In Vitro Differentiation of Human Placenta-Derived Multipotent Cells into Schwann-Like Cells

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Culture of Placenta-Derived Multipotent Stem Cells
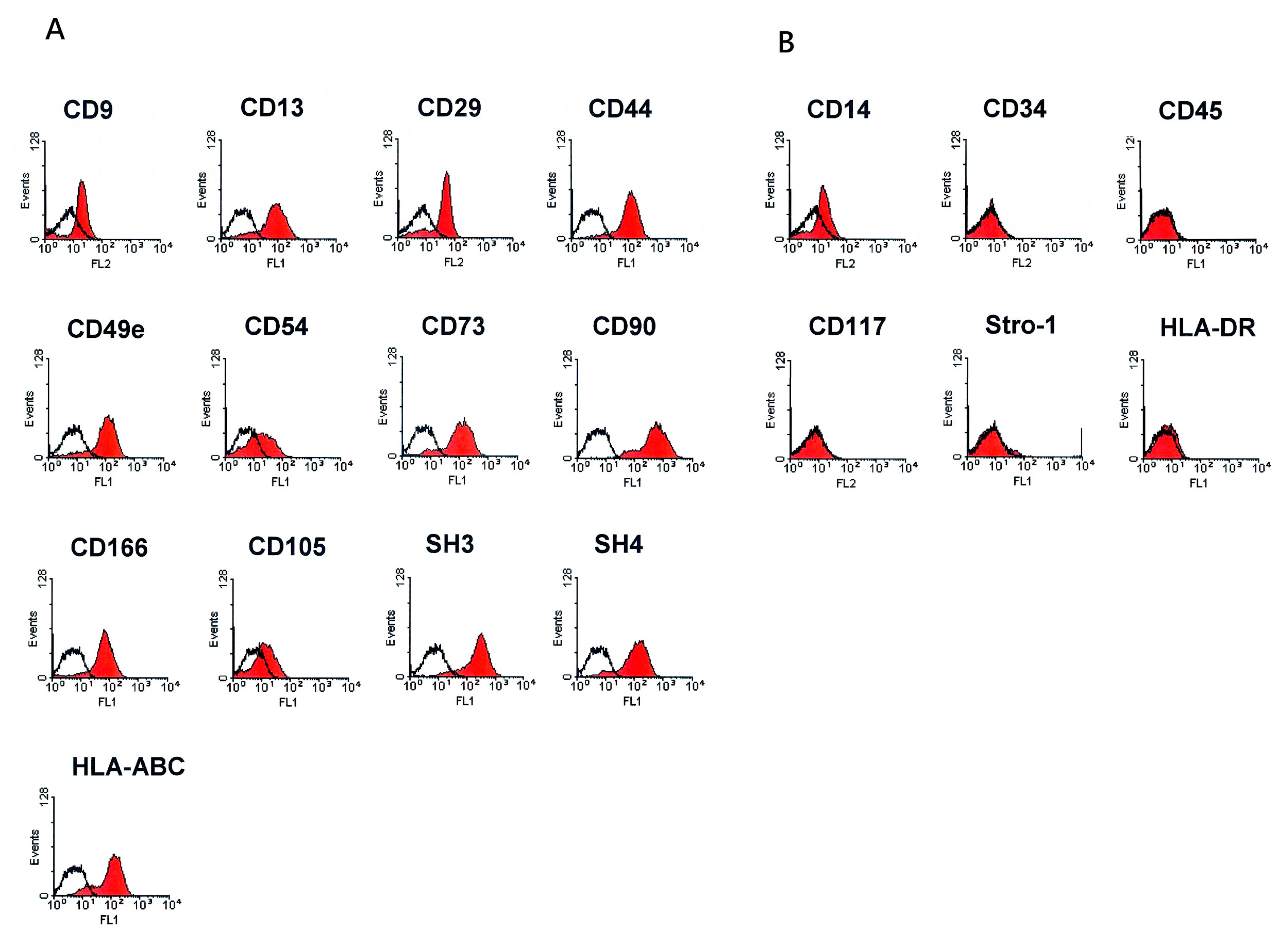
2.2. Flow Cytometry Analysis
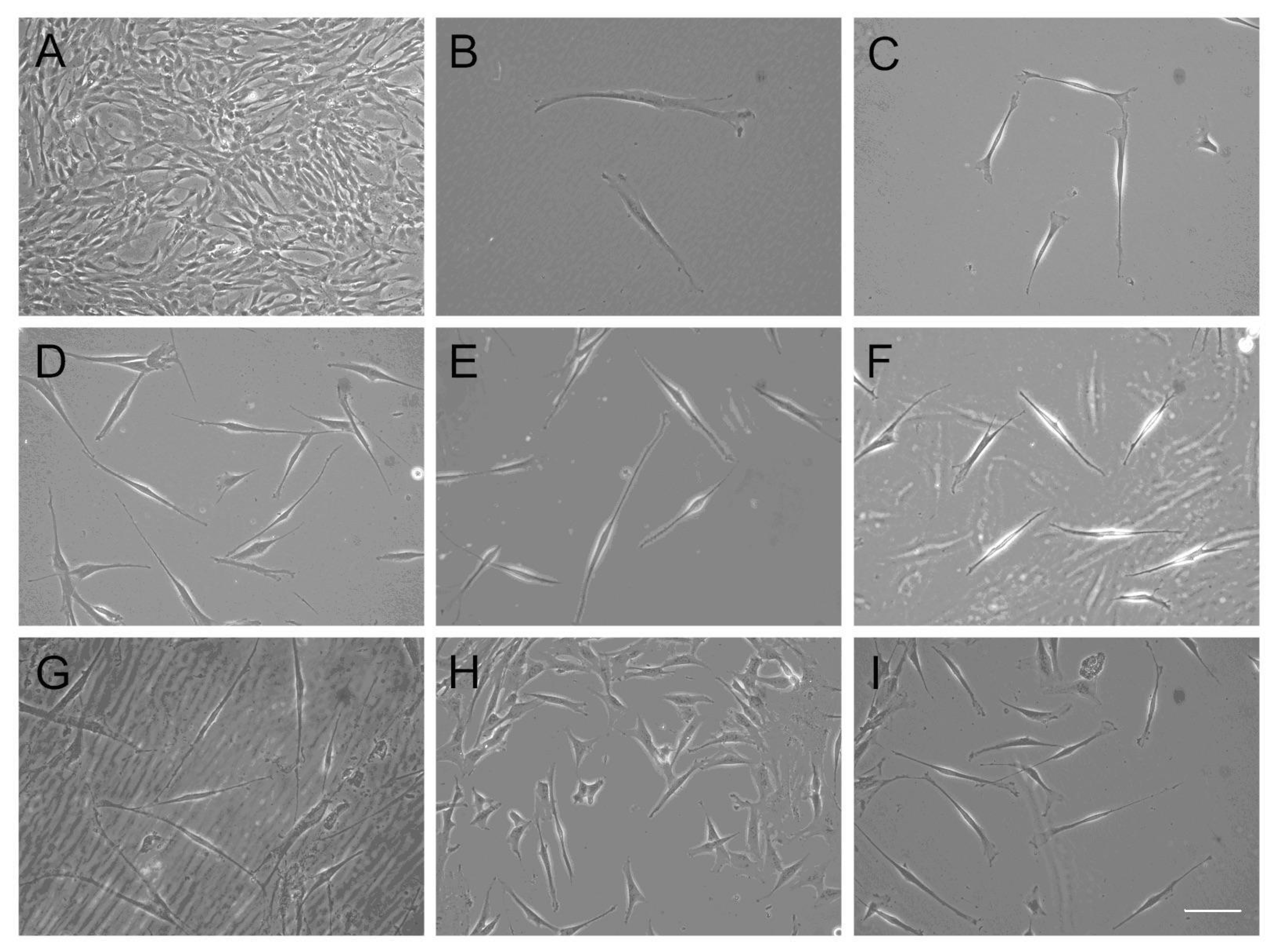
2.3. Induction of Differentiation into Schwann Cell-Like Cells
2.4. Quantitative Real-Time Polymerase Chain Reaction
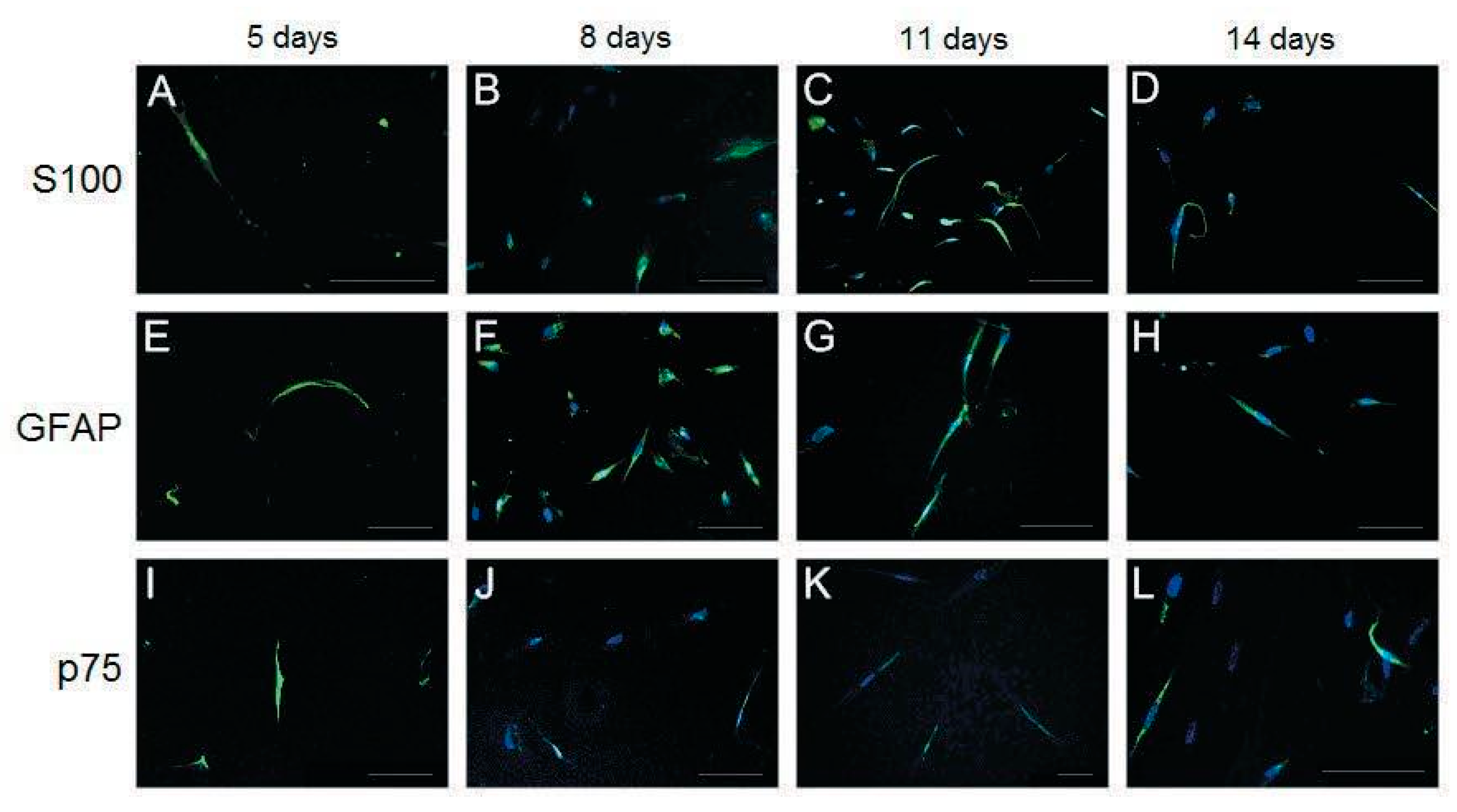
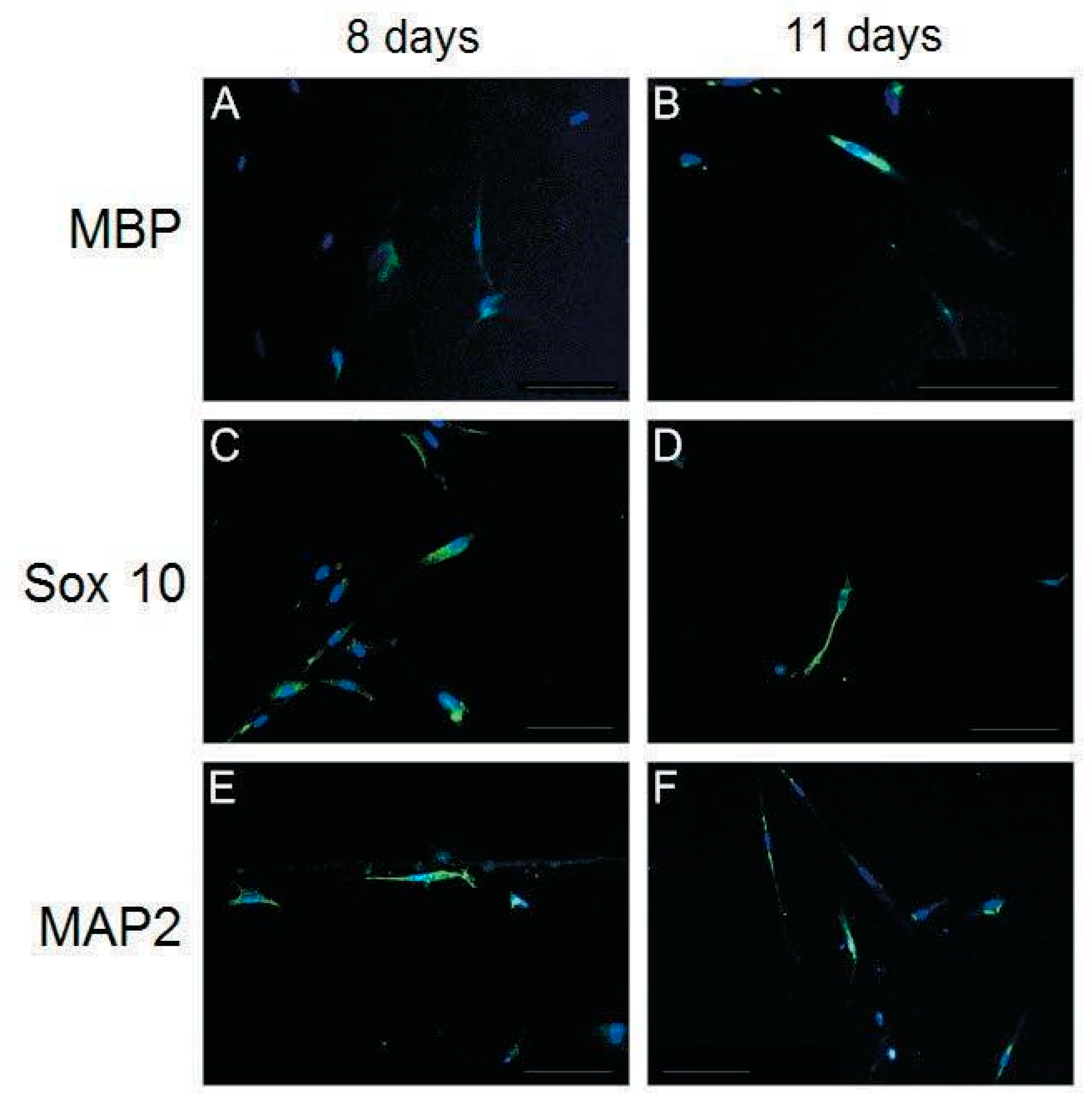
2.5. Immunofluorescence Analysis
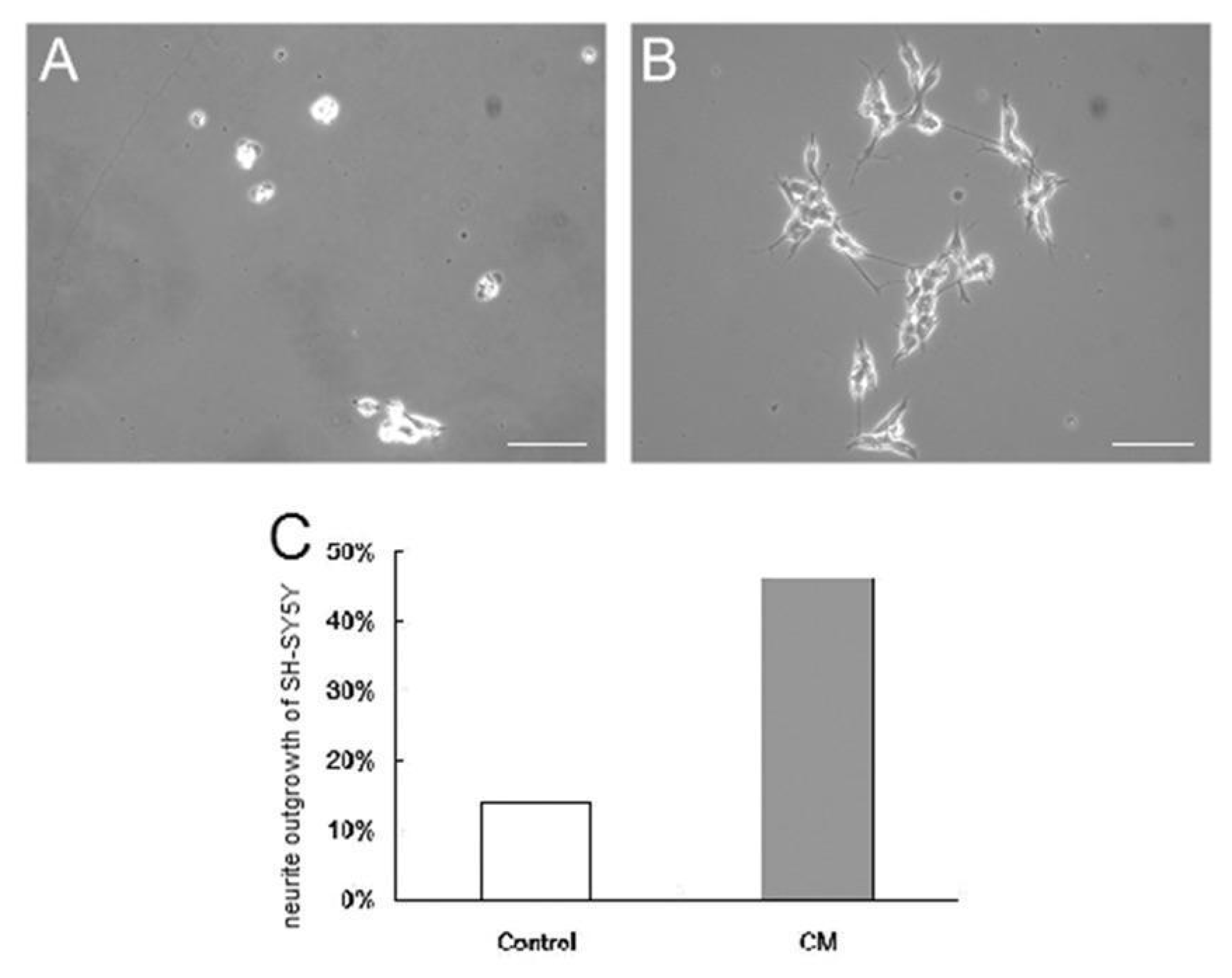
2.6. Conditioned Medium from Schwann Cell-Like Cells for SH-SY5Y Cell Culture
3. Results
3.1. Characterization of Human Placenta-Derived Multipotent Cells
3.2. Differentiation into Schwann Cell-Like Cells
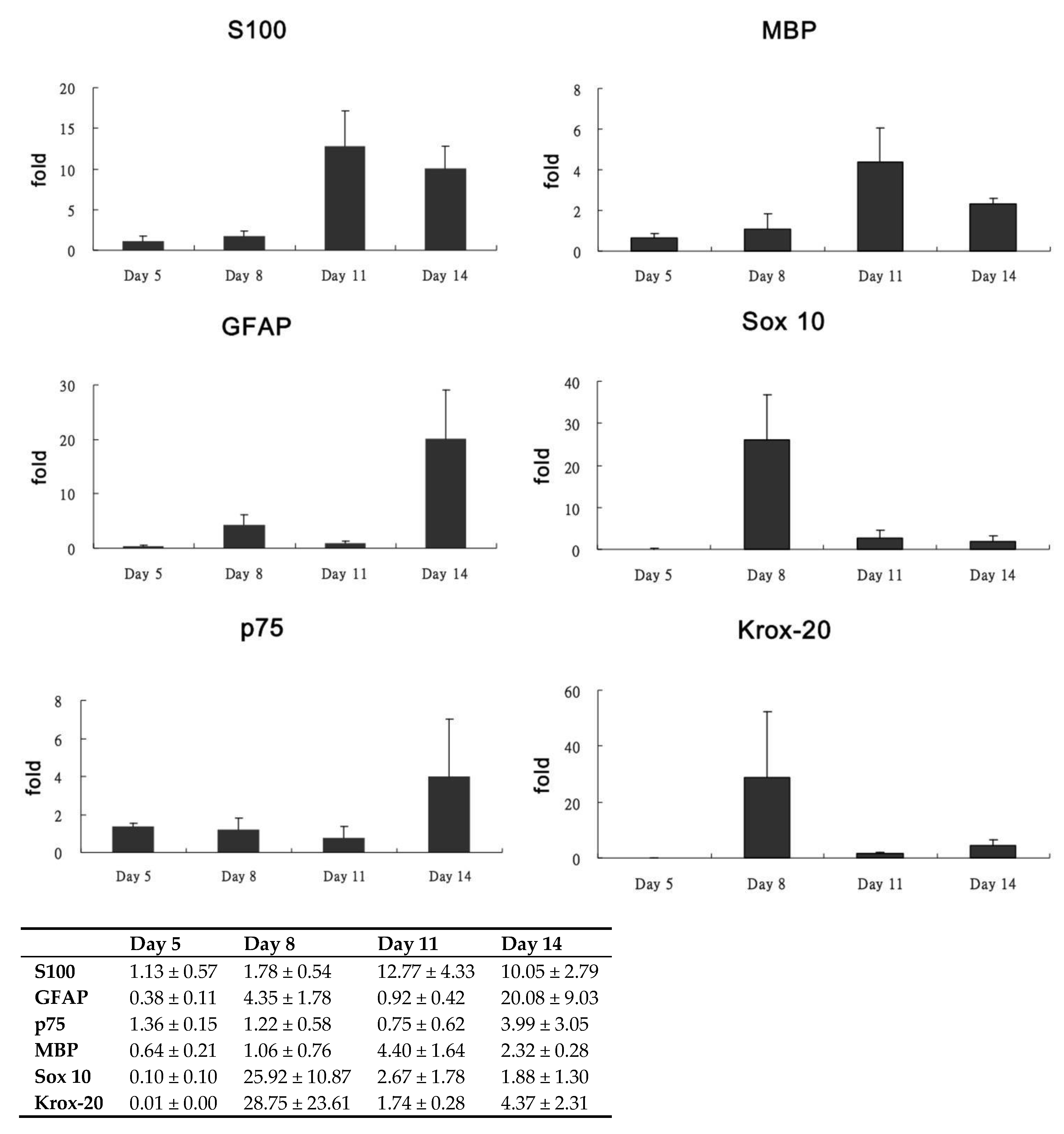
3.3. Characterization of Differentiated Placenta-Derived Multipotent Stem Cells
3.4. Culture of SH-SY5Y Cells in the Conditioned Medium from Differentiated Placenta-Derived Multipotent Stem Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Evans, G.R. Peripheral nerve injury: A review and approach to tissue engineered constructs. Anat. Rec. 2001, 263, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Wiberg, M.; Terenghi, G. Will it be possible to produce peripheral nerves? Surg. Technol. Int. 2003, 11, 303–310. [Google Scholar] [PubMed]
- Ruijs, A.C.J.; Jaquet, J.-B.; Kalmijn, S.; Giele, H.; Hovius, S.E.R. Median and Ulnar Nerve Injuries: A Meta-Analysis of Predictors of Motor and Sensory Recovery after Modern Microsurgical Nerve Repair. Plast. Reconstr. Surg. 2005, 116, 484–494. [Google Scholar] [CrossRef]
- Dezawa, M. Central and peripheral nerve regeneration by transplantation of Schwann cells and transdifferentiated bone marrow stromal cells. Anat. Sci. Int. 2002, 77, 12–25. [Google Scholar] [CrossRef]
- Frostick, S.P.; Yin, Q.; Kemp, G.J. Schwann cells, neurotrophic factors, and peripheral nerve regeneration. Microsurgery 1998, 18, 397–405. [Google Scholar] [CrossRef]
- Bhatheja, K.; Field, J. Schwann cells: Origins and role in axonal maintenance and regeneration. Int. J. Biochem. Cell Biol. 2006, 38, 1995–1999. [Google Scholar] [CrossRef]
- Harvey, A.R.; Plant, G.W.; Tan, M.M.L. Schwann cells and the regrowth of axons in the mammalian CNS: A review of transplantation studies in the rat visual system. Clin. Exp. Pharmacol. Physiol. 1995, 22, 569–579. [Google Scholar] [CrossRef]
- Bunge, M.B. Transplantation of purified populations of Schwann cells into lesioned adult rat spinal cord. J. Neurol. 1994, 242, S36–S39. [Google Scholar] [CrossRef]
- Keilhoff, G.; Goihl, A.; Langnäse, K.; Fansa, H.; Wolf, G. Transdifferentiation of mesenchymal stem cells into Schwann cell-like myelinating cells. Eur. J. Cell Biol. 2006, 85, 11–24. [Google Scholar] [CrossRef]
- Caddick, J.; Kingham, P.J.; Gardiner, N.J.; Wiberg, M.; Terenghi, G. Phenotypic and functional characteristics of mesenchymal stem cells differentiated along a Schwann cell lineage. Glia 2006, 54, 840–849. [Google Scholar] [CrossRef]
- Brohlin, M.; Mahay, D.; Novikov, L.N.; Terenghi, G.; Wiberg, M.; Shawcross, S.G.; Novikova, L.N. Characterisation of human mesenchymal stem cells following differentiation into Schwann cell-like cells. Neurosci. Res. 2009, 64, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Yen, B.L.; Chien, C.-C.; Chen, Y.-C.; Chen, J.-T.; Huang, J.-S.; Lee, F.-K.; Huang, H.-I. Placenta-Derived Multipotent Cells Differentiate into Neuronal and Glial Cells In Vitro. Tissue Eng. Part A 2008, 14, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Yen, B.L.; Huang, H.-I.; Chien, C.-C.; Jui, H.-Y.; Ko, B.; Yao, M.; Shun, C.-T.; Yen, M.-L.; Lee, M.-C.; Chen, Y.-C. Isolation of Multipotent Cells from Human Term Placenta. Stem Cells 2005, 23, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Matikainen, T.; Laine, J. Placenta—An alternative source of stem cells. Toxicol. Appl. Pharmacol. 2005, 207, 544–549. [Google Scholar] [CrossRef] [PubMed]
- Hall, F.L.; Fernyhough, P.; Ishii, D.N.; Vulliet, P.R. Suppression of nerve growth factor-directed neurite outgrowth in PC12 cells by sphingosine, an inhibitor of protein kinase C. J. Biol. Chem. 1988, 263, 4460–4466. [Google Scholar] [PubMed]
- Cheng, C.-C.; Chung, C.-A.; Su, L.-C.; Chien, C.-C.; Cheng, Y.-C. Osteogenic differentiation of placenta-derived multipotent cells in vitro. Taiwan. J. Obstet. Gynecol. 2014, 53, 187–192. [Google Scholar] [CrossRef]
- Chien, C.-C.; Yen, B.L.; Lee, F.-K.; Lai, T.-H.; Chen, Y.-C.; Chan, S.-H.; Huang, H.-I. In Vitro Differentiation of Human Placenta-Derived Multipotent Cells into Hepatocyte-Like Cells. Stem Cells 2006, 24, 1759–1768. [Google Scholar] [CrossRef]
- Woodbury, D.; Schwarz, E.J.; Prockop, D.J.; Black, I.B. Adult rat and human bone marrow stromal cells differentiate into neurons. J. Neurosci. Res. 2000, 61, 364–370. [Google Scholar] [CrossRef]
- Fraichard, A.; Chassande, O.; Bilbaut, G.; Dehay, C.; Savatier, P.; Samarut, J. In vitro differentiation of embryonic stem cells into glial cells and functional neurons. J. Cell Sci. 1995, 108 Pt 10, 3181–3188. [Google Scholar]
- Aquino, J.B.; Hjerling-Leffler, J.; Koltzenburg, M.; Edlund, T.; Villar, M.J.; Ernfors, P. In vitro and in vivo differentiation of boundary cap neural crest stem cells into mature Schwann cells. Exp. Neurol. 2006, 198, 438–449. [Google Scholar] [CrossRef]
- Mirsky, R.; Jessen, K.R.; Brennan, A.; Parkinson, D.; Dong, Z.; Meier, C.; Parmantier, E.; Lawson, D. Schwann cells as regulators of nerve development. J. Physiol. 2002, 96, 17–24. [Google Scholar] [CrossRef]
- Zorick, T.S.; Lemke, G. Schwann cell differentiation. Curr. Opin. Cell Biol. 1996, 8, 870–876. [Google Scholar] [CrossRef]
- Kim, H.A.; Ratner, N.; Roberts, T.M.; Stiles, C.D. Schwann Cell Proliferative Responses to cAMP andNf1Are Mediated by Cyclin D1. J. Neurosci. 2001, 21, 1110–1116. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rahmatullah, M.; Schroering, A.; Rothblum, K.; Stahl, R.C.; Urban, B.; Carey, D.J. Synergistic Regulation of Schwann Cell Proliferation by Heregulin and Forskolin. Mol. Cell. Biol. 1998, 18, 6245–6252. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Weir, M.D.; Xu, H.H. Effect of Cell Seeding Density on Proliferation and Osteodifferentiation of Umbilical Cord Stem Cells on Calcium Phosphate Cement-Fiber Scaffold. Tissue Eng. Part A 2011, 17, 2603–2613. [Google Scholar] [CrossRef] [PubMed]
- Mirsky, R.; Jessen, K.R. Schwann cell development, differentiation and myelination. Curr. Opin. Neurobiol. 1996, 6, 89–96. [Google Scholar] [CrossRef]
- Ghislain, J.; Charnay, P. Control of myelination in Schwann cells: A Krox20 cis -regulatory element integrates Oct6, Brn2 and Sox10 activities. EMBO Rep. 2006, 7, 52–58. [Google Scholar] [CrossRef]
- Lai, B.-Q.; Wang, J.-M.; Ling, E.-A.; Wu, J.-L.; Zeng, Y.-S. Graft of a Tissue-Engineered Neural Scaffold Serves as a Promising Strategy to Restore Myelination after Rat Spinal Cord Transection. Stem Cells Dev. 2014, 23, 910–921. [Google Scholar] [CrossRef]
- Ning, H.; Lin, G.; Lue, T.F.; Lin, C. Neuron-like differentiation of adipose tissue-derived stromal cells and vascular smooth muscle cells. Differentiation 2006, 74, 510–518. [Google Scholar] [CrossRef]
- Safford, K. Characterization of neuronal/glial differentiation of murine adipose-derived adult stromal cells. Exp. Neurol. 2004, 187, 319–328. [Google Scholar] [CrossRef]
- Mahay, D.; Terenghi, G.; Shawcross, S. Schwann cell mediated trophic effects by differentiated mesenchymal stem cells. Exp. Cell Res. 2008, 314, 2692–2701. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Chen, X.; Wang, X.; Liu, J.; Gu, X. Adult rat bone marrow stromal cells differentiate into Schwann cell-like cells in vitro. Vitr. Cell. Dev. Biol. Anim. 2008, 44, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Kingham, P.J.; Kalbermatten, D.F.; Mahay, D.; Armstrong, S.J.; Wiberg, M.; Terenghi, G. Adipose-derived stem cells differentiate into a Schwann cell phenotype and promote neurite outgrowth in vitro. Exp. Neurol. 2007, 207, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Britsch, S.; Goerich, D.E.; Riethmacher, D.; Peirano, R.I.; Rossner, M.; Nave, K.-A.; Birchmeier, C.; Wegner, M. The transcription factor Sox10 is a key regulator of peripheral glial development. Genes Dev. 2001, 15, 66–78. [Google Scholar] [CrossRef] [PubMed]
- Kuhlbrodt, K.; Herbarth, B.; Sock, E.; Hermans-Borgmeyer, I.; Wegner, M. Sox10, a Novel Transcriptional Modulator in Glial Cells. J. Neurosci. 1998, 18, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowski, J.L.; Rutkowski, J.L.; Yamashiro, D.J.; I Tennekoon, G.; Brodeur, G.M. Schwann cell-conditioned medium promotes neuroblastoma survival and differentiation. Cancer Res. 1998, 58, 4602–4606. [Google Scholar]
- Xu, Y.; Liu, Z.; Liu, L.; Zhao, C.; Xiong, F.; Zhou, C.; Li, Y.; Shan, Y.; Peng, F.; Zhang, C. Neurospheres from rat adipose-derived stem cells could be induced into functional Schwann cell-like cells in vitro. BMC Neurosci. 2008, 9, 21. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| S100β | AGACCAGGAAGGGGTGAGA | CCTCCCTTCCAGAATATTGGT |
| GFAP | GAAGGTTGAGTCGCTGGAGGAG | CGCTGTGAGGTCTGGCTTGG |
| p75 | CCAGCAGACCCATACGCAGAC | GCCAGATGTCGCCAGGTATCC |
| MBP | GCACAGAGACACGGGCATCC | CGGGCATGAGAAGGCAGAGG |
| Sox 10 | AACGGCGCCAGCAAAAGCAA | CGGGCATGAGAAGGCAGAGG |
| Krox-20 | CTCACGCCACTCTCCACCATC | CCTCACCGCCTCCACTTGC |
| GAPDH | ATGATTCTACCCACGGCAAG | CTGGAAGATGGTGATGGGTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Juan, C.-H.; Chen, M.-H.; Lin, F.-H.; Wong, C.-S.; Chien, C.-C.; Chen, M.-H. In Vitro Differentiation of Human Placenta-Derived Multipotent Cells into Schwann-Like Cells. Biomolecules 2020, 10, 1657. https://doi.org/10.3390/biom10121657
Juan C-H, Chen M-H, Lin F-H, Wong C-S, Chien C-C, Chen M-H. In Vitro Differentiation of Human Placenta-Derived Multipotent Cells into Schwann-Like Cells. Biomolecules. 2020; 10(12):1657. https://doi.org/10.3390/biom10121657
Chicago/Turabian StyleJuan, Chung-Hau, Mei-Hsiu Chen, Feng-Hui Lin, Chih-Shung Wong, Chih-Cheng Chien, and Ming-Hong Chen. 2020. "In Vitro Differentiation of Human Placenta-Derived Multipotent Cells into Schwann-Like Cells" Biomolecules 10, no. 12: 1657. https://doi.org/10.3390/biom10121657
APA StyleJuan, C.-H., Chen, M.-H., Lin, F.-H., Wong, C.-S., Chien, C.-C., & Chen, M.-H. (2020). In Vitro Differentiation of Human Placenta-Derived Multipotent Cells into Schwann-Like Cells. Biomolecules, 10(12), 1657. https://doi.org/10.3390/biom10121657