α-Glucosidase Inhibitory Activity of Tannat Grape Phenolic Extracts in Relation to Their Ripening Stages


 , and
, and
Abstract
1. Introduction
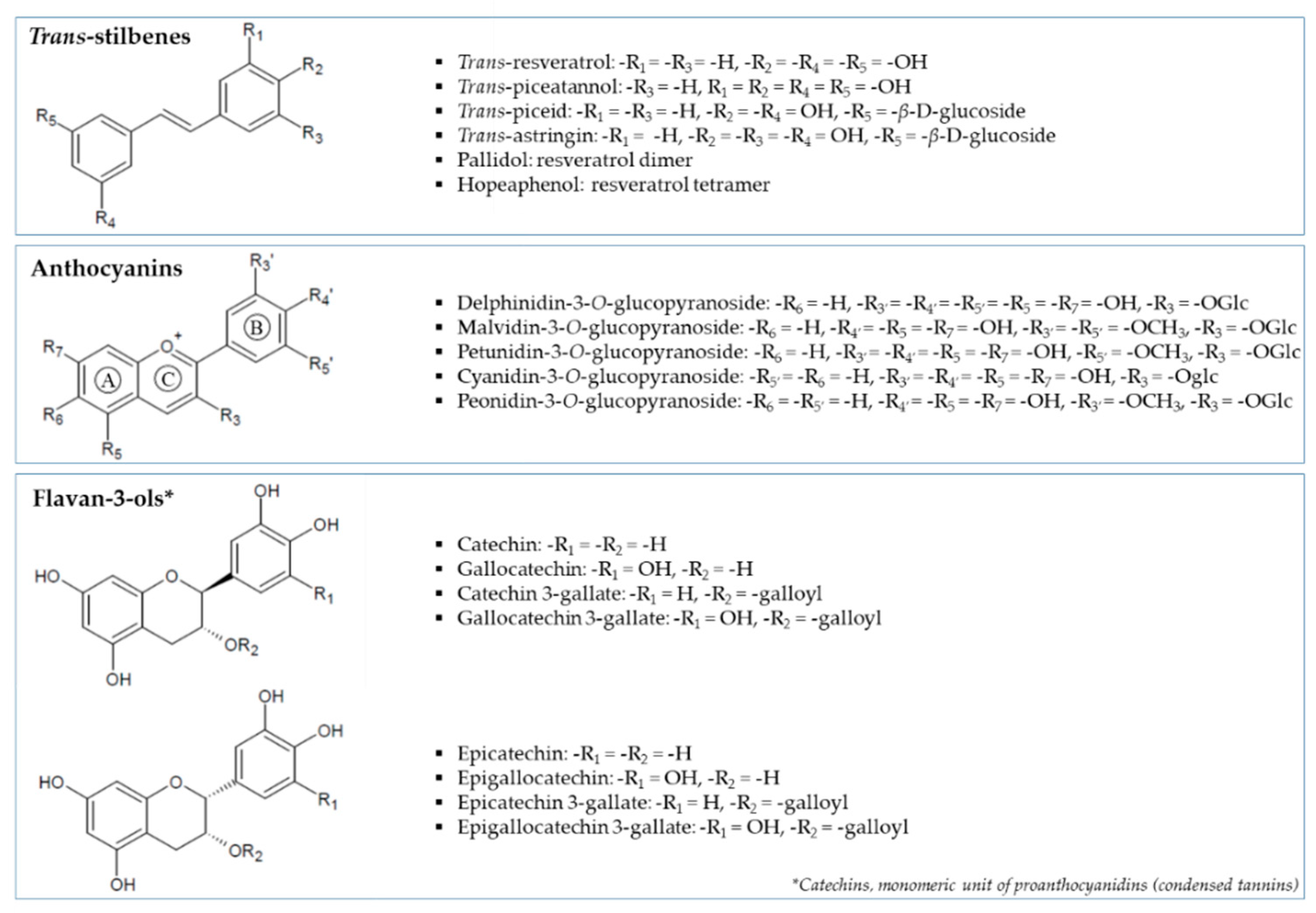
- To measure the stilbene, anthocyanin and flavan-3-ol content at the different ripening stages.
- To study the α-glucosidase inhibitory activity of seed and skin Tannat grape extracts at four ripening stages.
- To study the type of enzymatic inhibition by determining Lineweaver-Burk plots and kinetics constants calculations.
2. Materials and Methods
2.1. Materials
2.1.1. Chemicals and Reagents
2.1.2. Fruit Materials
2.2. Methods
2.2.1. Polyphenol Extraction
2.2.2. Ultra High Performance Liquid Chromatography-Mass Spectrometry (UHPLC-MS) Analysis of Stilbenes
2.2.3. UPLC-Photodiode Array (PDA)Analysis of Anthocyanins
2.2.4. UPLC-PDA Analysis of Flavan-3-ols
2.2.5. TPC Determination
2.2.6. α-Glucosidase Inhibition Assays
Set-up of the α-Glucosidase Inhibition Assays in Microplate Format
Data Interpretation
Application of the Microplate Assays to Kinetic Studies of α-Glucosidase Inhibition
2.2.7. Statistical Analysis
3. Results and Discussion
3.1. Evolution of Phenolic Composition in Tannat Grape during Maturation
3.1.1. TPC Analysis
3.1.2. Total Stilbene Content
3.1.3. Total Flavan-3-ol Content
3.1.4. Total Anthocyanin Content
3.2. Optimization and Performances of α-Glucosidase Inhibition Microplate Assays
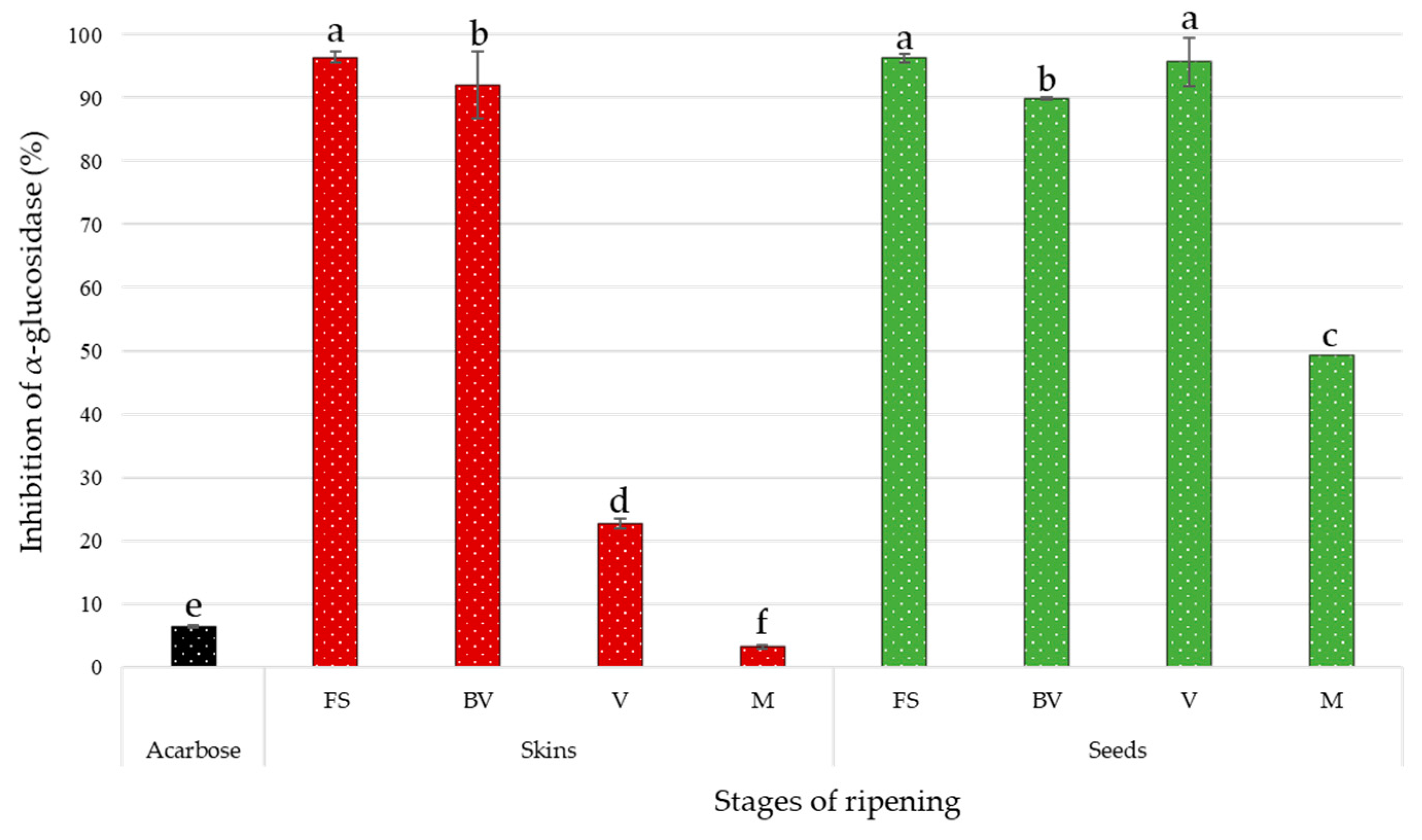
3.3. α-Glucosidase Inhibitory Activity of Grape Extracts
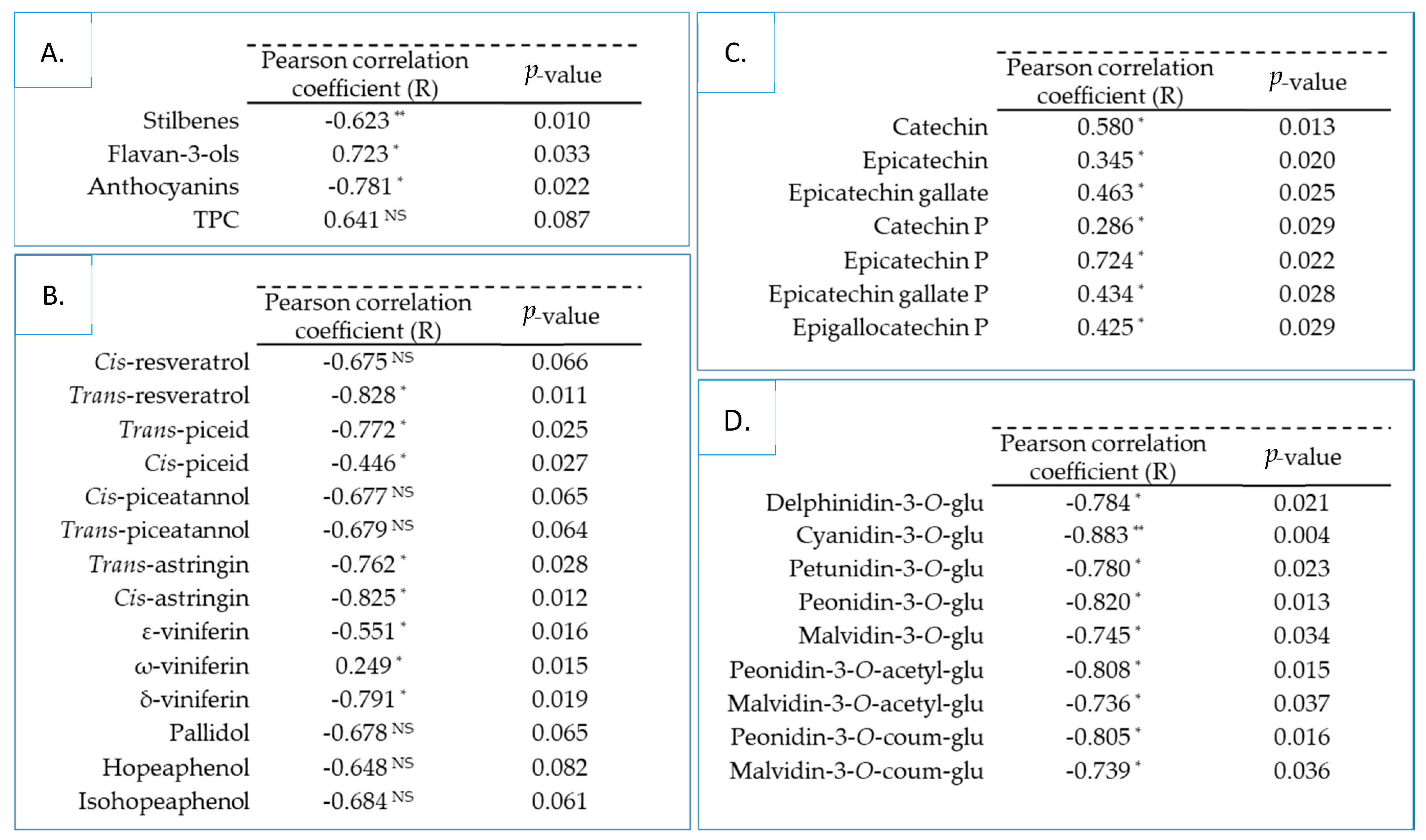
3.4. Correlation between Chemical Composition and α-Glucosidase Inhibitory Activity
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Federation International Diabetes IDF Diabetes Atlas Ninth. 2019. Available online: https://www.diabetesatlas.org/upload/resources/2019/IDF_Atlas_9th_Edition_2019.pdf (accessed on 16 July 2020).
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Res. Clin. Pract. 2019. [Google Scholar] [CrossRef] [PubMed]
- Hedrington, M.S.; Davis, S.N. Considerations when using alpha-glucosidase inhibitors in the treatment of type 2 diabetes. Expert Opin. Pharmacother. 2019, 20, 2229–2235. [Google Scholar] [CrossRef] [PubMed]
- Li, A.N.; Li, S.; Zhang, Y.J.; Xu, X.R.; Chen, Y.M.; Li, H. Bin Resources and biological activities of natural polyphenols. Nutrients 2014, 6, 6020–6047. [Google Scholar] [CrossRef]
- Kerem, Z.; Bilkis, I.; Flaishman, M.A.; Sivan, L. Antioxidant activity and inhibition of α-glucosidase by trans-resveratrol, piceid, and a novel trans-stilbene from the roots of Israeli Rumex bucephalophorus L. J. Agric. Food Chem. 2006, 54, 1243–1247. [Google Scholar] [CrossRef] [PubMed]
- Akkarachiyasit, S.; Charoenlertkul, P.; Yibchok-Anun, S.; Adisakwattana, S. Inhibitory activities of cyanidin and its glycosides and synergistic effect with acarbose against intestinal α-glucosidase and pancreatic α-amylase. Int. J. Mol. Sci. 2010, 11, 3387–3396. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, D.; Konishi, T. Anthocyanins and anthocyanin-rich extracts.pdf. Asia Pac. J. Clin. Nutr 2007, 16, 200–208. [Google Scholar] [CrossRef]
- Barik, S.K.; Russell, W.R.; Moar, K.M.; Cruickshank, M.; Scobbie, L.; Duncan, G.; Hoggard, N. The anthocyanins in black currants regulate postprandial hyperglycaemia primarily by inhibiting α-glucosidase while other phenolics modulate salivary α-amylase, glucose uptake and sugar transporters. J. Nutr. Biochem. 2020, 78, 108325. [Google Scholar] [CrossRef]
- Ochir, S.; Nishizawa, M.; Jae Park, B.; Ishii, K.; Kanazawa, T.; Funaki, M.; Yamagishi, T. Inhibitory effects of Rosa gallica on the digestive enzymes. J. Nat. Med. 2010, 64, 275–280. [Google Scholar] [CrossRef]
- Xiao, J.; Kai, G.; Yamamoto, K.; Chen, X. Advance in Dietary Polyphenols as α-Glucosidases Inhibitors: A Review on Structure-Activity Relationship Aspect. Crit. Rev. Food Sci. Nutr. 2013, 53, 818–836. [Google Scholar] [CrossRef]
- Lorrain, B.; Ky, I.; Pechamat, L.; Teissedre, P.L. Evolution of analysis of polyhenols from grapes, wines, and extracts. Molecules 2013, 18, 1076–1100. [Google Scholar] [CrossRef]
- Kong, F.; Qin, Y.; Su, Z.; Ning, Z.; Yu, S. Optimization of Extraction of Hypoglycemic Ingredients from Grape Seeds and Evaluation of α-Glucosidase and α-Amylase Inhibitory Effects In Vitro. J. Food Sci. 2018, 83, 1422–1429. [Google Scholar] [CrossRef] [PubMed]
- Lavelli, V.; Sri Harsha, P.S.C.; Ferranti, P.; Scarafoni, A.; Iametti, S. Grape skin phenolics as inhibitors of mammalian α-glucosidase and α-amylase—Effect of food matrix and processing on efficacy. Food Funct. 2016, 7, 1655–1663. [Google Scholar] [CrossRef] [PubMed]
- Hogan, S.; Zhang, L.; Li, J.; Sun, S.; Canning, C.; Zhou, K. Antioxidant rich grape pomace extract suppresses postprandial hyperglycemia in diabetic mice by specifically inhibiting alpha-glucosidase. Nutr. Metab. 2010, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kadouh, H.C.; Sun, S.; Zhu, W.; Zhou, K. α-Glucosidase inhibiting activity and bioactive compounds of six red wine grape pomace extracts. J. Funct. Foods 2016, 26, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hogan, S.; Li, J.; Sun, S.; Canning, C.; Zheng, S.J.; Zhou, K. Grape skin extract inhibits mammalian intestinal α-glucosidase activity and suppresses postprandial glycemic response in streptozocin-treated mice. Food Chem. 2011, 126, 466–471. [Google Scholar] [CrossRef]
- Sun, S.; Kadouh, H.C.; Zhu, W.; Zhou, K. Bioactivity-guided isolation and purification of α-glucosidase inhibitor, 6-O-D-glycosides, from Tinta Cão grape pomace. J. Funct. Foods 2016, 23, 573–579. [Google Scholar] [CrossRef]
- Yilmazer-Musa, M.; Griffith, A.M.; Michels, A.J.; Schneider, E.; Frei, B. Grape seed and tea extracts and catechin 3-gallates are potent inhibitors of α-amylase and α-glucosidase activity. J. Agric. Food Chem. 2012, 60, 8924–8929. [Google Scholar] [CrossRef]
- Zhang, A.J.; Rimando, A.M.; Mizuno, C.S.; Mathews, S.T. α-Glucosidase inhibitory effect of resveratrol and piceatannol. J. Nutr. Biochem. 2017, 47, 86–93. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, L.; Dong, Y.; Fang, Z.; Nisar, T.; Zhao, T.; Wang, Z.-C.; Guo, Y. Chemical compositions and α-glucosidase inhibitory effects of anthocyanidins from blueberry, blackcurrant and blue honeysuckle fruits. Food Chem. 2019, 299, 125102. [Google Scholar] [CrossRef]
- Fernández-Fernández, A.M.; Iriondo-DeHond, A.; Dellacassa, E.; Medrano-Fernandez, A.; del Castillo, M.D. Assessment of antioxidant, antidiabetic, antiobesity, and anti-inflammatory properties of a Tannat winemaking by-product. Eur. Food Res. Technol. 2019, 245, 1539–1551. [Google Scholar] [CrossRef]
- Zerbib, M.; Mazauric, J.P.; Meudec, E.; Le Guernevé, C.; Lepak, A.; Nidetzky, B.; Cheynier, V.; Terrier, N.; Saucier, C. New flavanol O-glycosides in grape and wine. Food Chem. 2018, 266, 441–448. [Google Scholar] [CrossRef]
- Loupit, G.; Prigent, S.; Franc, C.; De Revel, G.; Richard, T.; Cookson, S.J.; Fonayet, J.V. Polyphenol Profiles of Just Pruned Grapevine Canes from Wild Vitis Accessions and Vitis vinifera Cultivars. J. Agric. Food Chem. 2020. [Google Scholar] [CrossRef]
- Pérez-Magariño, S.; González-San José, M.L. Evolution of Flavanols, Anthocyanins, and Their Derivatives during the Aging of Red Wines Elaborated from Grapes Harvested at Different Stages of Ripening. J. Agric. Food Chem. 2004, 52, 1181–1189. [Google Scholar] [CrossRef]
- Giuffrè, A.M. HPLC-DAD detection of changes in phenol content of red berry skins during grape ripening. Eur. Food Res. Technol. 2013, 237, 555–564. [Google Scholar] [CrossRef]
- Kennedy, J.A.; Jones, G.P.; Pa, P. Analysis of Proanthocyanidin Cleavage Products Following Acid-Catalysis in the Presence of Excess. J. Agric. Food Chem. 2001, 49, 1740–1746. [Google Scholar] [CrossRef]
- Rayess, Y.E.; Barbar, R.; Wilson, E.A.; Bouajila, J. Analytical methods for wine polyphenols analysis and for their antioxidant activity evaluation. Wine Phenolic Compos. Classif. Heal. Benefits 2014, 71–101. [Google Scholar]
- Pramod, M.; Gurdeep, S.; Neetesh, J.; Gupta, M.K. In-vitro studies on inhibition of alpha amylase and alpha glucosidase by plant extracts of alternanthera Pungens kunth. J. Drug Ther. 2019, 8, 64–68. [Google Scholar]
- European Medicines Agency (EMEA) Guideline on Bioanalytical Method Validation 2015. Available online: https://www.ema.europa.eu/en/documents/scientific-guideline/guideline-bioanalytical-method-validation_en.pdf (accessed on 16 July 2020).
- Xu, C.; Zhang, Y.; Cao, L.; Lu, J. Phenolic compounds and antioxidant properties of different grape cultivars grown in China. Food Chem. 2010, 119, 1557–1565. [Google Scholar] [CrossRef]
- Obreque-Slier, E.; Peña-Neira, Á.; López-Solís, R.; Cáceres-Mella, A.; Toledo-Araya, H.; López-Rivera, A. Phenolic composition of skins from four Carmenet grape varieties (Vitis vinifera L.) during ripening. LWTFood Sci. Technol. 2013, 54, 404–413. [Google Scholar] [CrossRef]
- Ky, I.; Teissedre, P.L. Characterisation of Mediterranean grape pomace seed and skin extracts: Polyphenolic content and antioxidant activity. Molecules 2015, 20, 2190–2207. [Google Scholar] [CrossRef]
- Pantelić, M.M.; Dabić Zagorac, D.Č.; Davidović, S.M.; Todić, S.R.; Bešlić, Z.S.; Gašić, U.M.; Tešić, Ž.L.; Natić, M.M. Identification and quantification of phenolic compounds in berry skin, pulp, and seeds in 13 grapevine varieties grown in Serbia. Food Chem. 2016, 211, 243–252. [Google Scholar] [CrossRef]
- Mulero, J.; Pardo, F.; Zafrilla, P. Antioxidant activity and phenolic composition of organic and conventional grapes and wines. J. Food Compos. Anal. 2010, 23, 569–574. [Google Scholar] [CrossRef]
- Danilewicz, J.C. Folin-Ciocalteu, FRAP, and DPPH• assays for measuring polyphenol concentration in white wine. Am. J. Enol. Vitic. 2015, 66, 463–471. [Google Scholar] [CrossRef]
- Gil-Muñoz, R.; Fernández-Fernández, J.I.; Crespo-Villegas, O.; Garde-Cerdán, T. Elicitors used as a tool to increase stilbenes in grapes and wines. Food Res. Int. 2017, 98, 34–39. [Google Scholar] [CrossRef]
- Gatto, P.; Vrhovsek, U.; Muth, J.; Segala, C.; Romualdi, C.; Fontana, P.; Pruefer, D.; Stefanini, M.; Moser, C.; Mattivi, F.; et al. Ripening and genotype control stilbene accumulation in healthy grapes. J. Agric. Food Chem. 2008, 56, 11773–11785. [Google Scholar] [CrossRef]
- Geana, E.I.; Dinca, O.R.; Ionete, R.E.; Artem, V.; Niculescu, V.C. Monitoring trans-resveratrol in grape berry skins during ripening and in corresponding wines by HPLC. Food Technol. Biotechnol. 2015, 53, 73–80. [Google Scholar] [CrossRef]
- Wang, J.-F.; Ma, L.; Xi, H.-F.; Wang, L.-J.; Li, S.-H. Resveratrol synthesis under natural conditions and after UV-C irradiation in berry skin is associated with berry development stages in ‘Beihong’ (V. vinifera × V. amurensis). Food Chem. 2015, 168, 430–438. [Google Scholar] [CrossRef]
- Guerrero, R.F.; Puertas, B.; Fernández, M.I.; Palma, M.; Cantos-Villar, E. Induction of stilbenes in grapes by UV-C: Comparison of different subspecies of Vitis. Innov. Food Sci. Emerg. Technol. 2010, 11, 231–238. [Google Scholar] [CrossRef]
- Goldberg, D.M.; Ng, E.; Yan, J.; Karumanchiri, A.; Soleas, G.J.; Diamandis, E.P. Regional differences in resveratrol isomer concentrations of wines from various cultivars. J. Wine Res. 1996, 21, 13–24. [Google Scholar] [CrossRef]
- Spranger, I.; Sun, B.; Mateus, A.M.; Freitas, V.d.; Ricardo-da-Silva, J.M. Chemical characterization and antioxidant activities of oligomeric and polymeric procyanidin fractions from grape seeds. Food Chem. 2008, 108, 519–532. [Google Scholar] [CrossRef]
- Lorrain, B.; Chira, K.; Teissedre, P.L. Phenolic composition of Merlot and Cabernet-Sauvignon grapes from Bordeaux vineyard for the 2009-vintage: Comparison to 2006, 2007 and 2008 vintages. Food Chem. 2011, 126, 1991–1999. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.O. Phenolics and ripening in grape berries. Am. J. Enol. Vitic. 2006, 57, 249–256. [Google Scholar]
- Obreque-Slier, E.; Peña-Neira, Á.; López-SolíS, R.; Zamora-MaríN, F.; Ricardo-Da Silva, J.M.; Laureano, O. Comparative study of the phenolic composition of seeds and skins from carménère and cabernet sauvignon grape varieties (Vitis vinifera L.) during ripening. J. Agric. Food Chem. 2010, 58, 3591–3599. [Google Scholar] [CrossRef]
- Downey, M.O.; Harvey, J.S.; Robinson, S.P. Analysis of tannins in seeds and skins of Shiraz grapes throughout berry development. Aust. J. Grape Wine Res. 2003, 9, 15–27. [Google Scholar] [CrossRef]
- Kennedy, J.A.; Troup, G.J.; Pilbrow, J.R.; Hutton, D.R.; Hewitt, D.; Hunter, C.R.; Ristic, R.; Iland, P.G.; Jones, G.P. Development of seed polyphenols in berries from Vitis vinifera L. cv. Shiraz. Aust. J. Grape Wine Res. 2000, 6, 244–254. [Google Scholar] [CrossRef]
- Tkacz, K.; Wojdyło, A.; Nowicka, P.; Turkiewicz, I.; Golis, T. Characterization in vitro potency of biological active fractions of seeds, skins and flesh from selected Vitis vinifera L. cultivars and interspecific hybrids. J. Funct. Foods 2019, 56, 353–363. [Google Scholar] [CrossRef]
- Torchio, F.; Cagnasso, E.; Gerbi, V.; Rolle, L. Mechanical properties, phenolic composition and extractability indices of Barbera grapes of different soluble solids contents from several growing areas. Anal. Chim. Acta 2010, 660, 183–189. [Google Scholar] [CrossRef]
- Boido, E.; García-Marino, M.; Dellacassa, E.; Carrau, F.; Rivas-Gonzalo, J.C.; Escribano-Bailón, M.T. Characterisation and evolution of grape polyphenol profiles of Vitis vinifera L. cv. Tannat during ripening and vinification. Aust. J. Grape Wine Res. 2011, 17, 383–393. [Google Scholar] [CrossRef]
- Fanzone, M.; Zamora, F.; Jofré, V.; Assof, M.; Peña-Neira, Á. Phenolic composition of Malbec grape skins and seeds from valle de Uco (Mendoza, Argentina) during ripening. effect of cluster thinning. J. Agric. Food Chem. 2011, 59, 6120–6136. [Google Scholar] [CrossRef]
- De Pascual-Teresa, S.; Sanchez-Ballesta, M.T. Anthocyanins: From plant to health. Phytochem. Rev. 2008, 7, 281–299. [Google Scholar] [CrossRef]
- He, F.; Mu, L.; Yan, G.L.; Liang, N.N.; Pan, Q.H.; Wang, J.; Reeves, M.J.; Duan, C.Q. Biosynthesis of anthocyanins and their regulation in colored grapes. Molecules 2010, 15, 9057–9091. [Google Scholar] [CrossRef] [PubMed]
- Coklar, H. Antioxidant capacity and phenolic profile of berry, seed, and skin of Ekşikara (Vitis vinifera L) grape: Influence of harvest year and altitude. Int. J. Food Prop. 2017, 20, 2071–2087. [Google Scholar] [CrossRef]
- Rodríguez Montealegre, R.; Romero Peces, R.; Chacón Vozmediano, J.L.; Martínez Gascueña, J.; García Romero, E. Phenolic compounds in skins and seeds of ten grape Vitis vinifera varieties grown in a warm climate. J. Food Compos. Anal. 2006, 19, 687–693. [Google Scholar] [CrossRef]
- Yilmaz, Y.; Göksel, Z.; Erdoğan, S.S.; Öztürk, A.; Atak, A.; Özer, C. Antioxidant Activity and Phenolic Content of Seed, Skin and Pulp Parts of 22 Grape (Vitis vinifera L.) Cultivars (4 Common and 18 Registered or Candidate for Registration). J. Food Process. Preserv. 2015, 39, 1682–1691. [Google Scholar] [CrossRef]
- You, Q.; Chen, F.; Wang, X.; Luo, P.G.; Jiang, Y. Inhibitory effects of muscadine anthocyanins on α-glucosidase and pancreatic lipase activities. J. Agric. Food Chem. 2011, 59, 9506–9511. [Google Scholar] [CrossRef]
- Ogasawara, N.; Sasaki, M.; Itoh, Y. Cyanidin-3-rutinoside alleviates postprandial hyperglycemia and its synergism with acarbose by inhibition of intestinal α- glucosidase. J. Clin. Biochem. Nutr. 2011, 48, 154–160. [Google Scholar] [CrossRef]
- Matsui, T.; Tanaka, T.; Tamura, S.; Toshima, A.; Tamaya, K.; Miyata, Y.; Tanaka, K.; Matsumoto, K. A-Glucosidase Inhibitory Profile of Catechins and Theaflavins. J. Agric. Food Chem. 2007, 55, 99–105. [Google Scholar] [CrossRef]
- Gamberucci, A.; Konta, L.; Colucci, A.; Giunti, R.; Magyar, J.É.; Mandl, J.; Bánhegyi, G.; Benedetti, A.; Csala, M. Green tea flavonols inhibit glucosidase II. Biochem. Pharmacol. 2006, 72, 640–646. [Google Scholar] [CrossRef]
- Brown, A.; Anderson, D.; Racicot, K.; Pilkenton, S.J.; Apostolidis, E. Evaluation of Phenolic Phytochemical Enriched Commercial Plant Extracts on the In Vitro Inhibition of α-Glucosidase. Front. Nutr. 2017, 4, 56. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tannat Extracts | Stilbenes * (mg/kg FW) | Flavan-3-ols ** (g/kg FW) | Anthocyanins ** (g MGE/kg FW) | Total Polyphenols * (g GAE/kg FW) | |
|---|---|---|---|---|---|
| FS | seeds | nd | 35.24 ± 2.56 a | nd | 48.56 ± 1.74 ab |
| skins | 0.81 ± 0.20 b | 24.13 ± 6.68 b | nd | 30.29 ± 0.75 bc | |
| B | seeds | nd | 43.22 ± 2.59 a | nd | 58.44 ± 6.1 a |
| skins | 2.81 ± 1.35 ab | 16.23 ± 4.93 b | nd | 19.43 ± 1.87 c | |
| V | seeds | nd | 45.45 ± 6.48 a | nd | 65.29 ± 5.33 a |
| skins | 1.08 ± 0.08 b | 4.34 ± 0.47 c | 1.42 ± 0.27 b | 10.33 ± 1.04 c | |
| M | seeds | nd | 37.34 ± 3.36 a | nd | 47.87 ± 6.48 ab |
| skins | 5.18 ± 2.04 a | 4.63 ± 0.19 c | 8.70 ± 0.75 a | 27.07 ± 2.92 bc | |
| Polyphenolic Compounds | Skins | Seeds | ||||||
|---|---|---|---|---|---|---|---|---|
| FS | BV | V | M | FS | BV | V | M | |
| Stilbenes (mg/kg FW) | ||||||||
| Cis-resveratrol | 0.007 ± 0.001 | 0.011 ± 0.007 | 0.016 ± 0.007 | 0.101 ± 0.028 | - | - | - | - |
| Trans-resveratrol | 0.039 ± 0.011 | 0.014 ± 0.005 | 0.051 ± 0.007 | 0.062 ± 0.011 | - | - | - | - |
| Trans-piceid | 0.152 ± 0.033 | 0.151 ± 0.070 | 0.221 ± 0.005 | 0.746 ± 0.119 | - | - | - | - |
| Cis-piceid | 0.357 ± 0.056 | 0.309 ± 0.118 | 0.344 ± 0.026 | 1.390 ± 0.479 | - | - | - | - |
| Cis-piceatannol | - | - | 0.011 ± 0.003 | 0.152 ± 0.018 | - | - | - | - |
| Trans-piceatannol | - | - | 0.013 ± 0.002 | 0.163 ± 0.017 | - | - | - | - |
| Trans-astringin | 0.032 ± 0.008 | 0.044 ± 0.009 | 0.163 ± 0.020 | 1.273 ± 0.357 | - | - | - | - |
| Cis-astringin | 0.064 ± 0.036 | 0.108 ± 0.063 | 0.103 ± 0.017 | 0.487 ± 0.166 | - | - | - | - |
| ε-viniferin | 0.102 ± 0.039 | 1.669 ± 0.687 | 0.128 ± 0.016 | 0.469 ± 0.038 | - | - | - | - |
| ω-viniferin | 0.005 ± 0.001 | 0.418 ± 0.027 | 0.001 ± 0.000 | 0.029 ± 0.007 | - | - | - | - |
| δ-viniferin | 0.004 ± 0.000 | 0.037 ± 0.003 | 0.006 ± 0.002 | 0.221 ± 0.011 | - | - | - | - |
| Pallidol | 0.003 ± 0.001 | 0.010 ± 0.009 | 0.003 ± 0.000 | 0.042 ± 0.004 | - | - | - | - |
| Hopeaphenol | 0.008 ± 0.004 | 0.011 ± 0.004 | 0.001 ± 0.000 | 0.011 ± 0.009 | - | - | - | - |
| Isohopeaphenol | 0.040 ± 0.004 | 0.024 ± 0.016 | 0.029 ± 0.006 | 0.061 ± 0.002 | - | - | - | - |
| Anthocyanins (mg MGE/kg FW) | ||||||||
| Delphinidin-3-O-glu | - | - | 181.85 ± 24.31 | 1036.81 ± 76.09 | - | - | - | - |
| Cyanidin-3-O-glu | - | - | 134.13 ± 18.29 | 317.03 ± 18.71 | - | - | - | - |
| Petunidin-3-O-glu | - | - | 210.89 ± 29.55 | 1193.13 ± 75.74 | - | - | - | - |
| Peonidin-3-O-glu | - | - | 185.68 ± 26.25 | 694.38 ± 36.33 | - | - | - | - |
| Malvidin-3-O-glu | - | - | 455.83 ± 83.27 | 3609.86 ± 240.78 | - | - | - | - |
| Peonidin-3-O-acetyl-glu | - | - | 50.92 ± 2.70 | 224.57 ± 14.07 | - | - | - | - |
| Malvidin-3-O-acetyl-glu | - | - | 79.91 ± 6.48 | 778.08 ± 62.78 | - | - | - | - |
| Peonidin-3-O-coum-glu | - | - | 58.49 ± 6.81 | 237.19 ± 16.27 | - | - | - | - |
| Malvidin-3-O-coum-glu | - | - | 65.67 ± 8.54 | 604.70 ± 39.03 | - | - | - | - |
| Flavan-3-ols (mg/kg FW) | ||||||||
| Catechin | 1539.40 ± 454.85 | 731.77 ± 156.40 | 143.71 ± 34.34 | - | 1942.38 ± 260.92 | 4620.67 ± 290.95 | 5462.36 ± 331.06 | 2892.28 ± 237.32 |
| Epicatechin | - | - | - | - | 730.15 ± 126.63 | 3024.21 ± 384.69 | 4944.27 ± 303.71 | 2899.74 ± 292.72 |
| Epicatechin gallate | - | - | - | - | 2777.57 ± 353.07 | 4438.71 ± 297.00 | 4796.60 ± 405.72 | 2642.75 ± 199.44 |
| Catechin P | - | - | - | - | 2072.75 ± 178.05 | 2636.23 ± 149.28 | 2087.75 ± 156.28 | 1951.14 ± 297.05 |
| Epicatechin P | 16,142.07 ± 518.50 | 10,257.28 ± 276.46 | 4203.44 ± 305.93 | 4625.27 ± 135.65 | 20,745.92 ± 1206.95 | 21,957.21 ± 543.21 | 21,907.22 ± 971.52 | 21,338.75 ± 1436.20 |
| Epicatechin gallate P | - | - | - | - | 6972.59 ± 352.99 | 6539.24 ± 105.90 | 6251.56 ± 730.40 | 5619.76 ± 353.71 |
| Epigallocatechin P | 6451.77 ± 260.83 | 5245.72 ± 647.76 | - | - | - | - | - | - |
| Added Concentration of Grape Extracts (g/L) | Abs Value (Mean ± SR) | CVR (%) | SRW | CVRW (%) |
|---|---|---|---|---|
| 0 | 2.50 ± 0.10 | 3.5 | 0.2 | 8.8 |
| 0.25 | 1.90 ± 0.10 | 6.6 | 0.2 | 12.0 |
| 0.50 | 0.90 ± 0.10 | 11.5 | 0.2 | 26.1 |
| 1.00 | 0.30 ± 0.10 | 16.3 | 0.1 | 46.8 |
| [I] (mM) | Km (mM/L) | Vmax (1/min) |
|---|---|---|
| Control | ||
| 0 | 0.7395 | 0.1301 |
| Skin | ||
| 0.025 | 9.0839 | 0.2657 |
| 0.05 | 10.0296 | 0.0823 |
| 0.075 | 7.0813 | 0.0283 |
| 0.1 | 6.8055 | 0.0218 |
| Seed | ||
| 0.025 | 1.6743 | 0.1042 |
| 0.05 | 2.6553 | 0.0735 |
| 0.075 | 2.9287 | 0.0653 |
| 0.1 | 3.3618 | 0.0593 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dudoit, A.; Benbouguerra, N.; Richard, T.; Hornedo-Ortega, R.; Valls-Fonayet, J.; Coussot, G.; Saucier, C. α-Glucosidase Inhibitory Activity of Tannat Grape Phenolic Extracts in Relation to Their Ripening Stages. Biomolecules 2020, 10, 1088. https://doi.org/10.3390/biom10081088
Dudoit A, Benbouguerra N, Richard T, Hornedo-Ortega R, Valls-Fonayet J, Coussot G, Saucier C. α-Glucosidase Inhibitory Activity of Tannat Grape Phenolic Extracts in Relation to Their Ripening Stages. Biomolecules. 2020; 10(8):1088. https://doi.org/10.3390/biom10081088
Chicago/Turabian StyleDudoit, Auriane, Nawel Benbouguerra, Tristan Richard, Ruth Hornedo-Ortega, Josep Valls-Fonayet, Gaëlle Coussot, and Cédric Saucier. 2020. "α-Glucosidase Inhibitory Activity of Tannat Grape Phenolic Extracts in Relation to Their Ripening Stages" Biomolecules 10, no. 8: 1088. https://doi.org/10.3390/biom10081088
APA StyleDudoit, A., Benbouguerra, N., Richard, T., Hornedo-Ortega, R., Valls-Fonayet, J., Coussot, G., & Saucier, C. (2020). α-Glucosidase Inhibitory Activity of Tannat Grape Phenolic Extracts in Relation to Their Ripening Stages. Biomolecules, 10(8), 1088. https://doi.org/10.3390/biom10081088