Targeting MHC Regulation Using Polycyclic Polyprenylated Acylphloroglucinols Isolated from Garcinia bancana

 , , , , and
, , , , and 
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Reagents for Biological Assays
2.2. Plant Material
2.3. Extraction and Purification
2.4. GX Analysis and 2–3 Purity
2.5. NMR Experiments
2.6. Cell Culture and Treatments
2.7. Cell Immunostaining and Flow Cytometry
2.8. Cellular Viability Assay
2.9. Quantitative Real-Time RT-PCR
2.10. Western Blot Analysis
2.11. Data and Statistical Analysis
2.12. Molecular Docking
- Co-crystallized extracted ligand from the complex structure with no changes in conformation and configuration: the ligand (after hydrogens were added) was saved as a mol2 file without the presence of the protein and any further modification.
- Energy-minimized co-crystallized ligand extracted from the complex: the mol2 ligand structure was energetically minimized with the built-in MMFF94 function of LigandScout 4.4 and saved as another mol2 file.
- Energy-minimized reconstructed ligand: the structure of the ligand was rebuilt with ChemDraw Professional 16.0 [25], its 3D structure was then visualized from the SMILES code in the ligand-based view of LigandScout 4.4, after which the energy minimization step was carried out as described above. The fully processed ligand structure was output and saved as a mol2 file.
3. Results
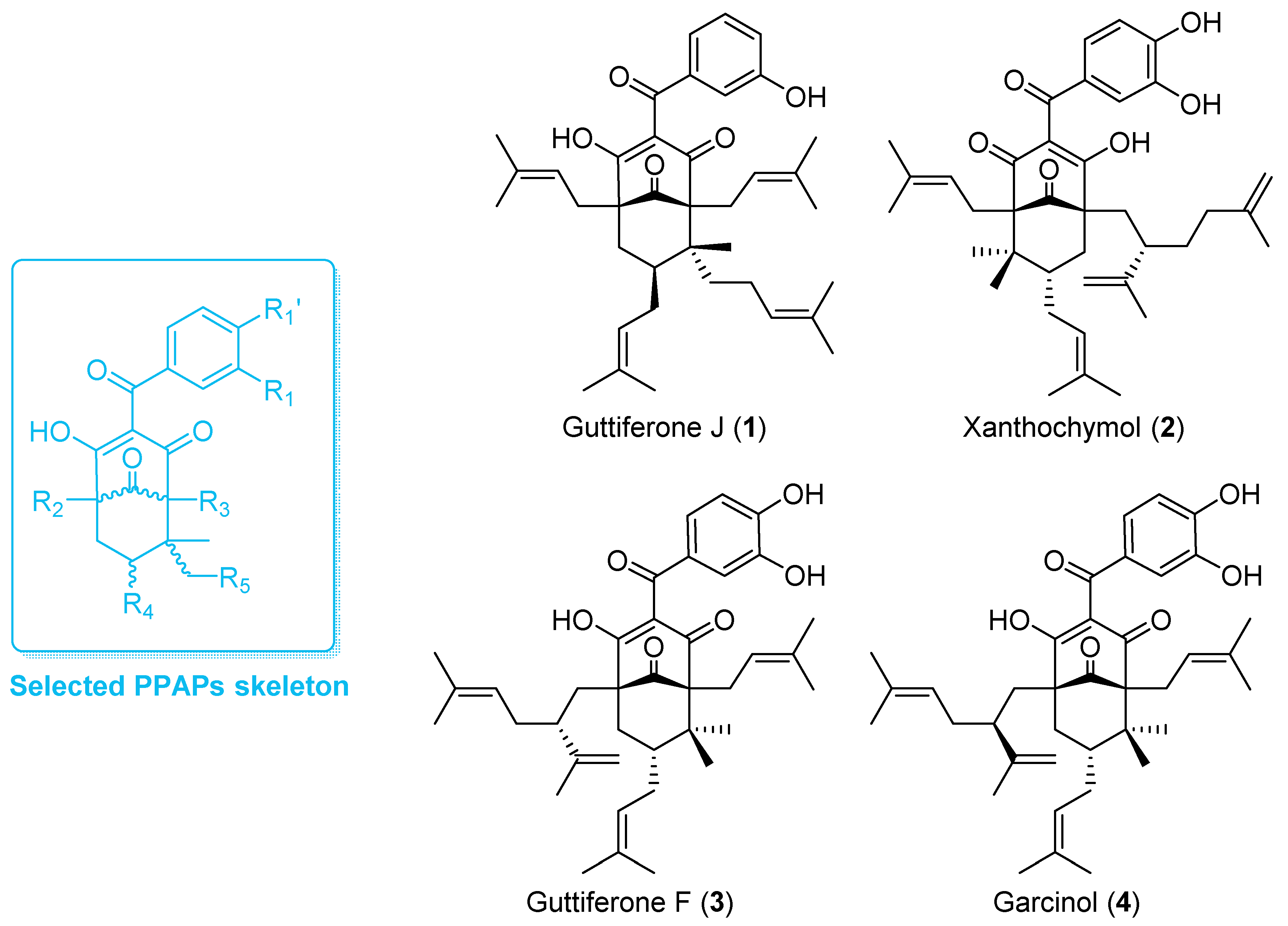
3.1. Phytochemical Analysis
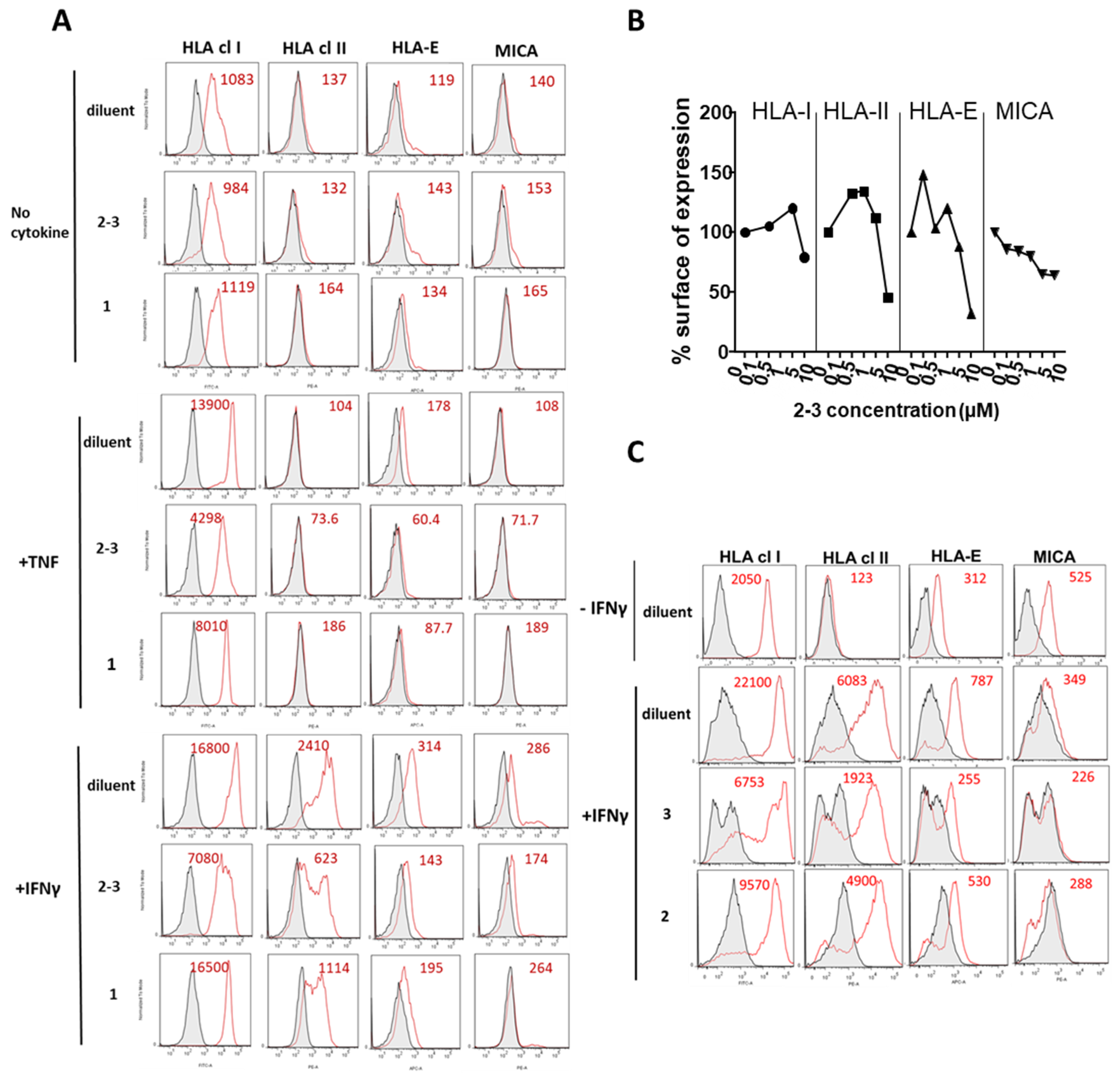
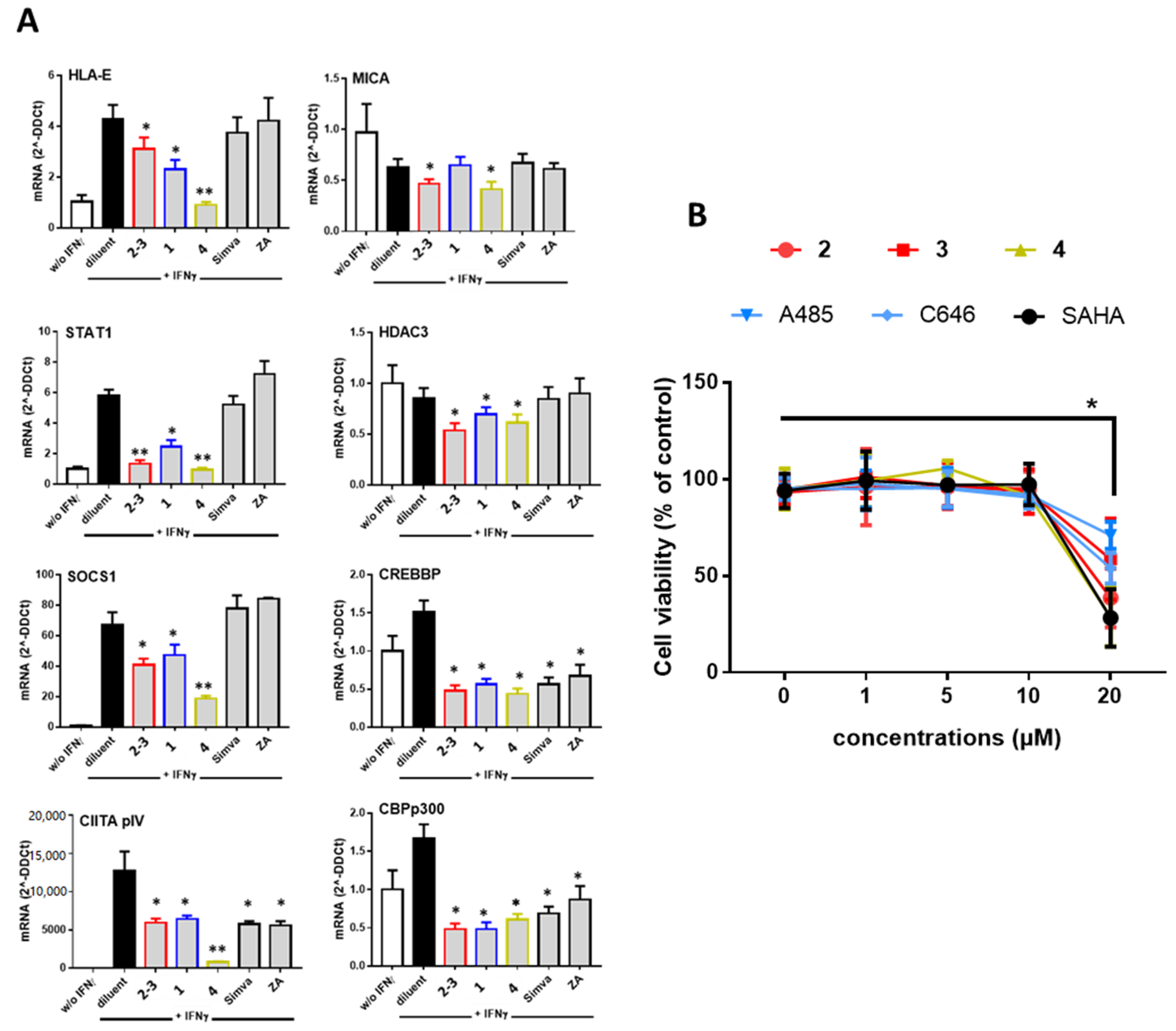
3.2. Comparative Inhibition of MHC Molecules Mediated by Guttiferones J (1) and F (3) and Xanthochymol (2)
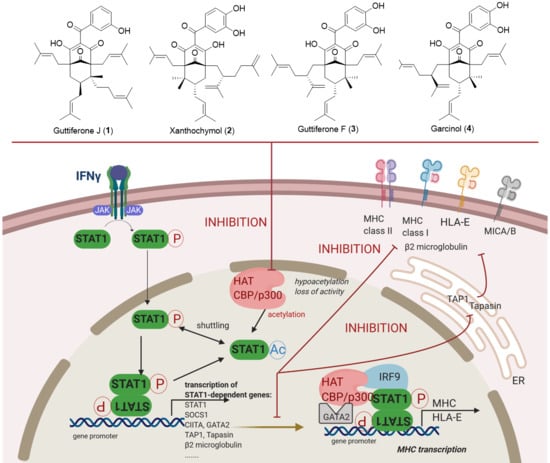
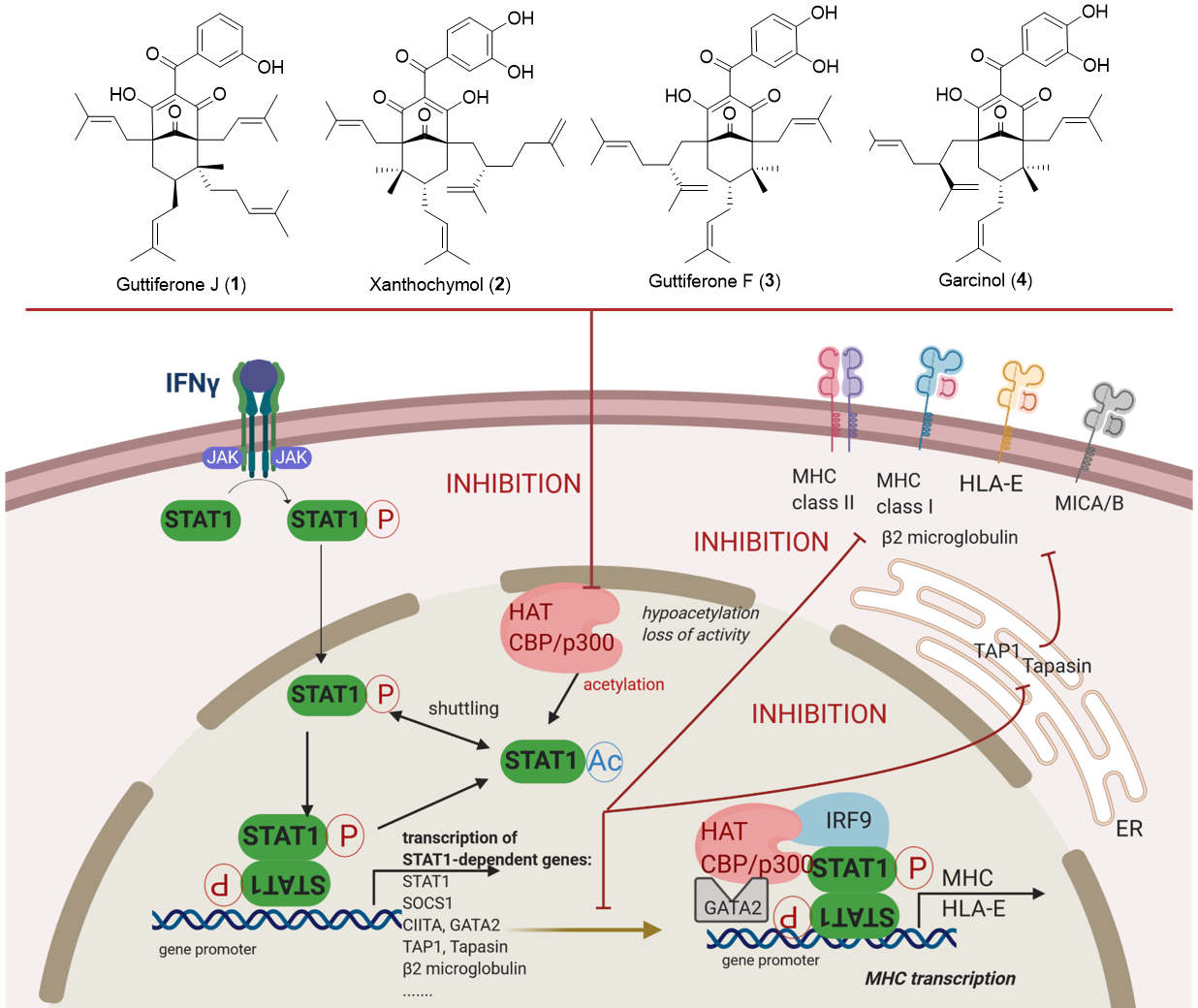
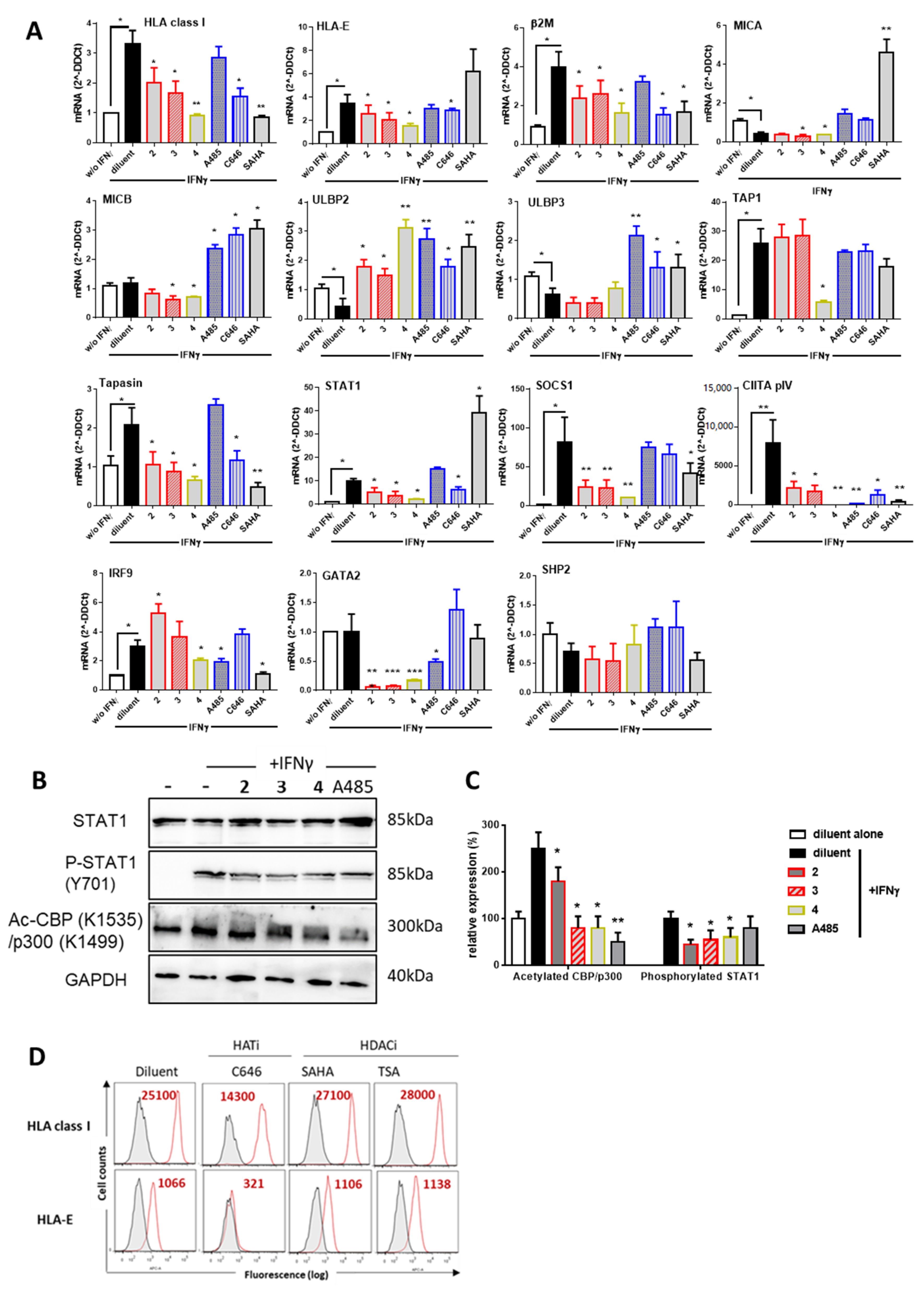
3.3. Guttiferones J (1) and F (3) and Xanthochymol (2) Are Novel Inhibitors of HAT CBP/p300 Activity, Which Impair IFNγ Signaling and Ultimately MHC Expression through the Inhibition of STAT1 Transcriptional Activities
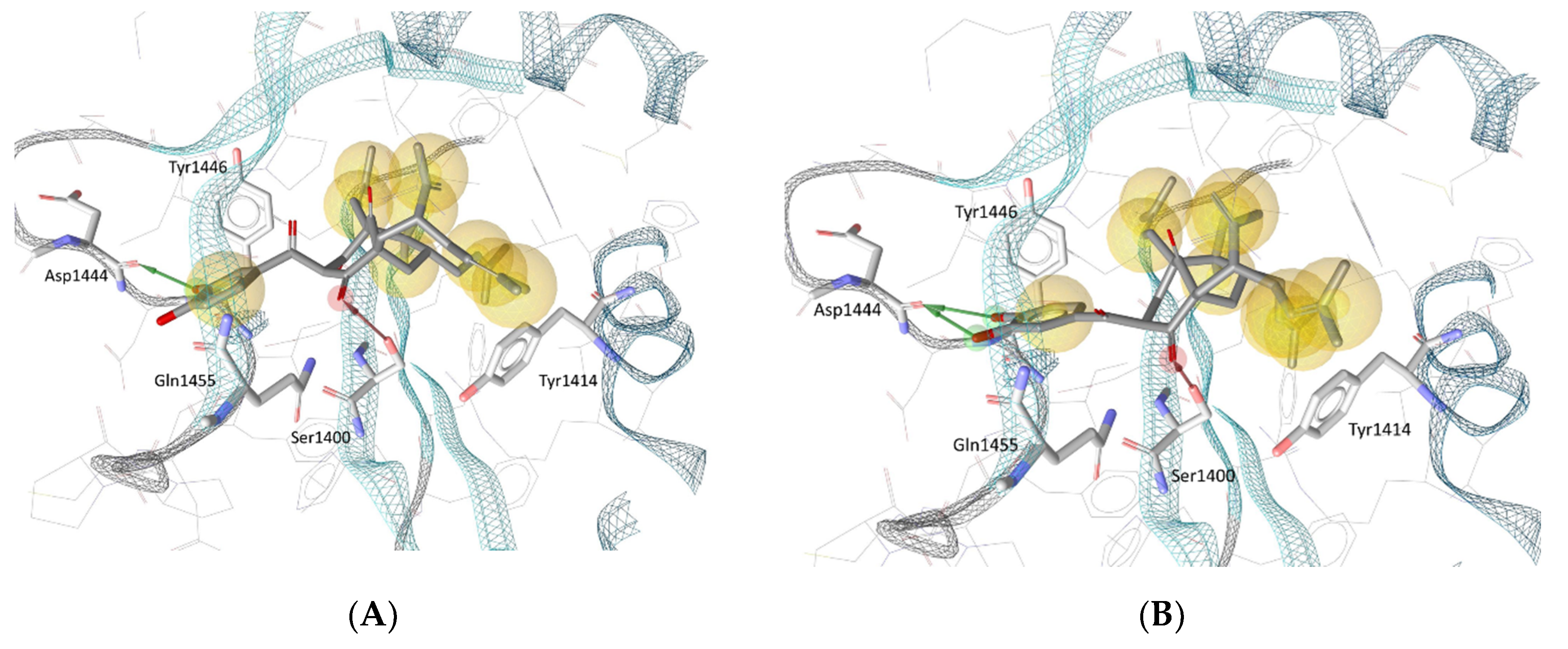
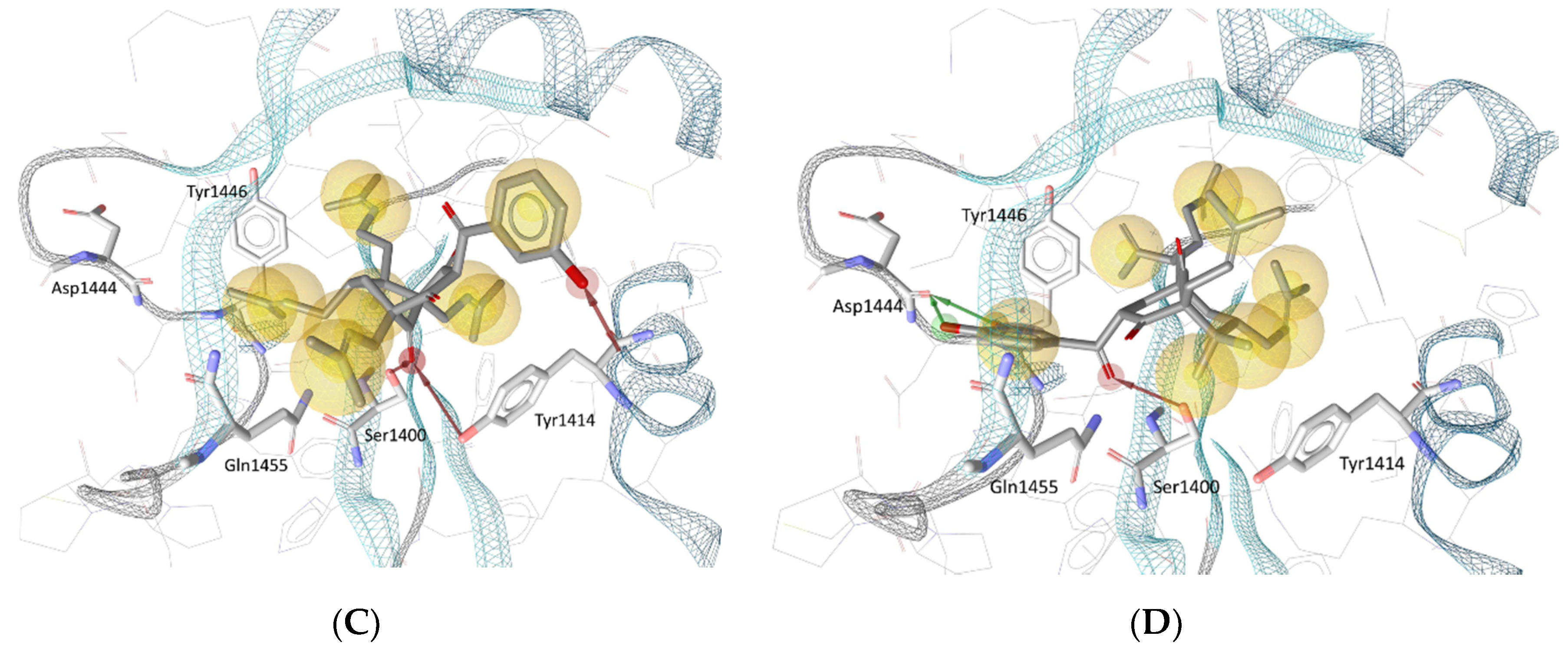
3.4. Molecular Docking in the Binding Site of Histone Acetyltransferase p300
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Janeway, C.A. How the immune system protects the host from infection. Microbes Infect. 2001, 3, 1167–1171. [Google Scholar] [CrossRef]
- Chaplin, D.D. Overview of the immune response. J. Allergy Clin. Immunol. 2010, 125, S3–S23. [Google Scholar] [CrossRef] [PubMed]
- Castro, F.; Cardoso, A.P.; Goncalves, R.M.; Serre, K.; Oliveira, M.J. Interferon-Gamma at the Crossroads of Tumor Immune Surveillance or Evasion. Front. Immunol. 2018, 9, 847. [Google Scholar] [CrossRef] [PubMed]
- Devaiah, B.N.; Singer, D.S. CIITA and Its Dual Roles in MHC Gene Transcription. Front. Immunol. 2013, 4, 476. [Google Scholar] [CrossRef] [PubMed]
- Gavlovsky, P.-J.; Tonnerre, P.; Guitton, C.; Charreau, B. Expression of MHC class I-related molecules MICA, HLA-E and EPCR shape endothelial cells with unique functions in innate and adaptive immunity. Hum. Immunol. 2016, 77, 1084–1091. [Google Scholar] [CrossRef]
- González, S.; Groh, V.; Spies, T. Immunobiology of Human NKG2D and Its Ligands. Curr. Top. Microbiol. Immunol. 2006, 298, 121–138. [Google Scholar] [CrossRef]
- Nausch, N.; Cerwenka, A. NKG2D ligands in tumor immunity. Oncogene 2008, 27, 5944–5958. [Google Scholar] [CrossRef]
- Jia, Y.; Pang, C.; Zhao, K.; Jiang, J.; Zhang, T.; Peng, J.; Sun, P.; Qian, Y. Garcinol Suppresses IL-1beta-Induced Chondrocyte Inflammation and Osteoarthritis via Inhibition of the NF-kappaB Signaling Pathway. Inflammation 2019, 42, 1754–1766. [Google Scholar] [CrossRef]
- Coupel, S.; Moreau, A.; Hamidou, M.; Horejsi, V.; Soulillou, J.P.; Charreau, B. Expression and release of soluble HLA-E is an immunoregulatory feature of endothelial cell activation. Blood 2007, 109, 2806–2814. [Google Scholar] [CrossRef] [PubMed]
- Jouand, N.; Bressollette-Bodin, C.; Gerard, N.; Giral, M.; Guerif, P.; Rodallec, A.; Oger, R.; Parrot, T.; Allard, M.; Cesbron-Gautier, A.; et al. HCMV triggers frequent and persistent UL40-specific unconventional HLA-E-restricted CD8 T-cell responses with potential autologous and allogeneic peptide recognition. PLoS Pathog. 2018, 14, e1007041. [Google Scholar] [CrossRef] [PubMed]
- Morvan, M.; Lanier, L.L. NK cells and cancer: You can teach innate cells new tricks. Nat. Rev. Cancer 2015, 16, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Bluestone, J.A. Mechanisms of tolerance. Immunol. Rev. 2011, 241, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Schmiedel, D.; Mandelboim, O. NKG2D Ligands-Critical Targets for Cancer Immune Escape and Therapy. Front. Immunol. 2018, 9, 2040. [Google Scholar] [CrossRef] [PubMed]
- Van Hall, T.; Andre, P.; Horowitz, A.; Ruan, D.F.; Borst, L.; Zerbib, R.; Narni-Mancinelli, E.; van der Burg, S.H.; Vivier, E. Monalizumab: Inhibiting the novel immune checkpoint NKG2A. J. Immunother. Cancer 2019, 7, 263. [Google Scholar] [CrossRef] [PubMed]
- Zingoni, A.; Molfetta, R.; Fionda, C.; Soriani, A.; Paolini, R.; Cippitelli, M.; Cerboni, C.; Santoni, A. NKG2D and Its Ligands: “One for All, All for One”. Front. Immunol. 2018, 9, 476. [Google Scholar] [CrossRef]
- Merza, J.; Mallet, S.; Litaudon, M.; Dumontet, V.; Seraphin, D.; Richomme, P. New cytotoxic guttiferone analogues from Garcinia virgata from New Caledonia. Planta Med. 2006, 72, 87–89. [Google Scholar]
- Rouger, C.; Pagie, S.; Derbré, S.; Le Ray, A.-M.; Richomme, P.; Charreau, B. Prenylated Polyphenols from Clusiaceae and Calophyllaceae with Immunomodulatory Activity on Endothelial Cells. PLoS ONE 2016, 11, e0167361. [Google Scholar] [CrossRef]
- Iinuma, M.; Tosa, H.; Tanaka, T.; Asai, F.; Kobayashl, Y.; Shimano, R.; Miyauchi, K.-I. Antibacterial Activity of Xanthones from Guttiferaeous Plants against Methicillin-resistant Staphylococcus aureus. J. Pharm. Pharmacol. 1996, 48, 861–865. [Google Scholar] [CrossRef]
- Fuller, R.W.; Blunt, J.W.; Boswell, J.L.; Cardellina, J.H.; Boyd, M.R. Guttiferone F, the First Prenylated Benzophenone fromAllanblackia stuhlmannii1. J. Nat. Prod. 1999, 62, 130–132. [Google Scholar] [CrossRef]
- Tonnerre, P.; Gerard, N.; Chatelais, M.; Charreau, B. MICA Gene Polymorphism in Kidney Allografts and Possible Impact of Functionally Relevant Variants. Transplant. Proc. 2010, 42, 4318–4321. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Battistuz, T.; Bhat, T.N.; Bluhm, W.F.; Bourne, P.E.; Burkhardt, K.; Feng, Z.; Gilliland, G.L.; Iype, L.; Jain, S.; et al. The Protein Data Bank. Acta Crystallogr. Sect. D Biol. Crystallogr. 2002, 58, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Verdonk, M.L.; Cole, J.C.; Hartshorn, M.J.; Murray, C.W.; Taylor, R.D. Improved protein-ligand docking using GOLD. Proteins: Struct. Funct. Bioinform. 2003, 52, 609–623. [Google Scholar] [CrossRef] [PubMed]
- Wolber, G.; Langer, T. LigandScout: 3-D Pharmacophores Derived from Protein-Bound Lingands and Their Use as Virtual Screening Filters. J. Chem. Inf. Model. 2005, 36, 160–169. [Google Scholar] [CrossRef]
- Chatterjee, A.; Yasmin, T.; Bagchi, D.; Stohs, S.J. The bactericidal effects of Lactobacillus acidophilus, garcinol and Protykin compared to clarithromycin, on Helicobacter pylori. Mol. Cell. Biochem. 2003, 243, 29–35. [Google Scholar] [CrossRef]
- Al-Qahtani, K.; Jabeen, B.; Sekirnik, R.; Riaz, N.; Claridge, T.D.W.; Schofield, C.J.; Mccullagh, J.S. The broad spectrum 2-oxoglutarate oxygenase inhibitor N-oxalylglycine is present in rhubarb and spinach leaves. Phytochemistry 2015, 117, 456–461. [Google Scholar] [CrossRef]
- Yang, X.-W.; Grossman, R.B.; Xu, G. Research Progress of Polycyclic Polyprenylated Acylphloroglucinols. Chem. Rev. 2018, 118, 3508–3558. [Google Scholar] [CrossRef]
- KewScience, Plants of the World Online. Garcinia L. 2020. Available online: http://plantsoftheworldonline.org/taxon/urn:lsid:ipni.org:names:19345-1 (accessed on 6 January 2020).
- Tropicos, Missouri Botanical Garden. 2020. Available online: https://www.tropicos.org (accessed on 6 January 2020).
- Bruguiere, A.; Derbre, S.; Coste, C.; Le Bot, M.; Siegler, B.; Leong, S.T.; Sulaiman, S.N.; Awang, K.; Richomme, P. (13)C-NMR dereplication of Garcinia extracts: Predicted chemical shifts as reliable databases. Fitoterapia 2018, 131, 59–64. [Google Scholar] [CrossRef]
- Hubert, J.; Nuzillard, J.M.; Purson, S.; Hamzaoui, M.; Borie, N.; Reynaud, R.; Renault, J.H. Identification of natural metabolites in mixture: A pattern recognition strategy based on (13)C NMR. Anal. Chem. 2014, 86, 2955–2962. [Google Scholar] [CrossRef]
- Li, J.; Gao, R.; Zhao, D.; Huang, X.; Chen, Y.; Gan, F.; Liu, H.; Yang, G. Separation and preparation of xanthochymol and guttiferone E by high performance liquid chromatography and high speed counter-current chromatography combined with silver nitrate coordination reaction. J. Chromatogr. A 2017, 1511, 143–148. [Google Scholar] [CrossRef]
- Roux, D.; Hadi, H.A.; Thoret, S.; Guenard, D.; Thoison, O.; Pais, M.; Sevenet, T. Structure-activity relationship of polyisoprenyl benzophenones from Garcinia pyrifera on the tubulin/microtubule system. J. Nat. Prod. 2000, 63, 1070–1076. [Google Scholar] [CrossRef] [PubMed]
- Coupel, S.; Leboeuf, F.; Boulday, G.; Soulillou, J.-P.; Charreau, B. RhoA Activation Mediates Phosphatidylinositol 3-Kinase-Dependent Proliferation of Human Vascular Endothelial Cells: An Alloimmune Mechanism of Chronic Allograft Nephropathy. J. Am. Soc. Nephrol. 2004, 15, 2429–2439. [Google Scholar] [CrossRef] [PubMed]
- Chauveau, A.; Tonnerre, P.; Pabois, A.; Gavlovsky, P.J.; Chatelais, M.; Coupel, S.; Charreau, B. Endothelial cell activation and proliferation modulate NKG2D activity by regulating MICA expression and shedding. J. Innate Immun. 2014, 6, 89–104. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanyam, K.; Altaf, M.; Varier, R.A.; Swaminathan, V.; Ravindran, A.; Sadhale, P.P.; Kundu, T.K. Polyisoprenylated benzophenone, garcinol, a natural histone acetyltransferase inhibitor, represses chromatin transcription and alters global gene expression. J. Biol. Chem. 2004, 279, 33716–33726. [Google Scholar] [CrossRef] [PubMed]
- Lasko, L.M.; Jakob, C.G.; Edalji, R.P.; Qiu, W.; Montgomery, D.; Digiammarino, E.L.; Hansen, T.M.; Risi, R.M.; Frey, R. Discovery of a selective catalytic p300/CBP inhibitor that targets lineage-specific tumours. Nature 2017, 550, 128–132. [Google Scholar] [CrossRef]
- Schindler, C.; Plumlee, C. Inteferons pen the JAK–STAT pathway. Semin. Cell Dev. Boil. 2008, 19, 311–318. [Google Scholar] [CrossRef]
- Hu, X.; Ivashkiv, L.B. Cross-regulation of signaling pathways by interferon-gamma: Implications for immune responses and autoimmune diseases. Immunity 2009, 31, 539–550. [Google Scholar] [CrossRef]
- Satoh, J.; Tabunoki, H. A Comprehensive Profile of ChIP-Seq-Based STAT1 Target Genes Suggests the Complexity of STAT1-Mediated Gene Regulatory Mechanisms. Gene Regul. Syst. Biol. 2013, 7, 41–56. [Google Scholar] [CrossRef]
- O’Shea, J.J.; Murray, P.J. Cytokine signaling modules in inflammatory responses. Immunity 2008, 28, 477–487. [Google Scholar] [CrossRef]
- Thompson, P.R.; Wang, D.; Wang, L.; Fulco, M.; Pediconi, N.; Zhang, D.; An, W.; Ge, Q.; Roeder, R.G.; Wong, J.; et al. Regulation of the p300 HAT domain via a novel activation loop. Nat. Struct. Mol. Biol. 2004, 11, 308–315. [Google Scholar] [CrossRef]
- Kramer, O.H.; Heinzel, T. Phosphorylation-acetylation switch in the regulation of STAT1 signaling. Mol. Cell. Endocrinol. 2010, 315, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, S. Regulation of STAT signaling by acetylation. Cell. Signal. 2013, 25, 1924–1931. [Google Scholar] [CrossRef] [PubMed]
- Min, W.; Pober, J.S.; Johnson, D.R. Kinetically coordinated induction of TAP1 and HLA class I by IFN-gamma: The rapid induction of TAP1 by IFN-gamma is mediated by Stat1 alpha. J. Immunol. 1996, 156, 3174–3183. [Google Scholar]
- Herberg, J.A.; Sgouros, J.; Jones, T.; Copeman, J.; Humphray, S.J.; Sheer, D.; Cresswell, P.; Beck, S.; Trowsdale, J. Genomic analysis of the Tapasin gene, located close to the TAP loci in the MHC. Eur. J. Immunol. 1998, 28, 459–467. [Google Scholar] [CrossRef]
- Van den Elsen, P.J.; Holling, T.M.; Kuipers, H.F.; van der Stoep, N. Transcriptional regulation of antigen presentation. Curr. Opin. Immunol. 2004, 16, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Leibowitz, M.S.; Srivastava, R.M.; Andrade Filho, P.A.; Egloff, A.M.; Wang, L.; Seethala, R.R.; Ferrone, S.; Ferris, R.L. SHP2 is overexpressed and inhibits pSTAT1-mediated APM component expression, T-cell attracting chemokine secretion, and CTL recognition in head and neck cancer cells. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2013, 19, 798–808. [Google Scholar] [CrossRef]
- Spilianakis, C.; Kretsovali, A.; Agalioti, T.; Makatounakis, T.; Thanos, D.; Papamatheakis, J. CIITA regulates transcription onset viaSer5-phosphorylation of RNA Pol II. EMBO J. 2003, 22, 5125–5136. [Google Scholar] [CrossRef]
- Oike, T.; Ogiwara, H.; Torikai, K.; Nakano, T.; Yokota, J.; Kohno, T. Garcinol, a histone acetyltransferase inhibitor, radiosensitizes cancer cells by inhibiting non-homologous end joining. Int. J. Radiat. Oncol. Biol. Phys. 2012, 84, 815–821. [Google Scholar] [CrossRef]
- Wang, B.; Lin, L.; Ai, Q.; Zeng, T.; Ge, P.; Zhang, L. HAT inhibitor, garcinol, exacerbates lipopolysaccharideinduced inflammation in vitro and in vivo. Mol. Med. Rep. 2016, 13, 5290–5296. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, M.; Zheng, D.; Zhang, H.; Lv, Y.; Zhang, L.; Tan, H.S.; Zhou, H.; Lao, Y.Z.; Xu, H.X. Garcinol inhibits esophageal cancer metastasis by suppressing the p300 and TGF-beta1 signaling pathways. Acta Pharmacol. Sin. 2020, 41, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Chan, H.M.; La Thangue, N.B. p300/CBP proteins: HATs for transcriptional bridges and scaffolds. J. Cell Sci. 2001, 114, 2363–2373. [Google Scholar] [PubMed]
- Celik, A.A.; Kraemer, T.; Huyton, T.; Blasczyk, R.; Bade-Doding, C. The diversity of the HLA-E-restricted peptide repertoire explains the immunological impact of the Arg107Gly mismatch. Immunogenetics 2016, 68, 29–41. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coste, C.; Gérard, N.; Dinh, C.P.; Bruguière, A.; Rouger, C.; Leong, S.T.; Awang, K.; Richomme, P.; Derbré, S.; Charreau, B. Targeting MHC Regulation Using Polycyclic Polyprenylated Acylphloroglucinols Isolated from Garcinia bancana. Biomolecules 2020, 10, 1266. https://doi.org/10.3390/biom10091266
Coste C, Gérard N, Dinh CP, Bruguière A, Rouger C, Leong ST, Awang K, Richomme P, Derbré S, Charreau B. Targeting MHC Regulation Using Polycyclic Polyprenylated Acylphloroglucinols Isolated from Garcinia bancana. Biomolecules. 2020; 10(9):1266. https://doi.org/10.3390/biom10091266
Chicago/Turabian StyleCoste, Chloé, Nathalie Gérard, Chau Phi Dinh, Antoine Bruguière, Caroline Rouger, Sow Tein Leong, Khalijah Awang, Pascal Richomme, Séverine Derbré, and Béatrice Charreau. 2020. "Targeting MHC Regulation Using Polycyclic Polyprenylated Acylphloroglucinols Isolated from Garcinia bancana" Biomolecules 10, no. 9: 1266. https://doi.org/10.3390/biom10091266
APA StyleCoste, C., Gérard, N., Dinh, C. P., Bruguière, A., Rouger, C., Leong, S. T., Awang, K., Richomme, P., Derbré, S., & Charreau, B. (2020). Targeting MHC Regulation Using Polycyclic Polyprenylated Acylphloroglucinols Isolated from Garcinia bancana. Biomolecules, 10(9), 1266. https://doi.org/10.3390/biom10091266