Molecular Dynamics of Chloroplast Membranes Isolated from Wild-Type Barley and a Brassinosteroid-Deficient Mutant Acclimated to Low and High Temperatures

 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Material and Methods
2.1. Plant Material, Experimental Design and Sampling
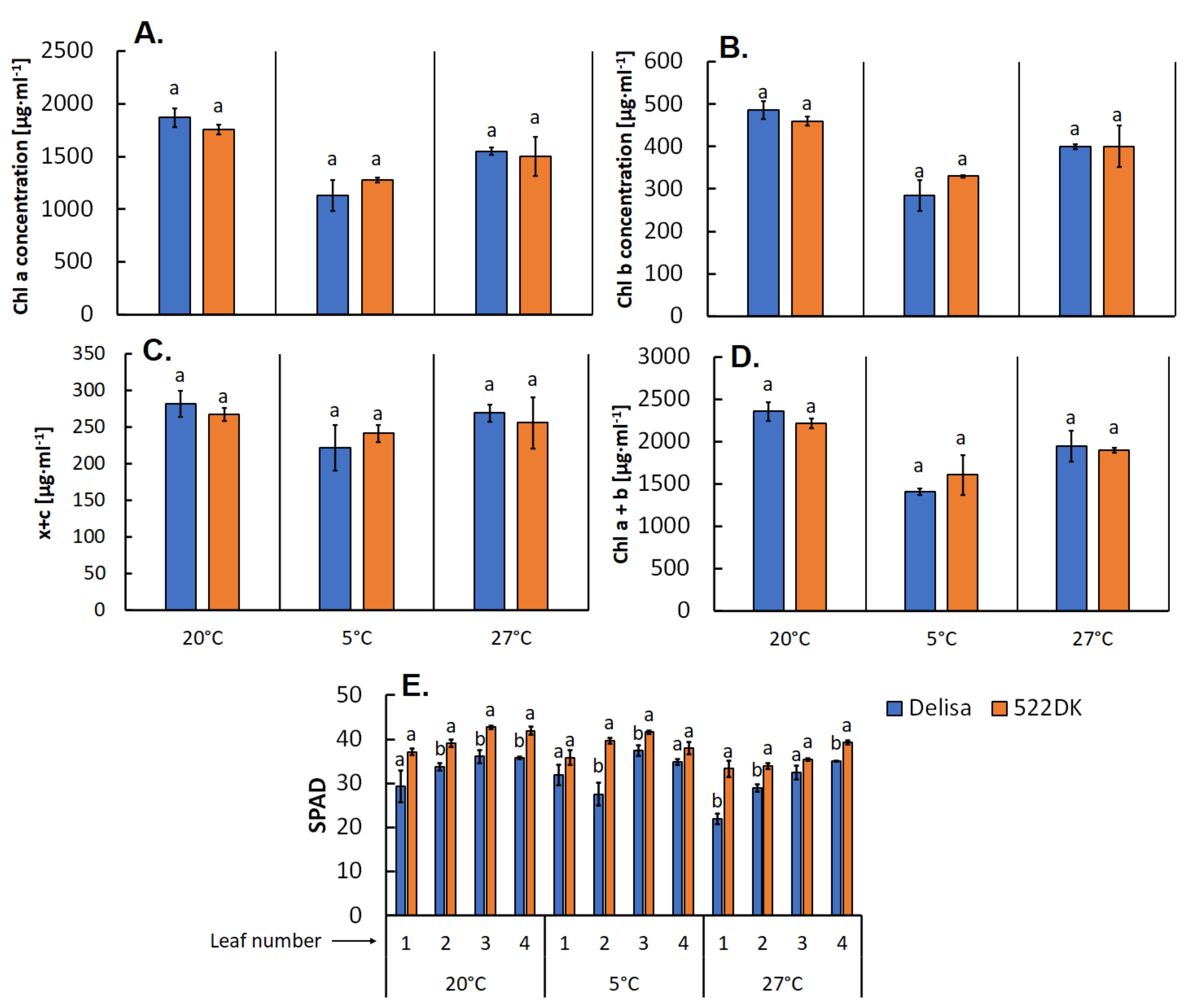
2.2. Leaf Greenness Measurement
2.3. Isolation of Chloroplasts from Barley Leaves
2.4. Analysis of the Protein Content in Chloroplasts
2.5. Analysis of the Fatty Acid Composition in Chloroplasts
2.6. Determining the Carotenoid and Chlorophyll Content in Chloroplasts
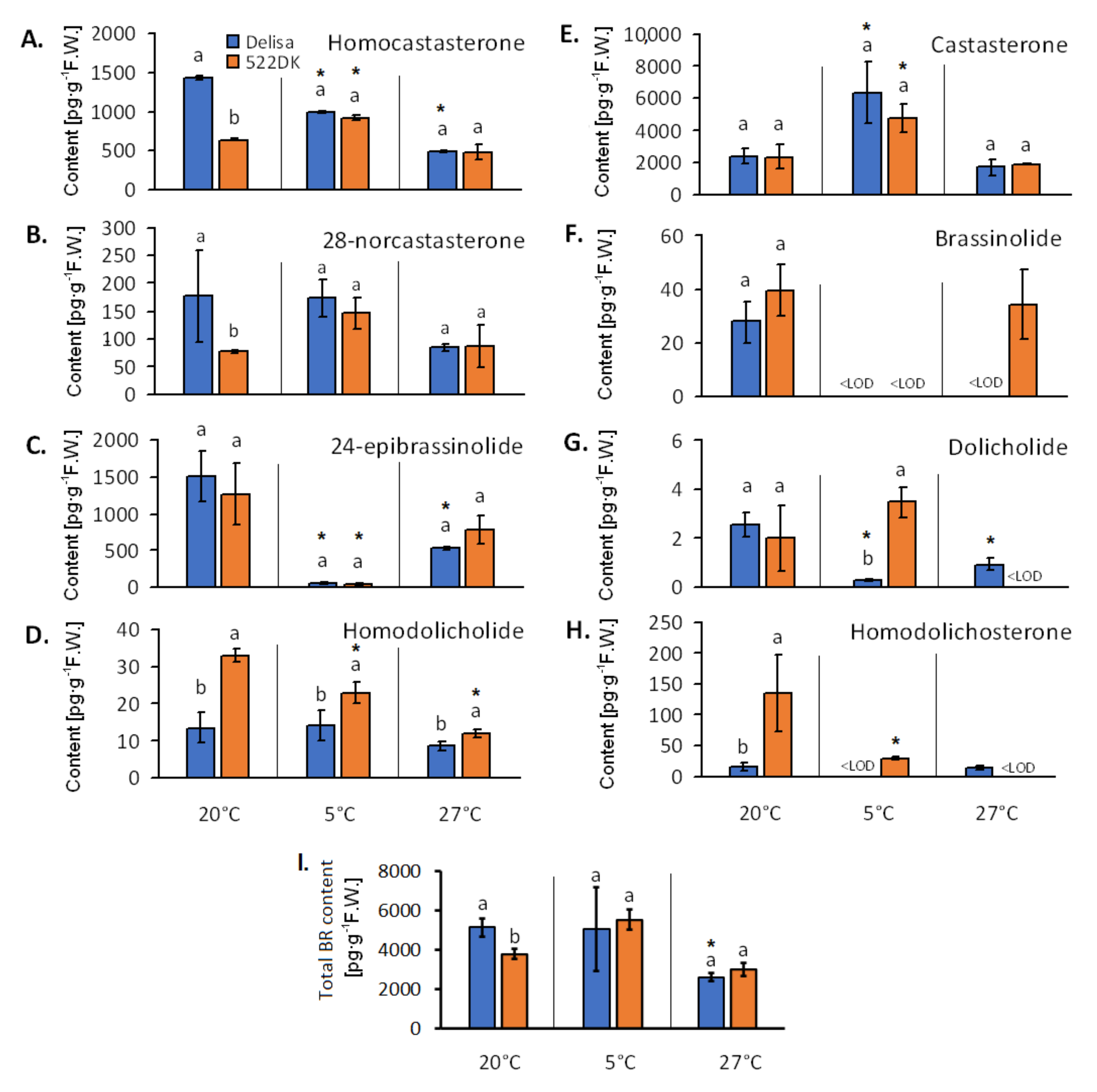
2.7. Analysis of the Brassinosteroids in the Chloroplasts
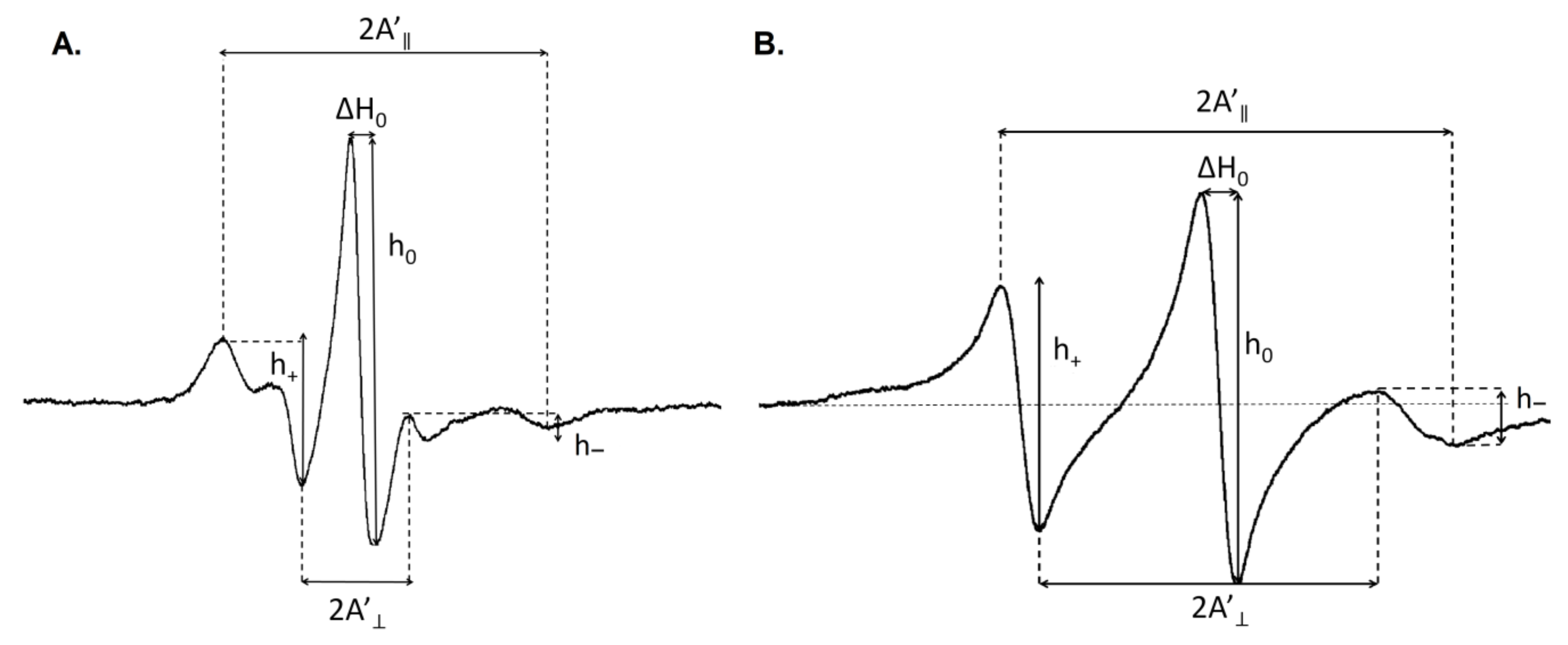
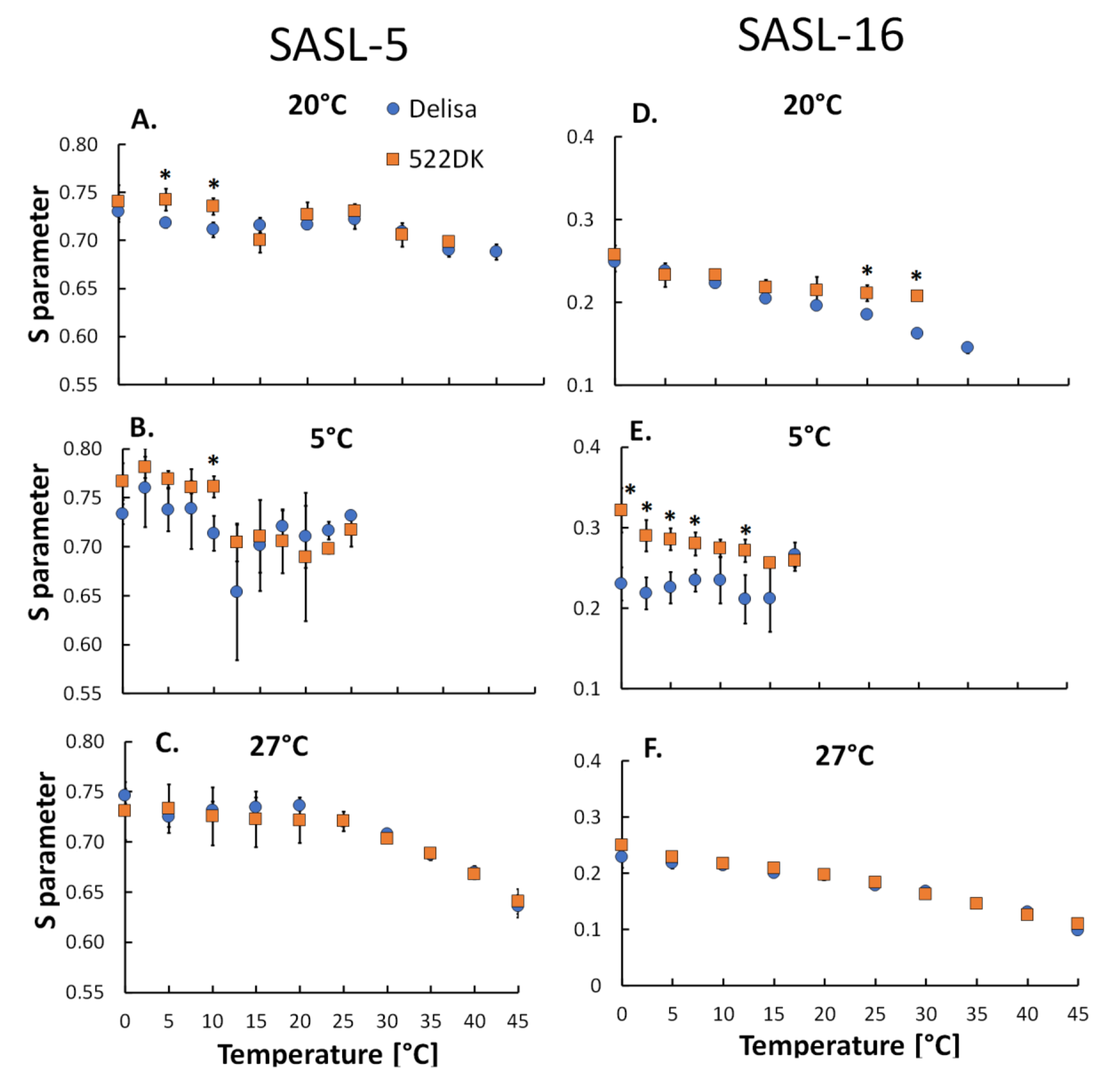
2.8. Electron Paramagnetic Resonance (EPR) Measurements
2.9. Statistical Analysis
3. Results and Discussion
3.1. Characteristic of Selected Chemical Components of the Chloroplast Membranes Isolated from Barley Grown at 20 °C, 5 °C, and 27 °C
3.1.1. Fatty Acid Composition of the Chloroplast Membranes
3.1.2. Carotenoid and Chlorophyll Content in the Chloroplast Membranes
3.1.3. Brassinosteroid Content of the Chloroplasts
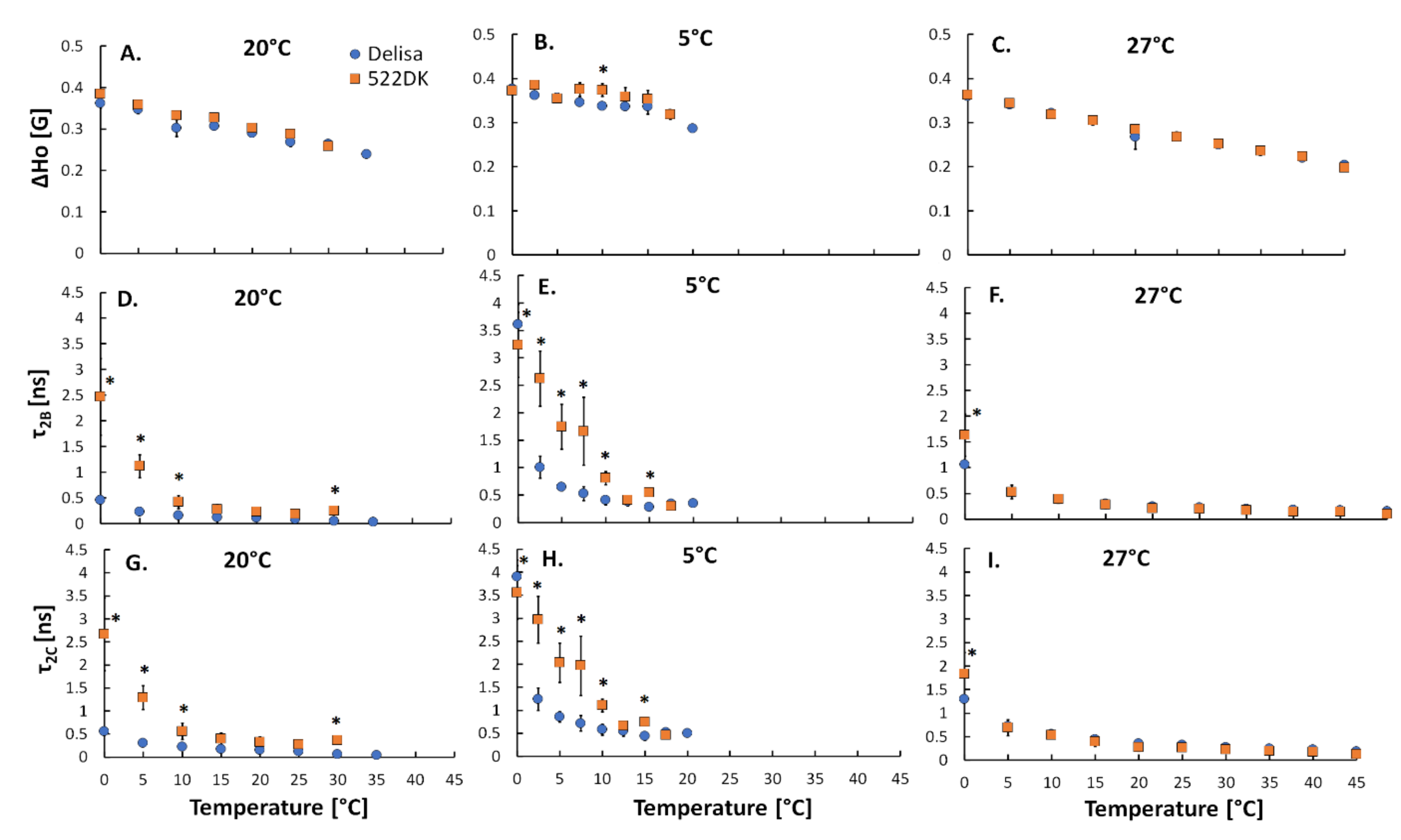
3.2. Molecular Dynamics of the Barley Chloroplasts Measured Using EPR Spectroscopy
4. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koyro, H.-W.; Ahmad, P.; Geissler, N. Abiotic stress responses in plants: An overview. In Environmental Adaptations and Stress Tolerance of Plants in the Era of Climate Change; Ahmad, P., Prasad, M., Eds.; Springer: New York, NY, USA, 2012; pp. 1–28. [Google Scholar]
- Hasanuzzaman, M.; Hossain, M.A.; da Silva, J.A.T.; Fujita, M. Plant response and tolerance to abiotic oxidative stress: Antioxidant defense is a key factor. In Crop Stress and Its Management: Perspectives and Strategies; Venkateswarlu, B., Shanker, A., Shanker, C., Maheswari, M., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 261–315. [Google Scholar]
- Prasad, M.N.V.; Rengel, Z. Plant acclimation and adaptation to natural and anthropogenic stress. Ann. N. Y. Acad. Sci. 1998, 851, 216–223. [Google Scholar] [CrossRef]
- Horváth, I.; Glatz, A.; Nakamoto, H.; Mishkind, M.L.; Munnik, T.; Saidi, Y.; Goloubinoff, P.; Harwood, J.L.; Vigh, L. Heat shock response in photosynthetic organisms: Membrane and lipid connections. Prog. Lipid Res. 2012, 51, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Gzyl-Malcher, B.; Zembala, M.; Filek, M. Effect of tocopherol on surface properties of plastid lipids originating from wheat calli cultivated in cadmium presence. Chem. Phys. Lipids 2010, 163, 74–81. [Google Scholar] [CrossRef]
- Strzałka, K.; Kostecka-Gugała, A.; Latowski, D. Carotenoids and environmental stress in plants: Significance of carotenoid-mediated modulation of membrane physical properties. Russ. J. Plant Physiol. 2003, 50, 168–173. [Google Scholar] [CrossRef]
- Mitchell, J.W.; Mandava, N.; Worley, J.F.; Plimmer, J.R.; Smith, M.V. Brassins—A new family of plant hormones from rape pollen. Nat. Cell Biol. 1970, 225, 1065–1066. [Google Scholar] [CrossRef]
- Grove, M.D.; Spencer, G.F.; Rohwedder, W.K.; Mandava, N.; Worley, J.F.; Warthen, J.D.; Steffens, G.L.; Flippen-Anderson, J.L.; Cook, J.C. Brassinolide, a plant growth-promoting steroid isolated from Brassica napus pollen. Nat. Cell Biol. 1979, 281, 216–217. [Google Scholar] [CrossRef]
- Bajguz, A.; Hayat, S. Effects of brassinosteroids on the plant responses to environmental stresses. Plant Physiol. Biochem. 2009, 47, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sadura, I.; Janeczko, A. Physiological and molecular mechanisms of brassinosteroid-induced tolerance to high and low temperature in plants. Biol. Plant. 2018, 62, 601–616. [Google Scholar] [CrossRef]
- Sadura, I.; Libik-Konieczny, M.; Jurczyk, B.; Gruszka, D.; Janeczko, A. Plasma membrane ATPase and the aquaporin HvPIP1 in barley brassinosteroid mutants acclimated to high and low temperature. J. Plant Physiol. 2020, 244, 153090. [Google Scholar] [CrossRef]
- Sadura, I.; Libik-Konieczny, M.; Jurczyk, B.; Gruszka, D.; Janeczko, A. HSP transcript and protein accumulation in brassinosteroid barley mutants acclimated to low and high temperatures. Int. J. Mol. Sci. 2020, 21, 1889. [Google Scholar] [CrossRef]
- Rudolphi-Szydło, E.; Sadura, I.; Filek, M.; Gruszka, D.; Janeczko, A. The impact of mutations in the HvCPD and HvBRI1 genes on the physicochemical properties of the membranes from barley acclimated to low/high temperatures. Cells 2020, 9, 1125. [Google Scholar] [CrossRef] [PubMed]
- Bojko, M.; Olchawa-Pajor, M.; Goss, R.; Schaller-Laudel, S.; Strzałka, K.; Latowski, D. Diadinoxanthin de-epoxidation as important factor in the short-term stabilization of diatom photosynthetic membranes exposed to different temperatures. Plant Cell Environ. 2018, 42, 1270–1286. [Google Scholar] [CrossRef] [PubMed]
- Strzałka, K.; Hara-Nishimura, I.; Nishimura, M. Changes in physical properties of vacuolar membrane during transformation of protein bodies into vacuoles in germinating pumpkin seeds. Biochim. Biophys. Acta (BBA) Biomembr. 1995, 1239, 103–110. [Google Scholar] [CrossRef]
- Jajić, I.; Wisniewska-Becker, A.; Sarna, T.; Jemioła-Rzemińska, M.; Strzałka, K. EPR spin labeling measurements of thylakoid membrane fluidity during barley leaf senescence. J. Plant Physiol. 2014, 171, 1046–1053. [Google Scholar] [CrossRef] [PubMed]
- Pottosin, I.; Shabala, S. Transport across chloroplast membranes: Optimizing photosynthesis for adverse environmental conditions. Mol. Plant 2016, 9, 356–370. [Google Scholar] [CrossRef]
- Gruszka, D.; Szarejko, I.; Maluszynski, M. Identification of barley DWARF gene involved in brassinosteroid synthesis. Plant Growth Regul. 2011, 65, 343–358. [Google Scholar] [CrossRef]
- Gruszka, D.; Gorniak, M.; Glodowska, E.; Wierus, E.; Oklestkova, J.; Janeczko, A.; Maluszynski, M.; Szarejko, I. A reverse-genetics mutational analysis of the barley HvDWARF gene results in identification of a series of alleles and mutants with short stature of various degree and disturbance in BR biosynthesis allowing a new insight into the process. Int. J. Mol. Sci. 2016, 17, 600. [Google Scholar] [CrossRef]
- Sadura, I.; Pociecha, E.; Dziurka, M.; Oklestkova, J.; Novak, O.; Gruszka, D.; Janeczko, A. Mutations in the HvDWARF, HvCPD and HvBRI1 genes-involved in brassinosteroid biosynthesis/signalling: Altered photosynthetic efficiency, hormonal homeostasis and tolerance to high/low temperatures in barley. J. Plant Growth Regul. 2019, 38, 1062–1081. [Google Scholar] [CrossRef]
- Pociecha, E.; Dziurka, M.; Oklestkova, J.; Janeczko, A. Brassinosteroids increase winter survival of winter rye (Secale cereale L.) by affecting photosynthetic capacity and carbohydrate metabolism during the cold acclimation process. Plant Growth Regul. 2016, 80, 127–135. [Google Scholar] [CrossRef]
- Janeczko, A.; Pociecha, E.; Dziurka, M.; Jurczyk, B.; Libik-Konieczny, M.; Oklestkova, J.; Novak, O.; Pilarska, M.; Filek, M.; Rudolphi-Skórska, E.; et al. Changes in content of steroid regulators during cold hardening of winter wheat—Steroid physiological/biochemical activity and impact on frost resistance. Plant Physiol. Biochem. 2019, 139, 215–228. [Google Scholar] [CrossRef]
- Fowler, D.B.; Limin, A.E.; Wang, S.-Y.; Ward, R. Relationship between low-temperature tolerance and vernalization response in wheat and rye. Can. J. Plant Sci. 1996, 76, 37–42. [Google Scholar] [CrossRef]
- Wu, X.; Yao, X.; Chen, J.; Zhu, Z.; Zhang, H.; Zha, D.S. Brassinosteroids protect photosynthesis and antioxidant system of eggplant seedlings from high-temperature stress. Acta Physiol. Plant. 2014, 36, 251–261. [Google Scholar] [CrossRef]
- Thussagunpanit, J.; Jutamanee, K.; Kaveeta, L.; Chai-Arree, W.; Pankean, P.; Homvisasevongsa, S.; Suksamrarn, A. Comparative effects of brassinosteroid and brassinosteroid mimic on improving photosynthesis, lipid peroxidation, and rice seed set under heat stress. J. Plant Growth Regul. 2015, 34, 320–331. [Google Scholar] [CrossRef]
- Block, M.A.; Dorne, A.J.; Joyard, J.; Douce, R. Preparation and characterization of membrane fractions enriched in outer and inner envelope membranes from spinach chloroplasts. II. Biochemical characterization. J. Biol. Chem. 1983, 258, 13281–13286. [Google Scholar]
- Filek, M.; Sieprawska, A.; Kościelniak, J.; Oklestkova, J.; Jurczyk, B.; Telk, A.; Biesaga-Kościelniak, J.; Janeczko, A. The role of chloroplasts in the oxidative stress that is induced by zearalenone in wheat plants—The functions of 24-epibrassinolide and selenium in the protective mechanisms. Plant Physiol. Biochem. 2019, 137, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Sedmak, J.; Grossberg, S.E. A rapid, sensitive, and versatile assay for protein using Coomassie brilliant blue G250. Anal. Biochem. 1977, 79, 544–552. [Google Scholar] [CrossRef]
- Trela, A.; Silska, G.; Chyc, M.; Latowski, D.; Kruk, J.; Szymańska, R. Tocochromanols and fatty acid composition in flax (Linum usitatissimum L.) accessions. Acta Soc. Bot. Pol. 2019, 88, 1–12. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Methods in Enzymology; Academic Press: Orlando, FL, USA, 1987; Volume 148, pp. 350–382. [Google Scholar]
- Oklestkova, J.; Tarkowská, D.; Eyer, L.; Elbert, T.; Marek, A.; Smržová, Z.; Novák, O.; Fránek, M.; Zhabinskii, V.N.; Strnad, M. Immunoaffinity chromatography combined with tandem mass spectrometry: A new tool for the selective capture and analysis of brassinosteroid plant hormones. Talanta 2017, 170, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Tarkowská, D.; Novák, O.; Oklestkova, J.; Strnad, M. The determination of 22 natural brassinosteroids in a minute sample of plant tissue by UHPLC–ESI–MS/MS. Anal. Bioanal. Chem. 2016, 408, 6799–6812. [Google Scholar] [CrossRef]
- Marsch, D. Electron spin resonance: Spin labels. In Membrane Spectroscopy; Grell, E., Ed.; Springer: Berlin/Heidelberg, Germany, 1981; Volume 31, pp. 51–137. [Google Scholar]
- Berliner, L.J. Spin labeling in enzymology: Spin-labeled enzymes and proteins. Methods Enzymol. 1978, 49, 418–480. [Google Scholar] [CrossRef]
- Mikami, K.; Murata, N. Membrane fluidity and the perception of environmental signals in cyanobacteria and plants. Prog. Lipid Res. 2003, 42, 527–543. [Google Scholar] [CrossRef]
- Los, D.A.; Murata, N. Membrane fluidity and its roles in the perception of environmental signals. Biochim. Biophys. Acta (BBA) Biomembr. 2004, 1666, 142–157. [Google Scholar] [CrossRef]
- Hölzl, G.; Dörmann, P. Chloroplast lipids and their biosynthesis. Annu. Rev. Plant Biol. 2019, 70, 51–81. [Google Scholar] [CrossRef] [PubMed]
- Schwertner, H.A.; Biale, J.B. Lipid composition of plant mitochondria and of chloroplasts. J. Lipid Res. 1973, 14, 235–242. [Google Scholar] [PubMed]
- Janeczko, A.; Hura, K.; Skoczowski, A.; Idzik, I.; Biesaga-Kościelniak, J.; Niemczyk, E. Temperature-dependent impact of 24-epibrassinolide on the fatty acid composition and sugar content in winter oilseed rape callus. Acta Physiol. Plant. 2009, 31, 71–79. [Google Scholar] [CrossRef]
- Fedina, E.; Yarin, A.; Mukhitova, F.; Blufard, A.; Chechetkin, I. Brassinosteroid-induced changes of lipid composition in leaves of Pisum sativum L. during senescence. Steroids 2017, 117, 25–28. [Google Scholar] [CrossRef]
- Filek, M.; Rudolphi-Skórska, E.; Sieprawska, A.; Kvasnica, M.; Janeczko, A. Regulation of the membrane structure by brassinosteroids and progesterone in winter wheat seedlings exposed to low temperature. Steroids 2017, 128, 37–45. [Google Scholar] [CrossRef]
- Janeczko, A.; Gruszka, D.; Pociecha, E.; Dziurka, M.; Filek, M.; Jurczyk, B.; Kalaji, H.M.; Kocurek, M.; Waligórski, P. Physiological and biochemical characterisation of watered and drought-stressed barley mutants in the HvDWARF gene encoding C6-oxidase involved in brassinosteroid biosynthesis. Plant Physiol. Biochem. 2016, 99, 126–141. [Google Scholar] [CrossRef]
- Havaux, M. Carotenoids as membrane stabilizers in chloroplasts. Trends Plant Sci. 1998, 3, 147–151. [Google Scholar] [CrossRef]
- Nie, S.; Huang, S.; Wang, S.; Cheng, D.; Liu, J.; Lv, S.; Li, Q.; Wang, X. Enhancing brassinosteroid signaling via overexpression of tomato (Solanum lycopersicum) SlBRI1 improves major agronomic traits. Front. Plant Sci. 2017, 8, 1386. [Google Scholar] [CrossRef]
- Kočová, M.; Rothová, O.; Holá, D.; Kvasnica, M.; Kohout, L. The effects of brassinosteroids on photosynthetic parameters in leaves of two field-grown maize inbred lines and their F1 hybrid. Biol. Plant. 2010, 54, 785–788. [Google Scholar] [CrossRef]
- Malaga, S.; Janeczko, A.; Janowiak, F.; Waligórski, P.; Oklestkova, J.; Dubas, E.; Krzewska, M.; Nowicka, A.; Surówka, E.; Rapacz, M.; et al. Involvement of homocastasterone, salicylic and abscisic acids in the regulation of drought and freezing tolerance in doubled haploid lines of winter barley. Plant Growth Regul. 2020, 90, 173–188. [Google Scholar] [CrossRef]
- Jørgensen, K.; Rasmussen, A.V.; Morant, M.; Nielsen, A.H.; Bjarnholt, N.; Zagrobelny, M.; Bak, S.; Møller, B.L. Metabolon formation and metabolic channeling in the biosynthesis of plant natural products. Curr. Opin. Plant Biol. 2005, 8, 280–291. [Google Scholar] [CrossRef]
- Cai, Y.; Tarin, M.; Fan, L.; Xie, D.; Rong, J.; He, T.; Chen, L.; Zheng, Y. Responses of photosynthesis, chloroplast ultrastructure, and antioxidant system of morinda officinalis how to exogenous 2,4-pibrassinolide treatments under high temperature stress. Appl. Ecol. Environ. Res. 2020, 18, 3981–4004. [Google Scholar] [CrossRef]
- Zhang, F.; Lu, K.; Gu, Y.; Zhang, L.; Li, W.; Tan, X. Effects of low-temperature stress and brassinolide application on the photosynthesis and leaf structure of tung tree seedlings. Front. Plant Sci. 2020, 10, 1767. [Google Scholar] [CrossRef]
- Krumova, S.; Zhiponova, M.; Dankov, K.; Velikova, V.; Balashev, K.; Andreeva, T.; Russinova, E.; Taneva, S. Brassinosteroids regulate the thylakoid membrane architecture and the photosystem II function. J. Photochem. Photobiol. B Biol. 2013, 126, 97–104. [Google Scholar] [CrossRef]
- Bojko, M.; Olchawa-Pajor, M.; Chyc, M.; Goss, R.; Schaller-Laudel, S.; Latowski, D. Acclimatization of Thalassiosira pseudonana photosynthetic membranes to environmental temperature changes. In Proceedings of the 3rd World Congress on New Technologies (NewTech’17), Rome, Italy, 6–8 June 2017. [Google Scholar] [CrossRef]
- Dzikovski, B.; Freed, J. Membrane fluidity. In Encyclopedia of Biophysics; Roberts, G.C.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1440–1446. [Google Scholar]
- Los, D.A.; Murata, N. Regulation of enzymatic activity and gene expression by membrane fluidity. Sci. Signal. 2000, 2000, pe1. [Google Scholar] [CrossRef]
- Staykova, M.; Stone, H.A. The role of the membrane confinement in the surface area regulation of cells. Commun. Integr. Biol. 2011, 4, 616–618. [Google Scholar] [CrossRef]
- Fujii, S.; Nagata, N.; Masuda, T.; Wada, H.; Kobayashi, K. Galactolipids are essential for internal membrane transformation during Etioplast-to-Chloroplast Differentiation. Plant Cell Physiol. 2019, 60, 1224–1238. [Google Scholar] [CrossRef]
- Inaba, M.; Suzuki, I.; Szalontai, B.; Kanesaki, Y.; Los, D.A.; Hayashi, H.; Murata, N. Gene-engineered rigidification of membrane lipids enhances the cold inducibility of gene expression in Synechocystis. J. Biol. Chem. 2003, 278, 12191–12198. [Google Scholar] [CrossRef]
- Li, B.; Zhang, C.; Cao, B.; Qin, G.; Wang, W.; Tian, S. Brassinolide enhances cold stress tolerance of fruit by regulating plasma membrane proteins and lipids. Amino Acids 2012, 43, 2469–2480. [Google Scholar] [CrossRef] [PubMed]
- Dou, X.; Li, Y.; Han, J.; Zarlenga, D.; Zhu, W.; Ren, X.; Dong, N.; Li, X.; Li, G. Cholesterol of lipid rafts is a key determinant for entry and post-entry control of porcine rotavirus infection. BMC Veter Res. 2018, 14, 45. [Google Scholar] [CrossRef] [PubMed]
- Beck, J.G.; Mathieu, D.; Loudet, C.; Buchoux, S.; Dufourc, E.J. Plant sterols in “rafts”: A better way to regulate membrane thermal shocks. FASEB J. 2007, 21, 1714–1723. [Google Scholar] [CrossRef] [PubMed]
- Uemura, M.; Tominaga, Y.; Nakagawara, C.; Shigematsu, S.; Minami, A.; Kawamura, Y. Responses of the plasma membrane to low temperatures. Physiol. Plant. 2006, 126, 81–89. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acids [%mol] | Growth Temperature | |||||
|---|---|---|---|---|---|---|
| 20 °C | 5 °C | 27 °C | ||||
| Delisa | 522DK | Delisa | 522DK | Delisa | 522DK | |
| 10:0 | <LOD | 0.79 | 0.47 a | 0.50 a | <LOD | 0.35 |
| 12:0 | 4.00 a | 3.93 a | 4.49 a | 4.37 a | 4.12 b | 4.60 a |
| 14:0 | 0.28 a | 0.25 a | 0.23 a | 0.19 a | 0.29 a | 0.21 b |
| 16:0 | 16.54 b | 18.76 a | 16.16 a | 17.41 a | 17.20 a | 16.11 a |
| 16:1 | 6.29 a | 4.95 a | 4.25 a | 4.23 a | 6.45 a | 3.36 a |
| 18:0 | 3.53 a | 3.24 a | 3.37 a | 3.47 a | 2.96 a | 2.59 a |
| 18:1 Δ9 cis | 1.05 b | 1.84 a | 0.93 b | 1.15 a | 1.36 a | 1.54 a |
| 18:2 Δ6 cis | 4.24 a | 4.32 a | 3.05 a | 3.01 a | 6.85 b | 7.64 a |
| 18:3 (3) | 63.87 a | 61.85 a | 66.45 a | 65.33 a | 60.43 a | 63.28 a |
| 20:1 | 0.19 a | 0.06 a | 0.34 a | 0.34 a | 0.34 a | 0.31 a |
| 18:3/18:2 | 15.09 a | 14.38 a | 22.29 a | 21.70 a | 8.84 a | 8.29 a |
| U/S | 3.11 a | 2.72 b | 3.01 a | 2.86 a | 3.08 a | 3.20 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadura, I.; Latowski, D.; Oklestkova, J.; Gruszka, D.; Chyc, M.; Janeczko, A. Molecular Dynamics of Chloroplast Membranes Isolated from Wild-Type Barley and a Brassinosteroid-Deficient Mutant Acclimated to Low and High Temperatures. Biomolecules 2021, 11, 27. https://doi.org/10.3390/biom11010027
Sadura I, Latowski D, Oklestkova J, Gruszka D, Chyc M, Janeczko A. Molecular Dynamics of Chloroplast Membranes Isolated from Wild-Type Barley and a Brassinosteroid-Deficient Mutant Acclimated to Low and High Temperatures. Biomolecules. 2021; 11(1):27. https://doi.org/10.3390/biom11010027
Chicago/Turabian StyleSadura, Iwona, Dariusz Latowski, Jana Oklestkova, Damian Gruszka, Marek Chyc, and Anna Janeczko. 2021. "Molecular Dynamics of Chloroplast Membranes Isolated from Wild-Type Barley and a Brassinosteroid-Deficient Mutant Acclimated to Low and High Temperatures" Biomolecules 11, no. 1: 27. https://doi.org/10.3390/biom11010027
APA StyleSadura, I., Latowski, D., Oklestkova, J., Gruszka, D., Chyc, M., & Janeczko, A. (2021). Molecular Dynamics of Chloroplast Membranes Isolated from Wild-Type Barley and a Brassinosteroid-Deficient Mutant Acclimated to Low and High Temperatures. Biomolecules, 11(1), 27. https://doi.org/10.3390/biom11010027