Circulating miRNAs: A New Opportunity in Bone Fragility



Abstract
1. Introduction
- Mechanical stress or biochemical stimuli are detected by osteocytes;
- Activation results in retraction of bone lining cells and digestion of the collagenous membrane by matrix metalloproteinases;
- Preosteoclasts are recruited and, following activation, become multinucleated osteoclasts mediating the bone resorption;
- Osteoblasts reach the resorption cavity producing new osteoid, which in turn calcifies.
2. Circulating miRNAs as Potential Biomarkers in Bone Fragility
2.1. Osteopenic/Osteoporotic Patients
2.2. Diabetic Patients
2.3. Antiosteoporosis Treatment
3. miRNAs in Human Circulating Monocytes
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Krabbe, S.; Transbøl, I.; Christiansen, C. Bone mineral homeostasis, bone growth, and mineralisation during years of pubertal growth: A unifying concept. Arch. Dis. Child. 1982, 57, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Buckwalter, J.A.; Glimcher, M.J.; Cooper, R.R.; Recker, R. Bone biology. I: Structure, blood supply, cells, matrix, and mineralization. Instr. Course Lect. 1996, 45, 371–386. [Google Scholar] [PubMed]
- Seeman, E. Bone quality: The material and structural basis of bone strength. J. Bone Miner. Metab. 2008, 26, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Fratzl, P.; Gupta, H.S.; Paschalis, E.P.; Roschger, P. Structure and mechanical quality of the collagen–mineral nano-composite in bone. J. Mater. Chem. 2004, 14, 2115–2123. [Google Scholar] [CrossRef]
- Florencio-Silva, R.; de Sasso, G.R.S.; Sasso-Cerri, E.; Simões, M.J.; Cerri, P.S. Biology of Bone Tissue: Structure, Function, and Factors That Influence Bone Cells. BioMed Res. Int. 2015, 2015, 421746. [Google Scholar] [CrossRef]
- Datta, H.K.; Ng, W.F.; Walker, J.A.; Tuck, S.P.; Varanasi, S.S. The cell biology of bone metabolism. J. Clin. Pathol. 2008, 61, 577–587. [Google Scholar] [CrossRef]
- Tranquilli Leali, P.; Doria, C.; Zachos, A.; Ruggiu, A.; Milia, F.; Barca, F. Bone fragility: Current reviews and clinical features. Clin. Cases Miner. Bone Metab. 2009, 6, 109–113. [Google Scholar]
- Materozzi, M.; Merlotti, D.; Gennari, L.; Bianciardi, S. The Potential Role of miRNAs as New Biomarkers for Osteoporosis. Int. J. Endocrinol. 2018, 2018, 2342860. [Google Scholar] [CrossRef]
- Qin, Y.-X.; Xia, Y.; Muir, J.; Lin, W.; Rubin, C.T. Quantitative ultrasound imaging monitoring progressive disuse osteopenia and mechanical stimulation mitigation in calcaneus region through a 90-day bed rest human study. J. Orthop. Transl. 2019, 18, 48–58. [Google Scholar] [CrossRef]
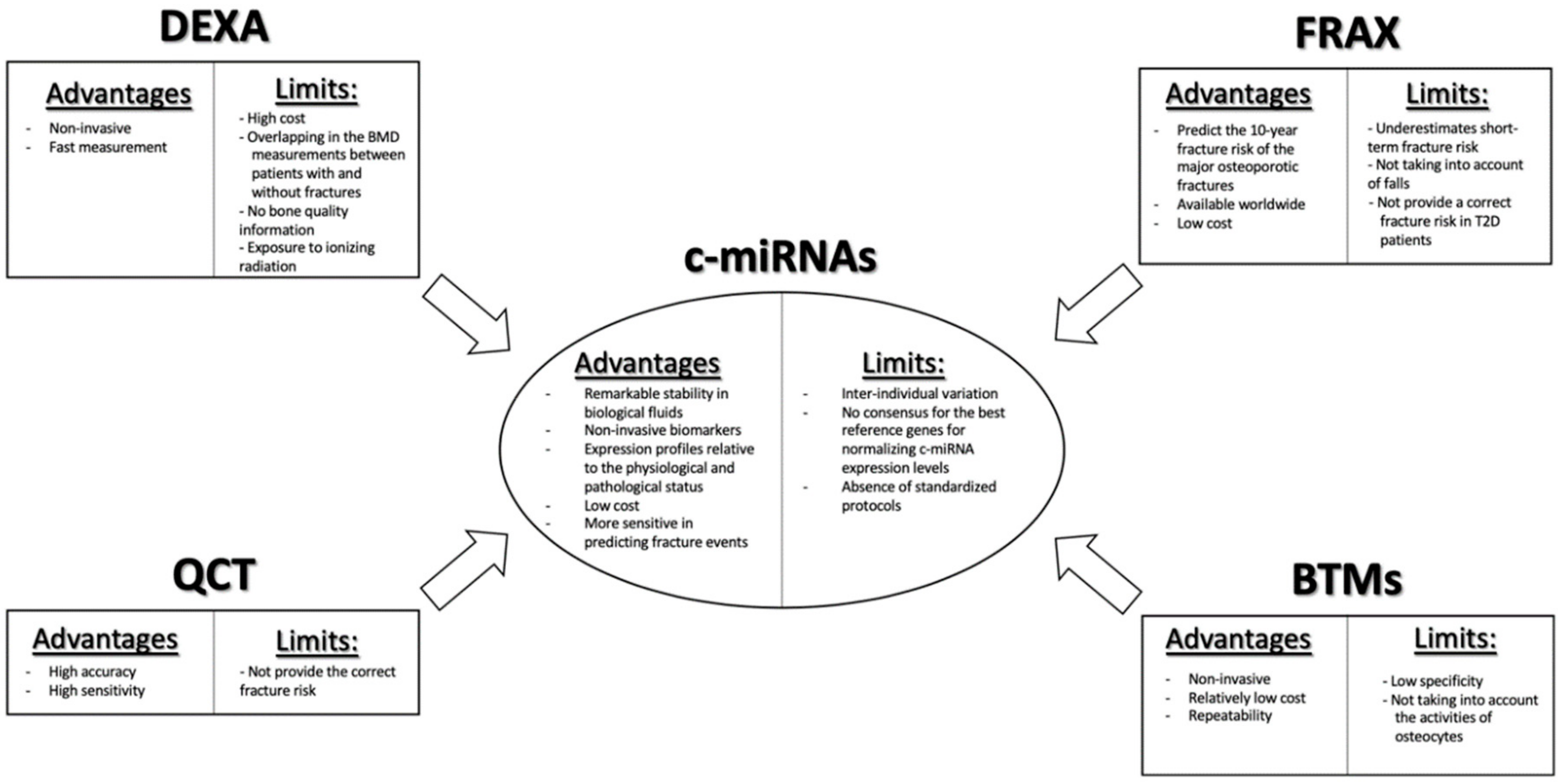
- Kanis, J.A.; Harvey, N.C.; Johansson, H.; Liu, E.; Vandenput, L.; Lorentzon, M.; Leslie, W.D.; McCloskey, E.V. A decade of FRAX: How has it changed the management of osteoporosis? Aging Clin. Exp. Res. 2020, 32, 187–196. [Google Scholar] [CrossRef]
- Leslie, W.D.; Rubin, M.R.; Schwartz, A.V.; Kanis, J.A. Type 2 diabetes and bone. J. Bone Miner. Res. 2012, 27, 2231–2237. [Google Scholar] [CrossRef] [PubMed]
- Watts, N.B.; Bilezikian, J.P.; Camacho, P.M.; Greenspan, S.L.; Harris, S.T.; Hodgson, S.F.; Kleerekoper, M.; Luckey, M.M.; McClung, M.R.; Pollack, R.P.; et al. American Association of Clinical Endocrinologists Medical Guidelines for Clinical Practice for the diagnosis and treatment of postmenopausal osteoporosis. Endocr. Pr. 2010, 16 (Suppl. 3), 1–37. [Google Scholar] [CrossRef]
- National Institute for Health and Care Excellence. Osteoporosis: Assessing the Risk of Fragility Fracture; National Institute for Health and Care Excellence: Clinical Guidelines; National Institute for Health and Care Excellence (UK): London, UK, 2017; ISBN 978-1-4731-2359-5. [Google Scholar]
- Fonseca, H.; Moreira-Gonçalves, D.; Coriolano, H.-J.A.; Duarte, J.A. Bone quality: The determinants of bone strength and fragility. Sports Med. 2014, 44, 37–53. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, G.; Lanteri, P.; Colombini, A.; Banfi, G. Blood biochemical markers of bone turnover: Pre-analytical and technical aspects of sample collection and handling. Clin. Chem. Lab. Med. 2012, 50, 771–789. [Google Scholar] [CrossRef] [PubMed]
- Garnero, P. New developments in biological markers of bone metabolism in osteoporosis. Bone 2014, 66, 46–55. [Google Scholar] [CrossRef]
- Hackl, M.; Heilmeier, U.; Weilner, S.; Grillari, J. Circulating microRNAs as novel biomarkers for bone diseases—Complex signatures for multifactorial diseases? Mol. Cell. Endocrinol. 2016, 432, 83–95. [Google Scholar] [CrossRef]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Wightman, B.; Ha, I.; Ruvkun, G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993, 75, 855–862. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. (Lausanne) 2018, 9, 402. [Google Scholar] [CrossRef]
- Flynt, A.S.; Lai, E.C. Biological principles of microRNA-mediated regulation: Shared themes amid diversity. Nat. Rev. Genet. 2008, 9, 831–842. [Google Scholar] [CrossRef]
- Carthew, R.W.; Sontheimer, E.J. Origins and Mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [PubMed]
- Malan-Müller, S.; Hemmings, S.M.J.; Seedat, S. Big effects of small RNAs: A review of microRNAs in anxiety. Mol. Neurobiol. 2013, 47, 726–739. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Huntzinger, E.; Izaurralde, E. Gene silencing by microRNAs: Contributions of translational repression and mRNA decay. Nat. Rev. Genet. 2011, 12, 99–110. [Google Scholar] [CrossRef]
- Ipsaro, J.J.; Joshua-Tor, L. From guide to target: Molecular insights into eukaryotic RNA-interference machinery. Nat. Struct. Mol. Biol. 2015, 22, 20–28. [Google Scholar] [CrossRef]
- Denli, A.M.; Tops, B.B.J.; Plasterk, R.H.A.; Ketting, R.F.; Hannon, G.J. Processing of primary microRNAs by the Microprocessor complex. Nature 2004, 432, 231–235. [Google Scholar] [CrossRef]
- Okada, C.; Yamashita, E.; Lee, S.J.; Shibata, S.; Katahira, J.; Nakagawa, A.; Yoneda, Y.; Tsukihara, T. A high-resolution structure of the pre-microRNA nuclear export machinery. Science 2009, 326, 1275–1279. [Google Scholar] [CrossRef]
- Yoda, M.; Kawamata, T.; Paroo, Z.; Ye, X.; Iwasaki, S.; Liu, Q.; Tomari, Y. ATP-dependent human RISC assembly pathways. Nat. Struct. Mol. Biol. 2010, 17, 17–23. [Google Scholar] [CrossRef]
- Khvorova, A.; Reynolds, A.; Jayasena, S.D. Functional siRNAs and miRNAs exhibit strand bias. Cell 2003, 115, 209–216. [Google Scholar] [CrossRef]
- Reid, G.; Kirschner, M.B.; van Zandwijk, N. Circulating microRNAs: Association with disease and potential use as biomarkers. Crit. Rev. Oncol. Hematol. 2011, 80, 193–208. [Google Scholar] [CrossRef]
- Ardekani, A.M.; Naeini, M.M. The Role of MicroRNAs in Human Diseases. Avicenna J. Med. Biotechnol. 2010, 2, 161–179. [Google Scholar] [PubMed]
- Chim, S.S.C.; Shing, T.K.F.; Hung, E.C.W.; Leung, T.-Y.; Lau, T.-K.; Chiu, R.W.K.; Lo, Y.M.D. Detection and characterization of placental microRNAs in maternal plasma. Clin. Chem. 2008, 54, 482–490. [Google Scholar] [CrossRef] [PubMed]
- Lawrie, C.H.; Gal, S.; Dunlop, H.M.; Pushkaran, B.; Liggins, A.P.; Pulford, K.; Banham, A.H.; Pezzella, F.; Boultwood, J.; Wainscoat, J.S.; et al. Detection of elevated levels of tumour-associated microRNAs in serum of patients with diffuse large B-cell lymphoma. Br. J. Haematol. 2008, 141, 672–675. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Guo, J.; Zhang, Y.; Chen, J.; Guo, X.; et al. Characterization of microRNAs in serum: A novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res. 2008, 18, 997–1006. [Google Scholar] [CrossRef]
- Cogswell, J.P.; Ward, J.; Taylor, I.A.; Waters, M.; Shi, Y.; Cannon, B.; Kelnar, K.; Kemppainen, J.; Brown, D.; Chen, C.; et al. Identification of miRNA changes in Alzheimer’s disease brain and CSF yields putative biomarkers and insights into disease pathways. J. Alzheimers Dis. 2008, 14, 27–41. [Google Scholar] [CrossRef]
- Gallo, A.; Tandon, M.; Alevizos, I.; Illei, G.G. The majority of microRNAs detectable in serum and saliva is concentrated in exosomes. PLoS ONE 2012, 7, e30679. [Google Scholar] [CrossRef]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; Huang, K.H.; Lee, M.J.; Galas, D.J.; Wang, K. The MicroRNA Spectrum in 12 Body Fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef]
- Zhou, Q.; Li, M.; Wang, X.; Li, Q.; Wang, T.; Zhu, Q.; Zhou, X.; Wang, X.; Gao, X.; Li, X. Immune-related MicroRNAs are Abundant in Breast Milk Exosomes. Int. J. Biol. Sci. 2011, 8, 118–123. [Google Scholar] [CrossRef]
- Da Silveira, J.C.; Veeramachaneni, D.N.R.; Winger, Q.A.; Carnevale, E.M.; Bouma, G.J. Cell-secreted vesicles in equine ovarian follicular fluid contain miRNAs and proteins: A possible new form of cell communication within the ovarian follicle. Biol. Reprod. 2012, 86, 71. [Google Scholar] [CrossRef]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. MicroRNAs are Transported in Plasma and Delivered to Recipient Cells by High-Density Lipoproteins. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Tabet, F.; Vickers, K.C.; Cuesta Torres, L.F.; Wiese, C.B.; Shoucri, B.M.; Lambert, G.; Catherinet, C.; Prado-Lourenco, L.; Levin, M.G.; Thacker, S.; et al. HDL-transferred microRNA-223 regulates ICAM-1 expression in endothelial cells. Nat. Commun. 2014, 5, 3292. [Google Scholar] [CrossRef] [PubMed]
- Turchinovich, A.; Weiz, L.; Burwinkel, B. Extracellular miRNAs: The mystery of their origin and function. Trends Biochem. Sci. 2012, 37, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Iftikhar, H.; Carney, G.E. Evidence and potential in vivo functions for biofluid miRNAs: From expression profiling to functional testing: Potential roles of extracellular miRNAs as indicators of physiological change and as agents of intercellular information exchange. BioEssays 2016, 38, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liang, H.; Zhang, J.; Zen, K.; Zhang, C.-Y. Secreted microRNAs: A new form of intercellular communication. Trends Cell Biol. 2012, 22, 125–132. [Google Scholar] [CrossRef]
- Cui, M.; Wang, H.; Yao, X.; Zhang, D.; Xie, Y.; Cui, R.; Zhang, X. Circulating MicroRNAs in Cancer: Potential and Challenge. Front. Genet. 2019, 10. [Google Scholar] [CrossRef]
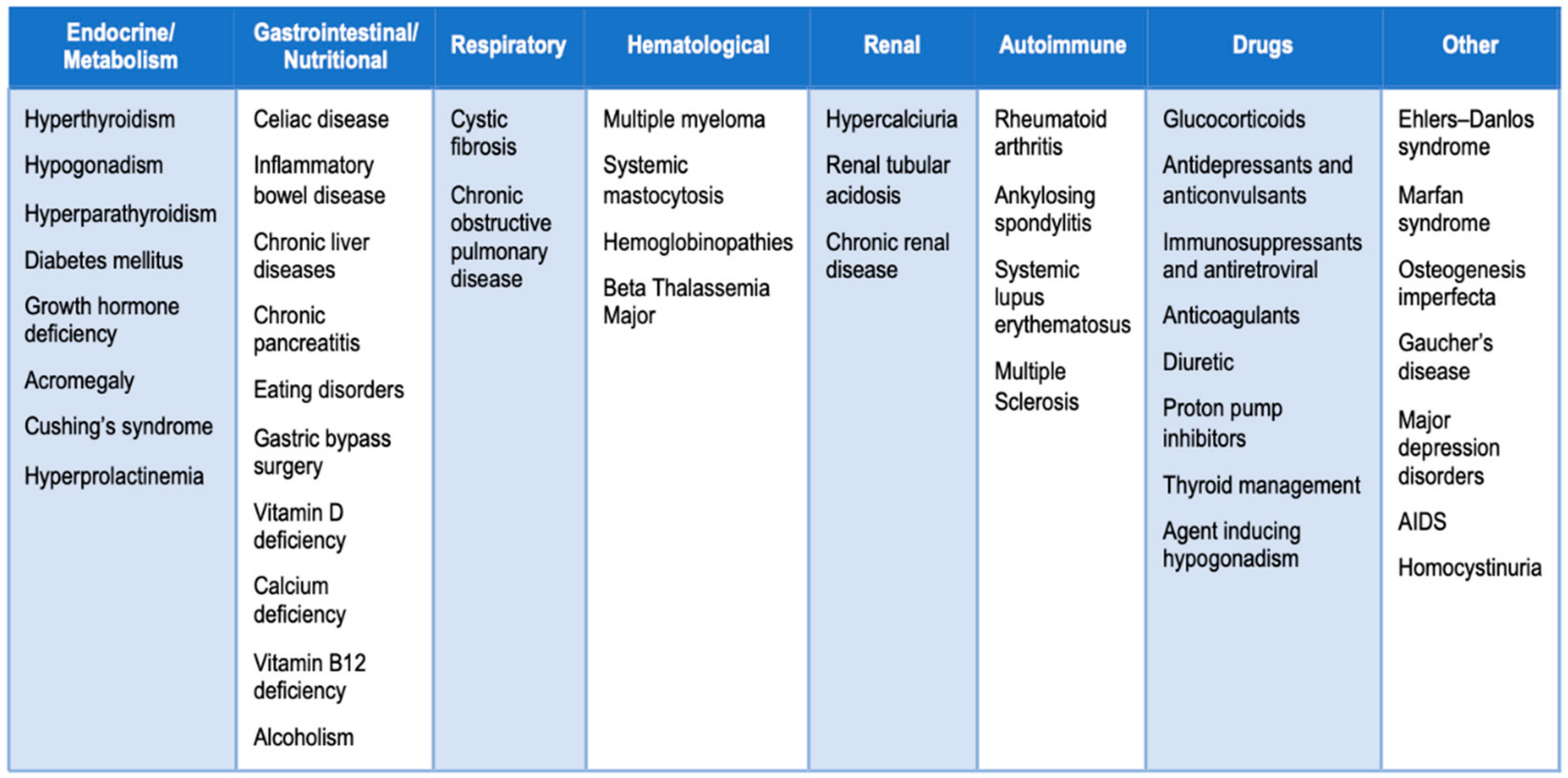
- Marcucci, G.; Brandi, M.L. Rare causes of osteoporosis. Clin. Cases Miner. Bone Metab. 2015, 12, 151–156. [Google Scholar] [CrossRef]
- Mirza, F.; Canalis, E. Secondary Osteoporosis: Pathophysiology and Management. Eur. J. Endocrinol. 2015, 173, R131–R151. [Google Scholar] [CrossRef]
- Seeliger, C.; Karpinski, K.; Haug, A.T.; Vester, H.; Schmitt, A.; Bauer, J.S.; van Griensven, M. Five freely circulating miRNAs and bone tissue miRNAs are associated with osteoporotic fractures. J. Bone Miner. Res. 2014, 29, 1718–1728. [Google Scholar] [CrossRef]
- Li, H.; Wang, Z.; Fu, Q.; Zhang, J. Plasma miRNA levels correlate with sensitivity to bone mineral density in postmenopausal osteoporosis patients. Biomarkers 2014, 19, 553–556. [Google Scholar] [CrossRef]
- Panach, L.; Mifsut, D.; Tarín, J.J.; Cano, A.; García-Pérez, M.Á. Serum Circulating MicroRNAs as Biomarkers of Osteoporotic Fracture. Calcif. Tissue Int. 2015, 97, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Weilner, S.; Skalicky, S.; Salzer, B.; Keider, V.; Wagner, M.; Hildner, F.; Gabriel, C.; Dovjak, P.; Pietschmann, P.; Grillari-Voglauer, R.; et al. Differentially circulating miRNAs after recent osteoporotic fractures can influence osteogenic differentiation. Bone 2015, 79, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Kocijan, R.; Muschitz, C.; Geiger, E.; Skalicky, S.; Baierl, A.; Dormann, R.; Plachel, F.; Feichtinger, X.; Heimel, P.; Fahrleitner-Pammer, A.; et al. Circulating microRNA Signatures in Patients With Idiopathic and Postmenopausal Osteoporosis and Fragility Fractures. J. Clin. Endocrinol. Metab. 2016, 101, 4125–4134. [Google Scholar] [CrossRef] [PubMed]
- Feichtinger, X.; Muschitz, C.; Heimel, P.; Baierl, A.; Fahrleitner-Pammer, A.; Redl, H.; Resch, H.; Geiger, E.; Skalicky, S.; Dormann, R.; et al. Bone-related Circulating MicroRNAs miR-29b-3p, miR-550a-3p, and miR-324-3p and their Association to Bone Microstructure and Histomorphometry. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, K.; Pang, Q.; Yang, C.; Zhang, H.; Wu, F.; Cao, H.; Liu, H.; Wan, Y.; Xia, W.; et al. Identification of suitable reference gene and biomarkers of serum miRNAs for osteoporosis. Sci. Rep. 2016, 6, 36347. [Google Scholar] [CrossRef]
- Bedene, A.; Mencej Bedrač, S.; Ješe, L.; Marc, J.; Vrtačnik, P.; Preželj, J.; Kocjan, T.; Kranjc, T.; Ostanek, B. MiR-148a the epigenetic regulator of bone homeostasis is increased in plasma of osteoporotic postmenopausal women. Wien Klin. Wochenschr. 2016, 128, 519–526. [Google Scholar] [CrossRef]
- Yavropoulou, M.P.; Anastasilakis, A.D.; Makras, P.; Tsalikakis, D.G.; Grammatiki, M.; Yovos, J.G. Expression of microRNAs that regulate bone turnover in the serum of postmenopausal women with low bone mass and vertebral fractures. Eur. J. Endocrinol. 2017, 176, 169–176. [Google Scholar] [CrossRef]
- Ramírez-Salazar, E.G.; Carrillo-Patiño, S.; Hidalgo-Bravo, A.; Rivera-Paredez, B.; Quiterio, M.; Ramírez-Palacios, P.; Patiño, N.; Valdés-Flores, M.; Salmerón, J.; Velázquez-Cruz, R. Serum miRNAs miR-140-3p and miR-23b-3p as potential biomarkers for osteoporosis and osteoporotic fracture in postmenopausal Mexican-Mestizo women. Gene 2018, 679, 19–27. [Google Scholar] [CrossRef]
- Mandourah, A.Y.; Ranganath, L.; Barraclough, R.; Vinjamuri, S.; Hof, R.V.; Hamill, S.; Czanner, G.; Dera, A.A.; Wang, D.; Barraclough, D.L. Circulating microRNAs as potential diagnostic biomarkers for osteoporosis. Sci. Rep. 2018, 8, 8421. [Google Scholar] [CrossRef]
- Chen, R.; Liao, X.; Chen, F.; Wang, B.; Huang, J.; Jian, G.; Huang, Z.; Yin, G.; Liu, H.; Jin, D. Circulating microRNAs, miR-10b-5p, miR-328-3p, miR-100 and let-7, are associated with osteoblast differentiation in osteoporosis. Int. J. Clin. Exp. Pathol. 2018, 11, 1383–1390. [Google Scholar]
- Wang, C.; He, H.; Wang, L.; Jiang, Y.; Xu, Y. Reduced miR-144-3p expression in serum and bone mediates osteoporosis pathogenesis by targeting RANK. Biochem. Cell Biol. 2018, 96, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Feurer, E.; Kan, C.; Croset, M.; Sornay-Rendu, E.; Chapurlat, R. Lack of Association Between Select Circulating miRNAs and Bone Mass, Turnover, and Fractures: Data From the OFELY Cohort. J. Bone Miner. Res. 2019, 34, 1074–1085. [Google Scholar] [CrossRef]
- Pickering, M.-E.; Millet, M.; Rousseau, J.-C.; Croset, M.; Szulc, P.; Borel, O.; Sornay Rendu, E.; Chapurlat, R. Selected serum microRNA, abdominal aortic calcification and risk of osteoporotic fracture. PLoS ONE 2019, 14, e0216947. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Bemben, M.G.; Bemben, D.A. Bone and muscle specific circulating microRNAs in postmenopausal women based on osteoporosis and sarcopenia status. Bone 2019, 120, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Yu, S.; Jin, R.; Xiao, Y.; Pan, M.; Pei, F.; Zhu, X.; Huang, H.; Zhang, Z.; Chen, S.; et al. Circulating miR-338 Cluster activities on osteoblast differentiation: Potential Diagnostic and Therapeutic Targets for Postmenopausal Osteoporosis. Theranostics 2019, 9, 3780–3797. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Hu, L.; Wang, S.; Huang, T.; Zhang, M.; Yang, M.; Zhen, W.; Yang, D.; Lu, W.; Guan, M.; et al. Circulating MicroRNA-19b Identified From Osteoporotic Vertebral Compression Fracture Patients Increases Bone Formation. J. Bone Miner. Res. 2020, 35, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Zarecki, P.; Hackl, M.; Grillari, J.; Debono, M.; Eastell, R. Serum microRNAs as novel biomarkers for osteoporotic vertebral fractures. Bone 2020, 130, 115105. [Google Scholar] [CrossRef]
- Bolamperti, S.; Villa, I.; Spinello, A.; Manfredini, G.; Mrak, E.; Mezzadri, U.; Ometti, M.; Fraschini, G.; Guidobono, F.; Rubinacci, A. Evidence for Altered Canonical Wnt Signaling in the Trabecular Bone of Elderly Postmenopausal Women with Fragility Femoral Fracture. BioMed Res. Int. 2016, 2016, 8169614. [Google Scholar] [CrossRef]
- Mäkitie, R.E.; Hackl, M.; Niinimäki, R.; Kakko, S.; Grillari, J.; Mäkitie, O. Altered MicroRNA Profile in Osteoporosis Caused by Impaired WNT Signaling. J. Clin. Endocrinol. Metab. 2018, 103, 1985–1996. [Google Scholar] [CrossRef]
- Walter, E.; Dellago, H.; Grillari, J.; Dimai, H.P.; Hackl, M. Cost-utility analysis of fracture risk assessment using microRNAs compared with standard tools and no monitoring in the Austrian female population. Bone 2018, 108, 44–54. [Google Scholar] [CrossRef]
- Ladang, A.; Beaudart, C.; Locquet, M.; Reginster, J.-Y.; Bruyère, O.; Cavalier, E. Evaluation of a Panel of MicroRNAs that Predicts Fragility Fracture Risk: A Pilot Study. Calcif. Tissue Int. 2019, 106, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Heilmeier, U.; Hackl, M.; Skalicky, S.; Weilner, S.; Schroeder, F.; Vierlinger, K.; Patsch, J.M.; Baum, T.; Oberbauer, E.; Lobach, I.; et al. Serum miRNA Signatures Are Indicative of Skeletal Fractures in Postmenopausal Women With and Without Type 2 Diabetes and Influence Osteogenic and Adipogenic Differentiation of Adipose Tissue-Derived Mesenchymal Stem Cells In Vitro. J. Bone Miner. Res. 2016, 31, 2173–2192. [Google Scholar] [CrossRef] [PubMed]
- Grieco, G.E.; Cataldo, D.; Ceccarelli, E.; Nigi, L.; Catalano, G.; Brusco, N.; Mancarella, F.; Ventriglia, G.; Fondelli, C.; Guarino, E.; et al. Serum Levels of miR-148a and miR-21-5p Are Increased in Type 1 Diabetic Patients and Correlated with Markers of Bone Strength and Metabolism. Noncoding RNA 2018, 4, 37. [Google Scholar] [CrossRef] [PubMed]
- Anastasilakis, A.D.; Yavropoulou, M.P.; Makras, P.; Sakellariou, G.T.; Papadopoulou, F.; Gerou, S.; Papapoulos, S.E. Increased osteoclastogenesis in patients with vertebral fractures following discontinuation of denosumab treatment. Eur. J. Endocrinol. 2017, 176, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Anastasilakis, A.D.; Makras, P.; Pikilidou, M.; Tournis, S.; Makris, K.; Bisbinas, I.; Tsave, O.; Yovos, J.G.; Yavropoulou, M.P. Changes of Circulating MicroRNAs in Response to Treatment with Teriparatide or Denosumab in Postmenopausal Osteoporosis. J. Clin. Endocrinol. Metab. 2018, 103, 1206–1213. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, L.; Moore, B.T.; Peng, X.-H.; Fang, X.; Lappe, J.M.; Recker, R.R.; Xiao, P. MiR-133a in human circulating monocytes: A potential biomarker associated with postmenopausal osteoporosis. PLoS ONE 2012, 7, e34641. [Google Scholar] [CrossRef]
- Cao, Z.; Moore, B.T.; Wang, Y.; Peng, X.-H.; Lappe, J.M.; Recker, R.R.; Xiao, P. MiR-422a as a potential cellular microRNA biomarker for postmenopausal osteoporosis. PLoS ONE 2014, 9, e97098. [Google Scholar] [CrossRef]
- Jiménez-Ortega, R.F.; Ramírez-Salazar, E.G.; Parra-Torres, A.Y.; Muñoz-Montero, S.A.; Rangel-Escareňo, C.; Salido-Guadarrama, I.; Rodriguez-Dorantes, M.; Quiterio, M.; Salmerón, J.; Velázquez-Cruz, R. Identification of microRNAs in human circulating monocytes of postmenopausal osteoporotic Mexican-Mestizo women: A pilot study. Exp. Ther. Med. 2017, 14, 5464–5472. [Google Scholar] [CrossRef]
- De-La-Cruz-Montoya, A.H.; Ramírez-Salazar, E.G.; Martínez-Aguilar, M.M.; González-de-la-Rosa, P.M.; Quiterio, M.; Abreu-Goodger, C.; Salmerón, J.; Velázquez-Cruz, R. Identification of miR-708-5p in peripheral blood monocytes: Potential marker for postmenopausal osteoporosis in Mexican-Mestizo population. Exp. Biol. Med. (Maywood) 2018, 243, 1027–1036. [Google Scholar] [CrossRef]
- Zhang, X.; Liang, H.; Kourkoumelis, N.; Wu, Z.; Li, G.; Shang, X. Comprehensive Analysis of lncRNA and miRNA Expression Profiles and ceRNA Network Construction in Osteoporosis. Calcif. Tissue Int. 2019. [Google Scholar] [CrossRef]
- Lassere, M.N. The Biomarker-Surrogacy Evaluation Schema: A review of the biomarker-surrogate literature and a proposal for a criterion-based, quantitative, multidimensional hierarchical levels of evidence schema for evaluating the status of biomarkers as surrogate endpoints. Stat. Methods Med. Res. 2008, 17, 303–340. [Google Scholar] [CrossRef] [PubMed]
- NIH Consensus Development Panel on Osteoporosis Prevention. Osteoporosis Prevention, Diagnosis, and Therapy. JAMA 2001, 285, 785–795. [Google Scholar] [CrossRef] [PubMed]
- Johnell, O.; Kanis, J.A. An estimate of the worldwide prevalence and disability associated with osteoporotic fractures. Osteoporos. Int. 2006, 17, 1726–1733. [Google Scholar] [CrossRef] [PubMed]
- Morrow, D.A.; de Lemos, J.A. Benchmarks for the Assessment of Novel Cardiovascular Biomarkers. Circulation 2007, 115, 949–952. [Google Scholar] [CrossRef] [PubMed]
- Marini, F.; Cianferotti, L.; Brandi, M.L. Epigenetic Mechanisms in Bone Biology and Osteoporosis: Can They Drive Therapeutic Choices? Int. J. Mol. Sci. 2016, 17, 1329. [Google Scholar] [CrossRef]
- Feng, Q.; Zheng, S.; Zheng, J. The emerging role of microRNAs in bone remodeling and its therapeutic implications for osteoporosis. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef]
- Zhou, Y.; Deng, H.-W.; Shen, H. Circulating monocytes: An appropriate model for bone-related study. Osteoporos. Int. 2015, 26, 2561–2572. [Google Scholar] [CrossRef]
- Terrinoni, A.; Calabrese, C.; Basso, D.; Aita, A.; Caporali, S.; Plebani, M.; Bernardini, S. The circulating miRNAs as diagnostic and prognostic markers. Clin. Chem. Lab. Med. 2019, 57, 932–953. [Google Scholar] [CrossRef]
- Hayes, J.; Peruzzi, P.P.; Lawler, S. MicroRNAs in cancer: Biomarkers, functions and therapy. Trends Mol. Med. 2014, 20, 460–469. [Google Scholar] [CrossRef]
- Kelch, S.; Balmayor, E.R.; Seeliger, C.; Vester, H.; Kirschke, J.S.; van Griensven, M. miRNAs in bone tissue correlate to bone mineral density and circulating miRNAs are gender independent in osteoporotic patients. Sci. Rep. 2017, 7, 15861. [Google Scholar] [CrossRef]
- Lennox, K.A.; Behlke, M.A. Chemical modification and design of anti-miRNA oligonucleotides. Gene Ther. 2011, 18, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Moldovan, L.; Batte, K.E.; Trgovcich, J.; Wisler, J.; Marsh, C.B.; Piper, M. Methodological challenges in utilizing miRNAs as circulating biomarkers. J. Cell. Mol. Med. 2014, 18, 371–390. [Google Scholar] [CrossRef] [PubMed]
- Donati, S.; Ciuffi, S.; Brandi, M.L. Human Circulating miRNAs Real-time qRT-PCR-based Analysis: An Overview of Endogenous Reference Genes Used for Data Normalization. Int. J. Mol. Sci. 2019, 20, 4353. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Biological Fluids | Study Population | Platforms for miRNAs Expression Profiling | Identified Candidate c-miRNAs | ROC Curve Analysis Data (AUC Value) | RGs Used for Normalizing qPCR Data | Study |
|---|---|---|---|---|---|---|
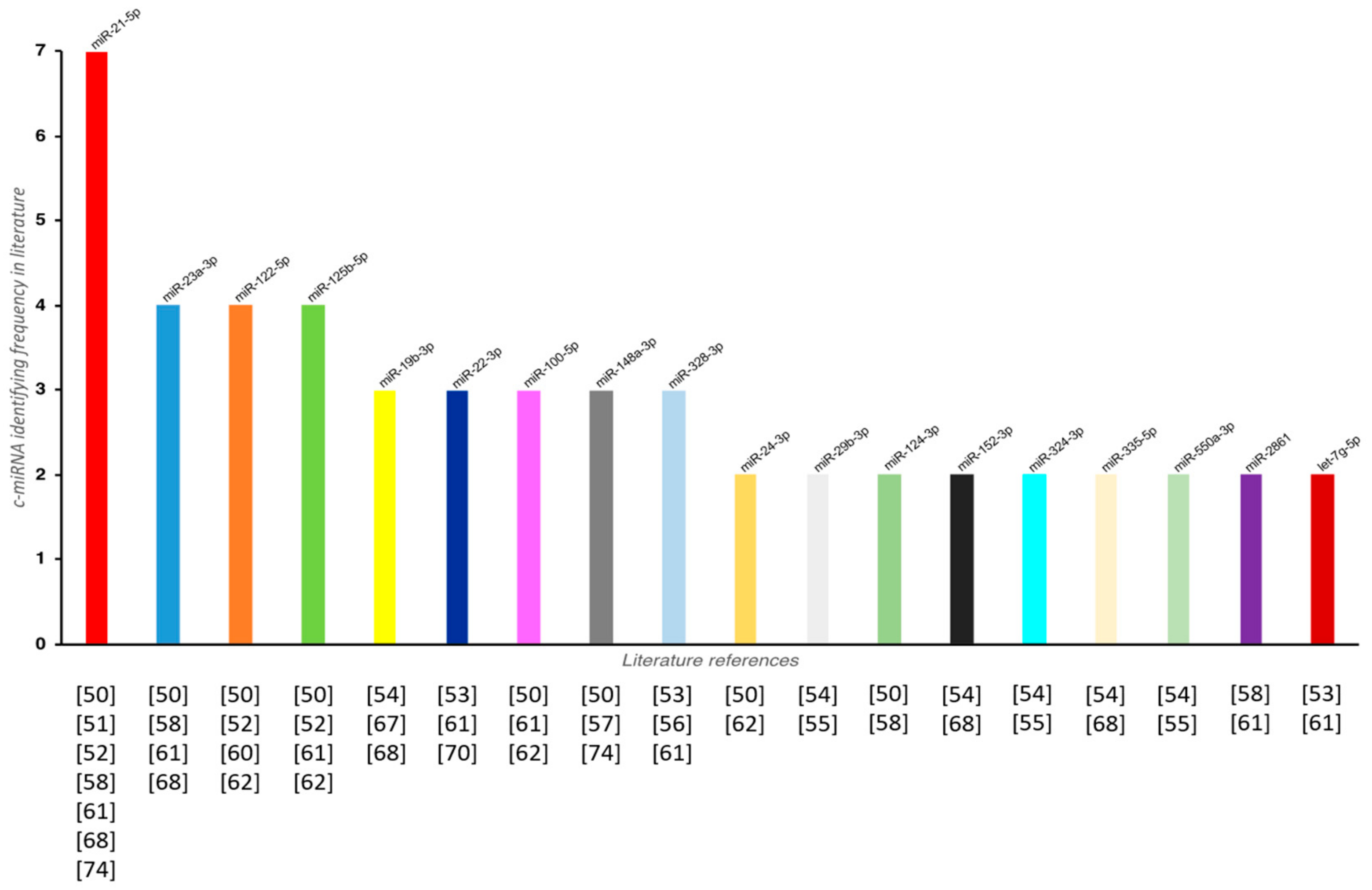
| Serum | OP (40), HC (40) | Prescreening: miRNA PCR arrays Validation: qPCR | miR-21-5p (↑), miR-23a-3p (↑), miR-24-3p (↑), miR-93 (↑), miR-100-5p (↑), miR-122-5p (↑), miR-124-3p (↑), miR-125b-5p (↑), miR-148a-3p (↑) | OP vs. HC 0.63 (miR-21-5p), 0.63 (miR-23a-3p), 0.63 (miR-24-3p), 0.68 (miR-93), 0.69 (miR-100-5p), 0.77 (miR-122-5p), 0.69 (miR-124-3p), 0.76 (miR-125b-5p), 0.61 (miR-148a-3p) | Average of SNORD96A and RNU6 | [50] |
| Plasma | Osteopenic (40), OP (40), HC (40) | qPCR | miR-21-5p (↓), miR-133a (↑) | / | miR-16 | [51] |
| Serum | OP with bone fractures (23), HC (17) | Prescreening: PCR Panel Validation: qPCR | miR-122-5p (↑), miR-125b-5p (↑), miR-21-5p (↑) | OP fracture vs. HC 0.87 (miR-122-5p), 0.76 (miR-125b-5p), 0.87 (miR-21-5p) | miR-93-5p | [52] |
| Serum | OP with fractures (19), HC (18) | Prescreening: qPCR Validation: qPCR | miR-22-3p (↑), miR-328-3p (↓), let-7g-5p (↓) | / | Global mean Ct value | [53] |
| Serum | Low-traumatic fractures (36), HC (39) | qPCR | miR-152-3p (↓), miR-30e-5p (↓), miR-140-5p (↓), miR-324-3p (↓), miR-19b-3p (↓), miR-335-5p (↑), miR-19a-3p (↓), miR-550a-3p (↓), miR-29b-3p (↓) | OP fracture vs. HC 0.962 (miR-152-3p), 0.959 (miR-30e-5p), 0.947 (miR-140-5p), 0.95 (miR-324-3p), 0.944 (miR-19b-3p), 0.939 (miR-335-5p), 0.929 (miR-19a-3p), 0.909 (miR-550a-3p), 0.838 (miR-29b-3p) | Global mean Ct value | [54] |
| Serum | Low-traumatic fractures (36) | qPCR | miR-29b-3p (↓), miR-324-3p (↓), miR-550a-3p (↓) | / | Global mean Ct value | [55] |
| Serum | OVX rats, rhesus monkeys, women with normal BMD (19), osteopenia (7), OP (10) | Prescreening: miRNA PCR arrays Validation: qPCR | miR-30b-5p (↓), miR-103-3p (↓), miR-142-3p (↓), miR-328-3p (↓) | OP vs. HC 0.926 (miR-30b-5p), 0.796 (miR-103-3p), 0.95 (miR-142-3p) | miR-25-3p | [56] |
| Plasma | OP (17), HC (57) | qPCR | miR-148a-3p (↑) | / | Mean of miR-16-5p and let-7a-5p | [57] |
| Serum | OP with vertebral fractures (35) OP (35), HC (30) | qPCR | miR-124-3p (↑), miR-2861 (↑), miR-21-5p (↓), miR-23a-3p (↓), miR-29a-3p (↓) | OP vs. osteopenic 0.66 (miR-21-5p), 0.63 (miR-23a-3p), 0.61 (miR-29a-3p) | SNORD95A, SNORD96A, RNU6-2 | [58] |
| Serum | Osteopenic (28), OP (46), OP with hip fractures (21), HC (42) | Prescreening: qPCR arrays Validation: qPCR | miR-23b-3p (↑), miR-140-3p (↑) | OP vs. HC: 0.69 (miR-23b-3p), 0.96 (miR-140-3p); OP fracture vs. HC: 0.8869 (miR-23b-3p), 0.92 (miR-140-3p) | RNU6 | [59] |
| Serum and Plasma | Osteopenic with fractures (15), Osteopenic w/o fractures (61), OP with fracture (18), OP w/o fractures (33), HC (12) | Prescreening: qPCR arrays Validation: qPCR | miR-122-5p (↓), miR-4516 (↓) | OP vs. HC 0.727 (miR-4516) 0.757 (miR-4516 + miR-122-5p) | SNORD96A, RNU6-6P | [60] |
| Serum | OP (9), HC (9) | qPCR | let-7g-5p (↓), miR-133a-5p (↓), miR-328-3p (↓), miR-22-3p (↓), miR-2861 (↓), miR-518d-5p (↓), miR-10b-5p (↑), miR-21-5p (↑), miR-125b-5p (↑), miR-23-3p (↑), miR-100-5p (↑) | OP vs. HC 0.8944 (let-7g-5p), 0.8656 (miR-328-3p), 0.8728 (miR-10b-5p), 0.8875 (miR-100) | U6 | [61] |
| Serum | OP (45), fractured non-OP (15) | qPCR | miR-24-3p (↑), miR-27a-3p (↑), miR-100-5p (↑), miR-125b-5p (↑), miR-122-5p (↑), miR-145 (↑), miR-144-3p (↓) | / | U6 | [62] |
| Serum | OP w/o fractures (559), OP with fractures (123) | qPCR | / | / | UniSP6, Mean of each miRNA | [63] |
| Serum | OP fractures (217), HC (217) | qPCR | / | / | miR-191-5p, miR-222-3p miR-361-5p | [64] |
| Serum | NN (13), OO (46), SOP (15), SP (1) | qPCR | / | / | Geometric mean of miR-16-5p, miR-93-5p, miR-191-5p | [65] |
| Serum | OP (15), HC (15) | qPCR | miR-338-3p (↑), miR-3065-5p (↑) | OP vs. HC 0.74 (miR-338-3p), 0.87 (miR-3065-5p) | Cel-miR-39-3p | [66] |
| Plasma | OP with vertebral compression fractures (30), OP w/o vertebral fractures (30), HC (30) | Prescreening: miRNA array analysis Validation: qPCR | miR-19b-3p (↓) | OP w/o fracture vs. HC 0.9280 (miR-19b-3p) OP fracture vs. HC 0.9505 (miR-19b-3p) | U6 | [67] |
| Serum | No vertebral fractures/Low BMD (35), Vertebral fractures/Low BMD receiving treatment for osteoporosis (17), Vertebral fractures/Low BMD w/o any treatment for osteoporosis (24), HC (40) | qPCR | miR-375 (↑), miR-532-3p (↑), miR-19b-3p (↑), miR-152-3p (↑), miR-23a-3p (↑), miR-335-5p (↑), miR-21-5p (↑) | / | UniSp4 | [68] |
| Serum | Fragility femoral fractures (25), Osteoarthritis (25) | qPCR | miR-130a (↑), mi-204 (↓) | / | snU6 | [69] |
| Serum | WNT1 Mutation-positive OP (12), WNT1 Mutation-negative OP (12) | qPCR | miR-18a-3p (↑), miR-223-3p (↑), miR-22-3p (↓), miR-31-5p (↓), miR-34a-5p (↓), miR-143-5p (↓), miR-423-5p (↓), miR-423-3p (↓) | / | Global mean Ct value | [70] |
| Serum | / | Markov model | OsteomiRTM test | / | / | [71] |
| Serum | Fracture group (17), HC (16) | qPCR | OsteomiR° test | OP fracture vs. HC 0.687 (OsteomiR test) | UniSp4 | [72] |
| Serum | DM (20), DMFx (20), nondiabetic OP women with fractures (20), HC (20) | Low-density qPCR array platform | miR-550a-5p (↑), miR-188-3p (↓), miR-382-3p (↓) | DMFX vs. DM: 10 candidate miRNAs panels (model of 4 miRNAs) have an AUC value between 0.922 and 0.965; OP vs. HC 10 candidate miRNAs panels (model of 4 miRNAs) have an AUC value between 0.972 and 0.991 | Ct values were computed using the second derivative maximum method provided with the LC480 II software. | [73] |
| Serum | T1D (15), HC (14) | qPCR | miR-148a-3p (↑), miR-21-5p (↑) | / | miR-191, ath-miR-159a | [74] |
| Serum | Dmab/Fx+ (5), Fx + (5), Dmab/Fx− (5) | qPCR | miR-503 (↓), miR-222-2 (↓) | / | SNORD95A, SNORD96A, RNU6-2 | [75] |
| Serum | Low bone mass women treated with either Dmab (30) or TPDT (30) | qPCR | miR-33-3p (↓) miR-133a-3p (↓) | / | SNORD95A, SNORD96A, RNU6-2 | [76] |
| Study Population | Platforms for miRNAs Expression Profiling | Identified Candidate miRNAs in Human Circulating Monocytes | ROC Curve Analysis Data (AUC Value) | Study |
|---|---|---|---|---|
| High BMD (10), low BMD (10) | Prescreening: miRNA array Validation: qPCR | miR-133a (↑) | / | [77] |
| High BMD (10), low BMD (10) | Prescreening: miRNA array Validation: qPCR | miR-422a (↑) | / | [78] |
| OP (6), HC (6) | Prescreening: microarray Validation: qPCR | miR-1270 (↑) | / | [79] |
| OP (7), HC (7) | Prescreening: small RNA-sequencing Validation: qPCR | miRN-708-5p (↑) | / | [80] |
| High hip BMD (5), low hip BMD (5) | Bioinformatic analysis from microarray data | 38 miRNAs (see text) | / | [81] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donati, S.; Ciuffi, S.; Palmini, G.; Brandi, M.L. Circulating miRNAs: A New Opportunity in Bone Fragility. Biomolecules 2020, 10, 927. https://doi.org/10.3390/biom10060927
Donati S, Ciuffi S, Palmini G, Brandi ML. Circulating miRNAs: A New Opportunity in Bone Fragility. Biomolecules. 2020; 10(6):927. https://doi.org/10.3390/biom10060927
Chicago/Turabian StyleDonati, Simone, Simone Ciuffi, Gaia Palmini, and Maria Luisa Brandi. 2020. "Circulating miRNAs: A New Opportunity in Bone Fragility" Biomolecules 10, no. 6: 927. https://doi.org/10.3390/biom10060927
APA StyleDonati, S., Ciuffi, S., Palmini, G., & Brandi, M. L. (2020). Circulating miRNAs: A New Opportunity in Bone Fragility. Biomolecules, 10(6), 927. https://doi.org/10.3390/biom10060927