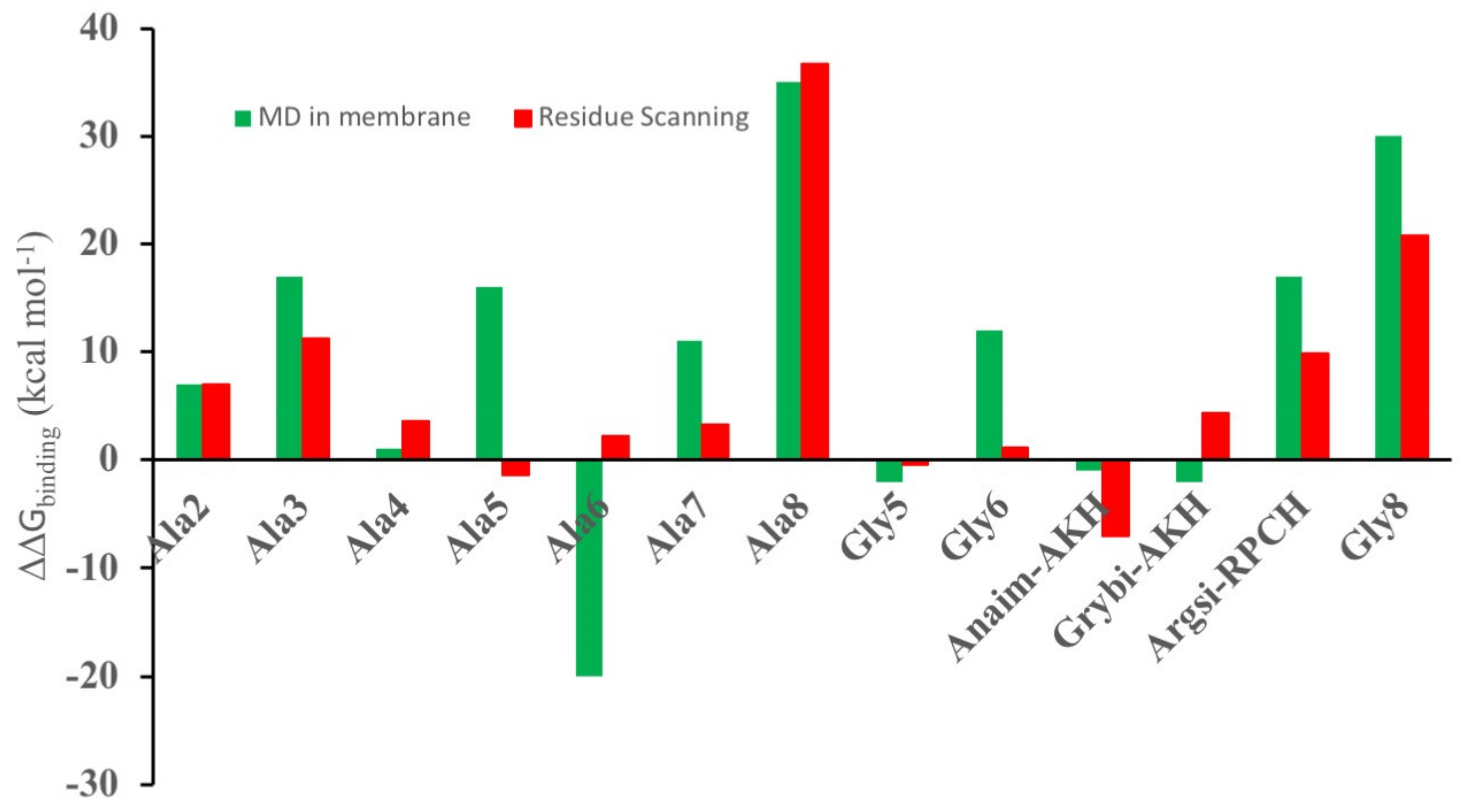
3.2. Comparison of Residue Scanning and Molecular Dynamics (MD)
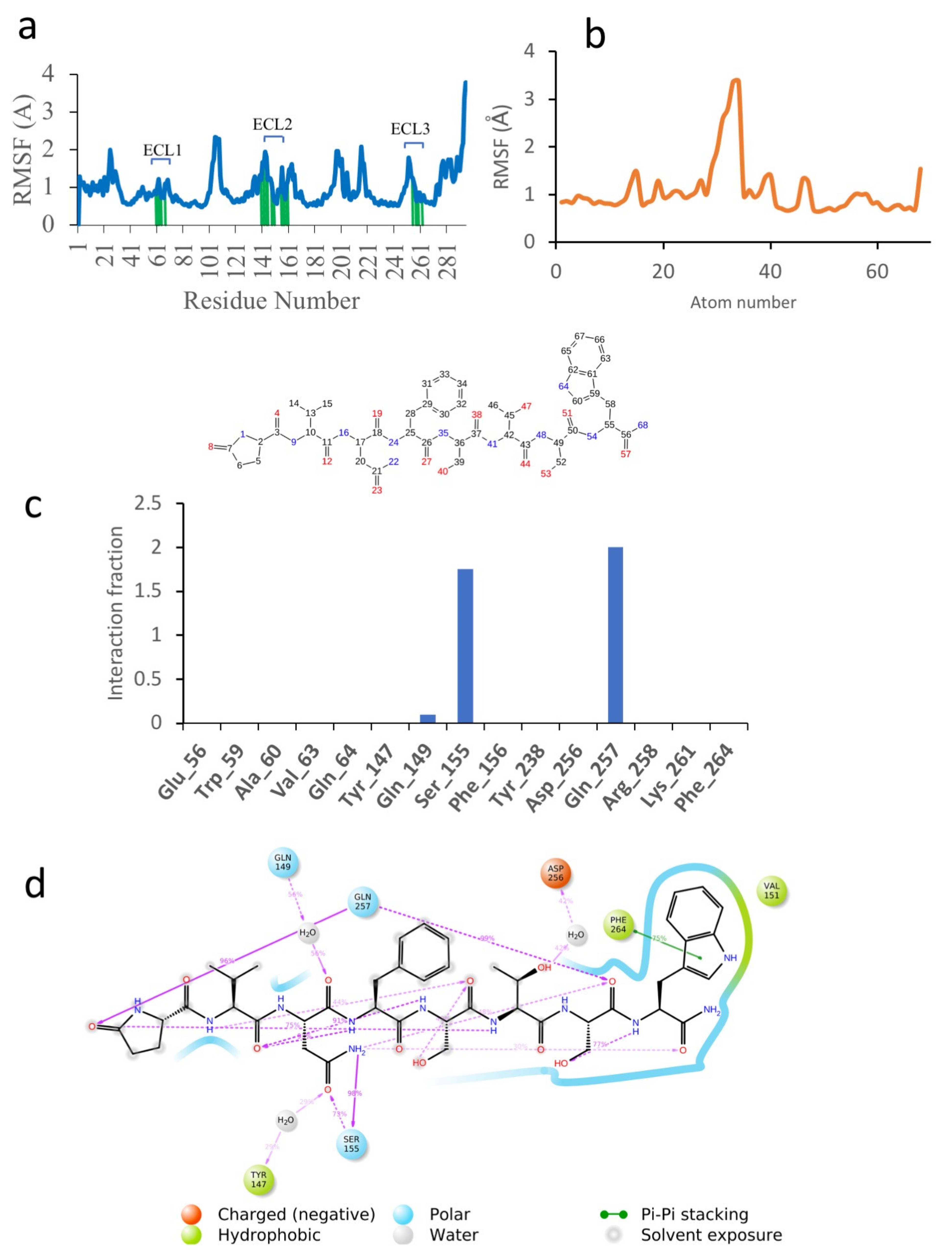
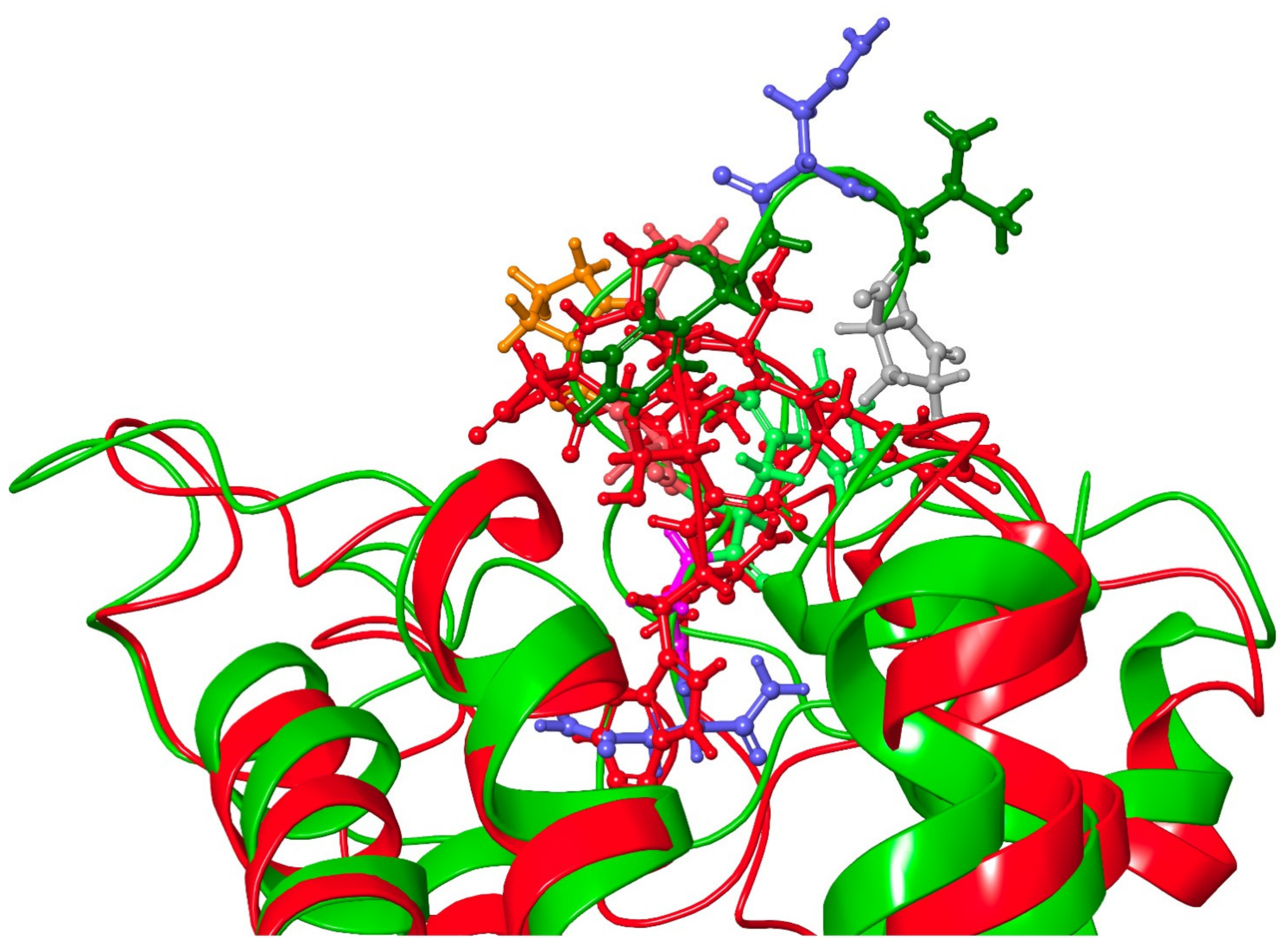
3.3. Molecular Dynamics
3.3.1. [Ala5]Dappu-RPCH
3.3.2. [Ala6]Dappu-RPCH
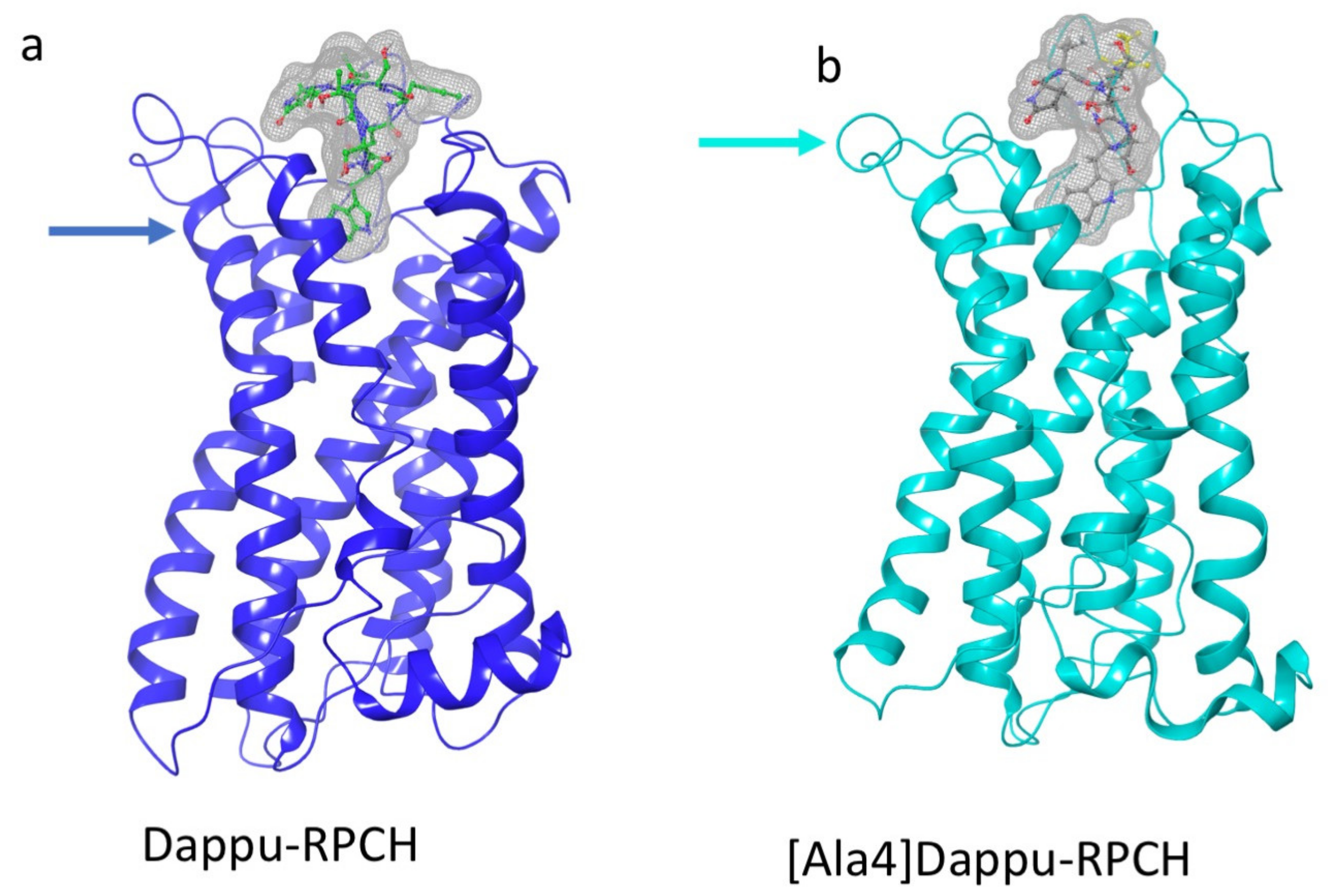
3.3.3. Dappu-RPCH
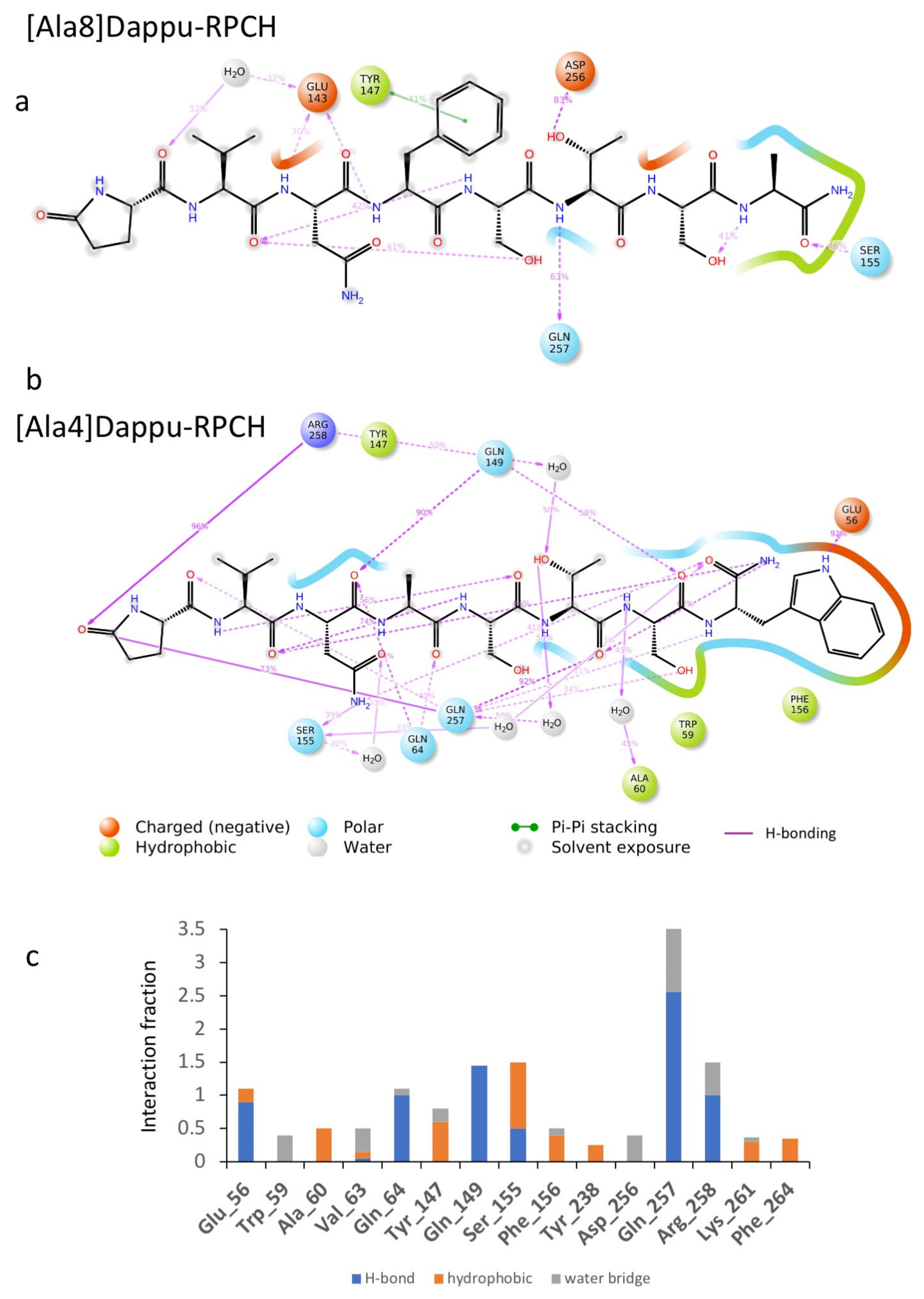
3.3.4. [Ace]Dappu-RPCH, [COOH]Dappu-RPCH and [Ala8]Dappu-RPCH
3.3.5. [Ala4]Dappu-RPCH
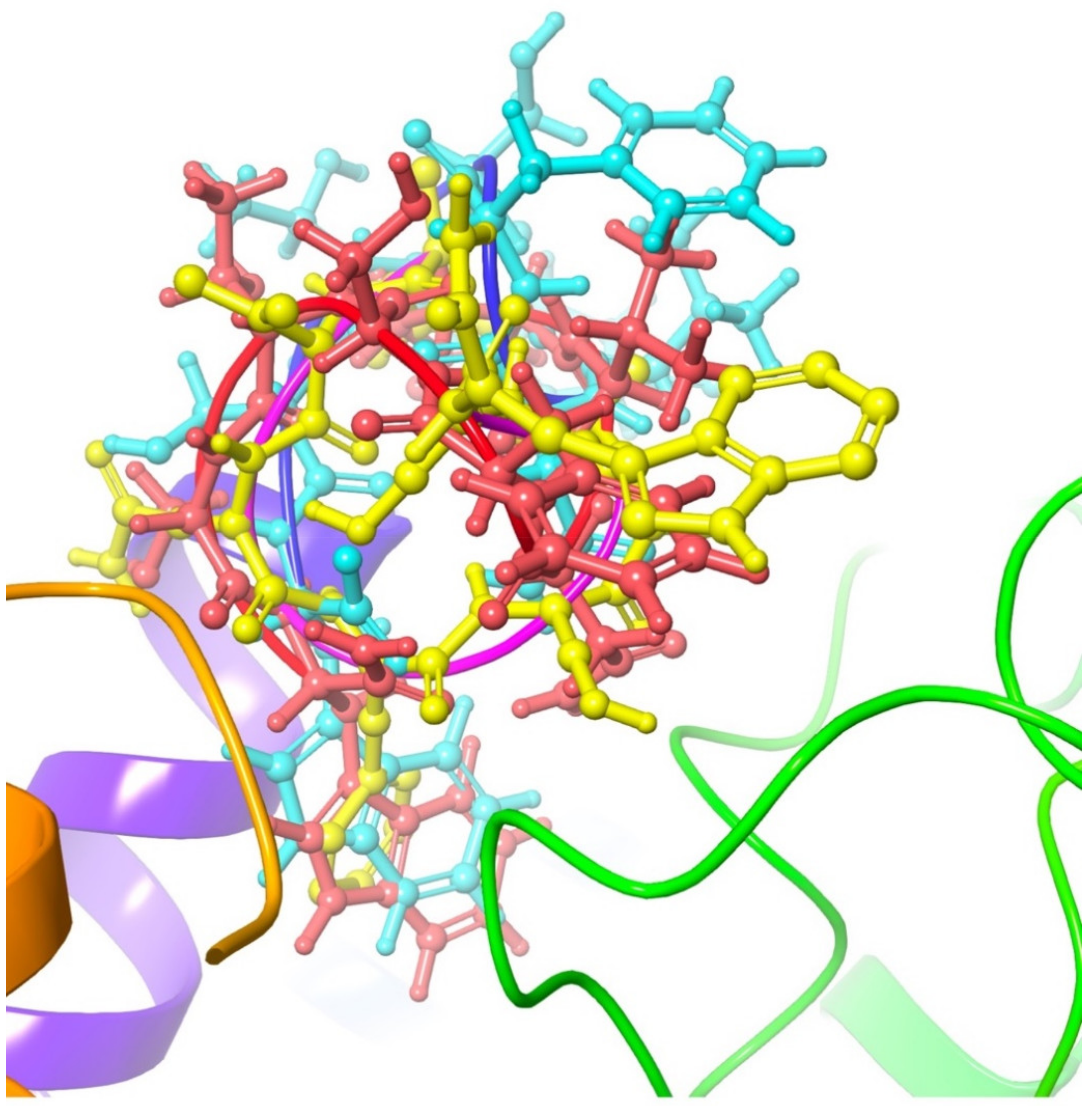
3.3.6. Placa-HrTH
3.3.7. Comparison with Gonadotropin Releasing Hormone GnRH
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Fernlund, P.; Josefsson, L. Crustacean color-change hormone: Amino acid sequence and chemical synthesis. Science 1972, 177, 173–175. [Google Scholar] [CrossRef]
- Gäde, G. The explosion of structural information on insect neuropeptides. In Progress in the Chemistry of Organic Natural Products; Herz, W., Kirby, G.W., Moore, R.E., Steglich, W., Tamm, C.H., Eds.; Springer: New York, NY, USA, 1997; Volume 71, pp. 1–128. [Google Scholar]
- Marco, H.G.; Gäde, G. Adipokinetic hormone: A hormone for all seasons. In Advances in Invertebrate (Neuro)Endocrinology: A Collection of Reviews in the Post- Genomic Era; Saleuddin, S., Lange, A.B., Orchard, I., Eds.; Apple Academic Press: Cambridge, MA, USA, 2020; Volume 2, pp. 126–170. [Google Scholar]
- Hansen, K.K.; Stafflinger, E.; Schneider, M.; Hauser, F.; Cazzamali, G.; Williamson, M.; Kollmann, M.; Schachtner, J.; Grimmelikhuijzen, C.J. Discovery of a novel insect neuropeptide signaling system closely related to the insect adipokinetic hormone and corazonin hormonal systems. J. Biol. Chem. 2010, 285, 10736–10747. [Google Scholar] [CrossRef]
- Roch, G.J.; Busby, E.R.; Sherwood, N.M. Evolution of GnRH: Diving deeper. Gen. Comp. Endocrinol. 2011, 171, 1–16. [Google Scholar] [CrossRef]
- Gäde, G.; Šimek, P.; Marco, H.G. An invertebrate [hydroxyproline]-modified neuropeptide: Further evidence for a close evolutionary relationship between insect adipokinetic hormone and mammalian gonadotropin hormone family. Biochem. Biophys. Res. Commun. 2011, 414, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Gäde, G.; Marco, H.G. The decapod red pigment-concentrating hormone (Panbo-RPCH) is the first identified neuropeptide of the order Plecoptera and is interpreted as homoplastic character state. Gen. Comp. Endocrinol. 2015, 221, 228–235. [Google Scholar] [CrossRef]
- Christie, A.E.; Cashman, C.R.; Brennan, H.R.; Ma, M.; Sousa, G.L.; Li, L.; Stemmler, E.A.; Dickinson, P.S. Identification of putative crustacean neuropeptides using in silico analyses of publicly accessible expressed sequence tags. Gen. Comp. Endocrinol. 2008, 156, 246–264. [Google Scholar] [CrossRef]
- Marco, H.G.; Gäde, G. Biological activity of the predicted red pigment-concentrating hormone of Daphnia pulex in a crustacean and an insect. Gen. Comp. Endocrinol. 2010, 166, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Christie, A.E.; McCoole, M.D.; Harmon, S.M.; Baer, K.N.; Lenz, P.H. Genomic analyses of the Daphnia pulex peptidome. Gen. Comp. Endocrinol. 2011, 171, 131–150. [Google Scholar] [CrossRef] [PubMed]
- Dircksen, H.; Neupert, S.; Predel, R.; Verleyen, P.; Huybrechts, J.; Strauss, J.; Hauser, F.; Stafflinge, E.; Schneider, M.; Grimmelikhuijzen, C.J.; et al. Genomics transcriptomics, and peptidomics of Daphnia pulex neuropeptides and protein hormones. J. Proteome Res. 2011, 10, 4478–4504. [Google Scholar] [CrossRef] [PubMed]
- Christie, A.E. Peptide discovery in the ectoparasitic crustacean Argulus siamensis: Identification of the first neuropeptides from a member of the Branchiura. Gen. Comp. Endocrinol. 2014, 204, 114–125. [Google Scholar] [CrossRef]
- Marco, H.G.; Verlinden, H.; VandenBroeck, J.; Gäde, G. Characterisation and pharmacological analysis of a crustacean G protein-coupled receptor: The red pigment-concentrating hormone receptor of Daphnia pulex. Sci. Rep. 2017, 7, 6851. [Google Scholar] [CrossRef]
- Soetaert, A.; Vandenbrouck, T.; van der Ven, K.; Maras, M.; van Remortel, P.; Blust, R.; De Coen, W.M. Molecular responses during cadmium-induced stress in Daphnia magna: Integration of differential gene expression with higher-level effects. Aquat. Toxicol. 2007, 83, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Baun, A.; Hartmann, N.B.; Grieger, K.; Kusk, K.O. Ecotoxicity of engineered nanoparticles to aquatic invertebrates: A brief review and recommendations for future toxicity testing. Ecotoxicology 2008, 17, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Christie, A.E.; McCoole, M.D. From genes to behavior: Investigations of neurochemical signaling come of age for the model crustacean Daphnia pulex. J. Exp. Biol. 2012, 215, 2535–2544. [Google Scholar] [CrossRef]
- Munoz, J.; Chaturvedi, A.; De Meester, L.; Weider, L.J. Characterization of genome-wide SNPs for the water flea Daphnia pulicaria generated by genotyping-by-sequencing (GBS). Sci. Rep. 2016, 6, 28569. [Google Scholar] [CrossRef]
- Audsley, N.; Down, R.E. G protein coupled receptors as targets for next generation pesticides. Insect Biochem. Mol. Biol. 2015, 67, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Verlinden, H.; Vleugels, R.; Zels, S.; Dillen, S.; Lenaerts, C.; Crabbé, K.; Spit, J.; Broeck, J.V. Receptors for neuronal or endocrine signalling molecules as potential targets for the control of insect pests. Adv. Insect Physiol. 2014, 46, 167–303. [Google Scholar]
- Jackson, G.E.; Pavadai, E.; Gäde, G.; Timol, Z.; Andersen, N.H. Data for the homology modelling of the red pigment-concentrating hormone receptor (Dappu-RPCHR) of the crustacean Daphnia pulex, and docking of its cognate agonist (Dappu-RPCH). Data Brief 2017, 15, 941–947. [Google Scholar] [CrossRef]
- Jackson, G.E.; Pavadai, E.; Gäde, G.; Timol, Z.; Andersen, N.H. Interaction of the red pigment-concentrating hormone of the crustacean Daphnia pulex, with its cognate receptor, Dappu-RPCHR: A nuclear magnetic resonance and modeling study. Int. J. Biol. Macromol. 2018, 106, 969–978. [Google Scholar] [CrossRef]
- Cherezov, V.; Rosenbaum, D.M.; Hanson, M.A.; Rasmussen, S.G.F.; Foon, S.T.; Kobilka, T.S.; Choi, H.J.; Kuhn, P.; Weis, W.I.; Kobilka, B.K.; et al. High-resolution crystal structure of an engineered humanß2-adrenergic G protein-coupled receptor. Science 2007, 318, 1258–1265. [Google Scholar] [CrossRef] [PubMed]
- Schrödinger Release 2021-1, Desmond Molecular Dynamics System, D.E. Shaw Research: New York, NY, USA, 2021. Maestro-Desmond Interoperability Tools, Schrödinger, New York, NY, USA. 2021. Available online: https://www.schrodinger.com/ (accessed on 12 March 2021).
- Sligar, S.G. Nanodiscs for Structural and Functional Studies of Membrane Proteins. Nat. Struct. Mol. Biol. 2016, 23, 481–486. [Google Scholar]
- Schrödinger Release 2021-1; Maestro, Schrödinger, LLC: New York, NY, USA, 2021.
- Genheden, S.; Ryde, U. The MM/PBSA and MM/GBSA methods to estimate ligand-binding affinities. Expert Opin. Drug Discov. 2015, 10, 449–461. [Google Scholar] [CrossRef]
- Gäde, G.; Janssens, M.P.E. Cicadas contain novel members of the AKH/RPCH family peptides with hypertrehalosaemic activity. Biol. Chem. Hoppe Seyler 1994, 375, 803–809. [Google Scholar] [PubMed]
- Flanagan, C.; Manilall, A. Gonadotropin-Releasing Hormone (GnRH) Receptor Structure and GnRH Binding. Front. Endocrinol. 2017, 8, 274. [Google Scholar]
- Ballesteros, J.A.; Weinstein, W. Integrated methods for the construction of three-dimensional models and computational probing of structure-function relations in G protein-coupled receptors. Methods Neurosci. 1995, 25, 366–428. [Google Scholar] [CrossRef]
- Cvicek, V.; Goddard, W.A., III; Abrol, R. Structure-based sequence alignment of the transmembrane domains of all human GPCRs: Phylogenetic, structural and functional implications. PLoS Comput. Biol. 2016, 12, e1004805. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide Name | Peptide Sequence | EC50 Value (M) | ||
|---|---|---|---|---|
| Dappu-RPCH | pQVNFSTSWamide | 6.45E-11 | ||
| [Ala2]Dappu-RPCH | pQANFSTSWamide | 1.77E-08 | ||
| [Ala3]Dappu-RPCH | pQVAFSTSWamide | 1.56E-07 | ||
| [Ala4]Dappu-RPCH | pQVNASTSWamide | 2.99E-07 | ||
| [Ala5]Dappu-RPCH | pQVNFATSWamide | 3.61E-09 | ||
| [Ala6]Dappu-RPCH | pQVNFSASWamide | 5.32E-10 | ||
| [Ala7]Dappu-RPCH | pQVNFSTAWamide | 3.55E-10 | ||
| [Ala8]Dappu-RPCH | pQVNFSTSAamide | 1.31E-07 | ||
| [Ace]Dappu-RPCH | [NAc-Ala]-VNFSTSWamide | 4.87E-08 | ||
| [COOH]Dappu-RPCH | pQVNFSTSW-OH | 1.74E-08 | ||
| Placa-HrTH | pQVNFSPSWGNamide | 6.64E-10 | ||
| [Pro6]Dappu-RPCH= Anaim-AKH | pQVNFSPSWamide | 2.24E-10 | ||
| [Gly7]Dappu-RPCH= Grybi-AKH | pQVNFSTGWamide | 6.67E-11 | ||
| [Lys7]Dappu-RPCH=Argsi-RPCH | pQVNFSTKWamide | 2.13E-8 | ||
| [Gly8]Dappu-RPCH | pQVNFSTSGamide | - | ||
| [Gly5]Dappu-RPCH | pQVNFGTSWamide | - | ||
| [Gly6]Dappu-RPCH | pQVNFSGSWamide | - | ||
| Peptide | Total | Coulombic | Hbond | Lipophilic | Solvation (GB) | Solvation (SA) | vdW |
|---|---|---|---|---|---|---|---|
| Dappu | −88(7) | −11(3) | −1.1(0.9) | −28(1) | 26(3) | −74(3) | 29(2) |
| COOH | −85(4) | −52(6) | −1.9(0.7) | −25(2) | 62(4) | −68(4) | 20(3) |
| ACE | −94(5) | −29(8) | −2.7(0.3) | −27(3) | 35(2) | −71(3) | 10(3) |
| Ala2 | −81(4) | −46(4) | −2.6(0.2) | −24(2) | 62(2) | −73(4) | 10(2) |
| Ala3 | −71(5) | −35(3) | −1.1(0.1) | −24(2) | 44(3) | −60(3) | 14(4) |
| Ala4 | −87(6) | −44(4) | −3.8(0.8) | −20(3) | 47(4) | −60(2) | 12(3) |
| Ala5 | −70(5) | −17(4) | −0.9(0.6) | −22(4) | 40(1) | −66(3) | 31(2) |
| Ala6 | −108(7) | −35(2) | −2.9(0.3) | −24(3) | 42(3) | −80(4) | 20(3) |
| Ala7 | −77(4) | −21(3) | −1.4(0.5) | −24(4) | 35(2) | −64(4) | 26(2) |
| Ala8 | −54(5) | −23(2) | −3.1(0.6) | −15(3) | 35(2) | −46(2) | 13(3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jackson, G.E.; Gäde, G. Insights into the Activation of a Crustacean G Protein-Coupled Receptor: Evaluation of the Red Pigment-Concentrating Hormone Receptor of the Water Flea Daphnia pulex (Dappu-RPCH R). Biomolecules 2021, 11, 710. https://doi.org/10.3390/biom11050710
Jackson GE, Gäde G. Insights into the Activation of a Crustacean G Protein-Coupled Receptor: Evaluation of the Red Pigment-Concentrating Hormone Receptor of the Water Flea Daphnia pulex (Dappu-RPCH R). Biomolecules. 2021; 11(5):710. https://doi.org/10.3390/biom11050710
Chicago/Turabian StyleJackson, Graham E., and Gerd Gäde. 2021. "Insights into the Activation of a Crustacean G Protein-Coupled Receptor: Evaluation of the Red Pigment-Concentrating Hormone Receptor of the Water Flea Daphnia pulex (Dappu-RPCH R)" Biomolecules 11, no. 5: 710. https://doi.org/10.3390/biom11050710
APA StyleJackson, G. E., & Gäde, G. (2021). Insights into the Activation of a Crustacean G Protein-Coupled Receptor: Evaluation of the Red Pigment-Concentrating Hormone Receptor of the Water Flea Daphnia pulex (Dappu-RPCH R). Biomolecules, 11(5), 710. https://doi.org/10.3390/biom11050710