RNF11 at the Crossroads of Protein Ubiquitination



Abstract
1. Introduction
2. RNF11, A Multifunctional RING-H2 Ligase
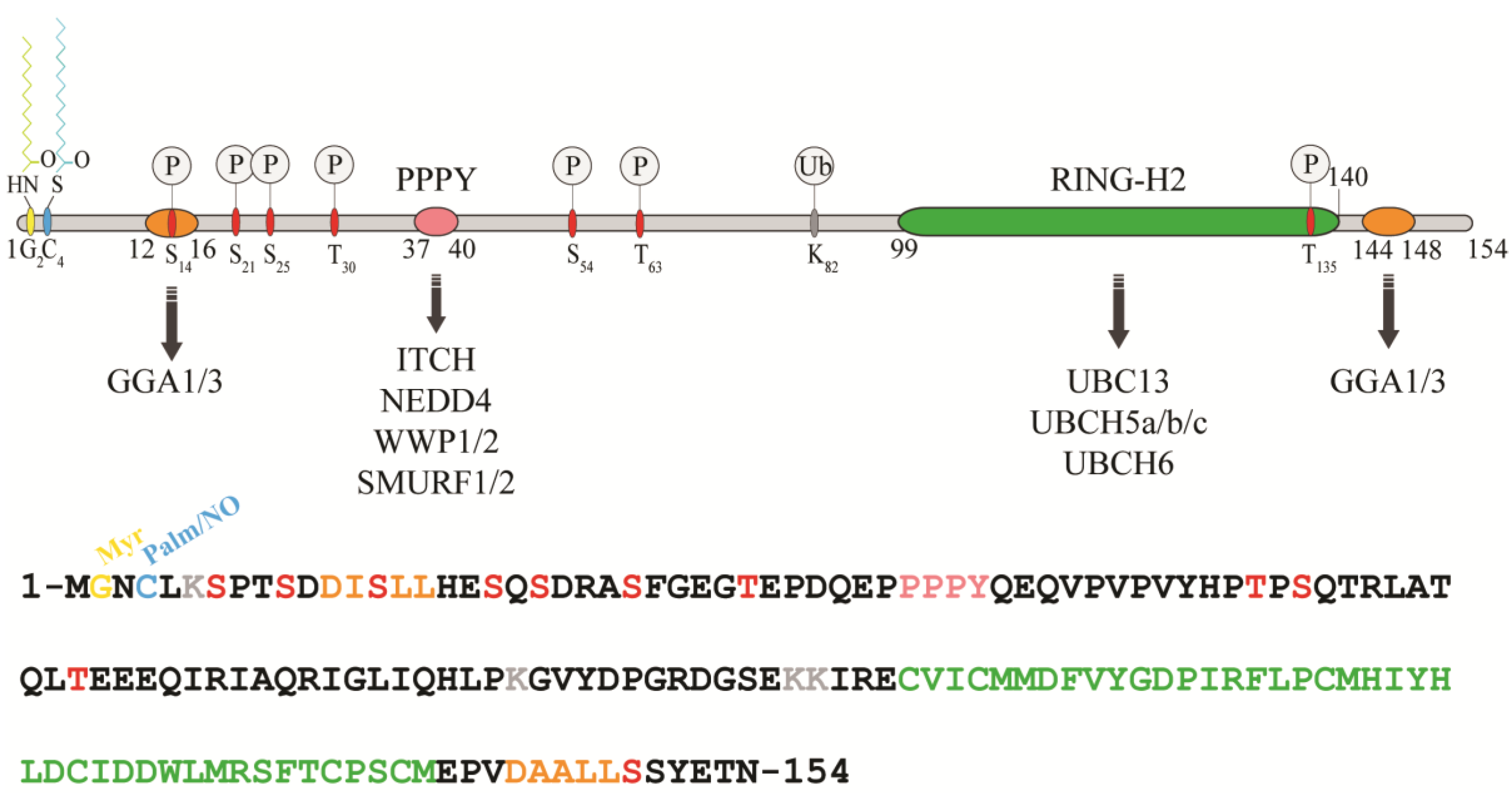
2.1. Domain Organization and Cellular Localization of RNF11
2.2. RNF11 Post-Translational Modifications and Functional Consequences
2.2.1. S-Nitrosylation
2.2.2. Ubiquitination
2.2.3. Sumoylation
2.2.4. Phosphorylation
2.3. The RNF11 Interaction Network
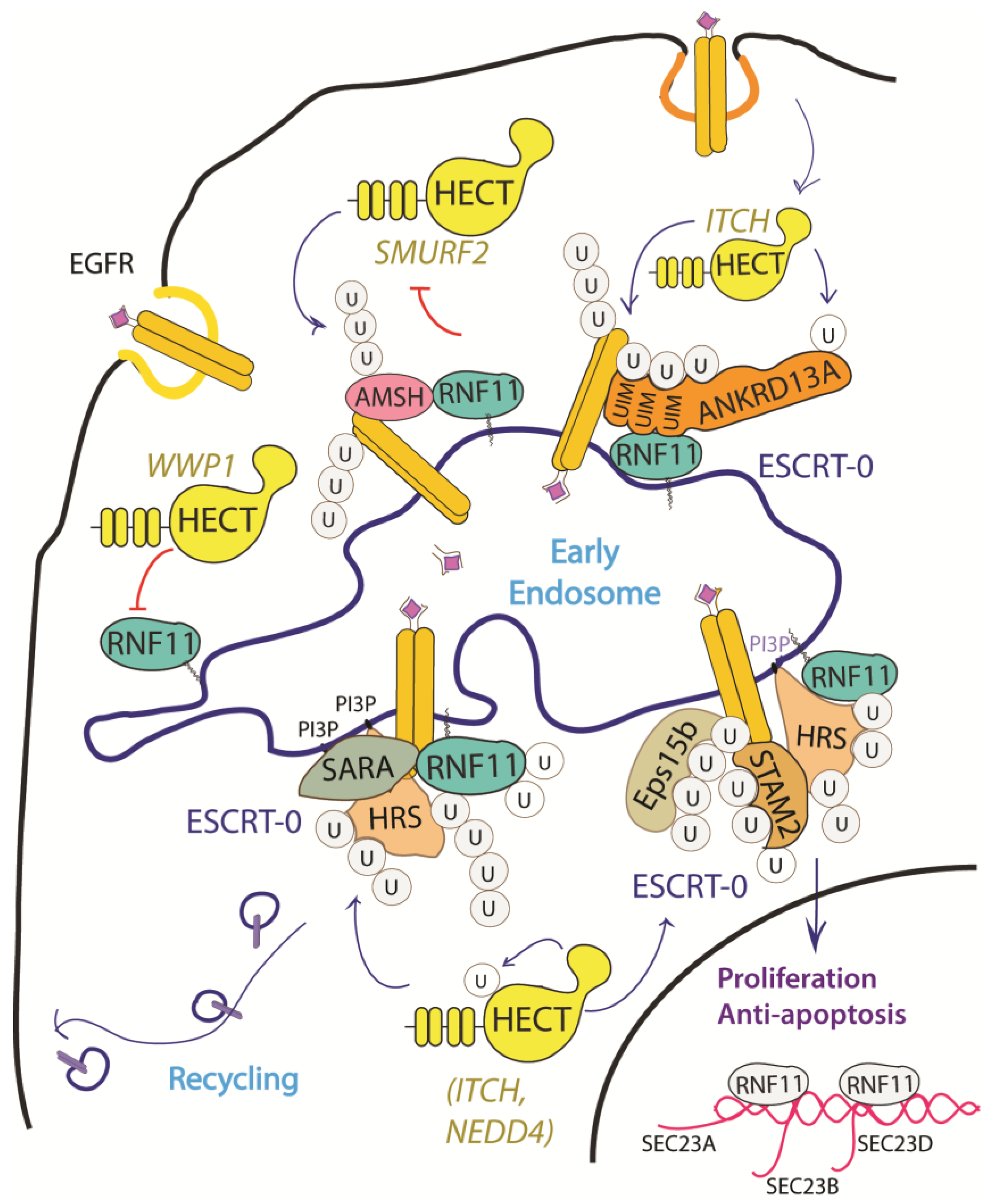
2.4. RNF11 in Intracellular Traffic, Signal Transduction and Oncogenesis
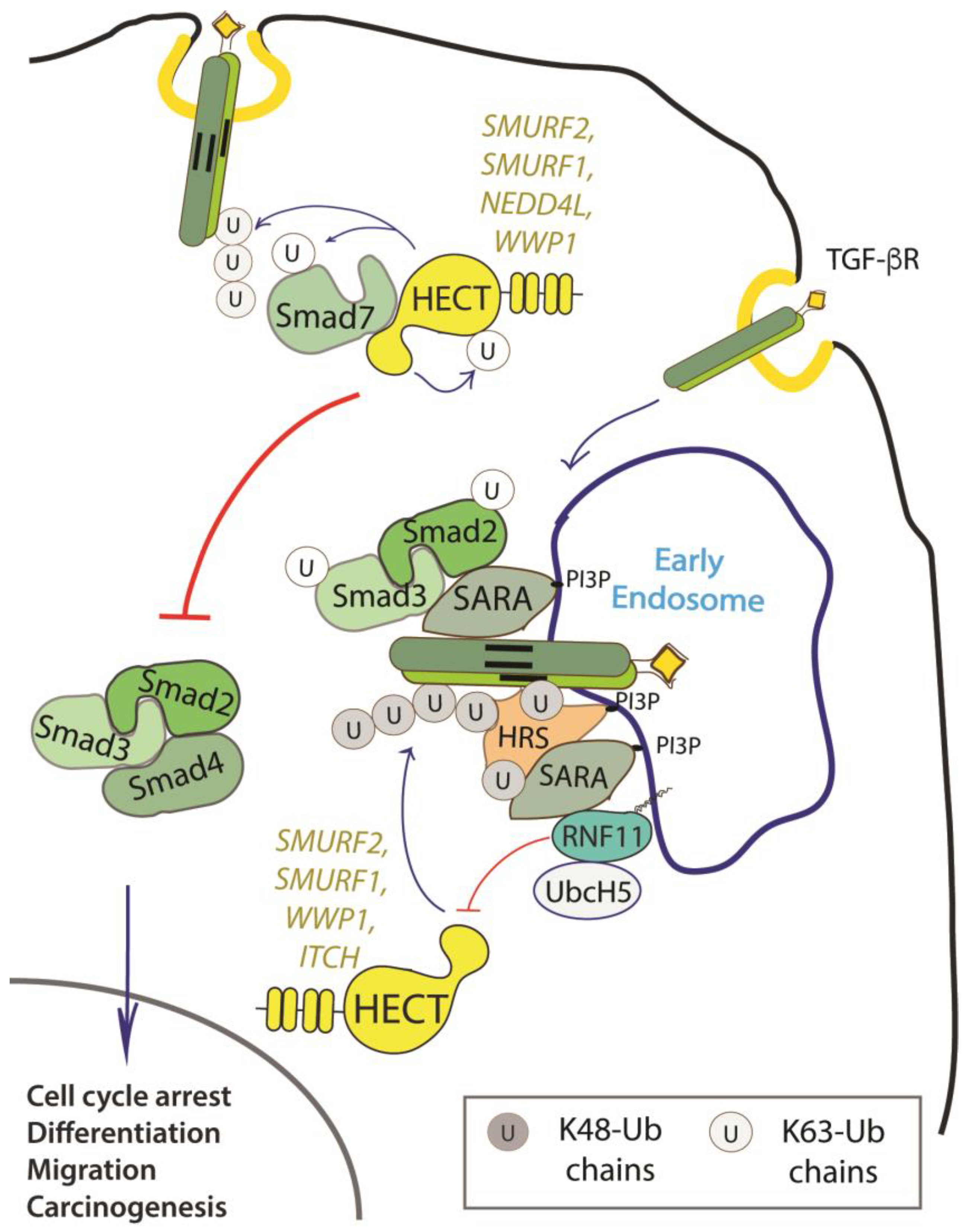
2.4.1. The TGF-Beta Signaling Pathway
2.4.2. The EGF Receptor Signaling Pathway
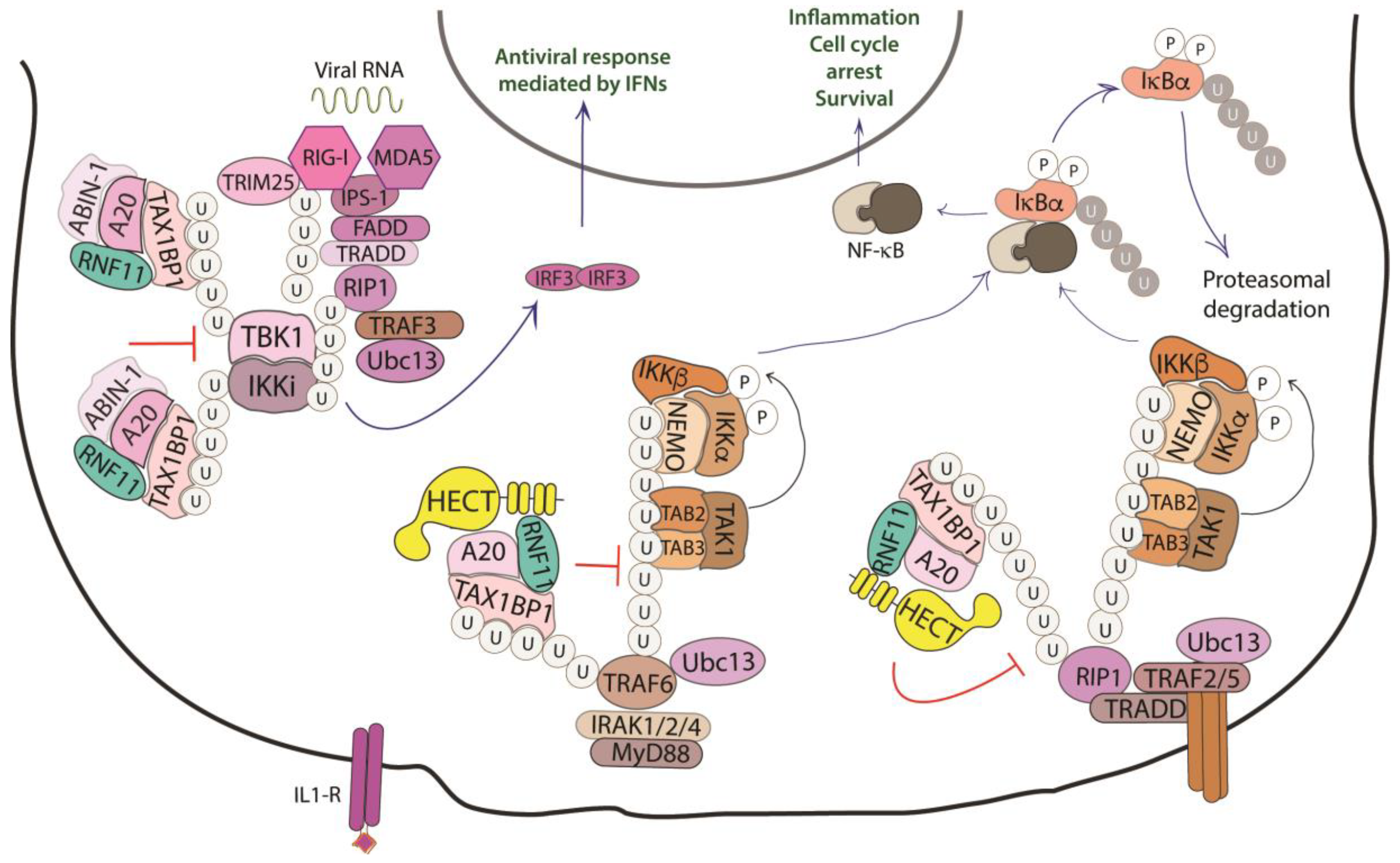
2.4.3. The NF-kB Signaling Pathway
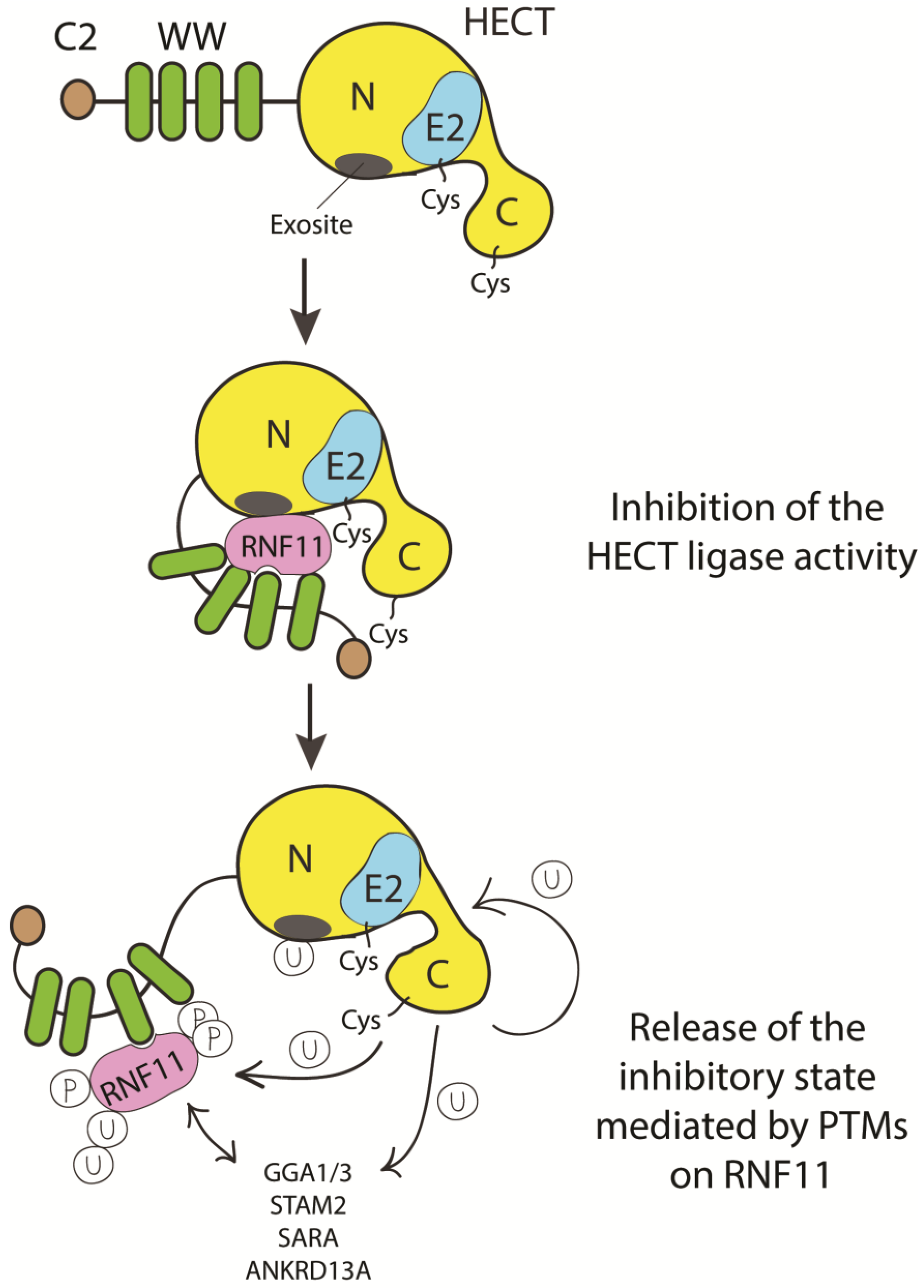
2.5. RNF11, a Novel Regulator of E2s and E3s Activity
2.6. Pathogenic Significance of RNF11 and Future Perspectives in Target Therapy
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schnell, J.D.; Hicke, L. Non-traditional functions of ubiquitin and ubiquitin-binding proteins. J. Biol. Chem. 2003, 278, 35857–35860. [Google Scholar] [CrossRef]
- Kimura, Y.; Tanaka, K. Regulatory mechanisms involved in the control of ubiquitin homeostasis. J. Biochem. 2010, 147, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Deshaies, R.J.; Joazeiro, C.A.P. RING domain E3 ubiquitin ligases. Annu. Rev. Biochem. 2009, 78, 399–434. [Google Scholar] [CrossRef] [PubMed]
- Metzger, M.B.; Pruneda, J.N.; Klevit, R.E.; Weissman, A.M. RING-type E3 ligases: Master manipulators of E2 ubiquitin-conjugating enzymes and ubiquitination. BBA Mol. Cell Res. 2014, 1843, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Spratt, D.E.; Walden, H.; Shaw, G.S. RBR E3 ubiquitin ligases: New structures, new insights, new questions. Biochem. J. 2014, 458, 421–437. [Google Scholar] [CrossRef]
- Scheffner, M.; Kumar, S. Mammalian HECT ubiquitin-protein ligases: Biological and pathophysiological aspects. BBA Mol. Cell Res. 2014, 1843, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Cadwell, K.; Coscoy, L. Ubiquitination on nonlysine residues by a viral E3 ubiquitin ligase. Science 2005, 309, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Ciechanover, A. N-terminal ubiquitination: More protein substrates join in. Trends Cell Biol. 2004, 14, 103–106. [Google Scholar] [CrossRef]
- Komander, D.; Rape, M. The ubiquitin code. Annu. Rev. Biochem. 2012, 81, 203–229. [Google Scholar] [CrossRef]
- Swatek, K.N.; Komander, D. Ubiquitin modifications. Cell Res. 2016, 26, 399–422. [Google Scholar] [CrossRef]
- Eletr, Z.M.; Wilkinson, K.D. Regulation of proteolysis by human deubiquitinating enzymes. BBA Mol. Cell Res. 2013, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Budhidarmo, R.; Zhu, J.; Middleton, A.J.; Day, C.L. The RING domain of RING Finger 11 (RNF11) protein binds Ubc13 and inhibits formation of polyubiquitin chains. FEBS Lett 2018, 592, 1434–1444. [Google Scholar] [CrossRef] [PubMed]
- Kitching, R.E.; Wong, M.J.; Koehler, D.; Burger, A.M.; Landberg, G.; Gish, G.; Seth, A.K. The RING-H2 protein RNF11 is differentially expressed in breast tumours and interacts with HECT-type E3 ligases. BBA Mol. Basis Dis. 2003, 1639, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Connor, M.K.; Seth, A.K. A central role for the ring finger protein RNF11 in ubiquitin-mediated proteolysis via interactions with E2s and E3s. Oncogene 2004, 23, 2089–2095. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Li, H.; Seth, A.K. An RNF11: Smurf2 complex mediates ubiquitination of the AMSH protein. Oncogene 2004, 23, 1801–1808. [Google Scholar] [CrossRef] [PubMed]
- Burger, A.M.; Amemiya, Y.; Kitching, R.E.; Seth, A.K. Novel RING E3 ubiquitin ligases in breast cancer. Neoplasia 2006, 8, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Santonico, E.; Belleudi, F.; Panni, S.; Torrisi, M.R.; Cesareni, G.; Castagnoli, L. Multiple modification and protein interaction signals drive the Ring finger protein 11 (RNF11) E3 ligase to the endosomal compartment. Oncogene 2010, 29, 5604–5618. [Google Scholar] [CrossRef]
- Santonico, E.; Mattioni, A.; Panni, S.; Belleudi, F.; Mattei, M.; Torrisi, M.R.; Castagnoli, L.; Cesareni, G. RNF11 is a GGA protein cargo and acts as a molecular adaptor for GGA3 ubiquitination mediated by Itch. Oncogene 2014, 34, 3377–3390. [Google Scholar] [CrossRef]
- Connor, M.K.; Azmi, P.B.; Subramaniam, V.; Li, H. Molecular characterization of Ring Finger Protein 11. Mol. Cancer Res. 2005, 3, 453–461. [Google Scholar] [CrossRef]
- Wright, J.D.; Mace, P.D.; Day, C.L. Noncovalent ubiquitin interactions regulate the catalytic activity of ubiquitin writers. Trends Biochem. Sci. 2016, 41, 924–937. [Google Scholar] [CrossRef]
- Trempe, J.-F.; Sauve, V.; Grenier, K.; Seirafi, M.; Tang, M.Y.; Menade, M.; Al-Abdul-Wahid, S.; Krett, J.; Wong, K.; Kozlov, G.; et al. Structure of Parkin Reveals Mechanisms for Ubiquitin Ligase Activation. Science 2013, 340, 1451–1455. [Google Scholar] [CrossRef] [PubMed]
- Rennie, M.L.; Chaugule, V.K.; Walden, H. Modes of allosteric regulation of the ubiquitination machinery. Curr. Opin. Struct. Biol. 2020, 62, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.D.; Mace, P.D.; Day, C.L. Secondary ubiquitin-RING docking enhances Arkadia and Ark2C E3 ligase activity. Nat. Struct. Mol. Biol. 2015, 23, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Kane, L.A.; Lazarou, M.; Fogel, A.I.; Li, Y.; Yamano, K.; Sarraf, S.A.; Banerjee, S.; Youle, R.J. PINK1 phosphorylates ubiquitin to activate Parkin E3 ubiquitin ligase activity. J. Cell Biol. 2014, 205, 143–153. [Google Scholar] [CrossRef]
- Kazlauskaite, A.; Kondapalli, C.; Gourlay, R.; Campbell, D.G.; Ritorto, M.S.; Hofmann, K.; Alessi, D.R.; Knebel, A.; Trost, M.; Muqit, M.M.K. Parkin is activated by PINK1-dependent phosphorylation of ubiquitin at Ser65. Biochem. J. 2014, 460, 127–141. [Google Scholar] [CrossRef]
- Koyano, F.; Okatsu, K.; Kosako, H.; Tamura, Y.; Go, E.; Kimura, M.; Kimura, Y.; Tsuchiya, H.; Yoshihara, H.; Hirokawa, T.; et al. Ubiquitin is phosphorylated by PINK1 to activate parkin. Nature 2014, 510, 162–166. [Google Scholar] [CrossRef]
- Farazi, T.A.; Waksman, G.; Gordon, J.I. The biology and enzymology of protein N-myristoylation. J. Biol. Chem. 2001, 276, 39501–39504. [Google Scholar] [CrossRef]
- Linder, M.E.; Deschenes, R.J. Palmitoylation: Policing protein stability and traffic. Nat. Rev. Mol. Cell Biol. 2007, 8, 74–84. [Google Scholar] [CrossRef]
- Colland, F.; Jacq, X.; Trouplin, V.; Mougin, C.; Groizeleau, C.; Hamburger, A.; Meil, A.; Wojcik, J.; Legrain, P.; Gauthier, J.-M. Functional Proteomics Mapping of a Human Signaling Pathway. Genome Res. 2004, 14, 1324–1332. [Google Scholar] [CrossRef]
- Kostaras, E.; Pedersen, N.M.; Stenmark, H.; Fotsis, T.; Murphy, C. SARA and RNF11 at the Crossroads of EGFR Signaling and Trafficking, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2014; Volume 535, pp. 225–247. [Google Scholar]
- Mattioni, A.; Boldt, K.; Auciello, G.; Komada, M.; Rappoport, J.Z.; Ueffing, M.; Castagnoli, L.; Cesareni, G.; Santonico, E. Ring Finger Protein 11 (RNF11) acts on ligand-activated EGFR via the direct interaction with the UIM region of ANKRD13 protein family. FEBS J. 2020, 1–46. [Google Scholar]
- Scharaw, S.; Iskar, M.; Ori, A.; Boncompain, G.; Laketa, V.; Poser, I.; Lundberg, E.; Perez, F.; Beck, M.; Bork, P.; et al. The endosomal transcriptional regulator RNF11 integrates degradation and transport of EGFR. J. Cell Biol. 2016, 215, 543–558. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, V.; Li, H.; Wong, M.J.; Kitching, R.E.; Attisano, L.; Wrana, J.L.; Zubovits, J.; Burger, A.M.; Seth, A.K. The RING-H2 protein RNF11 is overexpressed in breast cancer and is a target of Smurf2 E3 ligase. Br. J. Cancer 2003, 89, 1538–1544. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-I.; Giovinazzo, D.; Kang, H.C.; Lee, Y.; Jeong, J.S.; Doulias, P.-T.; Xie, Z.; Hu, J.; Ghasemi, M.; Ischiropoulos, H.; et al. Protein Microarray Characterization of the S-Nitrosoproteome. Mol. Cell. Proteom. 2014, 13, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Mengel, A.; Chaki, M.; Shekariesfahlan, A.; Lindermayr, C. Effect of nitric oxide on gene transcription—S-nitrosylation of nuclear proteins. Front. Plant Sci. 2013, 4, 293. [Google Scholar] [CrossRef]
- Wagner, S.A.; Beli, P.; Weinert, B.T.; Schölz, C.; Kelstrup, C.D.; Young, C.; Nielsen, M.L.; Olsen, J.V.; Brakebusch, C.; Choudhary, C. Proteomic Analyses Reveal Divergent Ubiquitylation Site Patterns in Murine Tissues. Mol. Cell. Proteom. 2012, 11, 1578–1585. [Google Scholar] [CrossRef]
- Markson, G.; Kiel, C.; Hyde, R.; Brown, S.; Charalabous, P.; Bremm, A.; Semple, J.; Woodsmith, J.; Duley, S.; Salehi-Ashtiani, K.; et al. Analysis of the human E2 ubiquitin conjugating enzyme protein interaction network. Genome Res. 2009, 19, 1905–1911. [Google Scholar] [CrossRef]
- Malonis, R.J.; Fu, W.; Jelcic, M.J.; Thompson, M.; Canter, B.S.; Tsikitis, M.; Esteva, F.J.; Sánchez, I. RNF11 sequestration of the E3 ligase SMURF2 on membranes antagonizes SMAD7 down-regulation of transforming growth factor β signaling. J. Biol. Chem. 2017, 292, 7435–7451. [Google Scholar] [CrossRef]
- Chen, C.; Zhou, Z.; Liu, R.; Li, Y.; Azmi, P.B.; Seth, A.K. The WW domain containing E3 ubiquitin protein ligase 1 upregulates ErbB2 and EGFR through RING finger protein 11. Oncogene 2008, 27, 6845–6855. [Google Scholar] [CrossRef]
- Azmi, P.B.; Seth, A.K. RNF11 is a multifunctional modulator of growth factor receptor signalling and transcriptional regulation. Eur. J. Cancer 2005, 41, 2549–2560. [Google Scholar] [CrossRef]
- Gâtel, P.; Brockly, F.; Reynes, C.; Pastore, M.; Hicheri, Y.; Cartron, G.; Piechaczyk, M.; Bossis, G. Ubiquitin and SUMO conjugation as biomarkers of acute myeloid leukemias response to chemotherapies. Life Sci. Alliance 2020, 3, e201900577-11. [Google Scholar] [CrossRef]
- Melchior, F.; Schergaut, M.; Pichler, A. SUMO: Ligases, isopeptidases and nuclear pores. Trends Biochem. Sci. 2003, 28, 612–618. [Google Scholar] [CrossRef] [PubMed]
- Gouw, M.; Michael, S.; Sámano-Sánchez, H.; Kumar, M.; Zeke, A.; Lang, B.; Bely, B.; Chemes, L.B.; Davey, N.E.; Deng, Z.; et al. The eukaryotic linear motif resource—2018 update. Nucleic Acids Res. 2017, 46, D428–D434. [Google Scholar] [CrossRef] [PubMed]
- Blom, N.; Sicheritz-Pontén, T.; Gupta, R.; Gammeltoft, S.; Brunak, S. Prediction of post-translational glycosylation and phosphorylation of proteins from the amino acid sequence. Proteomics 2004, 4, 1633–1649. [Google Scholar] [CrossRef]
- Huttlin, E.L.; Jedrychowski, M.P.; Elias, J.E.; Goswami, T.; Rad, R.; Beausoleil, S.A.; Villén, J.; Haas, W.; Sowa, M.E.; Gygi, S.P. A Tissue-Specific Atlas of Mouse Protein Phosphorylation and Expression. Cell 2010, 143, 1174–1189. [Google Scholar] [CrossRef] [PubMed]
- Weintz, G.; Olsen, J.V.; Frühauf, K.; Niedzielska, M.; Amit, I.; Jantsch, J.; Mages, J.; Frech, C.; Dölken, L.; Mann, M.; et al. The phosphoproteome of toll-like receptor-activated macrophages. Mol. Syst. Biol. 2010, 6, 371. [Google Scholar] [CrossRef]
- Wiśniewski, J.R.; Nagaraj, N.; Zougman, A.; Gnad, F.; Mann, M. Brain Phosphoproteome Obtained by a FASP-Based Method Reveals Plasma Membrane Protein Topology. J. Proteome Res. 2010, 9, 3280–3289. [Google Scholar] [CrossRef]
- Trinidad, J.C.; Barkan, D.T.; Gulledge, B.F.; Thalhammer, A.; Sali, A.; Schoepfer, R.; Burlingame, A.L. Global Identification and Characterization of Both O-GlcNAcylation and Phosphorylation at the Murine Synapse. Mol. Cell. Proteom. 2012, 11, 215–229. [Google Scholar] [CrossRef]
- Bian, Y.; Song, C.; Cheng, K.; Dong, M.; Wang, F.; Huang, J.; Sun, D.; Wang, L.; Ye, M.; Zou, H. An enzyme assisted RP-RPLC approach for in-depth analysis of human liver phosphoproteome. J. Proteom. 2014, 96, 253–262. [Google Scholar] [CrossRef]
- Bivona, T.G.; Quatela, S.E.; Bodemann, B.O.; Ahearn, I.M.; Soskis, M.J.; Mor, A.; Miura, J.; Wiener, H.H.; Wright, L.; Saba, S.G.; et al. PKC Regulates a Farnesyl-Electrostatic Switch on K-Ras that Promotes its Association with Bcl-Xl on Mitochondria and Induces Apoptosis. Mol. Cell 2006, 21, 481–493. [Google Scholar] [CrossRef]
- Bernassola, F.; Karin, M.; Ciechanover, A.; Melino, G. The HECT Family of E3 Ubiquitin Ligases: Multiple Players in Cancer Development. Cancer Cell 2008, 14, 10–21. [Google Scholar] [CrossRef]
- Dunn, R.; Klos, D.A.; Adler, A.S.; Hicke, L. The C2 domain of the Rsp5 ubiquitin ligase binds membrane phosphoinositides and directs ubiquitination of endosomal cargo. J. Cell Biol. 2004, 165, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Scheper, J.; Oliva, B.; Villà-Freixa, J.; Thomson, T.M. Analysis of electrostatic contributions to the selectivity of interactions between RING-finger domains and ubiquitin-conjugating enzymes. Proteins 2009, 74, 92–103. [Google Scholar] [CrossRef]
- Castagnoli, L.; Mandaliti, W.; Nepravishta, R.; Valentini, E.; Mattioni, A.; Procopio, R.; Iannuccelli, M.; Polo, S.; Paci, M.; Cesareni, G.; et al. Selectivity of the CUBAN domain in the recognition of ubiquitin and NEDD8. FEBS J. 2018, 286, 653–677. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, S. Structural mechanisms of HECT-type ubiquitin ligases. Biol. Chem. 2018, 399, 127–145. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.C.; Joggerst, B.; Laketa, V.; Verissimo, F.; Cetin, C.; Erfle, H.; Bexiga, M.G.; Singan, V.R.; Hériché, J.-K.; Neumann, B.; et al. Genome-wide RNAi screening identifies human proteins with a regulatory function in the early secretory pathway. Nat. Cell Biol. 2012, 14, 764–774. [Google Scholar] [CrossRef] [PubMed]
- Kostaras, E.; Sflomos, G.; Pedersen, N.M.; Stenmark, H.; Fotsis, T.; Murphy, C. SARA and RNF11 interact with each other and ESCRT-0 core proteins and regulate degradative EGFR trafficking. Oncogene 2012, 32, 5220–5232. [Google Scholar] [CrossRef] [PubMed]
- Hurley, J.H. Nipped in the Bud: How the AMSH MIT Domain Helps Deubiquitinate Lysosome-Bound Cargo. Structure 2011, 19, 1033–1035. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Roxrud, I.; Raiborg, C.; Pedersen, N.M.; Stang, E.; Stenmark, H. An endosomally localized isoform of Eps15 interacts with Hrs to mediate degradation of epidermal growth factor receptor. J. Cell Biol. 2008, 180, 1205–1218. [Google Scholar] [CrossRef]
- Henne, W.M.; Buchkovich, N.J.; Emr, S.D. The ESCRT Pathway. Dev. Cell 2011, 21, 77–91. [Google Scholar] [CrossRef]
- Acconcia, F.; Sigismund, S.; Polo, S. Ubiquitin in trafficking: The network at work. Exp. Cell Res. 2009, 315, 1610–1618. [Google Scholar] [CrossRef]
- Katz, M.; Shtiegman, K.; Tal-Or, P.; Yakir, L.; Mosesson, Y.; Harari, D.; Machluf, Y.; Asao, H.; Jovin, T.; Sugamura, K.; et al. Ligand-Independent Degradation of Epidermal Growth Factor Receptor Involves Receptor Ubiquitylation and Hgs, an Adaptor Whose Ubiquitin- Interacting Motif Targets Ubiquitylation by Nedd4. Traffic 2002, 3, 740–751. [Google Scholar] [CrossRef]
- Holleman, J.; Marchese, A. The ubiquitin ligase deltex-3l regulates endosomal sorting of the G protein–coupled receptor CXCR4. Mol. Biol. Cell 2014, 25, 1892–1904. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Baker, D.; ten Dijke, P. TGF-β-Mediated Epithelial-Mesenchymal Transition and Cancer Metastasis. IJMS 2019, 20, 2767. [Google Scholar] [CrossRef] [PubMed]
- Azmi, P.B.; Seth, A.K. The RING Finger Protein 11 Binds to Smad4 and Enhances Smad4-dependant TGF-β Signalling. Anticancer Res. 2009, 29, 2253–2264. [Google Scholar]
- Izzi, L.; Attisano, L. Ubiquitin-Dependent Regulation of TGβ Signaling in Cancer. Neoplasia 2006, 8, 677–688. [Google Scholar] [CrossRef]
- Itoh, S.; Dijke, P.T. Negative regulation of TGF-β receptor/Smad signal transduction. Curr. Opin. Cell Biol. 2007, 19, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Tomas, A.; Futter, C.E.; Eden, E.R. EGF receptor trafficking: Consequences for signaling and cancer. Trends Cell Biol. 2014, 24, 26–34. [Google Scholar] [CrossRef]
- McCullough, J.; Clague, M.J.; Urbé, S. AMSH is an endosome-associated ubiquitin isopeptidase. J. Cell Biol. 2004, 166, 487–492. [Google Scholar] [CrossRef]
- Zhi, X.; Chen, C. WWP1: A versatile ubiquitin E3 ligase in signaling and diseases. Cell. Mol. Life Sci. 2011, 69, 1425–1434. [Google Scholar] [CrossRef]
- Woelk, T.; Oldrini, B.; Maspero, E.; Confalonieri, S.; Cavallaro, E.; Di Fiore, P.P.; Polo, S. Molecular mechanisms of coupled monoubiquitination. Nat. Cell Biol. 2006, 8, 1246–1254. [Google Scholar] [CrossRef]
- Angers, A.; Ramjaun, A.R.; McPherson, P.S. The HECT domain ligase itch ubiquitinates endophilin and localizes to the trans-Golgi network and endosomal system. J. Biol. Chem. 2004, 279, 11471–11479. [Google Scholar] [CrossRef] [PubMed]
- Savio, M.G.; Wollscheid, N.; Cavallaro, E.; Algisi, V.; Di Fiore, P.P.; Sigismund, S.; Maspero, E.; Polo, S. USP9X Controls EGFR Fate by Deubiquitinating the Endocytic Adaptor Eps15. Curr. Biol. 2016, 26, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Wright, M.H.; Berlin, I.; Nash, P.D. Regulation of Endocytic Sorting by ESCRT–DUB-Mediated Deubiquitination. Cell Biochem. Biophys. 2011, 60, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Vallabhapurapu, S.; Karin, M. Regulation and Function of NF-κB Transcription Factors in the Immune System. Annu. Rev. Immunol. 2009, 27, 693–733. [Google Scholar] [CrossRef]
- Wertz, I.E.; Dixit, V.M. Signaling to NF-κB: Regulation by Ubiquitination. Cold Spring Harb. Perspect. Biol. 2010, 2, a003350. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, D.E.; McDaniel, D.K.; Ringel-Scaia, V.M.; Allen, I.C. Modulating Inflammation through the Negative Regulation of NF-κB Signaling. J. Leukoc. Biol. 2018, 103, 1131–1150. [Google Scholar] [CrossRef]
- Wertz, I.E.; ORourke, K.; Zhou, H.; Eby, M.; Aravind, L.; Seshagirl, S.; Wu, P.; Wiesmann, C.; Baker, R.; Boone, D.L.; et al. De-ubiquitination and ubiquitin ligase domains of A20 downregulate NF-kB signalling. Lett. Nat. 2004, 430, 694–699. [Google Scholar] [CrossRef]
- Shembade, N.; Parvatiyar, K.; Harhaj, N.S.; Harhaj, E.W. The ubiquitin-editing enzyme A20 requires RNF11 to downregulate NF-κB signalling. EMBO J. 2009, 28, 513–522. [Google Scholar] [CrossRef]
- Shembade, N.; Harhaj, N.S.; Parvatiyar, K.; Copeland, N.G.; Jenkins, N.A.; Matesic, L.E.; Harhaj, E.W. The E3 ligase Itch negatively regulates inflammatory signaling pathways by controlling the function of the ubiquitin-editing enzyme A20. Nat. Immunol. 2008, 9, 254–262. [Google Scholar] [CrossRef]
- Jacque, E.; Ley, S.C. RNF11, a new piece in the A20 puzzle. EMBO J. 2009, 28, 455–456. [Google Scholar] [CrossRef]
- Dalal, N.V.; Pranski, E.L.; Tansey, M.G.; Lah, J.J.; Levey, A.I.; Betarbet, R.S. RNF11 modulates microglia activation through NF-κB signalling cascade. Neurosci. Lett. 2012, 528, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Pranski, E.L.; Van Sanford, C.D.; Dalal, N.V.; Orr, A.L.; Karmali, D.; Cooper, D.S.; Costa, N.; Heilman, C.J.; Gearing, M.; Lah, J.J.; et al. Comparative distribution of protein components of the A20 ubiquitin-editing complex in normal human brain. Neurosci. Lett. 2012, 520, 104–109. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pranski, E.L.; Van Sanford, C.D.; Dalal, N.V.; Orr, A.L.; Karmali, D.; Cooper, D.S.; Gearing, M.; Lah, J.J.; Levey, A.I.; Betarbet, R.S. NF-κB activity is inversely correlated to RNF11 expression in Parkinson’s disease. Neurosci. Lett. 2013, 547, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Charoenthongtrakul, S.; Gao, L.; Parvatiyar, K.; Lee, D.; Harhaj, E.W. RING Finger Protein 11 Targets TBK1/IKKi Kinases to Inhibit Antiviral Signaling. PLoS ONE 2013, 8, e53717. [Google Scholar] [CrossRef]
- Bazzoni, F.; Rossato, M.; Fabbri, M.; Gaudiosi, D.; Mirolo, M.; Mori, L.; Tamassia, N.; Mantovani, A.; Cassatella, M.A.; Locati, M. Induction and regulatory function of miR-9 in human monocytes and neutrophils exposed to proinflammatory signals. Proc. Natl. Acad. Sci. USA 2009, 106, 5282–5287. [Google Scholar] [CrossRef]
- Boldin, M.P.; Taganov, K.D.; Rao, D.S.; Yang, L.; Zhao, J.L.; Kalwani, M.; Garcia-Flores, Y.; Luong, M.; Devrekanli, A.; Xu, J.; et al. miR-146a is a significant brake on autoimmunity, myeloproliferation, and cancer in mice. J. Exp. Med. 2011, 208, 1189–1201. [Google Scholar] [CrossRef]
- Stewart, C.R.; Marsh, G.A.; Jenkins, K.A.; Gantier, M.P.; Tizard, M.L.; Middleton, D.; Lowenthal, J.W.; Haining, J.; Izzard, L.; Gough, T.J.; et al. Promotion of Hendra Virus Replication by MicroRNA 146a. J. Virol. 2013, 87, 3782–3791. [Google Scholar] [CrossRef]
- Chen, Y.; Xu, H.; Liu, J.; Deng, Y.; Cheng, H.; Zhan, T.; Lu, X.; Liao, T.; Guo, L.; Zhu, S.; et al. Gga-miR-19b-3p Inhibits Newcastle Disease Virus Replication by Suppressing Inflammatory Response via Targeting RNF11 and ZMYND11. Front. Microbiol. 2019, 10, 2006. [Google Scholar] [CrossRef]
- Gantier, M.P.; Stunden, H.J.; McCoy, C.E.; Behlke, M.A.; Wang, D.; Kaparakis-Liaskos, M.; Sarvestani, S.T.; Yang, Y.H.; Xu, D.; Corr, S.C.; et al. A miR-19 regulon that controls NF-κB signaling. Nucleic Acids Res. 2012, 40, 8048–8058. [Google Scholar] [CrossRef]
- Sluimer, J.; Distel, B. Regulating the human HECT E3 ligases. Cell. Mol. Life Sci. 2018, 75, 3121–3141. [Google Scholar] [CrossRef]
- Verdecia, M.A.; Joazeiro, C.A.P.; Wells, N.J.; Ferrer, J.-L.; Bowman, M.E.; Hunter, T.; Noel, J.P. Conformational Flexibility Underlies Ubiquitin Ligation Mediated by the WWP1 HECT Domain E3 Ligase. Mol. Cell 2003, 11, 249–259. [Google Scholar] [CrossRef]
- Maspero, E.; Mari, S.; Valentini, E.; Musacchio, A.; Fish, A.; Pasqualato, S.; Polo, S. Structure of the HECT:ubiquitin complex and its role in ubiquitin chain elongation. EMBO Rep. 2011, 12, 342–349. [Google Scholar] [CrossRef]
- Maspero, E.; Valentini, E.; Mari, S.; Cecatiello, V.; Soffientini, P.; Pasqualato, S.; Polo, S. Structure of a ubiquitin-loaded HECT ligase reveals the molecular basis for catalytic priming. Nat. Struct. Mol. Biol. 2013, 20, 696–701. [Google Scholar] [CrossRef]
- Kamadurai, H.B.; Souphron, J.; Scott, D.C.; Duda, D.M.; Miller, D.J.; Stringer, D.; Piper, R.C.; Schulman, B.A. Insights into Ubiquitin Transfer Cascades from a Structure of a UbcH5B∼Ubiquitin-HECTNEDD4L Complex. Mol. Cell 2009, 36, 1095–1102. [Google Scholar] [CrossRef]
- Shembade, N.; Ma, A.; Harhaj, E.W. Inhibition of NF- B Signaling by A20 Through Disruption of Ubiquitin Enzyme Complexes. Science 2010, 327, 1135–1139. [Google Scholar] [CrossRef]
- Shi, C.-S.; Kehrl, J.H. Tumor necrosis factor (TNF)-induced germinal center kinase-related (GCKR) and stress-activated protein kinase (SAPK) activation depends upon the E2/E3 complex Ubc13-Uev1A/TNF receptor-associated factor 2 (TRAF2). J. Biol. Chem. 2003, 278, 15429–15434. [Google Scholar] [CrossRef]
- Bertrands, M.J.M.; Milutinovic, S.; Dickson, K.M.; Ho, W.C.; Boudreault, A.; Durkin, J.; Gillard, J.W.; Jaquith, J.B.; Morris, S.J.; Barker, P.A. cIAP1 and cIAP2 Facilitate Cancer Cell Survival by Functioning as E3 Ligases that Promote RIP1 Ubiquitination. Mol. Cell 2008, 30, 689–700. [Google Scholar] [CrossRef]
- Ohtake, F.; Tsuchiya, H.; Saeki, Y.; Tanaka, K. K63 ubiquitylation triggers proteasomal degradation by seeding branched ubiquitin chains. Proc. Natl. Acad. Sci. USA 2018, 115, E1401–E1408. [Google Scholar] [CrossRef]
- Kim, H.C.; Huibregtse, J.M. Polyubiquitination by HECT E3s and the Determinants of Chain Type Specificity. Mol. Cell. Biol. 2009, 29, 3307–3318. [Google Scholar] [CrossRef]
- French, M.E.; Kretzmann, B.R.; Hicke, L. Regulation of the RSP5 Ubiquitin Ligase by an Intrinsic Ubiquitin-binding Site. J. Biol. Chem. 2009, 284, 12071–12079. [Google Scholar] [CrossRef]
- Kim, W.; Bennett, E.J.; Huttlin, E.L.; Guo, A.; Li, J.; Possemato, A.; Sowa, M.E.; Rad, R.; Rush, J.; Comb, M.J.; et al. Systematic and Quantitative Assessment of the Ubiquitin-Modified Proteome. Mol. Cell 2011, 44, 325–340. [Google Scholar] [CrossRef] [PubMed]
- Ogunjimi, A.A.; Wiesner, S.; Briant, D.J.; Varelas, X.; Sicheri, F.; Forman-Kay, J.; Wrana, J.L. The Ubiquitin Binding Region of the Smurf HECT Domain Facilitates Polyubiquitylation and Binding of Ubiquitylated Substrates. J. Biol. Chem. 2010, 285, 6308–6315. [Google Scholar] [CrossRef] [PubMed]
- Sartelet, A.; Druet, T.; Michaux, C.; Fasquelle, C.; Géron, S.; Tamma, N.; Zhang, Z.; Coppieters, W.; Georges, M.; Charlier, C. A Splice Site Variant in the Bovine RNF11 Gene Compromises Growth and Regulation of the Inflammatory Response. PLoS Genet 2012, 8, e1002581. [Google Scholar] [CrossRef]
- Pranski, E.L.; Dalal, N.V.; Herskowitz, J.H.; Orr, A.L.; Roesch, L.A.; Fritz, J.J.; Heilman, C.J.; Lah, J.J.; Levey, A.I.; Betarbet, R.S. Neuronal RING finger protein 11 (RNF11) regulates canonical NF-κB signaling. J. Neuroinflamm. 2012, 9, 67. [Google Scholar] [CrossRef]
- Bentea, E.; Verbruggen, L.; Massie, A. The Proteasome Inhibition Model of Parkinson’s Disease. J. Parkinsons Dis. 2017, 7, 31–63. [Google Scholar] [CrossRef]
- Noureddine, M.A.; Li, Y.-J.; van der Walt, J.M.; Walters, R.; Jewett, R.M.; Xu, H.; Wang, T.; Walter, J.W.; Scott, B.L.; Hulette, C.; et al. Genomic convergence to identify candidate genes for Parkinson disease: SAGE analysis of the substantia nigra. Mov. Disord. 2005, 20, 1299–1309. [Google Scholar] [CrossRef]
- Hicks, A.A.; Pétursson, H.; Jónsson, T.; Stefánsson, H.; Jóhannsdóttir, H.S.; Sainz, J.; Frigge, M.L.; Kong, A.; Gulcher, J.R.; Stefánsson, K.; et al. A susceptibility gene for late-onset idiopathic Parkinson’s disease. Ann. Neurol. 2002, 52, 549–555. [Google Scholar] [CrossRef]
- Anderson, L.R.; Betarbet, R.S.; Gearing, M.; Gulcher, J.; Hicks, A.; Stefánsson, K.; Lah, J.J.; Levey, A.I. PARK10 Candidate RNF11 Is Expressed by Vulnerable Neurons and Localizes to Lewy Bodies in Parkinson Disease Brain. J. Neuropathol. Exp. Neurol. 2007, 66, 955–964. [Google Scholar] [CrossRef][Green Version]
- Müller, A.; Niederstadt, L.; Jonas, W.; Yi, C.-X.; Meyer, F.; Wiedmer, P.; Fischer, J.; Grötzinger, C.; Schürmann, A.; Tschöp, M.; et al. Ring Finger Protein 11 Inhibits Melanocortin 3 and 4 Receptor Signaling. Front. Endocrinol. 2016, 7, 109. [Google Scholar] [CrossRef]
- Seki, N.; Hattori, A.; Hayashi, A.; Kozuma, S.; Sasaki, M.; Suzuki, Y.; Sugano, S.; Muramatsu, M.-A.; Saito, T. Cloning and expression profile of mouse and human genes, Rnf11/RNF11, encoding a novel RING-H2 ¢nger protein. BBA Mol. Basis Dis. 1999, 1489, 421–427. [Google Scholar]
- Wang, J.-L.; Hu, Y.; Kong, X.; Wang, Z.-H.; Chen, H.-Y.; Xu, J.; Fang, J.-Y. Candidate microRNA Biomarkers in Human Gastric Cancer: A Systematic Review and Validation Study. PLoS ONE 2013, 8, e73683. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yang, H.-J.; Gao, S.-S.; Wang, M.; Shi, Y.; Cheng, B.-F.; Feng, Z.-W. Identification of a novel role of RING finger protein 11 promoting the metastasis of murine melanoma cells. Am. J. Transl. Res. 2015, 7, 1629–1635. [Google Scholar]
- Daviet, L.; Colland, F. Targeting ubiquitin specific proteases for drug discovery. Biochimie 2008, 90, 270–283. [Google Scholar] [CrossRef]
- Pal, A.; Young, M.A.; Donato, N.J. Emerging Potential of Therapeutic Targeting of Ubiquitin-Specific Proteases in the Treatment of Cancer. Cancer Res. 2014, 74, 4955–4966. [Google Scholar] [CrossRef]
- Guédat, P.; Colland, F. Patented small molecule inhibitors in the ubiquitin proteasome system. BMC Biochem. 2007, 8, S14. [Google Scholar] [CrossRef]
- Sa, J.K.; Hong, J.Y.; Lee, I.-K.; Kim, J.-S.; Sim, M.-H.; Kim, H.J.; An, J.Y.; Sohn, T.S.; Lee, J.H.; Bae, J.M.; et al. Comprehensive pharmacogenomic characterization of gastric cancer. Genome Med. 2020, 12, 1–12. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PTM | Residue | In Vivo Identification | Reference (PMID) | Predicted | |
|---|---|---|---|---|---|
| NetPhos3.1 | UbPred | ||||
| Myristoylation | Gly2 | ✓ | 20676133 | ||
| Palmitoylation | Cys4 | ✓ | 20676133 | ||
| Nitrosylation | Cys4 | ✓ | 24105792 | ||
| Ubiquitination | Lys6 | ✓ | |||
| Lys82 | ✓ | 22790023, 32274752 | |||
| Sumoylation | ND | ✓ | 32274752 | ||
| Phosphorylation | Ser7 | ✓ | |||
| Ser10 | ✓ | 22617229 | ✓ | ||
| Ser14 | ✓ | 27281782, 27251275, | ✓ | ||
| 25850435, 24275569, | |||||
| 23749302, 23312004, | |||||
| 23186163 | |||||
| Ser19 | ✓ | ||||
| Ser21 | ✓ | 20415495 | ✓ | ||
| Ser25 | ✓ | 25850435, 23684622, 19144319 | ✓ | ||
| Thr30 | ✓ | 24275569 | ✓ | ||
| Thr52 | ✓ | ||||
| Ser54 | ✓ | 24275569 | ✓ | ||
| Thr63 | ✓ | 22645316 | ✓ | ||
| Ser92 | ✓ | ||||
| Thr135 | ✓ | 16123141 | |||
| Ser149 | ✓ | ||||
| Yeast-Two-Hybrid | Mass Spectr. | Yeast-Two-Hybrid | Mass Spectr. | |||
|---|---|---|---|---|---|---|
| Gene Name | Binding Domain | Gene Name | Gene Name | Gene Name | Binding Domain | Gene Name |
| (A) E2, E3 and DUBs | (C) Effectors of anti-inflammatory response | |||||
| UBE2N | UBC | UBCH5c | TNFAIP3 | IKBKB | ||
| UBCH5a | UBC | UBCH6 | TAX1BP1 | CC-ZnF_C2H2 | CHUK | |
| UBCH5b | UBC | OPTN | ZnF_C2H2 | S100A8 | ||
| UBCH5c | UBC | NEMO | UBAN-ZnF_C2H2 | S100A9 | ||
| HERC1 | UBA-like | RNF216 | IBR | S100A11 | ||
| HERC2 | UBA-like | UBCH7 | (D) Membrane proteins and receptors | |||
| HUWE1 | UBCH9 | |||||
| CBL-b | UBA | UBC7 | CD45 | EGFR | ||
| WWP1 | WW | UBE2V1 | TGFBR1 | |||
| WWP2 | WW | hRul138 | TFRC | |||
| NEDD4 | WW | LISTERIN | CD44 | |||
| NEDD4-2 | WW | ITCH | ATP2B1 | |||
| SMURF1 | WW-HECT | IFITM3 | ||||
| SMURF2 | WW-HECT | RTN4 | ||||
| USP5 | UBA | (E) Other functions | ||||
| TNFAIP3 | ZnF_A20 | |||||
| RNF168 | UIM-MIU | NY-REN-25 | UBD | MYOF | ||
| USP5 | ERCC6 | UBD | MYOF | |||
| AMSH | QARS | DAP3 | ||||
| CUL1 | POLI | PDXP | ||||
| (B) Effectors and adaptors of intracellular traffic | KHNYN | NYN-CUBAN | SLC27A2 | |||
| CALCOCO2 | ZnF_C2H2 | PTP4A1 | ||||
| RPS27A | RAB6A | AUP1 | CUE | PHB | ||
| UBA52 | EXOSC10 | SPRTN | UBZ | PHB2 | ||
| EPN1 | UIM | RP42 | UBA | APOA2 | ||
| EPN2 | UIM | FLJ21588 | CUE | SHROOM3 | ||
| EPN3 | UIM | SDCBP | PDZ | CHCHD3 | ||
| ANKRD13A | UIM | SMAD4 | QPCTL | |||
| GGA1 | GAT | TXN | ||||
| GGA3 | GAT | LRRC59 | ||||
| TOM1L2 | GAT | SOAT1 | ||||
| ALIX | MOGS | |||||
| EPS15 | BRI3BP | |||||
| STAM2 | VHS-UIM-SH3-GAT | DRG1 | ||||
| Endofin | VCP | |||||
| ALIX | ALIX-LYPXL-bnd | DMD | ||||
| SARA | MSN | |||||
| UBQL-2 | UBA | |||||
| UBQL-4 | UBA | |||||
| MYO6 | MIU-MyUb | |||||
| NAF1 | ||||||
| RABEX5 | ||||||
| NDP52 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mattioni, A.; Castagnoli, L.; Santonico, E. RNF11 at the Crossroads of Protein Ubiquitination. Biomolecules 2020, 10, 1538. https://doi.org/10.3390/biom10111538
Mattioni A, Castagnoli L, Santonico E. RNF11 at the Crossroads of Protein Ubiquitination. Biomolecules. 2020; 10(11):1538. https://doi.org/10.3390/biom10111538
Chicago/Turabian StyleMattioni, Anna, Luisa Castagnoli, and Elena Santonico. 2020. "RNF11 at the Crossroads of Protein Ubiquitination" Biomolecules 10, no. 11: 1538. https://doi.org/10.3390/biom10111538
APA StyleMattioni, A., Castagnoli, L., & Santonico, E. (2020). RNF11 at the Crossroads of Protein Ubiquitination. Biomolecules, 10(11), 1538. https://doi.org/10.3390/biom10111538