Modulation of Polar Lipid Profiles in Chlorella sp. in Response to Nutrient Limitation

 , , , ,
, , , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Microalgal Strain and Culture Conditions
2.2. Bacterial Cell Enumeration (Flow Cytometry)
2.3. Particulate Organic Carbon (POC) and Nitrogen Analyses
2.4. Nutrient Analyses
2.5. Lipid Extraction
2.6. Polar Lipid Analyses (Liquid Chromatography–Electrospray Ionisation Mass Spectrometry (LC-ESI-MS/MS))
2.7. Fatty Acid Analyses (Gas Chromatography-Mass Spectrometry: GC-MS)
2.8. Data Analyses
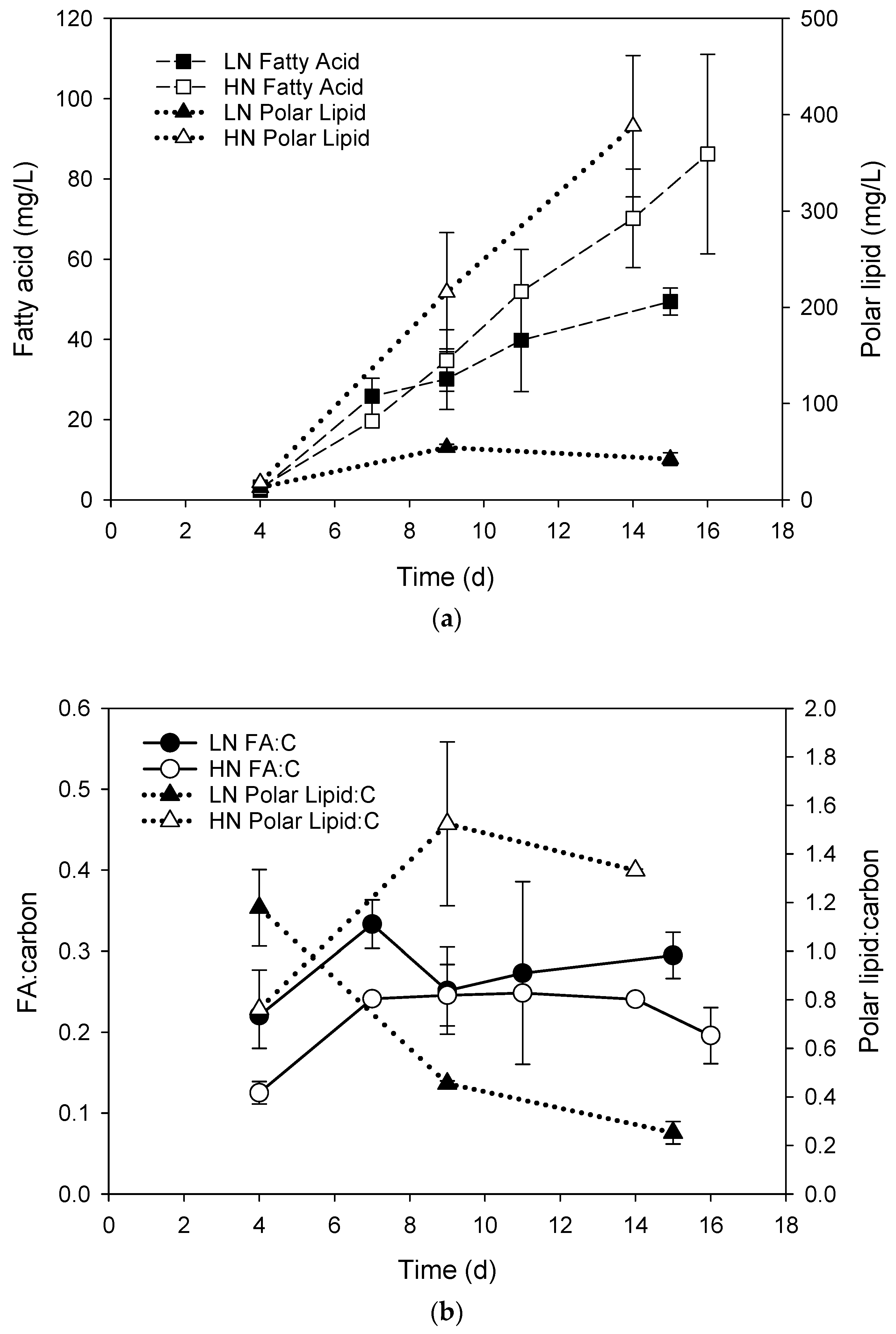
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Guschina, I.; Harwood, J. Algal lipids and effect of the environment on their biochemistry. In Lipids in Aquatic Ecosystems; Kainz, M., Brett, M.T., Arts, M.T., Eds.; Springer: New York, NY, USA, 2009; pp. 1–24. [Google Scholar] [CrossRef]
- Murata, N.; Siegenthaler, P.-A. Lipids in Photosynthesis: An Overview. In Lipids in Photosynthesis: Structure, Function and Genetics; Paul-André, S., Norio, M., Eds.; Springer: Berlin/Heidelberg, Germany, 1998; Volume 6, pp. 1–20. [Google Scholar]
- Van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Hölzl, G.; Dörmann, P. Structure and function of glycoglycerolipids in plants and bacteria. Prog. Lipid Res. 2007, 46, 225–243. [Google Scholar] [CrossRef] [PubMed]
- Benning, C. Biosynthesis and function of the sulfolipid sulfoquinovosyl diacylglycerol. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 1998, 49, 53–75. [Google Scholar] [CrossRef] [PubMed]
- Wada, H.; Mizusawa, N. The role of phosphatidylglycerol in photosynthesis. In Lipids in Photosynthesis: Essential and Regulatory Functions; Wada, H., Murata, N., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 243–263. [Google Scholar]
- Boudière, L.; Michaud, M.; Petroutsos, D.; Rébeillé, F.; Falconet, D.; Bastien, O.; Roy, S.; Finazzi, G.; Rolland, N.; Jouhet, J.; et al. Glycerolipids in photosynthesis: Composition, synthesis and trafficking. Biochim. Biophys. Acta (BBA) 2014, 1837, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Latowski, D.; Åkerlund, H.-E.; Strzałka, K. Violaxanthin De-Epoxidase, the Xanthophyll Cycle Enzyme, Requires Lipid Inverted Hexagonal Structures for Its Activity. Biochemistry 2004, 43, 4417–4420. [Google Scholar] [CrossRef] [PubMed]
- Harwood, J.L. Membrane lipids in algae. In Lipids in Photosynthesis: Structure, Function and Genetics; Siegenthaler, P.-A., Murata, N., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1998; pp. 53–64. [Google Scholar]
- Alonso, D.L.; Belarbi, E.H.; Fernandez-Sevilla, J.M.; Rodriguez-Ruiz, J.; Grima, E.M. Acyl lipid composition variation related to culture age and nitrogen concentration in continuous culture of the microalga Phaeodactylum Tricornutum. Phytochemistry 2000, 54, 461–471. [Google Scholar] [CrossRef]
- Fidalgo, J.P.; Cid, A.; Torres, E.; Sukenik, A.; Herrero, C. Effects of nitrogen source and growth phase on proximate biochemical composition, lipid classes and fatty acid profile of the marine microalga Isochrysis galbana. Aquaculture 1998, 166, 105–116. [Google Scholar] [CrossRef]
- Guiheneuf, F.; Fouqueray, M.; Mimouni, V.; Ulmann, L.; Jacquette, B.; Tremblin, G. Effect of UV stress on the fatty acid and lipid class composition in two marine microalgae Pavlova lutheri (Pavlovophyceae) and Odontella aurita (Bacillariophyceae). J. Appl. Phycol. 2010, 22, 629–638. [Google Scholar] [CrossRef]
- Guiheneuf, F.; Mimouni, V.; Ulmann, L.; Tremblin, G. Environmental factors affecting growth and omega 3 fatty acid composition in Skeletonema costatum. The influences of irradiance and carbon source. Diatom Res. 2008, 23, 93–103. [Google Scholar] [CrossRef]
- Guiheneuf, F.; Mimouni, V.; Ulmann, L.; Tremblin, G. Combined effects of irradiance level and carbon source on fatty acid and lipid class composition in the microalga Pavlova lutheri commonly used in mariculture. J. Exp. Mar. Biol. Ecol. 2009, 369, 136–143. [Google Scholar] [CrossRef]
- Liang, Y.; Beardall, J.; Heraud, P. Effects of nitrogen source and UV radiation on the growth, chlorophyll fluorescence and fatty acid composition of Phaeodactylum tricornutum and Chaetoceros muelleri (Bacillarlophyceae). J. Photochem. Photobiol. B Biol. 2006, 82, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Reitan, K.I.; Rainuzzo, J.R.; Olsen, Y. Effect of nutrient limitation on fatty-acid and lipid-content of marine microalgae. J. Phycol. 1994, 30, 972–979. [Google Scholar] [CrossRef]
- Plouguerné, E.; da Gama, B.A.P.; Pereira, R.C.; Barreto-Bergter, E. Glycolipids from seaweeds and their potential biotechnological applications. Front. Cell. Infect. Microbiol. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Hossain, Z.; Kurihara, H.; Hosokawa, M.; Takahashi, K. Growth inhibition and induction of differentiation and apoptosis mediated by sodium butyrate in caco-2 cells with algal glycolipids. In Vitro Cell. Dev. Biol. Anim. 2005, 41, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Imbs, T.I.; Ermakova, S.P.; Fedoreyev, S.A.; Anastyuk, S.D.; Zvyagintseva, T.N. Isolation of Fucoxanthin and Highly Unsaturated Monogalactosyldiacylglycerol from Brown Alga Fucus evanescens C Agardh and In Vitro Investigation of Their Antitumor Activity. Mar. Biotechnol. 2013, 15, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-J.; Sun Pan, B. Identification of Sulfoglycolipid Bioactivities and Characteristic Fatty Acids of Marine Macroalgae. J. Agric. Food Chem. 2012, 60, 8404–8410. [Google Scholar] [CrossRef] [PubMed]
- Banskota, A.H.; Stefanova, R.; Sperker, S.; Lall, S.P.; Craigie, J.S.; Hafting, J.T.; Critchley, A.T. Polar lipids from the marine macroalga Palmaria palmata inhibit lipopolysaccharide-induced nitric oxide production in RAW264.7 macrophage cells. Phytochemistry 2014, 101, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Jeong, S.Y.; Jung, H.A.; Choi, J.S.; Min, B.S.; Woo, M.H. Capsofulvesins A-C, cholinesterase inhibitors from Capsosiphon fulvescens. Chem. Pharm. Bull. 2012, 60, 1351–1358. [Google Scholar] [CrossRef] [PubMed]
- Cantillo-Ciau, Z.; Moo-Puc, R.; Quijano, L.; Freile-Pelegrín, Y. The Tropical Brown Alga Lobophora variegata: A Source of Antiprotozoal Compounds. Mar. Drugs 2010, 8, 1292–1304. [Google Scholar] [CrossRef] [PubMed]
- Treyvaud Amiguet, V.; Jewell, L.E.; Mao, H.; Sharma, M.; Hudson, J.B.; Durst, T.; Allard, M.; Rochefort, G.; Arnason, J.T. Antibacterial properties of a glycolipid-rich extract and active principle from Nunavik collections of the macroalgae Fucus evanescens C. Agardh (Fucaceae). Can. J. Microbiol. 2011, 57, 745–749. [Google Scholar] [CrossRef] [PubMed]
- Plouguerné, E.; Ioannou, E.; Georgantea, P.; Vagias, C.; Roussis, V.; Hellio, C.; Kraffe, E.; Stiger-Pouvreau, V. Anti-microfouling Activity of Lipidic Metabolites from the Invasive Brown Alga Sargassum muticum (Yendo) Fensholt. Mar. Biotechnol. 2010, 12, 52–61. [Google Scholar] [CrossRef] [PubMed]
- El Baz, F.K.; El Baroty, G.S.; Abd El Baky, H.H.; Abd El-Salam, O.I.; Ibrahim, E.A. Structural characterization and Biological Activity of Sulfolipids from selected Marine Algae. Grasas Aceites 2013, 64. [Google Scholar] [CrossRef]
- Al-Fadhli, A.; Wahidulla, S.; D’Souza, L. Glycolipids from the red alga Chondria armata (Kütz.) Okamura. Glycobiology 2006, 16, 902–915. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, C.; Yu, G.; Guan, H. Total Synthesis and Structure-Activity Relationship of Glycoglycerolipids from Marine Organisms. Mar. Drugs 2014, 12, 3634–3659. [Google Scholar] [CrossRef] [PubMed]
- Gunstone, F.D. Phospholipid Technologies and Applications; Woodhead Publishing: Sawston/Cambridge, UK, 2008. [Google Scholar]
- Safi, C.; Zebib, B.; Merah, O.; Pontalier, P.-Y.; Vaca-Garcia, C. Morphology, composition, production, processing and applications of Chlorella vulgaris: A review. Renew. Sustain. Energy Rev. 2014, 35, 265–278. [Google Scholar] [CrossRef]
- Guccione, A.; Biondi, N.; Sampietro, G.; Rodolfi, L.; Bassi, N.; Tredici, M.R. Chlorella for protein and biofuels: From strain selection to outdoor cultivation in a Green Wall Panel photobioreactor. Biotechnol. Biofuels 2014, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, Y.-F.; Chen, P.; Min, M.; Zhou, W.; Martinez, B.; Zhu, J.; Ruan, R. Characterization of a microalga Chlorella sp. well adapted to highly concentrated municipal wastewater for nutrient removal and biodiesel production. Bioresour. Technol. 2011, 102, 5138–5144. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Miao, X.; Wu, Q. High quality biodiesel production from a microalga Chlorella protothecoides by heterotrophic growth in fermenters. J. Biotechnol. 2006, 126, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Converti, A.; Casazza, A.A.; Ortiz, E.Y.; Perego, P.; Del Borghi, M. Effect of temperature and nitrogen concentration on the growth and lipid content of Nannochloropsis oculata and Chlorella vulgaris for biodiesel production. Chem. Eng. Process. 2009, 48, 1146–1151. [Google Scholar] [CrossRef]
- Liu, J.; Huang, J.; Sun, Z.; Zhong, Y.; Jiang, Y.; Chen, F. Differential lipid and fatty acid profiles of photoautotrophic and heterotrophic Chlorella zofingiensis: Assessment of algal oils for biodiesel production. Bioresour. Technol. 2011, 102, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Maeda, Y.; Yoshino, T.; Matsumoto, M.; Tanaka, T. Profiling of Polar Lipids in Marine Oleaginous Diatom Fistulifera solaris JPCC DA0580: Prediction of the Potential Mechanism for Eicosapentaenoic Acid-Incorporation into Triacylglycerol. Mar. Drugs 2014, 12, 3218–3230. [Google Scholar] [CrossRef] [PubMed]
- Cutignano, A.; Luongo, E.; Nuzzo, G.; Pagano, D.; Manzo, E.; Sardo, A.; Fontana, A. Profiling of complex lipids in marine microalgae by UHPLC/tandem mass spectrometry. Algal Res. 2016, 17, 348–358. [Google Scholar] [CrossRef]
- He, H.; Rodgers, R.P.; Marshall, A.G.; Hsu, C.S. Algae polar lipids characterized by online liquid chromatography coupled with hybrid linear quadrupole ion trap/fourier transform ion cyclotron resonance mass spectrometry. Energy Fuels 2011, 25, 4770–4775. [Google Scholar] [CrossRef]
- Lu, N.; Wei, D.; Chen, F.; Yang, S.T. Lipidomic profiling and discovery of lipid biomarkers in snow alga Chlamydomonas nivalis under salt stress. Eur. J. Lipid Sci. Technol. 2012, 114, 253–265. [Google Scholar] [CrossRef]
- Xu, J.L.; Chen, D.Y.; Yan, X.J.; Chen, J.J.; Zhou, C.X. Global characterization of the photosynthetic glycerolipids from a marine diatom Stephanodiscus sp by ultra performance liquid chromatography coupled with electrospray ionization-quadrupole-time of flight mass spectrometry. Anal. Chim. Acta 2010, 663, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Xu, J.; Jiang, Y.; Zhou, C.; Yu, X.; Zhong, Y.; Chen, J.; Yan, X. Lipidomic analysis can distinguish between two morphologically similar strains of Nannochloropsis oceanica. J. Phycol. 2015, 51, 264–276. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Chen, D.; Xu, J.; Zhou, C. Profiles of photosynthetic glycerolipids in three strains of Skeletonema determined by UPLC-Q-TOF-MS. J. Appl. Phycol. 2011, 23, 271–282. [Google Scholar] [CrossRef]
- Vieler, A.; Wilhelm, C.; Goss, R.; Süß, R.; Schiller, J. The lipid composition of the unicellular green alga Chlamydomonas reinhardtii and the diatom Cyclotella meneghiniana investigated by MALDI-TOF MS and TLC. Chem. Phys. Lipids 2007, 150, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Brewer, P.G.; Riley, J.P. The automatic determination of nitrate in sea water. Deep-Sea Res. 1965, 12, 765–772. [Google Scholar] [CrossRef]
- Zhang, J.Z.; Chi, J. Automated analysis of nanomolar concentrations of phosphate in natural waters, with liquid waveguide. Environ. Sci. Technol. 2002, 36, 1048–1053. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Popendorf, K.J.; Fredricks, H.F.; Van Mooy, B.A.S. Molecular ion-independent quantification of polar glycerolipid classes in marine plankton using triple quadrupole MS. Lipids 2013, 48, 185–195. [Google Scholar] [CrossRef] [PubMed]
- White, D.A.; Pagarette, A.; Rooks, P.; Ali, S.T. The effect of sodium bicarbonate supplementation on growth and biochemical composition of marine microalgae cultures. J. Appl. Phycol. 2013, 25, 153–165. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N.; Somerfied, P.J.; Warwick, R.M. Changes in Marine Communities: An Approach to Statistical Analysis and Interpretation, 3rd ed.; PRIMER-E: Plymouth, UK, 2014. [Google Scholar]
- Clarke, K.R.; Somerfield, P.J.; Chapman, M.G. On resemblance measures for ecological studies, including taxonomic dissimilarities and a zero-adjusted Bray-Curtis coefficient for denuded assemblages. J. Exp. Mar. Biol. Ecol. 2006, 330, 55–80. [Google Scholar] [CrossRef]
- Clarke, K.R.; Green, R.H. Statistical design and analysis for a biological effects study. Mar. Ecol. Prog. Ser. 1988, 46, 213–226. [Google Scholar] [CrossRef]
- Clarke, K.R.; Ainsworth, M. A method of linking multivariate community structure to environmental variables. Mar. Ecol. Prog. Ser. 1993, 92, 205–219. [Google Scholar] [CrossRef]
- Herndl, G.J.; Reinthaler, T.; Teira, E.; van Aken, H.; Veth, C.; Pernthaler, A.; Pernthaler, J. Contribution of Archaea to total prokaryotic production in the deep Atlantic Ocean. Appl. Environ. Microbiol. 2005, 71, 2303–2309. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, E.; Melo, T.; Moreira, A.S.P.; Alves, E.; Domingues, P.; Calado, R.; Abreu, M.H.; Domingues, M.R. Decoding bioactive polar lipid profile of the macroalgae Codium tomentosum from a sustainable IMTA system using a lipidomic approach. Algal Res. 2015, 12, 388–397. [Google Scholar] [CrossRef]
- Da Costa, E.; Silva, J.; Mendonça, S.H.; Abreu, M.H.; Domingues, M.R. Lipidomic Approaches towards Deciphering Glycolipids from Microalgae as a Reservoir of Bioactive Lipids. Mar. Drugs 2016, 14, 101. [Google Scholar] [CrossRef] [PubMed]
- Roessler, P. Environmental control of glycerolipid metabolism in microalgae: Commercial implications and future research directions. J. Phycol. 1990, 26, 393–399. [Google Scholar] [CrossRef]
- Martin, G.J.O.; Hill, D.R.A.; Olmstead, I.L.D.; Bergamin, A.; Shears, M.J.; Dias, D.A.; Kentish, S.E.; Scales, P.J.; Botte, C.Y.; Callahan, D.L. Lipid profile remodeling in response to nitrogen deprivation in the microalgae Chlorella sp. (Trebouxiophyceae) and Nannochloropsis sp. (Eustigmatophyceae). PLoS ONE 2014, 9, e103389. [Google Scholar] [CrossRef] [PubMed]
- Sung Ho, C.; Thompson, G.A. Properties of a fatty acyl hydrolase preferentially attacking monogalactosyldiacylglycerols in Dunaliella salina chloroplasts. Biochim. Biophys. Acta 1986, 878, 353–359. [Google Scholar] [CrossRef]
- Li, X.; Moellering, E.R.; Liu, B.; Johnny, C.; Fedewa, M.; Sears, B.B.; Kuo, M.-H.; Benning, C. A Galactoglycerolipid Lipase Is Required for Triacylglycerol Accumulation and Survival Following Nitrogen Deprivation in Chlamydomonas reinhardtii. Plant. Cell 2012, 24, 4670–4686. [Google Scholar] [CrossRef] [PubMed]
- Cai, T.; Park, S.Y.; Li, Y. Nutrient recovery from wastewater streams by microalgae: Status and prospects. Renew. Sustain. Energy Rev. 2013, 19, 360–369. [Google Scholar] [CrossRef]
- Dormann, P.; Holzl, P. The role of glycolipids in photosynthesis. In Lipids in Photosynthesis: Essential and Regulatory Functions; Wada, H., Murata, N., Eds.; Springer Science+ Business Media B.V.: Berlin/Heidelberg, Germany, 2009; pp. 265–282. [Google Scholar]
- Lu, N.; Wei, D.; Chen, F.; Yang, S.-T. Lipidomic profiling reveals lipid regulation in the snow alga Chlamydomonas nivalis in response to nitrate or phosphate deprivation. Process. Biochem. 2013, 48, 605–613. [Google Scholar] [CrossRef]
- Jouhet, J.; Maréchal, E.; Block, M.A. Glycerolipid transfer for the building of membranes in plant cells. Prog. Lipid Res. 2007, 46, 37–55. [Google Scholar] [CrossRef] [PubMed]
- Shimojima, M. Biosynthesis and functions of the plant sulfolipid. Prog. Lipid Res. 2011, 50, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Sato, N. Roles of the acidic lipids sulfoquinovosyl diacylglycerol and phosphatidylglycerol in photosynthesis: Their specificity and evolution. J. Plant. Res. 2004, 117, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Kates, M. Glycolipids of higher plants, algae, yeasts and fungi. In Handb Lipid Res; Kates, M., Ed.; Springer: Boston, MA, USA, 1990; Volume 6, pp. 235–320. [Google Scholar]
- Suen, Y.; Hubbard, J.S.; Holzer, G.; Tornabene, T.G. Total lipid production of the green alga Nannochloropsis sp. QII under different nitrogen regimes. J. Phycol. 1987, 23, 289–296. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid Class | Carbon: Double Bonds | LN | HN | |||||
|---|---|---|---|---|---|---|---|---|
| Acyl Groups | Day 4 | Day 9 | Day 15 | Day 4 | Day 9 | Day 14 | ||
| MGDG | 34:7 | 18:3/16:4 | 78.4 ± 0.5 | 11.1 ± 2.0 | 1.6 ± 0.5 | 78.5 ± 0.8 | 62.5 ± 1.3 | 58.6 ± 3.7 |
| MGDG | 34:6 | 18:3/16:3, 18:2/16:4 | 14.6 ± 0.2 | 39.9 ± 1.7 | 41.5 ± 2.8 | 15.0 ± 0.4 | 24.4 ± 1.1 | 20.8 ± 1.0 |
| MGDG | 34:5 | 18:3/16:2 | 0.6 ± 0.9 | 29.9 ± 3.3 | 28.2 ± 3.0 | 1.4 ± 0.1 | 5.6 ± 0.6 | 6.4 ± 1.0 |
| MGDG | 34:4 | 18:1/16:3 | 0.6 ± 0.8 | 12.0 ± 0.7 | 13.7 ± 0.8 | 1.6 ± 0.3 | 2.3 ± 0.3 | 3.8 ± 0.5 |
| MGDG | 34:3 | 18:1/16:2 | 0.0 ± 0.0 | 6.3 ± 0.8 | 10.1 ± 0.7 | 0.0 ± 0.0 | 0.8 ± 0.0 | 1.6 ± 0.4 |
| MGDG | 36:6 | 18:3/18:3 | 5.1 ± 0.7 | 0.7 ± 1.0 | 4.2 ± 0.2 | 3.5 ± 0.7 | 3.4 ± 0.5 | 5.8 ± 0.0 |
| DGDG | 34:7 | 18:3/16:4 | 14.6 ± 1.1 | 1.4 ± 0.6 | 0.0 ± 0.0 | 12.5 ± 0.7 | 20.1 ± 2.4 | 12.7 ± 0.9 |
| DGDG | 34:6 | 18:3/16:3 | 33.4 ± 2.4 | 22.3 ± 3.2 | 21.3 ± 0.4 | 31.3 ± 1.0 | 17.0 ± 1.4 | 14.5 ± 0.6 |
| DGDG | 34:5 | 18:2/16:3 | 10.2 ± 0.0 | 22.4 ± 1.0 | 21.7 ± 2.0 | 11.8 ± 0.5 | 10.2 ± 0.1 | 9.4 ± 0.8 |
| DGDG | 34:4 | 18:2/16:2 | 5.3 ± 0.0 | 9.0 ± 0.9 | 10.2 ± 0.2 | 9.5 ± 1.7 | 12.4 ± 1.0 | 10.9 ± 0.8 |
| DGDG | 34:3 | 18:3/16:0 | 16.1 ± 2.1 | 22.0 ± 1.4 | 21.4 ± 0.9 | 13.7 ± 0.5 | 11.4 ± 0.6 | 11.5 ± 1.1 |
| DGDG | 34:2 | 18:2/16:0 | 7.5 ± 0.8 | 14.6 ± 0.7 | 15.2 ± 0.5 | 8.2 ± 0.3 | 12.0 ± 0.7 | 14.9 ± 0.1 |
| DGDG | 34:1 | 18:1/16:0 | 5.6 ± 0.2 | 8.4 ± 0.2 | 10.3 ± 1.4 | 5.8 ± 0.7 | 8.7 ± 0.2 | 8.8 ± 0.5 |
| DGDG | 36:6 | 18:3/18:3 | 6.0 ± 0.2 | 0.0 ± 0.0 | 0.0 ± 0.0 | 5.4 ± 1.3 | 5.6 ± 1.3 | 8.9 ± 0.6 |
| DGDG | 36:5 | 18:3/18:2 | 1.3 ± 0.1 | 0.0 ± 0.0 | 0.0 ± 0.0 | 1.8 ± 0.5 | 1.8 ± 0.3 | 4.7 ± 0.3 |
| DGDG | 36:4 | 18:1/18:3 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.8 ± 1.1 | 3.5 ± 0.2 |
| SQDG | 32:0 | 16:0/16:0 | 37.5 ± 1.0 | 38.9 ± 2.5 | 37.9 ± 2.4 | 36.2 ± 1.3 | 32.5 ± 0.9 | 30.0 ± 3.7 |
| SQDG | 34:3 | 18:3/16:0 | 45.4 ± 0.2 | 27.1 ± 2.1 | 26.5 ± 0.7 | 46.2 ± 0.4 | 46.9 ± 3.3 | 45.5 ± 0.9 |
| SQDG | 34:2 | 18:2/16:2 | 12.0± 0.0 | 18.2 ± 0.1 | 17.0 ± 0.1 | 12.1 ± 0.2 | 14.5 ± 2.3 | 17.4 ± 2.0 |
| SQDG | 34:1 | 18:1/16:0 | 5.1 ± 1.2 | 15.8 ± 0.3 | 18.6 ± 1.8 | 5.5 ± 1.9 | 6.1 ± 0.2 | 7.1 ± 0.8 |
| PG | 32:1 | 16:0/16:1 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 10.3 ± 0.2 | 3.2 ± 0.2 | 1.3 ± 0.1 |
| PG | 34:4 | 18:3/16:1 | 60.7 ± 0.0 | 31.3 ± 5.0 | 0.0 ± 0.0 | 10.8 ± 3.4 | 31.2 ± 2.5 | 33.2 ± 0.9 |
| PG | 34:3 | 18:3/16:0, 18:2/16:1 | 23.4 ± 0.7 | 23.5 ± 0.6 | 0.0 ± 0.0 | 1.8 ± 2.5 | 22.3 ± 0.5 | 24.0 ± 3.7 |
| PG | 34:2 | 18:1/16:1, 18:2/16:0 | 8.1 ± 1.0 | 16.5 ± 0.7 | 23.2 ± 8.0 | 43.9 ± 0.7 | 23.2 ± 2.4 | 21.6 ± 1.0 |
| PG | 34:1 | 18:1/16:0 | 7.8 ± 0.2 | 28.7 ± 5.0 | 76.8 ± 8.0 | 28.0 ± 0.1 | 14.8 ± 0.1 | 13.2 ± 1.4 |
| PG | 36:2 | 18:1/18:1 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 5.2 ± 0.3 | 5.3 ± 0.6 | 6.7 ± 2.2 |
| PE | 32:1 | 16:0/16:1 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 14.2 ± 0.0 | 10.8 ± 1.1 | 10.1 ± 1.7 |
| PE | 33:1 | 16:0/17:1 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.4 ± 0.2 | 0.8 ± 0.3 | 1.1 ± 0.0 |
| PE | 34:2 | 18:1/16:1 | 2.3 ± 0.3 | 13.3 ± 2.3 | 3.7 ± 5.2 | 50.7 ± 2.1 | 48.3 ± 0.3 | 47.5 ± 1.0 |
| PE | 34:1 | 18:1/16:0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 28.4 ± 0.1 | 22.5 ± 2.3 | 21.3 ± 1.6 |
| PE | 34:0 | 18:0/16:0 | 11.2 ± 1.6 | 0.0 ± 0.0 | 17.5 ± 11.0 | 1.4 ± 2.0 | 2.5 ± 0.6 | 2.5 ± 0.0 |
| PE | 36:5 | 18:2/18:3 | 46.2 ± 0.6 | 22.0 ± 1.4 | 6.6 ± 9.3 | 0.7 ± 0.2 | 2.9 ± 0.0 | 1.7 ± 1.1 |
| PE | 36:4 | 18:1/18:3, 18:2/18:2 | 29.7 ± 1.9 | 36.6 ± 0.4 | 27.2 ± 2.6 | 0.0 ± 0.0 | 2.2 ± 0.0 | 1.4 ± 2.0 |
| PE | 36:3 | 18:2/18:1 | 10.6 ± 0.6 | 28.0 ± 3.3 | 29.6 ± 1.1 | 0.0 ± 0.0 | 0.4 ± 0.5 | 0.7 ± 1.0 |
| PE | 36:2 | 18:1/18:1 | 0.0 ± 0.0 | 0.0 ± 0.0 | 15.5 ± 10.6 | 2.3 ± 0.4 | 3.3 ± 0.0 | 5.5 ± 0.1 |
| PC | 34:4 | 18:3/16:1 | 5.3 ± 0.1 | 1.8 ± 0.8 | 1.8 ± 0.0 | 4.4 ± 0.2 | 6.5 ± 1.2 | 5.1 ± 0.0 |
| PC | 34:3 | 18:3/16:0, 18:2/16:1 | 15.1 ± 0.2 | 6.7 ± 1.0 | 6.2 ± 1.7 | 13.5 ±1.8 | 9.6 ± 0.2 | 10.4 ± 0.7 |
| PC | 34:2 | 18:2/16:0, 18:1/16:1 | 3.5 ± 0.0 | 10.4 ± 1.7 | 10.3 ± 0.2 | 5.3 ± 0.3 | 3.3 ± 0.3 | 4.7 ± 0.9 |
| PC | 36:6 | 18:3/18:3 | 36.1 ± 1.2 | 15.1 ± 4.3 | 14.3 ± 2.6 | 36.4 ± 0.1 | 36.4 ± 0.1 | 28.4 ± 6.8 |
| PC | 36:5 | 18:3/18:2 | 22.8 ± 0.2 | 26.6 ± 1.3 | 23.5 ± 4.9 | 20.7 ± 0.2 | 20.9 ± 1.5 | 18.7 ± 0.2 |
| PC | 36:4 | 18:3/18:1 | 11.6 ± 0.7 | 18.7 ± 0.1 | 20.0 ±2.1 | 11.3 ± 0.5 | 11.7 ± 0.6 | 17.0 ± 1.9 |
| PC | 36:3 | 18:2/18:1 | 4.1 ± 0.2 | 10.9 ± 2.3 | 13.1 ± 0.5 | 4.0 ± 0.6 | 4.9 ± 0.1 | 8.9 ± 1.3 |
| PC | 36:2 | 18:1/18:1 | 1.7 ± 0.2 | 9.8 ± 0.3 | 11.1 ± 3.2 | 4.4 ± 0.8 | 3.3 ± 0.6 | 6.7 ± 2.1 |
| DGTS | 34:4 | 18:4/16:0 | 45.0 ± 0.1 | 25.7 ± 1.7 | 24.3 ± 0.8 | 44.6 ± 2.7 | 33.1 ± 1.2 | 31.6 ± 0.5 |
| DGTS | 34:3 | 18:3/16:0 | 16.3 ± 0.6 | 23.1 ± 1.0 | 28.8 ± 1.0 | 14.4 ± 0.9 | 11.1 ± 0.2 | 12.1 ± 1.5 |
| DGTS | 34:2 | 18:2/16:0 | 1.8 ± 0.0 | 12.0 ± 2.1 | 15.0 ± 0.3 | 0.0 ± 0.0 | 1.4 ± 1.9 | 3.5 ± 1.8 |
| DGTS | 36:7 | 18:3/18:4 | 14.8 ± 0.9 | 3.8 ± 0.4 | 2.3 ± 0.1 | 15.6 ± 0.8 | 23.7 ± 1.7 | 19.6 ± 4.6 |
| DGTS | 36:6 | 18:3/18:3 | 14.1 ± 0.2 | 9.1 ± 0.8 | 6.3 ± 0.7 | 14.8 ± 1.6 | 16.9 ± 0.3 | 15.2 ± 0.7 |
| DGTS | 36:5 | 18:3/18:2 | 6.3 ± 0.6 | 14.5 ± 0.8 | 11.3 ± 0.1 | 7.8 ± 1.0 | 9.2 ± 0.5 | 11.0 ± 1.0 |
| DGTS | 36:4 | 18:3/18:1 | 1.7 ± 0.1 | 11.4 ± 0.5 | 12.0 ± 0.2 | 2.9 ± 0.2 | 4.6 ± 0.5 | 6.9 ± 1.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
White, D.A.; Rooks, P.A.; Kimmance, S.; Tait, K.; Jones, M.; Tarran, G.A.; Cook, C.; Llewellyn, C.A. Modulation of Polar Lipid Profiles in Chlorella sp. in Response to Nutrient Limitation. Metabolites 2019, 9, 39. https://doi.org/10.3390/metabo9030039
White DA, Rooks PA, Kimmance S, Tait K, Jones M, Tarran GA, Cook C, Llewellyn CA. Modulation of Polar Lipid Profiles in Chlorella sp. in Response to Nutrient Limitation. Metabolites. 2019; 9(3):39. https://doi.org/10.3390/metabo9030039
Chicago/Turabian StyleWhite, Daniel A., Paul A. Rooks, Susan Kimmance, Karen Tait, Mark Jones, Glen A. Tarran, Charlotte Cook, and Carole A. Llewellyn. 2019. "Modulation of Polar Lipid Profiles in Chlorella sp. in Response to Nutrient Limitation" Metabolites 9, no. 3: 39. https://doi.org/10.3390/metabo9030039
APA StyleWhite, D. A., Rooks, P. A., Kimmance, S., Tait, K., Jones, M., Tarran, G. A., Cook, C., & Llewellyn, C. A. (2019). Modulation of Polar Lipid Profiles in Chlorella sp. in Response to Nutrient Limitation. Metabolites, 9(3), 39. https://doi.org/10.3390/metabo9030039