
Update on Thiamine Triphosphorylated Derivatives and Metabolizing Enzymatic Complexes


Abstract
1. Introduction
- Thiamine was reported to be released during electrical nerve stimulation [11].

2. Thiamine Triphosphate: Occurrence, Synthesis, Regulation, and Role
2.1. Occurrence of ThTP in Living Organisms
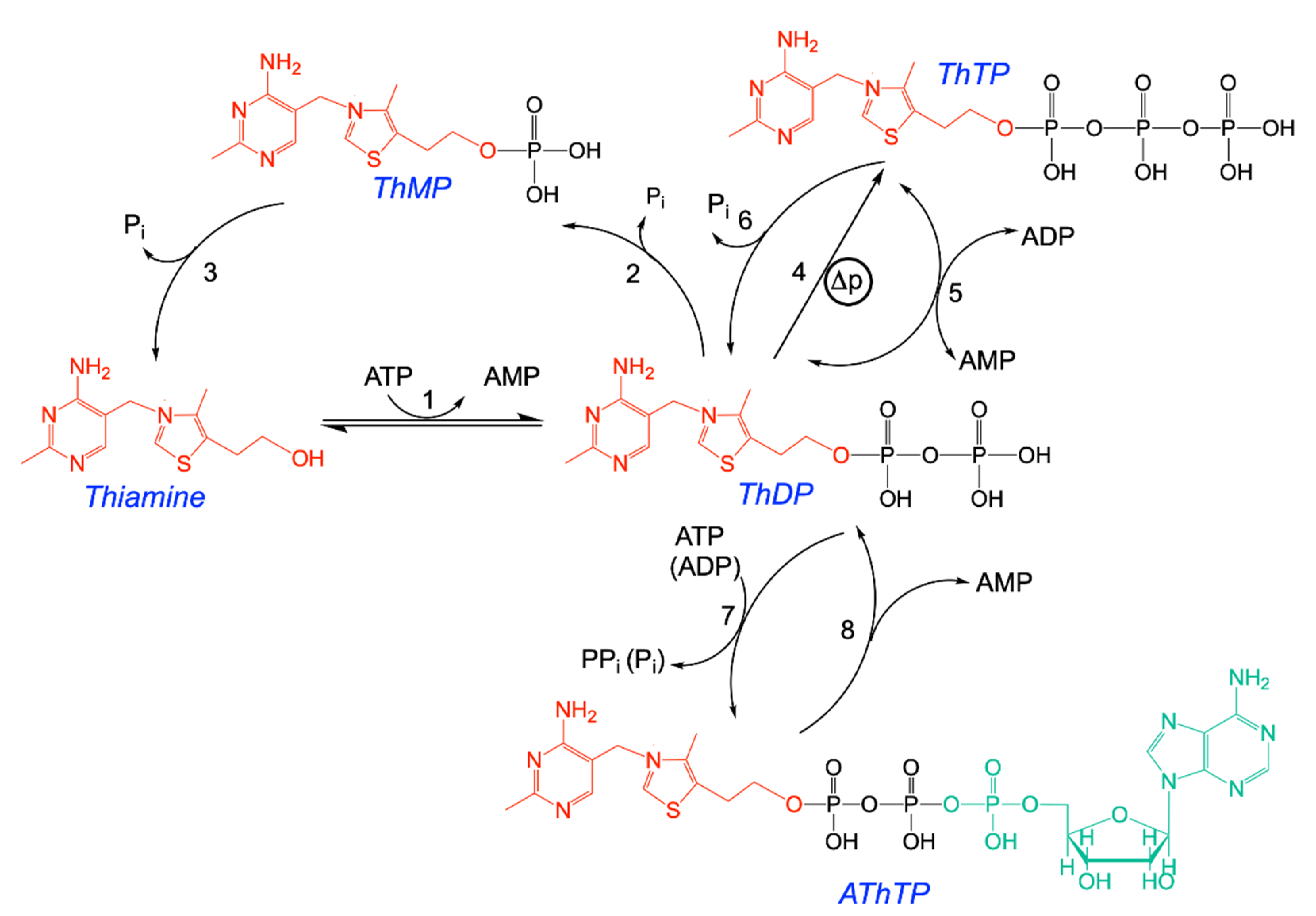
2.2. Two Different Mechanisms of Synthesis of ThTP: Adenylate Kinase and ATP Synthase
2.3. Regulation of Intracellular ThTP Concentrations
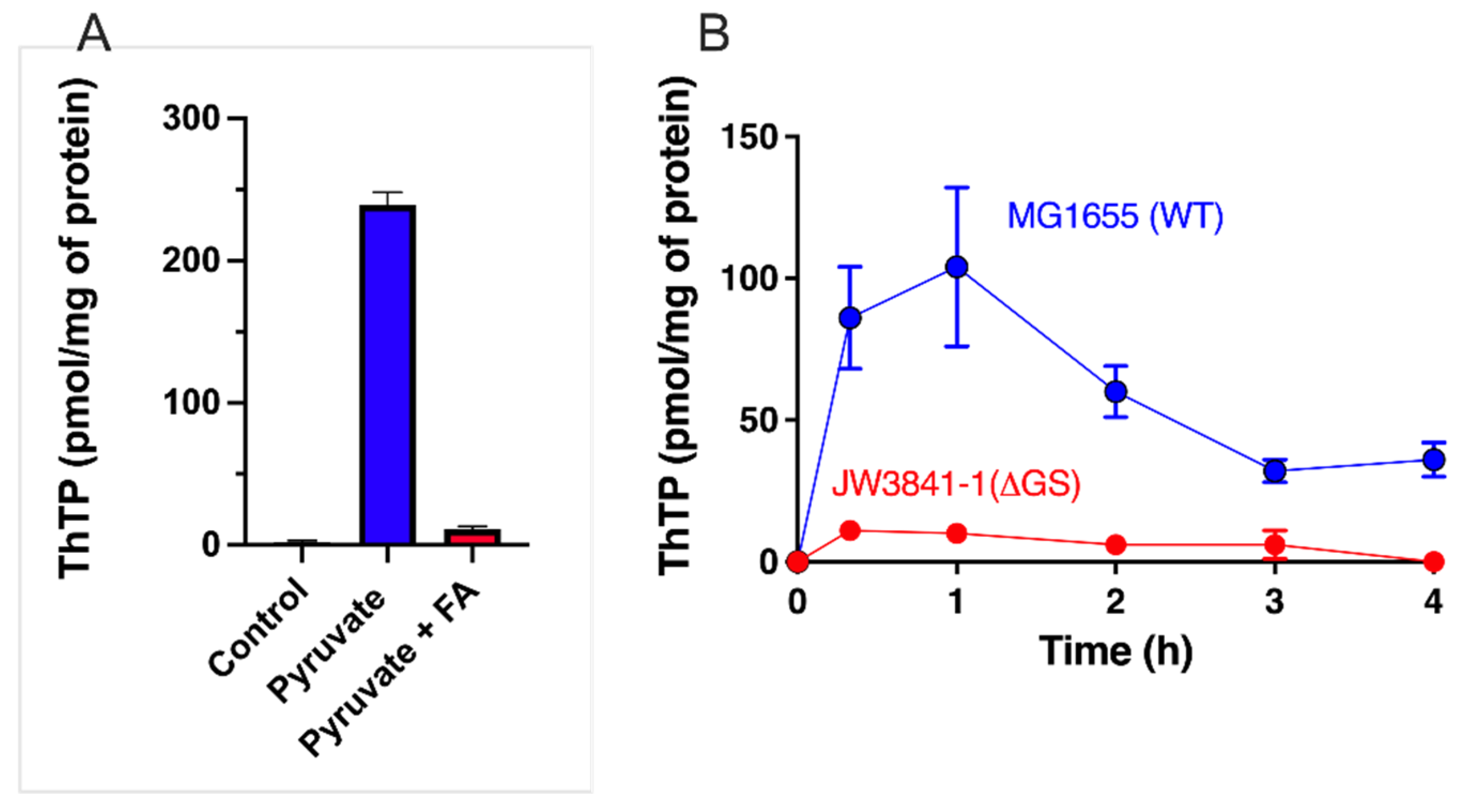
2.4. The Special Case of ThTP Synthesis in E. coli
2.5. Possible Physiological Roles of ThTP in Nerve Tissue
2.5.1. A Specific Role of ThTP in Nerve Excitability
- The electrical stimulation of isolated nerves leads to a release of thiamine, probably resulting from a dephosphorylation shift of higher thiamine phosphoester, such as ThDP and ThTP [11].
2.5.2. ThTP and Neurotransmitter Release
2.5.3. ThTP and Protein Phosphorylation
2.5.4. ThTP and Membrane Chloride Permeability
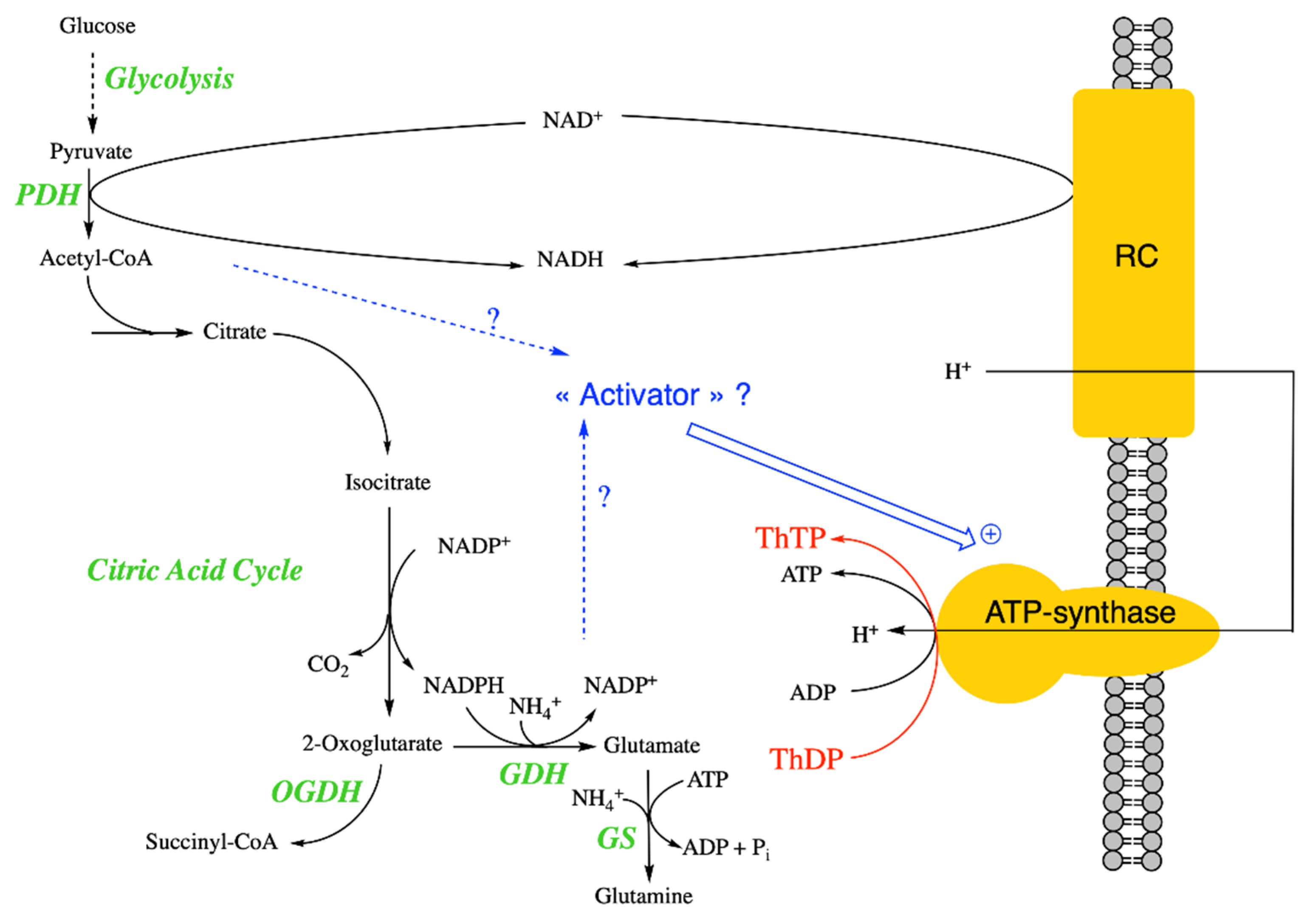
2.5.5. ThTP and Glutamate Dehydrogenase
3. Thiamine Triphosphatases
3.1. Mammalian Membrane-Associated ThTPase
3.2. Soluble 25-kDa ThTPase (THTPA)
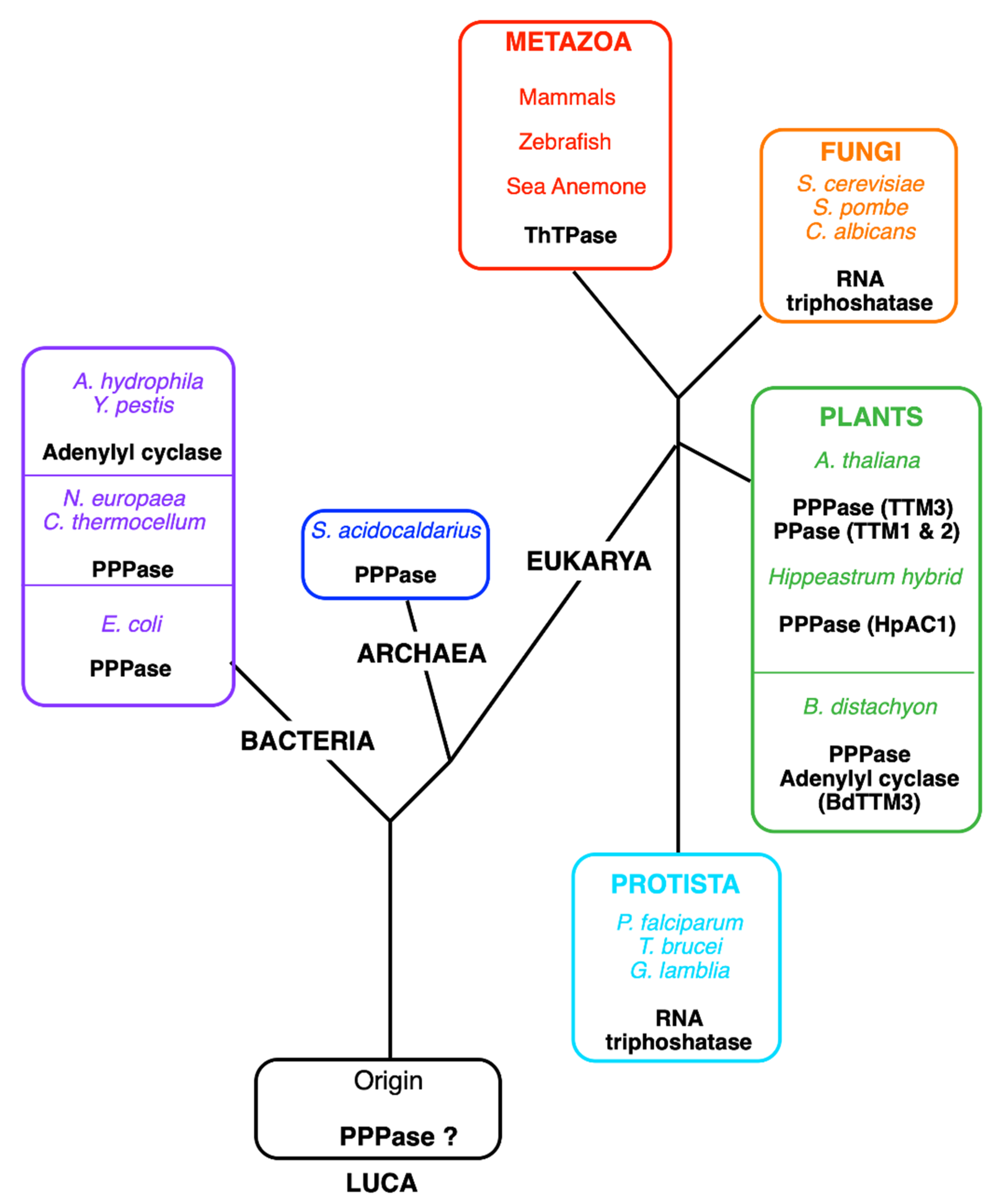
3.2.1. Characterization, Structure, and Evolution of THTPA

3.2.2. Possible Physiological Roles of THTPA and Regulation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | AK1 (µmol min−1 mg−1 Protein) | THTPA (nmol min−1 g−1 ww) | ThTP (nmol g−1 ww) |
|---|---|---|---|
| Fish E. electricus electric organ E. electricus brain T. marmorata electric organ | — — — | n. d. 1 n. d. 1 n. d. 1 | 3.9 ± 0.5 (87%) 1 0.37 ± 0.05 (7.1%) 1 45 ± 4 (38%) 2 |
| Birds Chicken skeletal muscle Chicken brain | 1.6 ± 0.1 3 — | n. d. n. d. | 3.7 (71) 4 0.92 (8.4%) 4 |
| Mammals Pig skeletal muscle Mouse skeletal muscle (wt) Mouse skeletal muscle (AK−/−) Mouse brain (wt) Mouse brain (AK−/−) | 1.03 + 0.15 3 1.3 ± 0.2 3 — 0.16 ± 0.2 3 0.02 ± 0.01 3 | 12.5 ± 0.5 4,6 42 3 — 280 ± 30 5 — | 20 (64%) 4 0.026 ± 0.006 (0.35%) 3 0.026 ± 0.008 (0.36%) 3 0.009 ± 0.003 (0.09%) 3 0.014 ± 0.01 (0.14%) 3 |
4. Adenylated Thiamine Nucleotides
4.1. Discovery, Chemical Synthesis, and Chemical Properties
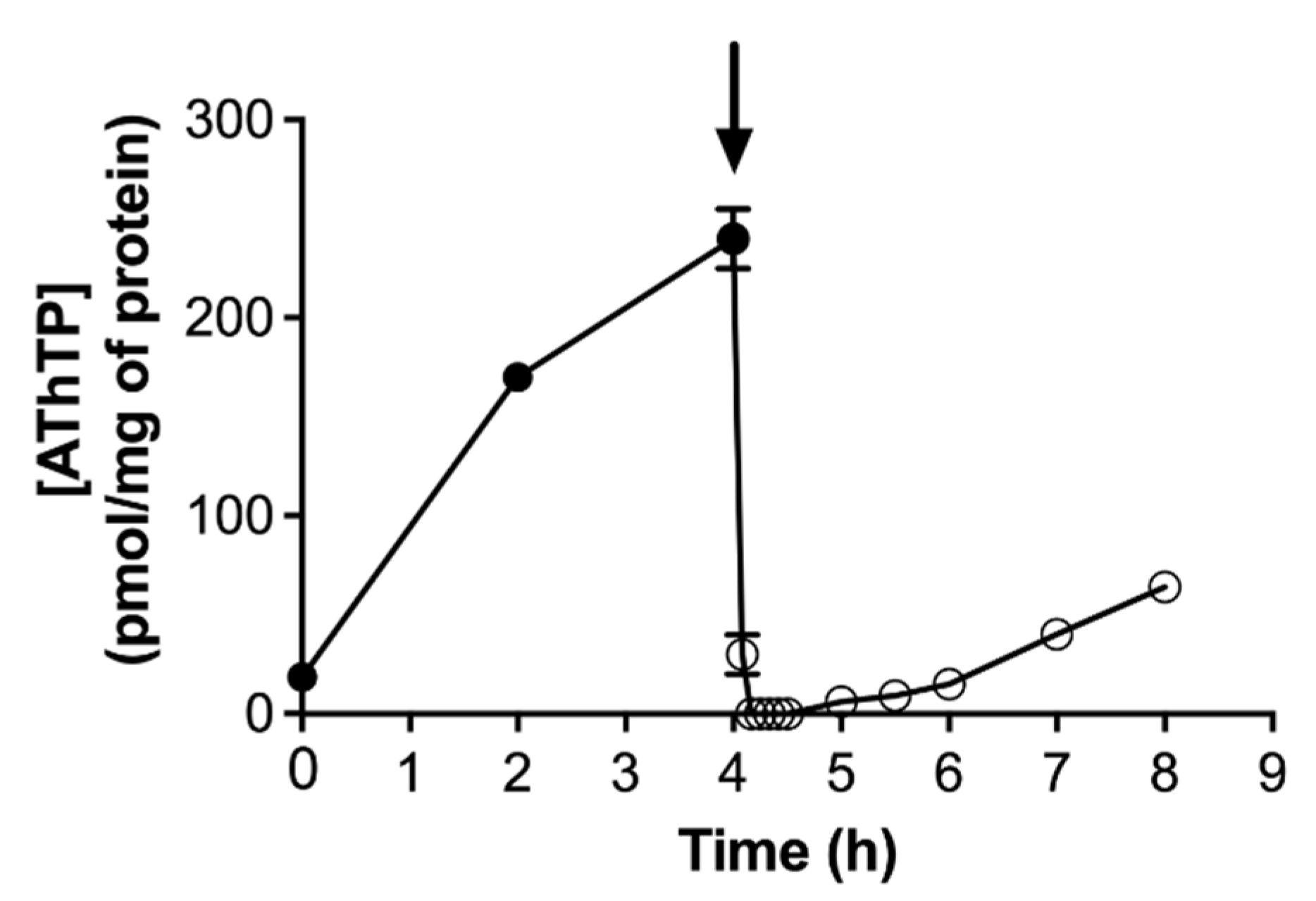
4.2. Regulation of AThTP Synthesis in E. coli
- It is not the absence of a carbon source per se that triggers AThTP synthesis, but the absence of its metabolization. Hence, AThTP accumulates in the presence of glucose when glycolysis is inhibited by iodoacetate, or in the presence of lactate when the respiratory chain is inhibited by KCN.
- AThTP synthesis probably requires a low molecular weight factor seemingly synthesized from pyruvate. Indeed, uncouplers, such as CCCP, induce ThTP synthesis only in the presence of pyruvate or a pyruvate-yielding substrate (D-glucose or L-lactate).
- ThTP is an inhibitor of AThTP synthesis, explaining at least in part that both compounds never accumulate together.
4.3. AThTP Is Synthesized by a High Molecular Weight Complex in E. coli
4.4. AThTP Hydrolysis in E. coli
4.5. AThTP Hydrolysis in Animal Tissues
4.6. Physiological Roles of Adenylated Thiamine Derivatives
5. Conclusions
- The first was the discovery that in E. coli, ThTP is induced under conditions of amino acid starvation in the presence of a carbon source, leading to the oxidation of pyruvate [28]. These results obtained from E. coli cells cannot necessarily be extrapolated to animal cells. Indeed, there are no known conditions where animals reversibly accumulate ThTP (we do not consider here constitutive synthesis by adenylate kinase in skeletal muscle and electric organs due to inactive THTPA).
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AK1 | Adenylate kinase 1 (myokinase) |
| AThDP | Adenosine thiamine diphosphate |
| AThTP | Adenosine thiamine triphosphate |
| GDH | Glutamate dehydrogenase |
| GS | Glutamine synthetase |
| ThDP | Thiamine diphosphate |
| ThMP | Thiamine monophosphate |
| ThTP | Thiamine triphosphate |
| THTPA | 25-kDa ThTPase |
| ThTPase | Thiamine triphosphatase |
| TPK1 | Thiamine diphosphokinase 1 |
| TTM | Triphosphate Tunnel Metalloenzyme |
References
- Kluger, R.; Tittmann, K. Thiamin Diphosphate Catalysis: Enzymic and Nonenzymic Covalent Intermediates. Chem. Rev. 2008, 108, 1797–1833. [Google Scholar] [CrossRef]
- Vogel, C.; Pleiss, J. The Modular Structure of ThDP-Dependent Enzymes. Proteins 2014, 82, 2523–2537. [Google Scholar] [CrossRef]
- Bunik, V.I.; Tylicki, A.; Lukashev, N.V. Thiamin Diphosphate-Dependent Enzymes: From Enzymology to Metabolic Regulation, Drug Design and Disease Models. FEBS J. 2013, 280, 6412–6442. [Google Scholar] [CrossRef]
- Bettendorff, L. 10. Thiamine. In Basic Nutrition and Metabolism; Present Knowledge in Nutrition 11th Edition; Marriott, B., Birt, D., Stalling, V., Yates, A., Eds.; Academic Press: Cambridge, MA, USA, 2020; Volume 1, p. 676. ISBN 978-0-323-66162-1. [Google Scholar]
- Abdou, E.; Hazell, A.S. Thiamine Deficiency: An Update of Pathophysiologic Mechanisms and Future Therapeutic Considerations. Neurochem. Res. 2015, 40, 353–361. [Google Scholar] [CrossRef]
- Whitfield, K.C.; Smith, G.; Chamnan, C.; Karakochuk, C.D.; Sophonneary, P.; Kuong, K.; Dijkhuizen, M.A.; Hong, R.; Berger, J.; Green, T.J.; et al. High Prevalence of Thiamine (Vitamin B1) Deficiency in Early Childhood among a Nationally Representative Sample of Cambodian Women of Childbearing Age and Their Children. PLoS Negl. Trop. Dis. 2017, 11, e0005814. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.R.; Fischer, P.R.; Thacher, T.D.; Topazian, M.D.; Bourassa, M.W.; Combs, G.F. Thiamin Deficiency in Low- and Middle-Income Countries: Disorders, Prevalences, Previous Interventions and Current Recommendations. Nutr. Health 2019, 25, 127–151. [Google Scholar] [CrossRef]
- Smith, T.J.; Johnson, C.R.; Koshy, R.; Hess, S.Y.; Qureshi, U.A.; Mynak, M.L.; Fischer, P.R. Thiamine Deficiency Disorders: A Clinical Perspective. Ann. N. Y. Acad. Sci. 2020. [Google Scholar] [CrossRef]
- Todd, K.; Butterworth, R.F. Mechanisms of Selective Neuronal Cell Death Due to Thiamine Deficiency. Ann. N. Y. Acad. Sci. 1999, 893, 404–411. [Google Scholar] [CrossRef]
- Gibson, G.; Nielsen, P.; Mykytyn, V.; Carlson, K.; Blass, J. Regionally Selective Alterations in Enzymatic Activities and Metabolic Fluxes during Thiamin Deficiency. Neurochem. Res. 1989, 14, 17–24. [Google Scholar] [CrossRef]
- Gurtner, H.P. Aneurin Und Nervenerregung. Versuche Mit 35S Markiertem Aneurin and Aneurinantimetaboliten. Helv. Physiol. Acta 1961, (Suppl. XI), 1–47. [Google Scholar]
- Bettendorff, L. Thiamine in Excitable Tissues: Reflections on a Non-Cofactor Role. Metab. Brain Dis. 1994, 9, 183–209. [Google Scholar] [CrossRef] [PubMed]
- Aleshin, V.A.; Mkrtchyan, G.V.; Bunik, V.I. Mechanisms of Non-Coenzyme Action of Thiamine: Protein Targets and Medical Significance. Biochem. Biokhimiia 2019, 84, 829–850. [Google Scholar] [CrossRef] [PubMed]
- Mkrtchyan, G.; Aleshin, V.; Parkhomenko, Y.; Kaehne, T.; Luigi Di Salvo, M.; Parroni, A.; Contestabile, R.; Vovk, A.; Bettendorff, L.; Bunik, V. Molecular Mechanisms of the Non-Coenzyme Action of Thiamin in Brain: Biochemical, Structural and Pathway Analysis. Sci. Rep. 2015, 5, 12583. [Google Scholar] [CrossRef]
- Makarchikov, A.F.; Lakaye, B.; Gulyai, I.E.; Czerniecki, J.; Coumans, B.; Wins, P.; Grisar, T.; Bettendorff, L. Thiamine Triphosphate and Thiamine Triphosphatase Activities: From Bacteria to Mammals. Cell Mol. Life Sci. 2003, 60, 1477–1488. [Google Scholar] [CrossRef]
- Bettendorff, L.; Lakaye, B.; Kohn, G.; Wins, P. Thiamine Triphosphate: A Ubiquitous Molecule in Search of a Physiological Role. Metab. Brain Dis. 2014, 29, 1069–1082. [Google Scholar] [CrossRef] [PubMed]
- Bettendorff, L.; Wirtzfeld, B.; Makarchikov, A.F.; Mazzucchelli, G.; Frédérich, M.; Gigliobianco, T.; Gangolf, M.; De Pauw, E.; Angenot, L.; Wins, P. Discovery of a Natural Thiamine Adenine Nucleotide. Nat. Chem. Biol. 2007, 3, 211–212. [Google Scholar] [CrossRef]
- Bettendorff, L.; Wins, P. Thiamin Diphosphate in Biological Chemistry: New Aspects of Thiamin Metabolism, Especially Triphosphate Derivatives Acting Other than as Cofactors. FEBS J. 2009, 276, 2917–2925. [Google Scholar] [CrossRef]
- Rossi-Fanelli, A.; Siliprandi, N.; Fasella, P. On the Presence of the Triphosphothiamine (TPT) in the Liver. Science 1952, 116, 711–713. [Google Scholar] [CrossRef]
- Kiessling, K.-H. Thiamine Triphosphate in Bakers’ Yeast. Nature 1953, 172, 1187–1188. [Google Scholar] [CrossRef] [PubMed]
- Greiling, H.; Kiesow, L. Zur Biochemie Des Thiamintriphosphorsäure IV. Mitt.: Das Vorkommen von Thiamintriphosphate Im Tierischen Organismus. Z. Naturforsch. 1958, 13, 251–252. [Google Scholar] [CrossRef]
- Rossi-Fanelli, A.; Ipata, P.L.; Fasella, P. On the Distribution and Transformation of Thiamine and Its Phosphoric Esters in Biological Material. Biochem. Biophys. Res. Commun. 1961, 4, 23–27. [Google Scholar] [CrossRef]
- Bettendorff, L.; Wins, P. Thiamine Derivatives in Excitable Tissues: Metabolism, Deficiency and Neurodegenerative Diseases. Recent Res. Devel. Neurochem. 1999, 2, 37–62. [Google Scholar]
- Bettendorff, L. The Compartmentation of Phosphorylated Thiamine Derivatives in Cultured Neuroblastoma Cells. Biochim. Biophys Acta 1994, 1222, 7–14. [Google Scholar] [CrossRef]
- Bettendorff, L.; Wins, P.; Lesourd, M. Subcellular Localization and Compartmentation of Thiamine Derivatives in Rat Brain. Biochim. Biophys. Acta 1994, 1222, 1–6. [Google Scholar] [CrossRef]
- Gangolf, M.; Czerniecki, J.; Radermecker, M.; Detry, O.; Nisolle, M.; Jouan, C.; Martin, D.; Chantraine, F.; Lakaye, B.; Wins, P.; et al. Thiamine Status in Humans and Content of Phosphorylated Thiamine Derivatives in Biopsies and Cultured Cells. PLoS ONE 2010, 5, e13616. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, T.; Tonomura, H.; Baba, A.; Iwata, H. Tissue Difference in Cellular Localization of Thiamine Phosphate Esters. Comp. Biochem. Physiol. B 1989, 94, 405–409. [Google Scholar] [CrossRef]
- Lakaye, B.; Wirtzfeld, B.; Wins, P.; Grisar, T.; Bettendorff, L. Thiamine Triphosphate, a New Signal Required for Optimal Growth of Escherichia coli during Amino Acid Starvation. J. Biol. Chem. 2004, 279, 17142–17147. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, M.; Loubéry, S.; Fitzpatrick, T.B. On the Nature of Thiamine Triphosphate in Arabidopsis. Plant Direct 2020, 4, e00258. [Google Scholar] [CrossRef]
- Chernikevich, I.P.; Luchko, V.; Voskoboev, A.I.; Ostrovsky, Y.M. Purification and Properties of ATP:Thiamine Diphosphate Phosphotransferase from Brewer’s Yeast. Biokhimiya 1984, 49, 899–907. [Google Scholar]
- Shikata, H.; Koyama, S.; Egi, Y.; Yamada, K.; Kawasaki, T. Cytosolic Adenylate Kinase Catalyzes the Synthesis of Thiamin Triphosphate from Thiamin Diphosphate. Biochem. Int. 1989, 18, 933–941. [Google Scholar] [PubMed]
- Gangolf, M.; Wins, P.; Thiry, M.; El Moualij, B.; Bettendorff, L. Thiamine Triphosphate Synthesis in Rat Brain Occurs in Mitochondria and Is Coupled to the Respiratory Chain. J. Biol. Chem. 2010, 285, 583–594. [Google Scholar] [CrossRef]
- Egi, Y.; Koyama, S.; Shikata, H.; Yamada, K.; Kawasaki, T. Content of Thiamin Phosphate Esters in Mammalian Tissues - an Extremely High Concentration of Thiamin Triphosphate in Pig Skeletal Muscle. Biochem. Int. 1986, 12, 385–390. [Google Scholar] [PubMed]
- Brune, M.; Schumann, R.; Wittinghofer, F. Cloning and Sequencing of the Adenylate Kinase Gene (Adk) of Escherichia coli. Nucleic Acids Res. 1985, 13, 7139–7151. [Google Scholar] [CrossRef] [PubMed]
- Gigliobianco, T.; Lakaye, B.; Makarchikov, A.F.; Wins, P.; Bettendorff, L. Adenylate Kinase-Independent Thiamine Triphosphate Accumulation under Severe Energy Stress in Escherichia coli. BMC Microbiol. 2008, 8, 16. [Google Scholar] [CrossRef] [PubMed]
- Makarchikov, A.F.; Wins, P.; Janssen, E.; Wieringa, B.; Grisar, T.; Bettendorff, L. Adenylate Kinase 1 Knockout Mice Have Normal Thiamine Triphosphate Levels. Biochim. Biophys. Acta 2002, 1592, 117–121. [Google Scholar] [CrossRef][Green Version]
- Gigliobianco, T.; Gangolf, M.; Lakaye, B.; Pirson, B.; von Ballmoos, C.; Wins, P.; Bettendorff, L. An Alternative Role of FoF1-ATP Synthase in Escherichia coli: Synthesis of Thiamine Triphosphate. Sci. Rep. 2013, 3, 1071. [Google Scholar] [CrossRef]
- Miyoshi, K.; Egi, Y.; Shioda, T.; Kawasaki, T. Evidence for in Vivo Synthesis of Thiamin Triphosphate by Cytosolic Adenylate Kinase in Chicken Skeletal Muscle. J. Biochem. Tokyo 1990, 108, 267–270. [Google Scholar] [CrossRef]
- Eder, L.; Dunant, Y. Thiamine and Cholinergic Transmission in the Electric Organ of Torpedo. I. Cellular Localization and Functional Changes of Thiamine and Thiamine Phosphate Esters. J. Neurochem. 1980, 35, 1278–1286. [Google Scholar] [CrossRef]
- Bettendorff, L.; Michel-Cahay, C.; Grandfils, C.; De Rycker, C.; Schoffeniels, E. Thiamine Triphosphate and Membrane-Associated Thiamine Phosphatases in the Electric Organ of Electrophorus Electricus. J. Neurochem. 1987, 49, 495–502. [Google Scholar] [CrossRef]
- Delvaux, D.; Kerff, F.; Murty, M.R.; Lakaye, B.; Czerniecki, J.; Kohn, G.; Wins, P.; Herman, R.; Gabelica, V.; Heuze, F.; et al. Structural Determinants of Specificity and Catalytic Mechanism in Mammalian 25-KDa Thiamine Triphosphatase. Biochim. Biophys. Acta 2013, 1830, 4513–4523. [Google Scholar] [CrossRef]
- Szyniarowski, P.; Lakaye, B.; Czerniecki, J.; Makarchikov, A.F.; Wins, P.; Margineanu, I.; Coumans, B.; Grisar, T.; Bettendorff, L. Pig Tissues Express a Catalytically Inefficient 25-KDa Thiamine Triphosphatase: Insight in the Catalytic Mechanisms of This Enzyme. Biochim. Biophys. Acta 2005, 1725, 93–102. [Google Scholar] [CrossRef]
- Zimatkina, T.I.; Chernikevich, I.P.; Zimatkin, S.M.; Deitrich, R.A. Thiamine Status in Liver and Brain of Rats Genetically Selected for Different Sensitivity to Hypnotic Effect of Alcohol [In Process Citation]. Alcohol. Clin. Exp. Res. 2000, 24, 1620–1624. [Google Scholar] [CrossRef]
- Wu, C.-C.; Ekanem, T.I.; Phan, N.N.; Loan, D.T.T.; Hou, S.-Y.; Lee, K.-H.; Wang, C.-Y. Gene Signatures and Prognostic Analyses of the Tob/BTG Pituitary Tumor-Transforming Gene (PTTG) Family in Clinical Breast Cancer Patients. Int. J. Med. Sci. 2020, 17, 3112–3124. [Google Scholar] [CrossRef] [PubMed]
- Liebecq, C.; Peters, R.A. The Toxicity of Fluoroacetate and the Tricarboxylic Acid Cycle. 1949. Biochim. Biophys. Acta 1989, 1000, 254–269. [Google Scholar] [PubMed]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia coli K-12 in-Frame, Single-Gene Knockout Mutants: The Keio Collection. Mol. Syst. Biol. 2006, 2, 2006.0008. [Google Scholar] [CrossRef]
- Gigliobianco, T. Biosynthèse des Dérivés Triphosphorylés de la Thiamine Chez Escherichia coli. Ph.D. Thesis, University of Liège, Liège, Belgium, 2012. [Google Scholar]
- Armingol, E.; Tobar, E.; Cabrera, R. Understanding the Impact of the Cofactor Swapping of Isocitrate Dehydrogenase over the Growth Phenotype of Escherichia coli on Acetate by Using Constraint-Based Modeling. PLoS ONE 2018, 13, e0196182. [Google Scholar] [CrossRef]
- Bettendorff, L.; Schoffeniels, E.; Naquet, R.; Silva-Barrat, C.; Riche, D.; Menini, C. Phosphorylated Thiamine Derivatives and Cortical Activity in the Baboon Papio Papio: Effect of Intermittent Light Stimulation. J. Neurochem. 1989, 53, 80–87. [Google Scholar] [CrossRef]
- Yamashita, H.; Zhang, Y.X.; Nakamura, S. The Effects of Thiamin and Its Phosphate Esters on Dopamine Release in the Rat Striatum. Neurosci. Lett. 1993, 158, 229–231. [Google Scholar] [CrossRef]
- Nghiêm, H.O.; Bettendorff, L.; Changeux, J.P. Specific Phosphorylation of Torpedo 43K Rapsyn by Endogenous Kinase(s) with Thiamine Triphosphate as the Phosphate Donor. FASEB J. 2000, 14, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Cooper, J.R.; Pincus, J.H. The Role of Thiamine in Nervous Tissue. Neurochem. Res. 1979, 4, 223–239. [Google Scholar] [CrossRef]
- Bettendorff, L.; Peeters, M.; Wins, P.; Schoffeniels, E. Metabolism of Thiamine Triphosphate in Rat Brain: Correlation with Chloride Permeability. J. Neurochem. 1993, 60, 423–434. [Google Scholar] [CrossRef]
- Bettendorff, L.; Hennuy, B.; De Clerck, A.; Wins, P. Chloride Permeability of Rat Brain Membrane Vesicles Correlates with Thiamine Triphosphate Content. Brain Res. 1994, 652, 157–160. [Google Scholar] [CrossRef]
- Bettendorff, L.; Hennuy, B.; Wins, P.; Schoffeniels, E. Thiamin and Derivatives as Modulators of Rat Brain Chloride Channels. Neuroscience 1993, 52, 1009–1017. [Google Scholar] [CrossRef]
- Bettendorff, L.; Kolb, H.A.; Schoffeniels, E. Thiamine Triphosphate Activates an Anion Channel of Large Unit Conductance in Neuroblastoma Cells. J. Membr. Biol. 1993, 136, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Sabirov, R.Z.; Islam, M.R.; Okada, T.; Merzlyak, P.G.; Kurbannazarova, R.S.; Tsiferova, N.A.; Okada, Y. The ATP-Releasing Maxi-Cl Channel: Its Identity, Molecular Partners and Physiological/Pathophysiological Implications. Life Basel Switz. 2021, 11, 509. [Google Scholar] [CrossRef]
- Fitzpatrick, T.B.; Chapman, L.M. The Importance of Thiamine (Vitamin B1) in Plant Health: From Crop Yield to Biofortification. J. Biol. Chem. 2020, 295, 12002–12013. [Google Scholar] [CrossRef]
- Murai, A.; Katsura, E. Thiamine Triphosphatase Activity of Myosin and Accelerating Effect of Thiamine Di- and Tri-Phosphates on Superprecipitation of Actomyosin. J. Nutr. Sci. Vitaminol. Tokyo 1975, 21, 169–181. [Google Scholar] [CrossRef]
- Shioda, T.; Kawasaki, T. Thiamin Triphosphate Does Not Affect Contraction of Skinned Fibers. J. Nutr. Sci. Vitaminol. Tokyo 1992, 38, 529–533. [Google Scholar] [CrossRef][Green Version]
- Barchi, R.L.; Braun, P.E. A Membrane-Associated Thiamine Triphosphatase from Rat Brain. Properties of the Enzyme. J. Biol. Chem. 1972, 247, 7668–7673. [Google Scholar] [CrossRef]
- Barchi, R.L. Membrane Thiamine Triphosphatase from Rat Brain: Inhibition by ATP and ADP. J. Neurochem. 1976, 26, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Bettendorff, L.; Wins, P.; Schoffeniels, E. Thiamine Triphosphatase from Electrophorus Electric Organ Is Anion-Dependent and Irreversibly Inhibited by 4,4′-Diisothiocyanostilbene-2,2′disulfonic Acid. Biochem. Biophys. Res. Commun. 1988, 154, 942–947. [Google Scholar] [CrossRef]
- Bettendorff, L.; Grandfils, C.; Wins, P.; Schoffeniels, E. Thiamine Triphosphatase in the Membranes of the Main Electric Organ of Electrophorus Electricus: Substrate-Enzyme Interactions. J. Neurochem. 1989, 53, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, T.; Tonomura, H.; Baba, A.; Iwata, H. Membrane-Associated Thiamine Triphosphatase in Rat Skeletal Muscle. Int J. Biochem. 1991, 23, 1111–1114. [Google Scholar] [PubMed]
- Bettendorff, L.; Longree, I.; Wins, P.; Schoffeniels, E. Solubilization of Thiamine Triphosphatase from the Electric Organ of Electrophorus Electricus. Biochim. Biophys. Acta 1991, 1073, 69–76. [Google Scholar] [CrossRef]
- Kolas, I.K.; Makarchikov, A.F. Properties of chicken liver membrane-associated thiamine triphosphatase. Ukr. Biochem. J. 2015, 87, 37–46. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sidorova, A.A.; Stepanenko, S.P.; Parkhomenko Iu, M. Characteristics of Thiamine Triphosphatase from Neural Cells Plasma Membranes. Ukr. Biokhim. Zh. 2009, 81, 57–65. [Google Scholar]
- Parkhomenko, I.M.; Strokina, A.A.; Pilipchuk, S.; Stepanenko, S.P.; Chekhovskaia, L.I.; Donchenko, G.V. Existence of Two Different Active Sites on Thiamine Binding Protein in Plasma Membranes of Synaptosomes. Ukr. Biokhim. Zh. 2010, 82, 34–41. [Google Scholar]
- Hashitani, Y.; Cooper, J.R. The Partial Purification of Thiamine Triphosphatase from Rat Brain. J. Biol. Chem. 1972, 247, 2117–2119. [Google Scholar] [CrossRef]
- Rusina, I.M.; Makarchikov, A.F. Thiamine Triphosphatase Activity in Mammalian Mitochondria. Ukr. Biokhim. Zh. 2003, 75, 63–68. [Google Scholar]
- Makarchikov, A.F.; Chernikevich, I.P. Purification and Characterization of Thiamine Triphosphatase from Bovine Brain. Biochim. Biophys. Acta 1992, 1117, 326–332. [Google Scholar] [CrossRef]
- Lakaye, B.; Makarchikov, A.F.; Antunes, A.F.; Zorzi, W.; Coumans, B.; De Pauw, E.; Wins, P.; Grisar, T.; Bettendorff, L. Molecular Characterization of a Specific Thiamine Triphosphatase Widely Expressed in Mammalian Tissues. J. Biol. Chem. 2002, 277, 13771–13777. [Google Scholar] [CrossRef]
- Lakaye, B.; Verlaet, M.; Dubail, J.; Czerniecki, J.; Bontems, S.; Makarchikov, A.F.; Wins, P.; Piette, J.; Grisar, T.; Bettendorff, L. Expression of 25 KDa Thiamine Triphosphatase in Rodent Tissues Using Quantitative PCR and Characterization of Its MRNA. Int. J. Biochem. Cell Biol. 2004, 36, 2032–2041. [Google Scholar] [CrossRef] [PubMed]
- Iyer, L.M.; Aravind, L. The Catalytic Domains of Thiamine Triphosphatase and CyaB-like Adenylyl Cyclase Define a Novel Superfamily of Domains That Bind Organic Phosphates. BMC Genom. 2002, 3, 33. [Google Scholar] [CrossRef]
- Gong, C.; Smith, P.; Shuman, S. Structure-Function Analysis of Plasmodium RNA Triphosphatase and Description of a Triphosphate Tunnel Metalloenzyme Superfamily That Includes Cet1-like RNA Triphosphatases and CYTH Proteins. RNA 2006, 12, 1468–1474. [Google Scholar] [CrossRef] [PubMed]
- Delvaux, D.; Murty, M.R.; Gabelica, V.; Lakaye, B.; Lunin, V.V.; Skarina, T.; Onopriyenko, O.; Kohn, G.; Wins, P.; De Pauw, E.; et al. A Specific Inorganic Triphosphatase from Nitrosomonas Europaea: Structure and Catalytic Mechanism. J. Biol. Chem. 2011, 286, 34023–34035. [Google Scholar] [CrossRef] [PubMed]
- Kohn, G.; Delvaux, D.; Lakaye, B.; Servais, A.C.; Scholer, G.; Fillet, M.; Elias, B.; Derochette, J.M.; Crommen, J.; Wins, P.; et al. High Inorganic Triphosphatase Activities in Bacteria and Mammalian Cells: Identification of the Enzymes Involved. PLoS ONE 2012, 7, e43879. [Google Scholar]
- Moeder, W.; Garcia-Petit, C.; Ung, H.; Fucile, G.; Samuel, M.A.; Christendat, D.; Yoshioka, K. Crystal Structure and Biochemical Analyses Reveal That the Arabidopsis Triphosphate Tunnel Metalloenzyme, AtTTM3, Is a Tripolyphosphatase and Is Involved in Root Development. Plant J. 2013, 76, 615–626. [Google Scholar] [CrossRef] [PubMed]
- Kleinboelting, S.; Miehling, J.; Steegborn, C. Crystal Structure and Enzymatic Characterization of the Putative Adenylyl Cyclase HpAC1 from Hippeastrum Reveal Dominant Triphosphatase Activity. J. Struct. Biol. 2020, 212, 107649. [Google Scholar] [CrossRef]
- Ung, H.; Karia, P.; Ebine, K.; Ueda, T.; Yoshioka, K.; Moeder, W. Triphosphate Tunnel Metalloenzyme Function in Senescence Highlights a Biological Diversification of This Protein Superfamily. Plant Physiol. 2017, 175, 473–485. [Google Scholar] [CrossRef]
- Vogt, M.S.; Ngouoko Nguepbeu, R.R.; Mohr, M.K.F.; Albers, S.-V.; Essen, L.-O.; Banerjee, A. The Archaeal Triphosphate Tunnel Metalloenzyme SaTTM Defines Structural Determinants for the Diverse Activities in the CYTH Protein Family. J. Biol. Chem. 2021, 297, 100820. [Google Scholar] [CrossRef]
- Bettendorff, L.; Wins, P. Thiamine Triphosphatase and the CYTH Superfamily of Proteins. FEBS J. 2013, 280, 6443–6455. [Google Scholar] [CrossRef]
- Świeżawska, B.; Duszyn, M.; Kwiatkowski, M.; Jaworski, K.; Pawełek, A.; Szmidt-Jaworska, A. Brachypodium Distachyon Triphosphate Tunnel Metalloenzyme 3 Is Both a Triphosphatase and an Adenylyl Cyclase Upregulated by Mechanical Wounding. FEBS Lett. 2020, 594, 1101–1111. [Google Scholar] [CrossRef] [PubMed]
- Makarchikov, A.F. The rate of molecular evolution of mammalian soluble thiamine triphosphatase. Ukr. Biokhim. Zh. 1999 2009, 81, 26–42. [Google Scholar] [PubMed]
- Song, J.; Bettendorff, L.; Tonelli, M.; Markley, J.L. Structural Basis for the Catalytic Mechanism of Mammalian 25-KDa Thiamine Triphosphatase. J. Biol. Chem. 2008, 283, 10939–10948. [Google Scholar] [CrossRef] [PubMed]
- Ung, H.; Moeder, W.; Yoshioka, K. Arabidopsis Triphosphate Tunnel Metalloenzyme2 Is a Negative Regulator of the Salicylic Acid-Mediated Feedback Amplification Loop for Defense Responses. Plant Physiol. 2014, 166, 1009–1021. [Google Scholar] [CrossRef]
- Sehnal, D.; Bittrich, S.; Deshpande, M.; Svobodová, R.; Berka, K.; Bazgier, V.; Velankar, S.; Burley, S.K.; Koča, J.; Rose, A.S. Mol* Viewer: Modern Web App for 3D Visualization and Analysis of Large Biomolecular Structures. Nucleic Acids Res. 2021, 49, W431–W437. [Google Scholar] [CrossRef]
- Czerniecki, J.; Chanas, G.; Verlaet, M.; Bettendorff, L.; Makarchikov, A.F.; Leprince, P.; Wins, P.; Grisar, T.; Lakaye, B. Neuronal Localization of the 25-KDa Specific Thiamine Triphosphatase in Rodent Brain. Neuroscience 2004, 125, 833–840. [Google Scholar] [CrossRef]
- Kovacevic, Z.; Fu, D.; Richardson, D.R. The Iron-Regulated Metastasis Suppressor, Ndrg-1: Identification of Novel Molecular Targets. Biochim. Biophys. Acta 2008, 1783, 1981–1992. [Google Scholar] [CrossRef]
- Suryo Rahmanto, Y.; Dunn, L.; Richardson, D. Identification of Distinct Changes in Gene Expression after Modulation of Melanoma Tumor Antigen P97 (Melanotransferrin) in Multiple Models in Vitro and in Vivo. Carcinogenesis 2007, 26, 2172–2183. [Google Scholar] [CrossRef]
- Terkelsen, T.; Pernemalm, M.; Gromov, P.; Børresen-Dale, A.-L.; Krogh, A.; Haakensen, V.D.; Lethiö, J.; Papaleo, E.; Gromova, I. High-Throughput Proteomics of Breast Cancer Interstitial Fluid: Identification of Tumor Subtype-Specific Serologically Relevant Biomarkers. Mol. Oncol. 2021, 15, 429–461. [Google Scholar] [CrossRef]
- Linster, C.L.; Van Schaftingen, E.; Hanson, A.D. Metabolite Damage and Its Repair or Pre-Emption. Nat. Chem. Biol. 2013, 9, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Kohn, G. Metabolic Significance of Inorganic Triphosphate, Thiamine Triphosphate and Their Hydrolyzing Enzyme; University of Liege: Liege, Belgium, 2015. [Google Scholar]
- Bettendorff, L.; Peeters, M.; Jouan, C.; Wins, P.; Schoffeniels, E. Determination of Thiamin and Its Phosphate Esters in Cultured Neurons and Astrocytes Using an Ion-Pair Reversed-Phase High-Performance Liquid Chromatographic Method. Anal. Biochem. 1991, 198, 52–59. [Google Scholar] [CrossRef]
- Frédérich, M.; Delvaux, D.; Gigliobianco, T.; Gangolf, M.; Dive, G.; Mazzucchelli, G.; Elias, B.; De Pauw, E.; Angenot, L.; Wins, P.; et al. Thiaminylated Adenine Nucleotides. Chemical Synthesis, Structural Characterization and Natural Occurrence. FEBS J. 2009, 276, 3256–3268. [Google Scholar] [CrossRef] [PubMed]
- Monteverde, D.R.; Gómez-Consarnau, L.; Suffridge, C.; Sañudo-Wilhelmy, S.A. Life’s Utilization of B Vitamins on Early Earth. Geobiology 2017, 15, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Makarchikov, A.F.; Saroka, T.V.; Kudyrka, T.G.; Kolas, I.K.; Luchko, T.A.; Rusina, I.M.; Gurinovich, V.A. Adenosine Thiamine Triphosphate and Adenosine Thiamine Triphosphate Hydrolase Activity in Animal Tissues. Ukr Biochem. J. 2018, 90, 52–63. [Google Scholar] [CrossRef]
- Hofer, A.; Marques, E.; Kieliger, N.; Gatter, S.-K.N.; Jordi, S.; Ferrari, E.; Hofmann, M.; Fitzpatrick, T.B.; Hottiger, M.O.; Jessen, H.J. Chemoselective Dimerization of Phosphates. Org. Lett. 2016, 18, 3222–3225. [Google Scholar] [CrossRef] [PubMed]
- Möhler, M.; Höfer, K.; Jäschke, A. Synthesis of 5′-Thiamine-Capped RNA. Molecules 2020, 25, 5492. [Google Scholar] [CrossRef]
- Makarchikov, A.F.; Brans, A.; Bettendorff, L. Thiamine Diphosphate Adenylyl Transferase from E. Coli: Functional Characterization of the Enzyme Synthesizing Adenosine Thiamine Triphosphate. BMC Biochem. 2007, 8, 17. [Google Scholar] [CrossRef]
- Hofmann, M. On the Role of Triphosphorylated Thiamine Compounds in Arabidopsis Thaliana. Ph.D. Thesis, Université de Genève, Genève, Switzerland, 2019. [Google Scholar]
- Gigliobianco, T.; Lakaye, B.; Wins, P.; El Moualij, B.; Zorzi, W.; Bettendorff, L. Adenosine Thiamine Triphosphate Accumulates in Escherichia coli Cells in Response to Specific Conditions of Metabolic Stress. BMC Microbiol. 2010, 10, 148. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Yamamoto, D.; Sato, T.; Tanaka, S.; Usui, K.; Manabe, M.; Aoki, Y.; Iwashima, Y.; Saito, Y.; Mino, Y.; et al. Adenosine Thiamine Triphosphate (AThTP) Inhibits Poly(ADP-Ribose) Polymerase-1 (PARP-1) Activity. J. Nutr. Sci. Vitaminol. Tokyo 2011, 57, 192–196. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bettendorff, L. Update on Thiamine Triphosphorylated Derivatives and Metabolizing Enzymatic Complexes. Biomolecules 2021, 11, 1645. https://doi.org/10.3390/biom11111645
Bettendorff L. Update on Thiamine Triphosphorylated Derivatives and Metabolizing Enzymatic Complexes. Biomolecules. 2021; 11(11):1645. https://doi.org/10.3390/biom11111645
Chicago/Turabian StyleBettendorff, Lucien. 2021. "Update on Thiamine Triphosphorylated Derivatives and Metabolizing Enzymatic Complexes" Biomolecules 11, no. 11: 1645. https://doi.org/10.3390/biom11111645
APA StyleBettendorff, L. (2021). Update on Thiamine Triphosphorylated Derivatives and Metabolizing Enzymatic Complexes. Biomolecules, 11(11), 1645. https://doi.org/10.3390/biom11111645