
The Role of Nuclear Insulin and IGF1 Receptors in Metabolism and Cancer




Abstract
1. Overview of the Insulin-Like Growth Factor System
2. Classical Concepts on Hormonal Signaling and Internalization
3. Historical Aspects of Nuclear InsR/IGF1R Translocation
4. Proteomic Profiling Identified the IGF1R in Nuclear Fractions of Breast Cancer Cells
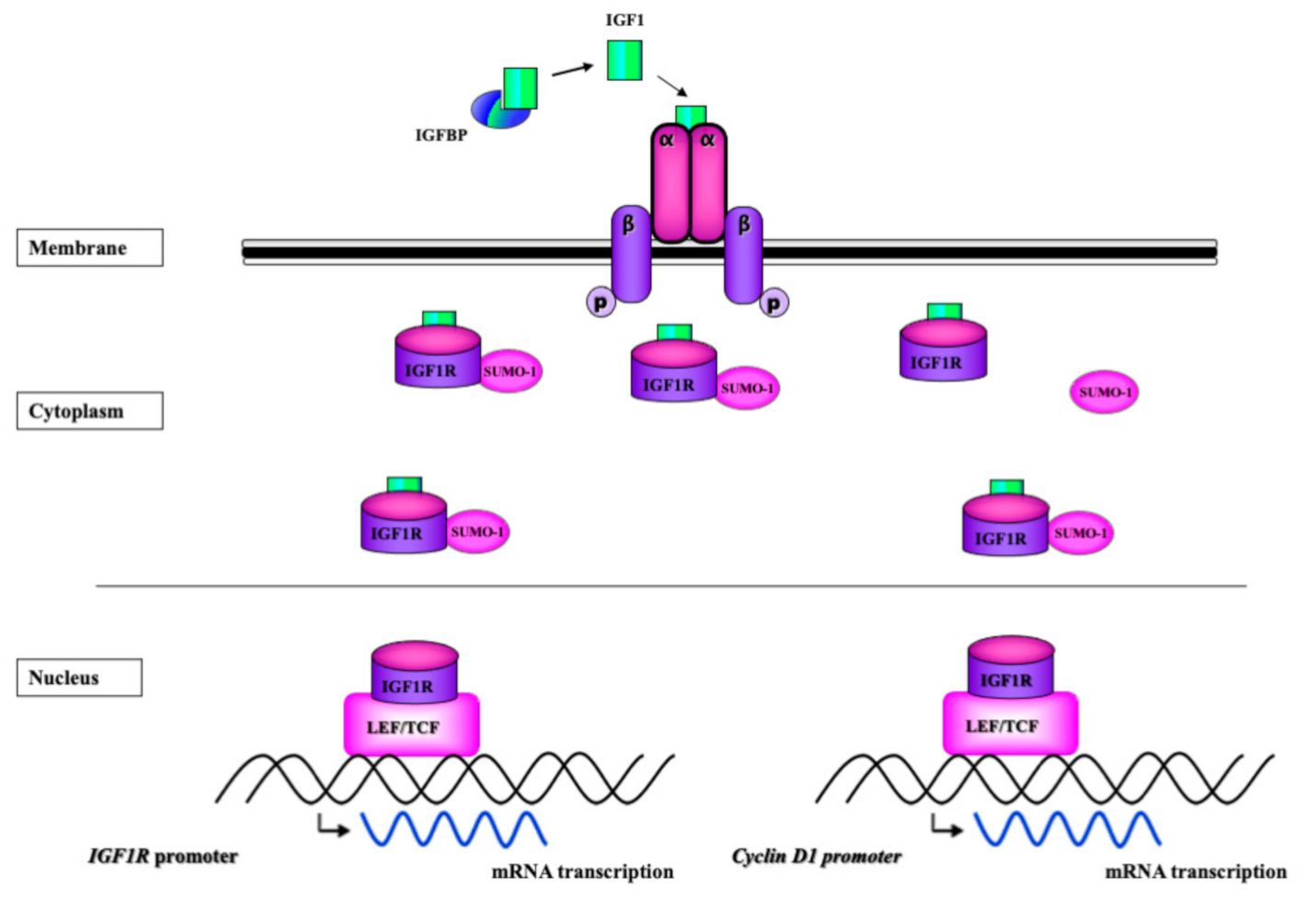
5. Potential Mechanisms of InsR/IGF1R Nuclear Translocation
6. Transcriptional Activity of InsR and IGF1R
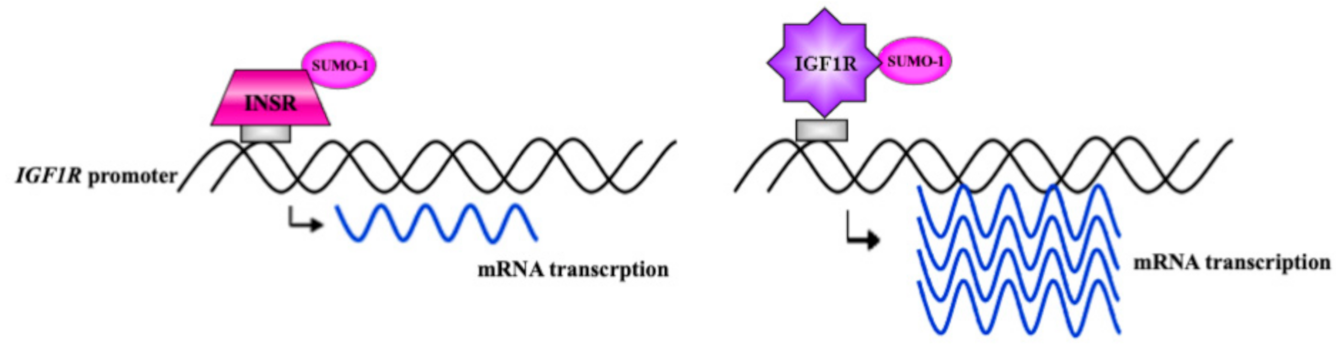
7. Autoregulation of IGF1R Gene Expression by IGF1R
8. Autoregulation of InsR Gene Expression by InsR
9. Is Nuclear InsR/IGF1R Translocation Limited to Malignant Cells?
10. Nuclear Localization of Hybrid InsR/IGF1R
11. IGF1R Localization in the Golgi Apparatus
12. Nuclear Localization Is a Common Theme among Tyrosine Kinase Receptors
13. The Role of Nuclear InsR/IGF1R in Cancer
14. The Role of Nuclear InsR/IGF1R in Metabolism
15. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yakar, S.; Adamo, M.L. Insulin-like growth factor 1 physiology: Lessons from mouse models. Endocrinol. Metab. Clin. N. Am. 2012, 41, 231–247. [Google Scholar] [CrossRef] [PubMed]
- Yakar, S.; Werner, H.; Rosen, C.J. 40 YEARS OF IGF1: Insulin-like growth factors: Actions on the skeleton. J. Mol. Endocrinol. 2018, 61, T115–T137. [Google Scholar] [CrossRef] [PubMed]
- Longo, V.; Finch, C. Evolutionary medicine: From dwarf model systems to healthy centenarians? Science 2003, 299, 1342–1346. [Google Scholar] [CrossRef] [PubMed]
- Bentov, I.; Werner, H. Insulin-like growth factors and breast cancer. Current Med. Lit. Breast Cancer 2009, 21, 113–120. [Google Scholar]
- Salmon, W.D.; Daughaday, W.H. A hormonally controlled serum factor which stimulates sulfate incorporation by cartilage in vitro. J. Lab. Clin. Med. 1957, 49, 825–836. [Google Scholar]
- Rosenfeld, R.G. Insulin-like Growth Factors and the Basis of Growth. N. Engl. J. Med. 2003, 349, 2184–2186. [Google Scholar] [CrossRef]
- Moschos, S.J.; Mantzoros, C.S. The Role of the IGF System in Cancer: From Basic to Clinical Studies and Clinical Applications. Oncology 2002, 63, 317–332. [Google Scholar] [CrossRef]
- Leroith, D. Clinical relevance of systemic and local IGF-I: Lessons from animal models. Pediatr. Endocrinol. Rev. 2008, 5, 739–743. [Google Scholar]
- Sarfstein, R.; Maor, S.; Reizner, N.; Abramovitch, S.; Werner, H. Transcriptional regulation of the insulin-like growth factor-I receptor gene in breast cancer. Mol. Cell. Endocrinol. 2006, 252, 241–246. [Google Scholar] [CrossRef]
- De Meyts, P.; Whittaker, J. Structural biology of insulin and IGF1 receptors: Implications for drug design. Nat. Rev. Drug Discov. 2002, 1, 769–783. [Google Scholar] [CrossRef]
- Werner, H.; Weinstein, D.; Bentov, I. Similarities and differences between insulin and IGF-I: Structures, receptors, and signalling pathways. Arch. Physiol. Biochem. 2008, 114, 17–22. [Google Scholar] [CrossRef]
- Werner, H.; Sarfstein, R. Transcriptional and epigenetic control of IGF1R gene expression: Implications in metabolism and cancer. Growth Horm. IGF Res. 2014, 24, 112–118. [Google Scholar] [CrossRef]
- Baker, J.; Liu, J.-P.; Robertson, E.J.; Efstratiadis, A. Role of insulin-like growth factors in embryonic and postnatal growth. Cell 1993, 75, 73–82. [Google Scholar] [CrossRef]
- Bach, L. Insulin-like growth factor binding proteins—An update. Pediatr. Endocrinol. Rev. 2015, 13, 521–530. [Google Scholar]
- Holly, J.; Perks, C. The Role of Insulin-Like Growth Factor Binding Proteins. Neuroendocrinology 2006, 83, 154–160. [Google Scholar] [CrossRef]
- Domene, H.; Bengolea, S.V.; Martinez, A.S.; Ropelato, M.G.; Pennisi, P.; Scaglia, P.; Heinrich, J.; Jasper, H. Deficiency of the circulating IGF system associated with inactivation of the acid-labile subunit gene. N. Engl. J. Med. 2004, 350, 570–577. [Google Scholar] [CrossRef]
- Baxter, R.C. IGF binding proteins in cancer: Mechanistic and clinical insights. Nat. Rev. Cancer 2014, 14, 329–341. [Google Scholar] [CrossRef]
- Cai, Q.; Dozmorov, M.; Oh, Y. IGFBP-3/IGFBP-3 Receptor System as an Anti-Tumor and Anti-Metastatic Signaling in Cancer. Cells 2020, 9, 1261. [Google Scholar] [CrossRef]
- Ravid, D.; Maor, S.; Werner, H.; Liscovitch, M. Caveolin-1 inhibits anoikis and promotes survival signaling in cancer cells. Adv. Enzym. Regul. 2006, 46, 163–175. [Google Scholar] [CrossRef]
- Traub, L.M.; Bonifacino, J.S. Cargo Recognition in Clathrin-Mediated Endocytosis. Cold Spring Harb. Perspect. Biol. 2013, 5, a016790. [Google Scholar] [CrossRef]
- Janssen, J.A. New Insights from IGF-IR Stimulating Activity Analyses: Pathological Considerations. Cells 2020, 9, 862. [Google Scholar] [CrossRef] [PubMed]
- Irannejad, R.; Tsvetanova, N.G.; Lobingier, B.T.; von Zastrow, M. Effects of endocytosis on receptor-mediated signaling. Curr. Opin. Cell Biol. 2015, 35, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Goldfine, I.D.; Smith, G.J. Binding of insulin to isolated nuclei. Proc. Natl. Acad. Sci. USA 1976, 73, 1427–1431. [Google Scholar] [CrossRef] [PubMed]
- Goldfine, I.D.; Vigneri, R.; Cohen, D.; Pliam, N.B.; Kahn, C.R. Intracellular binding sites for insulin are immunologically distinct from those on the plasma membrane. Nat. Cell Biol. 1977, 269, 698–700. [Google Scholar] [CrossRef] [PubMed]
- Vigneri, R.; Goldfine, I.; Wong, K.; Smith, G.; Pezzino, V. The nuclear envelope. The major site of insulin binding in rat liver nuclei. J. Biol. Chem. 1978, 253, 2098–2103. [Google Scholar] [CrossRef]
- Horvath, A. Insulin binding sites on rat liver nuclear membranes: Biochemical and immunofluorescent studies. J. Cell Physiol. 1978, 97, 37–47. [Google Scholar] [CrossRef]
- Bergeron, J.; Posner, B.; Josefsberg, Z.; Sikstrom, R. Intracellular polypeptide hormone receptors. The demonstration of specific binding sites for insulin and human growth hormone in Golgi fractions isolated from the liver of female rats. J. Biol. Chem. 1978, 253, 4058–4066. [Google Scholar] [CrossRef]
- Podlecki, D.A.; Smith, R.M.; Kao, M.; Tsai, P.; Huecksteadt, T.; Brandenburg, D.; Lasher, R.S.; Jarett, L.; Olefsky, J.M. Nuclear translocation of the insulin receptor. A possible mediator of insulin’s long term effects. J. Biol. Chem. 1987, 262, 3362–3368. [Google Scholar] [CrossRef]
- Chen, C.-W.; Oberley, T.D.; Roy, D. Inhibition of stilbene estrogen-induced cell proliferation of renal epithelial cells through the modulation of insulin-like growth factor-I receptor expression. Cancer Lett. 1996, 105, 51–59. [Google Scholar] [CrossRef]
- Sun, H.; Tu, X.; Prisco, M.; Wu, A.; Casiburi, I.; Baserga, R. Insulin-Like Growth Factor I Receptor Signaling and Nuclear Translocation of Insulin Receptor Substrates 1 and 2. Mol. Endocrinol. 2003, 17, 472–486. [Google Scholar] [CrossRef]
- Baserga, R. The decline and fall of the IGF-I receptor. J. Cell. Physiol. 2012, 228, 675–679. [Google Scholar] [CrossRef]
- Sarfstein, R.; Belfiore, A.; Werner, H. Identification of Insulin-Like Growth Factor-I Receptor (IGF-IR) Gene Promoter-Binding Proteins in Estrogen Receptor (ER)-Positive and ER-Depleted Breast Cancer Cells. Cancers 2010, 2, 233–261. [Google Scholar] [CrossRef]
- Sarfstein, R.; Pasmanik-Chor, M.; Yeheskel, A.; Edry, L.; Shomron, N.; Warman, N.; Wertheimer, E.; Maor, S.; Shochat, L.; Werner, H. Insulin-like Growth Factor-I Receptor (IGF-IR) Translocates to Nucleus and Autoregulates IGF-IR Gene Expression in Breast Cancer Cells. J. Biol. Chem. 2012, 287, 2766–2776. [Google Scholar] [CrossRef]
- Maor, S.; Mayer, D.; Yarden, I.R.; Lee, A.V.; Sarfstein, R.; Werner, H.; Papa, M.Z. Estrogen receptor regulates insulin-like growth factor-I receptor gene expression in breast tumor cells: Involvement of transcription factor Sp1. J. Endocrinol. 2006, 191, 605–612. [Google Scholar] [CrossRef]
- Vecchione, A.; Marchese, A.; Henry, P.; Rotin, D.; Morrione, A. The Grb10/Nedd4 Complex Regulates Ligand-Induced Ubiquitination and Stability of the Insulin-Like Growth Factor I Receptor. Mol. Cell. Biol. 2003, 23, 3363–3372. [Google Scholar] [CrossRef]
- Aleksic, T.; Chitnis, M.M.; Perestenko, O.V.; Gao, S.; Thomas, P.H.; Turner, G.D.; Protheroe, A.S.; Howarth, M.; Macaulay, V.M. Type 1 Insulin-like Growth Factor Receptor Translocates to the Nucleus of Human Tumor Cells. Cancer Res. 2010, 70, 6412–6419. [Google Scholar] [CrossRef]
- Warsito, D.; Sjöström, S.; Andersson, S.; Larsson, O.; Sehat, B. Nuclear IGF1R is a transcriptional co-activator of LEF1/TCF. EMBO Rep. 2012, 13, 244–250. [Google Scholar] [CrossRef]
- Packham, S.; Warsito, D.; Lin, Y.; Sadi, S.; Karlsson, R.; Sehat, B.; Larsson, O. Nuclear translocation of IGF-1R via p150(Glued) and an importin-β/RanBP2-dependent pathway in cancer cell. Oncogene 2015, 34, 2227–2238. [Google Scholar] [CrossRef]
- Sehat, B.; Tofigh, A.; Lin, Y.; Trocmé, E.; Liljedahl, U.; Lagergren, J.; Larsson, O. SUMOylation Mediates the Nuclear Translocation and Signaling of the IGF-1 Receptor. Sci. Signal. 2010, 3, ra10. [Google Scholar] [CrossRef]
- Sarfstein, R.; Werner, H. Minireview: Nuclear Insulin and Insulin-like Growth Factor-1 Receptors: A Novel Paradigm in Signal Transduction. Endocrinology 2013, 154, 1672–1679. [Google Scholar] [CrossRef]
- Lin, Y.; Liu, H.; Waraky, A.; Haglund, F.; Agarwal, P.; Jernberg-Wiklund, H.; Warsito, D.; Larsson, O. SUMO-modified insulin-like growth factor 1 receptor (IGF-1R) increases cell cycle progression and cell proliferation. J. Cell. Physiol. 2017, 232, 2722–2730. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Lin, Y.; Badin, M.; Vasilcanu, D.; Strömberg, T.; Jernberg-Wiklund, H.; Sehat, B.; Larsson, O. Over-accumulation of nuclear IGF-1 receptor in tumor cells requires elevated expression of the receptor and the SUMO-conjugating enzyme Ubc9. Biochem. Biophys. Res. Commun. 2011, 404, 667–671. [Google Scholar] [CrossRef] [PubMed]
- Solomon-Zemler, R.; Sarfstein, R.; Werner, H. Nuclear insulin-like growth factor-1 receptor (IGF1R) displays proliferative and regulatory activities in non-malignant cells. PLoS ONE 2017, 12, e0185164. [Google Scholar] [CrossRef] [PubMed]
- Jamwal, G.; Singh, G.; Dar, M.S.; Singh, P.; Bano, N.; Syed, S.H.; Sandhu, P.; Akhter, Y.; Monga, S.P. Identification of a unique loss-of-function mutation in IGF1R and a crosstalk between IGF1R and Wnt/β-catenin signaling pathways. Biochim. Biophys. ACTA (BBA) Bioenerg. 2018, 1865, 920–931. [Google Scholar] [CrossRef]
- Warsito, D.; Lin, Y.; Gnirck, A.-C.; Sehat, B.; Larsson, O. Nuclearly translocated insulin-like growth factor 1 receptor phosphorylates histone H3 at tyrosine 41 and induces SNAI2 expression via Brg1 chromatin remodeling protein. Oncotarget 2016, 7, 42288–42302. [Google Scholar] [CrossRef]
- Sarfstein, R.; Werner, H. Tumor suppressor p53 regulates insulin receptor (INSR) gene expression via direct binding to the INSR promoter. Oncotarget 2020, 11, 2424–2437. [Google Scholar] [CrossRef]
- Solomon-Zemler, R.; Pozniak, Y.; Geiger, T.; Werner, H. Identification of nucleolar protein NOM1 as a novel nuclear IGF1R-interacting protein. Mol. Genet. Metab. 2019, 126, 259–265. [Google Scholar] [CrossRef]
- Belfiore, A.; Frasca, F.; Pandini, G.; Sciacca, L.; Vigneri, R. Insulin Receptor Isoforms and Insulin Receptor/Insulin-Like Growth Factor Receptor Hybrids in Physiology and Disease. Endocr. Rev. 2009, 30, 586–623. [Google Scholar] [CrossRef]
- Soos, A.M.; Whittaker, J.; Lammers, R.; Ullrich, A.; Siddle, K. Receptors for insulin and insulin-like growth factor-I can form hybrid dimers. Characterisation of hybrid receptors in transfected cells. Biochem. J. 1990, 270, 383–390. [Google Scholar] [CrossRef]
- Bailyes, E.M.; Navé, B.T.; Soos, M.A.; Orr, S.R.; Hayward, A.C.; Siddle, K. Insulin receptor/IGF-I receptor hybrids are widely distributed in mammalian tissues: Quantification of individual receptor species by selective immunoprecipitation and immunoblotting. Biochem. J. 1997, 327, 209–215. [Google Scholar] [CrossRef]
- Pandini, G.; Frasca, F.; Mineo, R.; Sciacca, L.; Vigneri, R.; Belfiore, A. Insulin/Insulin-like Growth Factor I Hybrid Receptors Have Different Biological Characteristics Depending on the Insulin Receptor Isoform Involved. J. Biol. Chem. 2002, 277, 39684–39695. [Google Scholar] [CrossRef]
- Wu, Y.-C.; Zhu, M.; Robertson, D.M. Novel nuclear localization and potential function of insulin-like growth factor-1 receptor/insulin receptor bybrid in corneal epithelial cells. PLoS ONE 2012, 7, e42483. [Google Scholar]
- Titone, R.; Zhu, M.; Robertson, D.M. Insulin mediates de novo nuclear accumulation of the IGF-1/insulin Hybrid Receptor in corneal epithelial cells. Sci. Rep. 2018, 8, 4378. [Google Scholar] [CrossRef]
- Rieger, L.; O’Shea, S.; Godsmark, G.; Stanicka, J.; Kelly, G.; O’Connor, R. IGF-1 receptor activity in the Golgi of migratory cancer cells depends on adhesion-dependent phosphorylation of Tyr1250 and Tyr1251. Sci. Signal. 2020, 13, eaba3176. [Google Scholar] [CrossRef]
- Yu, L.; Wang, H.; Guo, Z.; Li, F.; Cui, H. Role of nucleolar protein NOM1 in pancreatic islet β cell apoptosis in diabetes. Exp. Ther. Med. 2016, 12, 2275–2280. [Google Scholar] [CrossRef][Green Version]
- Alexandrov, A.; Colognori, D.; Steitz, J.A. Human eIF4AIII interacts with an eIF4G-like partner, NOM1, revealing an evolutionarily conserved function outside the exon junction complex. Genes Dev. 2011, 25, 1078–1090. [Google Scholar] [CrossRef]
- Gunawardena, S.R.; Ruis, B.L.; Meyer, J.A.; Kapoor, M.; Conklin, K.F. NOM1 Targets Protein Phosphatase I to the Nucleolus. J. Biol. Chem. 2008, 283, 398–404. [Google Scholar] [CrossRef]
- Wang, Y.-N.; Yamaguchi, H.; Hsu, J.-M.; Hung, M.-C. Nuclear trafficking of the epidermal growth factor receptor family membrane proteins. Oncogene 2010, 29, 3997–4006. [Google Scholar] [CrossRef]
- Lin, S.-Y.; Makino, K.; Xia, W.; Matin, A.; Wen, Y.; Kwong, K.Y.; Bourguignon, L.; Hung, M.-C. Nuclear localization of EGF receptor and its potential new role as a transcription factor. Nat. Cell Biol. 2001, 3, 802–808. [Google Scholar] [CrossRef]
- Dittmann, K.; Mayer, C.; Czemmel, S.; Huber, S.M.; Rodemann, H.P. New roles for nuclear EGFR in regulating the stability and translation of mRNAs associated with VEGF signaling. PLoS ONE 2017, 12, e0189087. [Google Scholar] [CrossRef]
- Hsu, S.-C.; Miller, S.A.; Wang, Y.; Hung, M.-C. Nuclear EGFR is required for cisplatin resistance and DNA repair. Am. J. Transl. Res. 2009, 1, 249–258. [Google Scholar] [PubMed]
- Aslam, M.I.; Hettmer, S.; Abraham, J.; Latocha, D.; Soundararajan, A.; Huang, E.T.; Goros, M.W.; Michalek, J.E.; Wang, S.; Mansoor, A.; et al. Dynamic and Nuclear Expression of PDGFRα and IGF-1R in Alveolar Rhabdomyosarcoma. Mol. Cancer Res. 2013, 11, 1303–1313. [Google Scholar] [CrossRef] [PubMed]
- Baron, O.; Förthmann, B.; Lee, Y.-W.; Terranova, C.; Ratzka, A.; Stachowiak, E.K.; Grothe, C.; Claus, P.; Stachowiak, M.K. Cooperation of Nuclear Fibroblast Growth Factor Receptor 1 and Nurr1 Offers New Interactive Mechanism in Postmitotic Development of Mesencephalic Dopaminergic Neurons. J. Biol. Chem. 2012, 287, 19827–19840. [Google Scholar] [CrossRef] [PubMed]
- Codony-Servat, J.; Cuatrecasas, M.; Asensio, E.; Montironi, C.; Martínez-Cardús, A.; Marín-Aguilera, M.; Horndler, C.; Martínez-Balibrea, E.; Rubini, M.; Jares, P.; et al. Nuclear IGF-1R predicts chemotherapy and targeted therapy resistance in metastatic colorectal cancer. Br. J. Cancer 2017, 117, 1777–1786. [Google Scholar] [CrossRef]
- Clément, F.; Martin, A.; Venara, M.; Calcagno, M.D.L.; Mathó, C.; Maglio, S.; Lombardi, M.G.; Bergadá, I.; A Pennisi, P. Type 1 IGF Receptor Localization in Paediatric Gliomas: Significant Association with WHO Grading and Clinical Outcome. Horm. Cancer 2018, 9, 205–214. [Google Scholar] [CrossRef]
- Aleksic, T.; Gray, N.; Wu, X.; Rieunier, G.; Osher, E.; Mills, J.; Verrill, C.; Bryant, R.J.; Han, C.; Hutchinson, K.; et al. Nuclear IGF1R Interacts with Regulatory Regions of Chromatin to Promote RNA Polymerase II Recruitment and Gene Expression Associated with Advanced Tumor Stage. Cancer Res. 2018, 78, 3497–3509. [Google Scholar] [CrossRef]
- Guerard, M.; Robin, T.; Perron, P.; Hatat, A.-S.; David-Boudet, L.; Vanwonterghem, L.; Busser, B.; Coll, J.-L.; Lantuejoul, S.; Eymin, B.; et al. Nuclear translocation of IGF1R by intracellular amphiregulin contributes to the resistance of lung tumour cells to EGFR-TKI. Cancer Lett. 2018, 420, 146–155. [Google Scholar] [CrossRef]
- Yang, C.; Zhang, Y.; Chen, Y.; Ragaller, F.; Liu, M.; Corvigno, S.; Dahlstrand, H.; Carlson, J.; Chen, Z.; Näsman, A.; et al. Nuclear IGF1R interact with PCNA to preserve DNA replication after DNA-damage in a variety of human cancers. PLoS ONE 2020, 15, e0236291. [Google Scholar] [CrossRef]
- Asmane, I.; Watkin, E.; Alberti, L.; Duc, A.; Marec-Berard, P.; Ray-Coquard, I.; Cassier, P.; Decouvelaere, A.-V.; Ranchère, D.; Kurtz, J.-E.; et al. Insulin-like growth factor type 1 receptor (IGF-1R) exclusive nuclear staining: A predictive biomarker for IGF-1R monoclonal antibody (Ab) therapy in sarcomas. Eur. J. Cancer 2012, 48, 3027–3035. [Google Scholar] [CrossRef]
- Björner, S.; Rosendahl, A.H.; Simonsson, M.; Markkula, A.; Jirström, K.; Borgquist, S.; Rose, C.; Ingvar, C.; Jernström, H. Body mass index influences the prognostic impact of combined nuclear insulin receptor and estrogen receptor expression in primary breast cancer. Front. Endocrinol. 2017, 8, 332. [Google Scholar] [CrossRef]
- Hancock, M.L.; Meyer, R.C.; Mistry, M.; Khetani, R.S.; Wagschal, A.; Shin, T.; Sui, S.J.H.; Näär, A.M.; Flanagan, J.G. Insulin Receptor Associates with Promoters Genome-wide and Regulates Gene Expression. Cell 2019, 177, 722–736.e22. [Google Scholar] [CrossRef]
- Hoa, N.; Tsui, S.; Afifiyan, N.F.; Hikim, A.S.; Li, B.; Douglas, R.S.; Smith, T.J. Nuclear Targeting of IGF-1 Receptor in Orbital Fibroblasts from Graves’ Disease: Apparent Role of ADAM17. PLoS ONE 2012, 7, e34173. [Google Scholar] [CrossRef]
- Rieger, L.; O’Connor, R.O. Controlled signaling-Insulin-like growth factor receptor endocytosis and presence at intracellular compartments. Front. Endocrinol. 2021, 11, 620013. [Google Scholar] [CrossRef]
- Poreba, E.; Durzynska, J. Nuclear localization and actions of the insulin-like growth factor 1 (IGF-1) system components: Transcriptional regulation and DNA damage response. Mutat. Res. 2020, 784, 108307. [Google Scholar] [CrossRef]
- Werner, H.; Bruchim, I. IGF-1 and BRCA1 signalling pathways in familial cancer. Lancet Oncol. 2012, 13, e537–e544. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Gene Names | Protein Names | Student’s t-Test Difference | Student’s t-Test p-Value |
|---|---|---|---|
| ALDH18A1 | Delta-1-pyrroline-5-carboxylate synthase | −6.61981074 | 0.000628706 |
| TJP2 | Tight junction protein ZO-2 | −6.070006053 | 1.25 × 10−8 |
| SIPA1L1 | Signal-induced proliferation-associated 1-like protein 1 | −5.783857346 | 9.09 × 10−7 |
| RPS29 | 40S ribosomal protein S29 | −5.609598796 | 3.73 × 10−5 |
| VAPB | Vesicle-associated membrane protein-associated protein B/C | −4.180510203 | 6.64 ×10−6 |
| TJP1 | Tight junction protein ZO-1 | −4.132112503 | 0.005198435 |
| C17orf85 | Uncharacterized protein C17orf85 | −4.00877889 | 0.001092293 |
| NUP88 | Nuclear pore complex protein Nup88 | −3.874612808 | 5.29 × 10−6 |
| IGF1R | Insulin-like growth factor 1 receptor | −3.842482885 | 0.000196745 |
| FAM162A | Protein FAM162A | −3.818694433 | 3.46 × 10−6 |
| SPRR1B | Cornifin-B | −3.621960322 | 0.000569671 |
| TJAP1 | Tight junction-associated protein 1 | −3.406965892 | 0.000618073 |
| TUBB | Tubulin beta chain | −3.353492101 | 0.000981451 |
| RAP1A | Ras-related protein Rap-1A | −3.285950343 | 7.10 × 10−5 |
| NOM1 | Nucleolar MIF4G domain-containing protein 1 | −3.033599218 | 3.15 × 10−5 |
| PABPC1 | Polyadenylate-binding protein 1 | −2.579547246 | 0.000578437 |
| ZNF687 | Zinc finger protein 687 | −2.193822225 | 0.001073949 |
| SART1 | U4/U6.U5 tri-snRNP-associated protein 1 | −0.605529785 | 0.001747125 |
| NHP2L1 | NHP2-like protein 1 | −0.575263341 | 0.00718376 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Werner, H.; Sarfstein, R.; Laron, Z. The Role of Nuclear Insulin and IGF1 Receptors in Metabolism and Cancer. Biomolecules 2021, 11, 531. https://doi.org/10.3390/biom11040531
Werner H, Sarfstein R, Laron Z. The Role of Nuclear Insulin and IGF1 Receptors in Metabolism and Cancer. Biomolecules. 2021; 11(4):531. https://doi.org/10.3390/biom11040531
Chicago/Turabian StyleWerner, Haim, Rive Sarfstein, and Zvi Laron. 2021. "The Role of Nuclear Insulin and IGF1 Receptors in Metabolism and Cancer" Biomolecules 11, no. 4: 531. https://doi.org/10.3390/biom11040531
APA StyleWerner, H., Sarfstein, R., & Laron, Z. (2021). The Role of Nuclear Insulin and IGF1 Receptors in Metabolism and Cancer. Biomolecules, 11(4), 531. https://doi.org/10.3390/biom11040531