Production of Polyhydroxyalkanoates by Two Halophilic Archaeal Isolates from Chott El Jerid Using Inexpensive Carbon Sources


Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Maintenance of Isolates
2.2. Screening of Potential Halophilic PHA Producers by Staining Procedures
2.3. Screening of PHA Synthase Genes by Degenerate Polymerase Chain Reaction
2.4. Staining of Promising Cells with Nile Red
2.5. Optimization of Growth Conditions of the Potential Halophilic PHA Producers
2.6. Effect of Carbon Sources on PHA Production by Potential Halophilic PHA Producers
2.7. Measurement of Cell Dry Weight (CDW)
2.8. Analysis of PHA in Dried Cells by Gas Chromatography (GC)
2.9. Polymer Extraction and Quantitative Analysis of PHA
3. Results
3.1. Screening of PHA-Producing Isolates from Chott El Jerid
3.2. Growth Kinetics of Potential Halophilic PHA Producers
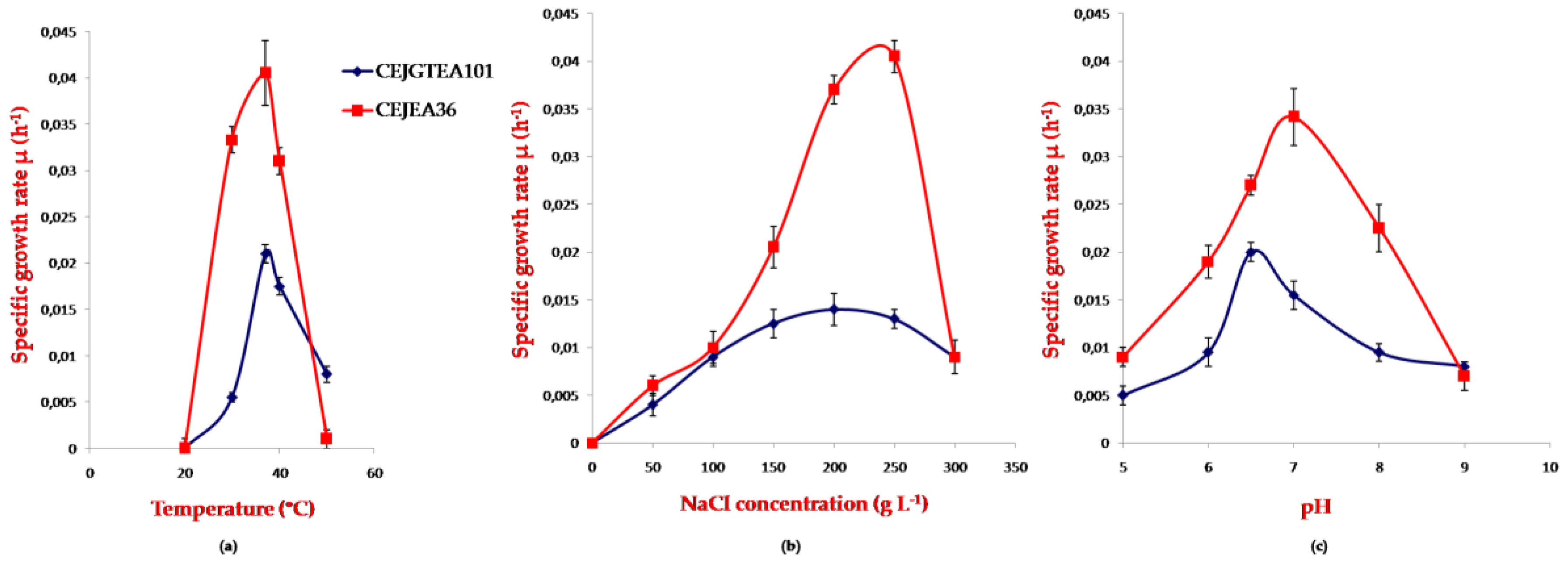
3.3. Effect of Temperature, pH, Salinity on Growth of Potential Halophilic PHA Producers
3.4. Effect of Carbon Sources on Growth and PHA Production by Potential Halophilic PHA Producers
3.5. Yield of PHA Extracted from Potential Halophilic PHA Producers
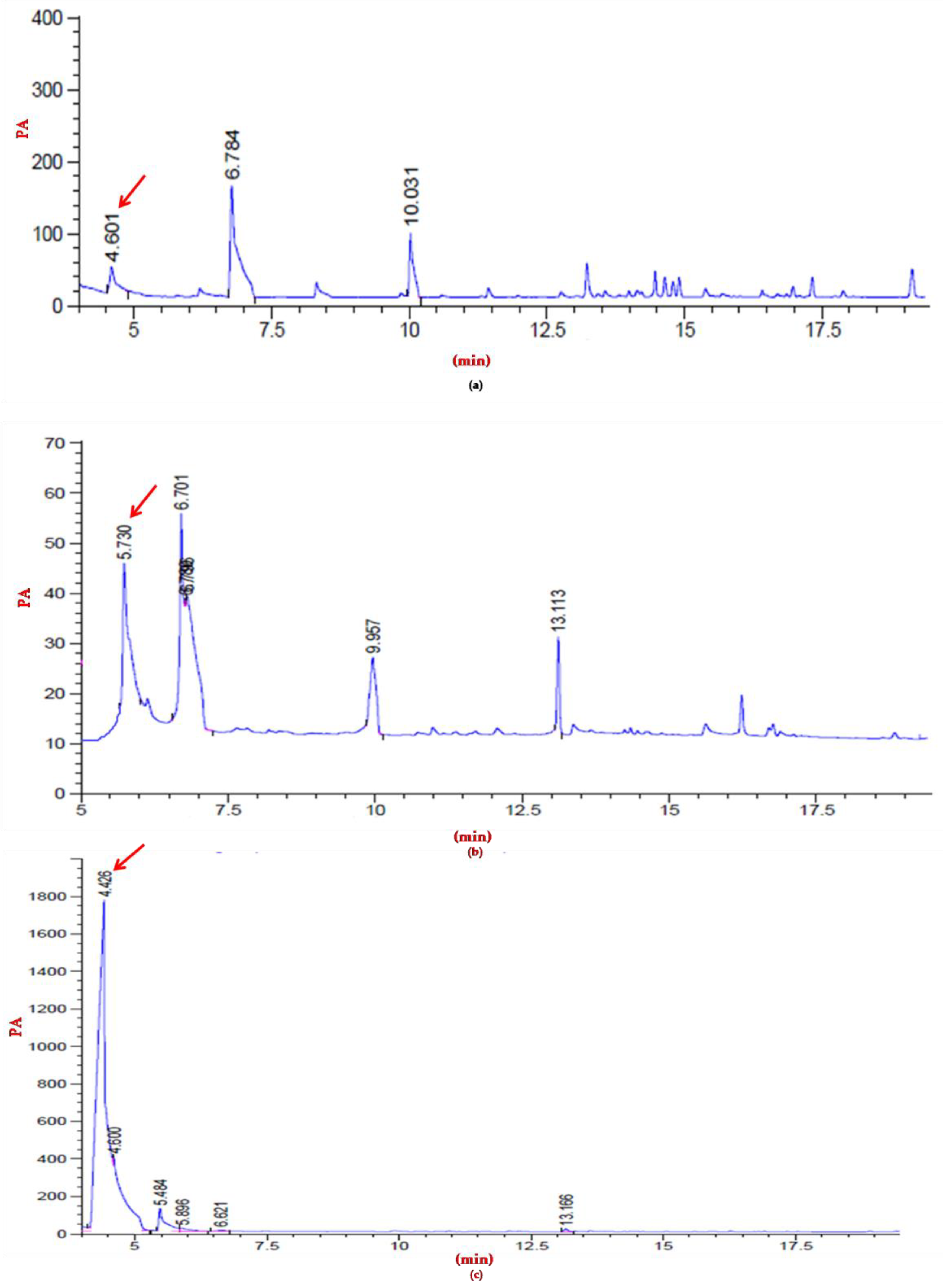
3.6. Polymer Characterization
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Anderson, A.J.; Dawes, E.A. Occurrence, metabolism, metabolic role, and industrial uses of bacterial polyhydroxyalkanoates. Microbiol. Mol. Biol. Rev. 1990, 54, 450–472. [Google Scholar] [CrossRef]
- Jendrossek, D.; Pfeiffer, D. New insights in the formation of polyhydroxyalkanoate granules (carbonosomes) and novel functions of poly(3-hydroxybutyrate). Environ. Microbiol. 2014, 16, 2357–2373. [Google Scholar] [CrossRef] [PubMed]
- Steinbüchel, A.; Füchtenbusch, B. Bacterial and other biological systems for polyester production. Trends Biotechnol. 1998, 16, 419–427. [Google Scholar] [CrossRef]
- Babruwad, P.R.; Prabhu, S.U.; Upadhyaya, K.P.; Hungund, B.S. Production and characterization of thermostable polyhydroxybutyrate from Bacillus cereus PW3A. J. Biochem. Technol. 2015, 6, 990–995. [Google Scholar]
- Wei, Y.H.; Chen, W.C.; Huang, C.K.; Wu, H.S.; Sun, Y.M.; Lo, C.W.; Janarthanan, O.M. Screening and evaluation of polyhydroxybutyrate-producing strains from indigenous isolate Cupriavidus taiwanensis strains. Int. J. Mol. Sci. 2011, 12, 252–265. [Google Scholar] [CrossRef]
- Rathi, D.N.; Amir, H.G.; Abed, R.M.M.; Kosugi, A.; Arai, T.; Sulaiman, O.; Hashim, R.; Sudesh, K. Polyhydroxyalkanoate biosynthesis and simplified polymer recovery by a novel moderately halophilic bacterium isolated from hypersaline microbial mats. J. Appl. Microbiol. 2012, 114, 384–395. [Google Scholar] [CrossRef]
- Poli, A.; Di Donato, P.; Abbamondi, G.R.; Nicolaus, B. Synthesis, production, and biotechnological applications of exopolysaccharides and polyhydroxyalkanoates by Archaea. Archaea 2011, 2011, 1–13. [Google Scholar] [CrossRef]
- Taran, M. Utilization of petrochemical wastewater for the production of poly(3-hydroxybutyrate) by Haloarcula sp. IRU1. J. Hazard. Mater. 2011, 188, 26–28. [Google Scholar] [CrossRef]
- Salgaonkar, B.B.; Mani, K.; Bragança, J.M. Accumulation of polyhydroxyalkanoates by halophilic archaea isolated from traditional solar salterns of India. Extremophiles 2013, 17, 787–795. [Google Scholar] [CrossRef]
- Oren, A. Industrial and environmental applications of halophilic microorganisms. Environ. Technol. 2010, 31, 825–834. [Google Scholar] [CrossRef]
- Karray, F.; Ben Abdallah, M.; Kallel, N.; Hamza, M.; Fakhfakh, M.; Sayadi, S. Extracellular hydrolytic enzymes produced by halophilic bacteria and archaea isolated from hypersaline lake. Mol. Biol. Rep. 2018, 45, 1297–1309. [Google Scholar] [CrossRef] [PubMed]
- Nicolaus, B.; Lama, L.; Esposito, E.; Manca, M.C.; Improta, R.; Bellitti, M.R.; Duckworth, A.W.; Grant, W.D.; Gambacorta, A. Haloarcula spp able to biosynthesize exo- and endopolymers. J. Ind. Microbiol. Biotechnol. 1999, 23, 489–496. [Google Scholar] [CrossRef]
- Ben Abdallah, M.; Karray, F.; Mhiri, N.; Mei, N.; Quéméneur, M.; Cayol, J.L.; Erauso, G.; Tholozan, J.L.; Alazard, D.; Sayadi, S. Prokaryotic diversity in a Tunisian hypersaline lake, Chott El Jerid. Extremophiles 2016, 20, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Spiekermann, P.; Rehm, B.H.A.; Kalscheuer, R.; Baumeister, D.; Steinbüchel, A. A sensitive, viable-colony staining method using Nile red for direct screening of bacteria that accumulate polyhydroxyalkanoic acids and other lipid storage compounds. Arch. Microbiol. 1999, 171, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Gonzalez, J.E.; Willis, L.B.; Walker, G.C. A Novel Screening Method for Isolating Exopolysaccharide-Deficient Mutants. Appl. Environ. Microbiol. 1998, 64, 4600–4602. [Google Scholar] [CrossRef] [PubMed]
- Solaiman, D.K.Y.; Ashby, R.D.; Foglia, T.A. Rapid and specific identification of medium-chain-length polyhydroxyalkanoate synthase gene by polymerase chain reaction. Appl. Microbiol. Biotechnol. 2000, 53, 690–694. [Google Scholar] [CrossRef]
- Han, J.; Hou, J.; Liu, H.; Cai, S.; Feng, B.; Zhou, J.; Xiang, H. Wide distribution among halophilic archaea of a novel polyhydroxyalkanoate synthase subtype with homology to bacterial type III synthases. Appl. Environ. Microbiol. 2010, 76, 7811–7819. [Google Scholar] [CrossRef]
- Salgaonkar, B.B.; Mani, K.; Braganca, J.M. Characterization of polyhydroxyalkanoates accumulated by a moderately halophilic salt pan isolate Bacillus megaterium strain H16. J. Appl. Microbiol. 2013, 114, 1347–1356. [Google Scholar] [CrossRef]
- APHA; AWWA; WEF. Standard methods for the examination of water and wastewater, 23nd ed.; American Public Health Association; American Water Works Association; Water Environment Federation: Washington, DC, USA, 2005; pp. 1–1504. [Google Scholar]
- Brandl, H.; Gross, R.A.; Lenz, R.W.; Fuller, R.C. Pseudomonas oleovorans as a source of Poly(beta-Hydroxyalkanoates) for potential applications as biodegradable polyesters. Appl. Environ. Microbiol. 1988, 54, 1977–1982. [Google Scholar] [CrossRef]
- Bhattacharyya, A.; Pramanik, A.; Maji, S.; Haldar, S.; Mukhopadhyay, U.; Mukherjee, J. Utilization of vinasse for production of poly-3-(hydroxybutyrate-co-hydroxyvalerate) by Haloferax mediterranei. AMB Express 2012, 2, 34. [Google Scholar] [CrossRef]
- Law, J.H.; Slepecky, R.A. Assay of poly-{beta}-hydroxybutyric acid. J. Bacteriol. 1961, 82, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Danis, O.; Ogan, A.; Tatlican, P.; Attar, A.; Cakmakci, E.; Mertoglu, B.; Birbir, M. Preparation of poly(3-hydroxybutyrate-co-hydroxyvalerate) films from halophilic archaea and their potential use in drug delivery. Extremophiles 2015, 19, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Di Donato, P.; Fiorentino, G.; Anzelmo, G.; Tommonaro, G.; Nicolaus, B.; Poli, A. Re-use of vegetable wastes as cheap substrates for extremophile biomass production. Waste and Biomass Valorization 2011, 2, 103–111. [Google Scholar] [CrossRef]
- Kumar, V.; Tiwari, S.K. Halocin diversity among halophilic archaea and their applications. In Microbial Diversity in Ecosystem Sustainability and Biotechnological Applications; Springer Singapore: Singapore, 2019; pp. 497–532. [Google Scholar]
- Amaro, T.M.M.M.; Rosa, D.; Comi, G.; Iacumin, L. Prospects for the use of whey for polyhydroxyalkanoate (PHA) production. Front. Microbiol. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Koller, M. Polyhydroxyalkanoate biosynthesis at the edge of water activitiy-haloarchaea as biopolyester factories. Bioengineering 2019, 6, 34. [Google Scholar] [CrossRef]
- Pramanik, A.; Mitra, A.; Arumugam, M.; Bhattacharyya, A.; Sadhukhan, S.; Ray, A.; Haldar, S.; Mukhopadhyay, U.K.; Mukherjee, J. Utilization of vinasse for the production of polyhydroxybutyrate by Haloarcula marismortui. AMB Express 2012, 57, 71–79. [Google Scholar] [CrossRef]
- Mahansaria, R.; Dhara, A.; Saha, A.; Haldar, S.; Mukherjee, J. Production enhancement and characterization of the polyhydroxyalkanoate produced by Natrinema ajinwuensis (as synonym) ≡ Natrinema altunense strain RM-G10. Int. J. Biol. Macromol. 2018, 107, 1480–1490. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Sugar Wastewater |
|---|---|
| TOC (g L−1) | 5.73 ± 0.8 |
| TN (g L−1) | 0.054 ± 0.02 |
| TS (g L−1) | 19 ± 1.2 |
| VS (g L−1) | 17.45 ± 0.8 |
| pH | 7.4 |
| Salinity (g L−1) | 22.06 ± 0.6 |
| Isolates | Cell Dry Weight (g L−1) | Dry weight Extracted PHA a (g L−1) | PHA Accumulation (wt %) | Nature of PHA | Carbon Source | References |
|---|---|---|---|---|---|---|
| CEJGTEA101 | 0.5 | 0.175 | 35 | PHB | Glucose | This study |
| Natrinema pallidum strain 1KYS1 | 0.174 | 0.075 | 53.14 | PHBV | Corn starch | [23] |
| 2.219 | 0.055 | 2.48 | PHBV | Sucrose | ||
| 0.457 | 0.091 | 19.92 | PHBV | Whey | ||
| 0.371 | 0.039 | 10.5 | PHBV | Melon | ||
| 2.55 | 0.077 | 3.02 | PHBV | Apple | ||
| 3.858 | 0.464 | 12.03 | PHBV | Tomato | ||
| CEJEA36 | 0.5 | 0.125 | 25 | PHV | Glucose | This study |
| Haloterrigena hispanica strain FP1 | 0.6 | n.d. | 0.13 | PHB | Carrot waste | [24] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ben Abdallah, M.; Karray, F.; Sayadi, S. Production of Polyhydroxyalkanoates by Two Halophilic Archaeal Isolates from Chott El Jerid Using Inexpensive Carbon Sources. Biomolecules 2020, 10, 109. https://doi.org/10.3390/biom10010109
Ben Abdallah M, Karray F, Sayadi S. Production of Polyhydroxyalkanoates by Two Halophilic Archaeal Isolates from Chott El Jerid Using Inexpensive Carbon Sources. Biomolecules. 2020; 10(1):109. https://doi.org/10.3390/biom10010109
Chicago/Turabian StyleBen Abdallah, Manel, Fatma Karray, and Sami Sayadi. 2020. "Production of Polyhydroxyalkanoates by Two Halophilic Archaeal Isolates from Chott El Jerid Using Inexpensive Carbon Sources" Biomolecules 10, no. 1: 109. https://doi.org/10.3390/biom10010109
APA StyleBen Abdallah, M., Karray, F., & Sayadi, S. (2020). Production of Polyhydroxyalkanoates by Two Halophilic Archaeal Isolates from Chott El Jerid Using Inexpensive Carbon Sources. Biomolecules, 10(1), 109. https://doi.org/10.3390/biom10010109