The Effects of Doxorubicin-based Chemotherapy and Omega-3 Supplementation on Mouse Brain Lipids

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Experiments and Diets
2.1.1. Study 1
2.1.2. Study 2
2.2. Chemicals
2.3. Targeted SPM Analysis (Study 1)
2.3.1. Brain Tissue Extraction
2.3.2. UHPLC-MS/MS Analyses
2.3.3. Statistical Analysis
2.4. Untargeted Lipidomics (Study 2)
2.4.1. Hippocampal Tissue Extraction
2.4.2. UHPLC-MS Analyses
2.4.3. Data Processing
2.5. Statistical Analysis
2.6. Compound Identification
3. Results
3.1. SPM Detection and Quantification in Whole Brain Extract
3.2. Untargeted Lipidomics
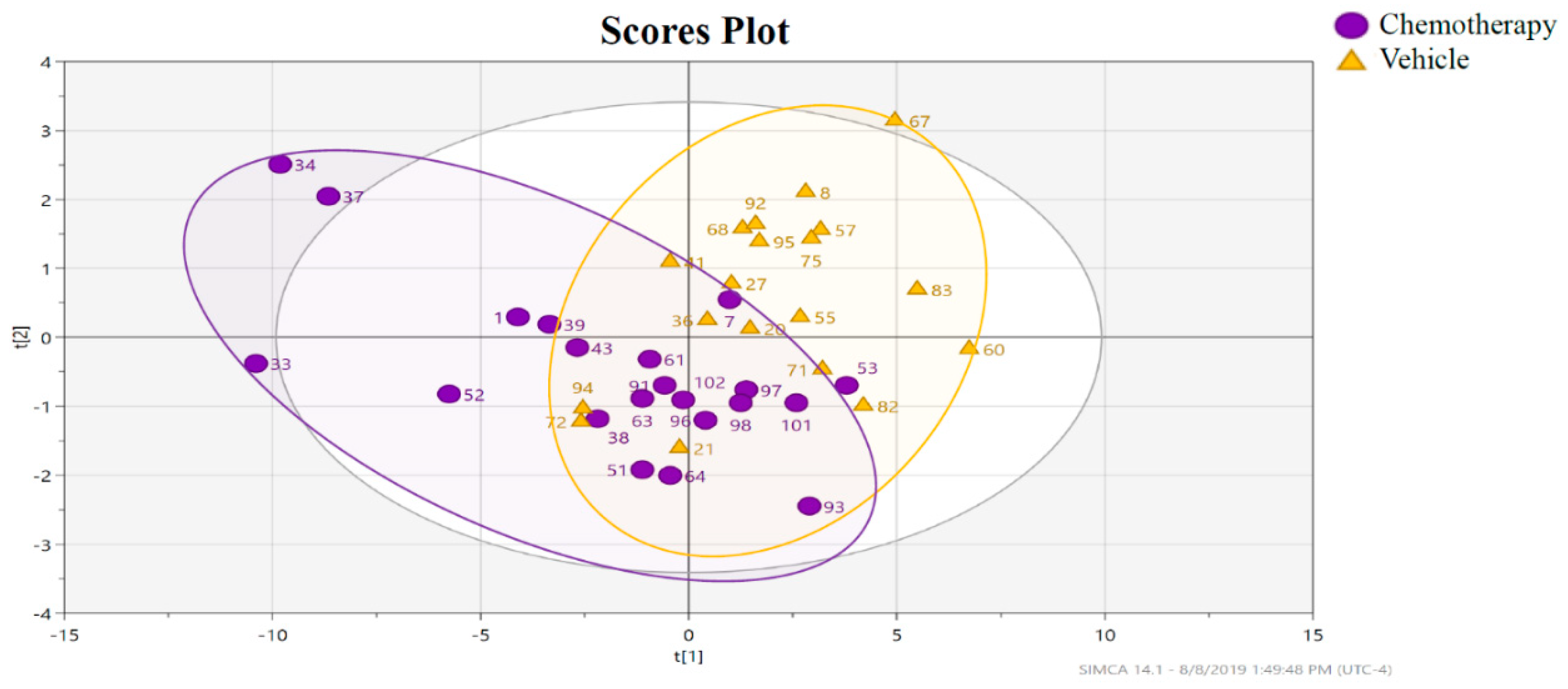
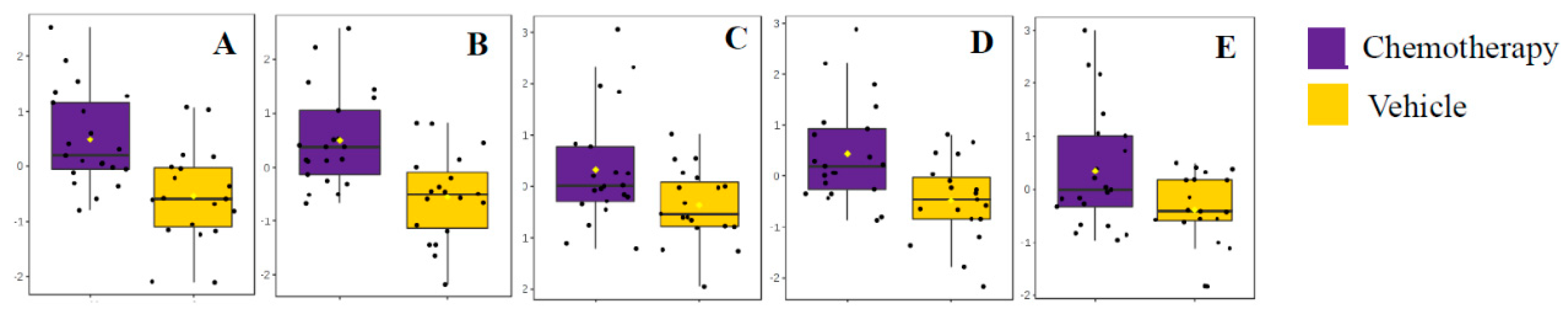
3.2.1. Impact of Chemotherapy on Hippocampal Lipidome
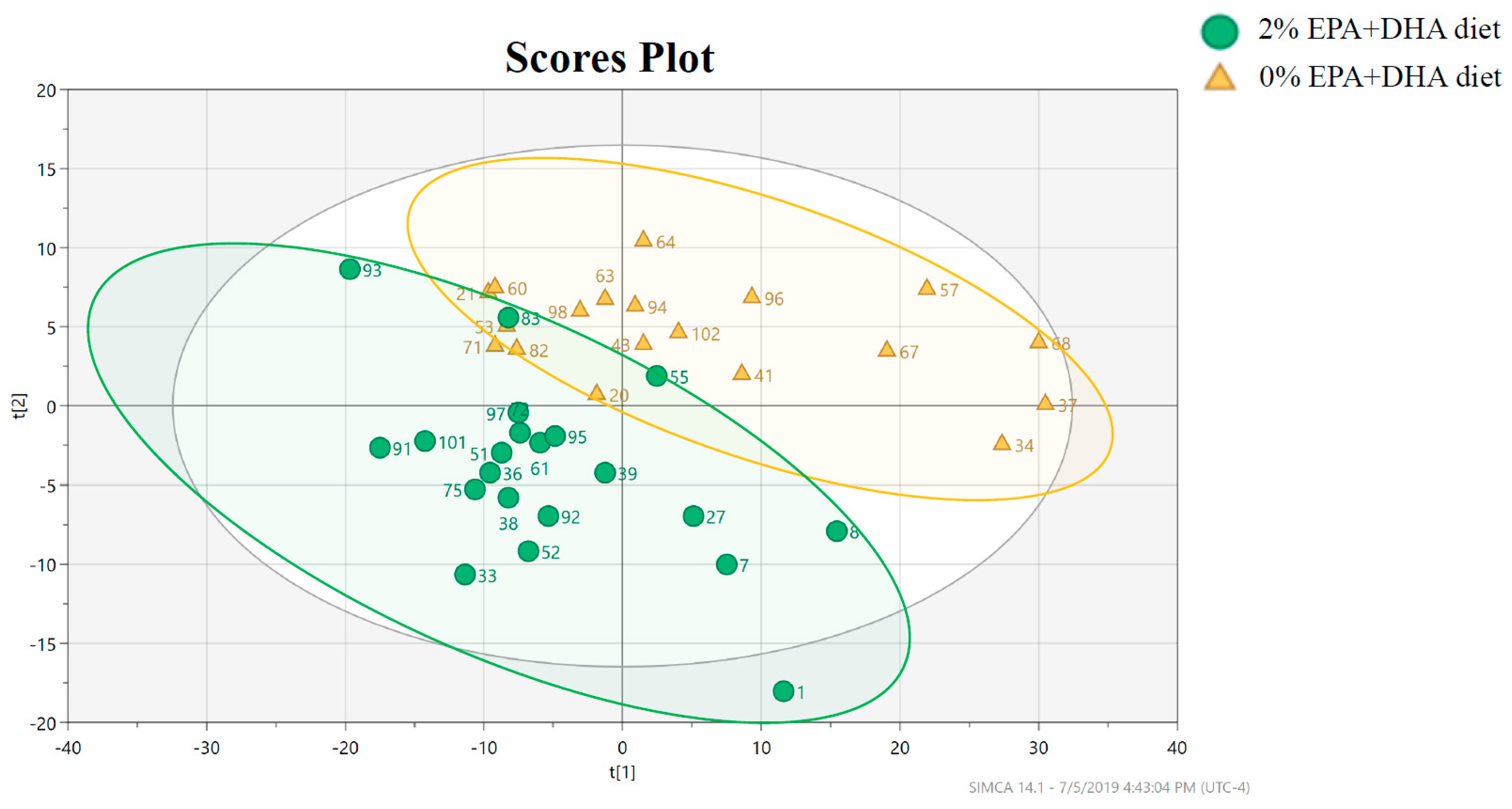
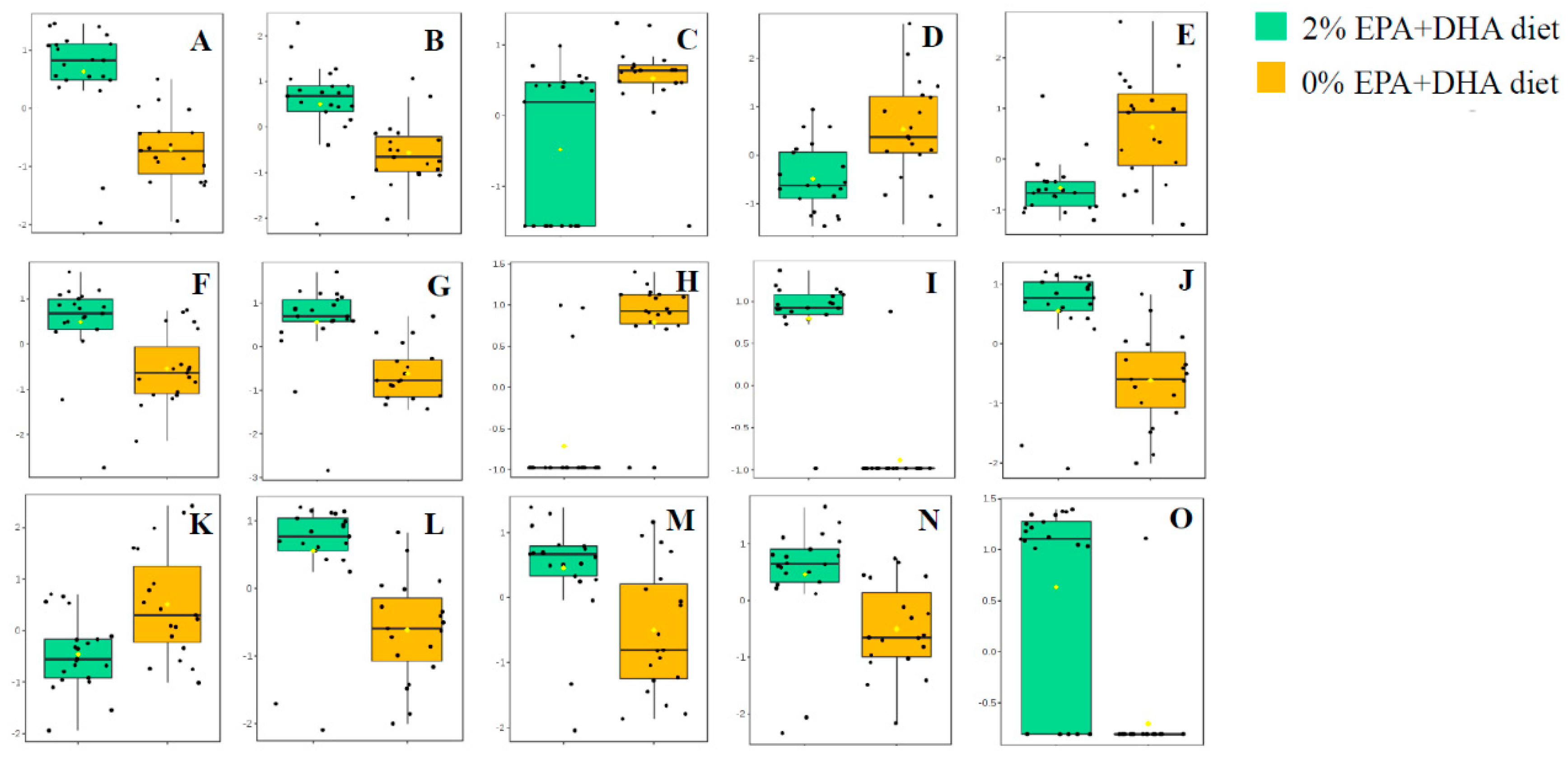
3.2.2. Impact of EPA + DHA Supplementation on the Hippocampal Lipidome
3.2.3. Interaction between the Impact of Chemotherapy and of Omega-3 Supplementation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Amarante, M.K.; de Sousa Pereira, N.; Vitiello, G.A.F.; Watanabe, M.A.E. Involvement of a mouse mammary tumor virus (MMTV) homologue in human breast cancer: Evidence for, against and possible causes of controversies. Microb. Pathog. 2019, 130, 283–294. [Google Scholar] [CrossRef] [PubMed]
- World Health Organisation. Latest global cancer data. Int. Agency Res. Cancer 2018, 263, 13–15. [Google Scholar]
- Mackey, J.R.; Martin, M.; Pienkowski, T.; Rolski, J.; Guastalla, J.P.; Sami, A.; Glaspy, J.; Juhos, E.; Wardley, A.; Fornander, T.; et al. Adjuvant docetaxel, doxorubicin, and cyclophosphamide in node-positive breast cancer: 10-year follow-up of the phase 3 randomised BCIRG 001 trial. Lancet Oncol. 2013, 14, 72–80. [Google Scholar] [CrossRef]
- Carvalho, C.; Santos, R.; Cardoso, S.; Correia, S.; Oliveira, P.; Santos, M.; Moreira, P. Doxorubicin: The good, the bad and the ugly effect. Curr. Med. Chem. 2009, 16, 3267–3285. [Google Scholar] [CrossRef] [PubMed]
- Wefel, J.S.; Saleeba, A.K.; Buzdar, A.U.; Meyers, C.A. Acute and late onset cognitive dysfunction associated with chemotherapy in women with breast cancer. Cancer 2010, 116, 3348–3356. [Google Scholar] [CrossRef] [PubMed]
- Brezden, C.B.; Phillips, K.-A.; Abdolell, M.; Bunston, T.; Tannock, I.F. Cognitive function in breast cancer patients receiving adjuvant chemotherapy. J. Clin. Oncol. 2000, 18, 2695–2701. [Google Scholar] [CrossRef] [PubMed]
- Schagen, S.B.; Van Dam, F.S.A.M.; Muller, M.J.; Boogerd, W.; Lindeboom, J.; Bruning, P.F. Cognitive deficits after postoperative adjuvant chemotherapy for breast carcinoma. Cancer 1999, 85, 640–650. [Google Scholar] [CrossRef]
- Shilling, V.; Jenkins, V.; Morris, R.; Deutsch, G.; Bloomfield, D. The effects of adjuvant chemotherapy on cognition in women with breast cancer—Preliminary results of an observational longitudinal study. Breast 2005, 14, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Jansen, C.E.; Dodd, M.J.; Miaskowski, C.A.; Dowling, G.A.; Kramer, J. Preliminary results of a longitudinal study of changes in cognitive function in breast cancer patients undergoing chemotherapy with doxorubicin and cyclophosphamide. Psychooncology 2008, 17, 1189–1195. [Google Scholar] [CrossRef] [PubMed]
- Orchard, T.S.; Gaudier-Diaz, M.M.; Phuwamongkolwiwat-Chu, P.; Andridge, R.; Lustberg, M.B.; Bomser, J.; Cole, R.M.; Belury, M.A.; Devries, A.C. Low sucrose, omega-3 enriched diet has region-specific effects on neuroinflammation and synaptic function markers in a mouse model of doxorubicin-based chemotherapy. Nutrients 2018, 10, 2004. [Google Scholar] [CrossRef]
- Tangpong, J.; Cole, M.P.; Sultana, R.; Joshi, G.; Estus, S.; Vore, M.; St. Clair, W.; Ratanachaiyavong, S.; St. Clair, D.K.; Butterfield, D.A. Adriamycin-induced, TNF-α-mediated central nervous system toxicity. Neurobiol. Dis. 2006, 23, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Bigotte, L.; Olsson, Y. Cytofluorescence localization of adriamycin in the nervous system. Acta Neuropathol. 2004, 58, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Seruga, B.; Zhang, H.; Bernstein, L.J.; Tannock, I.F. Cytokines and their relationship to the symptoms and outcome of cancer. Nat. Rev. Cancer 2008, 8, 887–899. [Google Scholar] [CrossRef]
- Tangpong, J.; Cole, M.P.; Sultana, R.; Estus, S.; Vore, M.; St. Clair, W.; Ratanachaiyavong, S.; St. Clair, D.K.; Butterfield, D.A. Adriamycin-mediated nitration of manganese superoxide dismutase in the central nervous system: Insight into the mechanism of chemobrain. J. Neurochem. 2007, 100, 191–201. [Google Scholar] [CrossRef]
- Kirkpatrick, L.; Brady, S. Basic Neurochemistry: Molecular, Cellular and Medical Aspects; Elsevier: Oxford, UK, 2008; ISBN 978-1-4939-3772-1. [Google Scholar]
- Serhan, C.N. Pro-resolving lipid mediators are leads for resolution physiology. Nature 2014, 510, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Finocchiaro, C.; Segre, O.; Fadda, M.; Monge, T.; Scigliano, M.; Schena, M.; Tinivella, M.; Tiozzo, E.; Catalano, M.G.; Pugliese, M.; et al. Effect of n-3 fatty acids on patients with advanced lung cancer: A double-blind, placebo-controlled study. Br. J. Nutr. 2012, 108, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Alfano, C.M.; Imayama, I.; Neuhouser, M.L.; Kiecolt-Glaser, J.K.; Smith, A.W.; Meeske, K.; McTiernan, A.; Bernstein, L.; Baumgartner, K.B.; Ulrich, C.M.; et al. Fatigue, inflammation, and -γ3 and γ-6 fatty acid intake among breast cancer survivors. J. Clin. Oncol. 2012, 30, 1280–1287. [Google Scholar] [CrossRef] [PubMed]
- American Cancer Society. Cancer Treatment & Survivorship Facts & Figures 2016-2017; American Cancer Society: Atlanta, GA, USA, 2019. [Google Scholar]
- Villa, A.; Rizzi, N.; Vegeto, E.; Ciana, P.; Maggi, A. Estrogen accelerates the resolution of inflammation in macrophagic cells. Sci. Rep. 2015, 5, 15224. [Google Scholar] [CrossRef] [PubMed]
- Loiola, R.A.; Wickstead, E.S.; Solito, E.; McArthur, S. Estrogen promotes pro-resolving microglial behavior and phagocytic cell clearance through the actions of annexin A1. Front. Endocrinol. 2019, 10, 420. [Google Scholar] [CrossRef] [PubMed]
- Gubbels Bupp, M.R. Sex, the aging immune system, and chronic disease. Cell. Immunol. 2015, 294, 102–110. [Google Scholar] [CrossRef]
- Lustberg, M.B.; Orchard, T.S.; Reinbolt, R.; Andridge, R.; Pan, X.; Belury, M.; Cole, R.; Logan, A.; Layman, R.; Ramaswamy, B.; et al. Randomized placebo-controlled pilot trial of omega 3 fatty acids for prevention of aromatase inhibitor-induced musculoskeletal pain. Breast Cancer Res. Treat. 2018, 167, 709–718. [Google Scholar] [CrossRef] [PubMed]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2007, 22, 659–661. [Google Scholar] [CrossRef] [PubMed]
- Colas, R.A.; Shinohara, M.; Dalli, J.; Chiang, N.; Serhan, C.N. Identification and signature profiles for pro-resolving and inflammatory lipid mediators in human tissue. Am. J. Physiol. Physiol. 2014, 307, C39–C54. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 10 July 2019).
- Ash, M.; Chang, A.; Heiberger, R.; Kerns, G.J.; Lancelot, R.; Lesnoff, M.; Muenchen, R.; Murdoch, D.; Neuwirth, E.; Putler, D.; et al. Package ‘ Rcmdr ’. Available online: http://CRAN.R-project.org/package=Rcmdr (accessed on 14 November 2018).
- Bates, D.; Maechler, M.; Bolker, B. lme4: Linear Mixed-Effects Models Using S4 Classes (R Package Version 0.999999-0). Available online: https://www.scirp.org/(S(351jmbntvnsjt1aadkposzje))/reference/ReferencesPapers.aspx?ReferenceID=1334359 (accessed on 9 September 2019).
- Bozek, K.; Wei, Y.; Yan, Z.; Liu, X.; Xiong, J.; Sugimoto, M.; Tomita, M.; Pääbo, S.; Sherwood, C.C.; Hof, P.R.; et al. Organization and evolution of brain lipidome revealed by large-scale analysis of human, chimpanzee, macaque, and mouse tissues. Neuron 2015, 85, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Psychogios, N.; Young, N.; Wishart, D.S. MetaboAnalyst: A web server for metabolomic data analysis and interpretation. Nucleic Acids Res. 2009, 37, 652–660. [Google Scholar] [CrossRef] [PubMed]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; Vandergheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data-independent MS/MS deconvolution for comprehensive metabolome analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Tsugawa, H.; Kind, T.; Nakabayashi, R.; Yukihira, D.; Tanaka, W.; Cajka, T.; Saito, K.; Fiehn, O.; Arita, M. Hydrogen rearrangement rules: Computational MS/MS fragmentation and structure elucidation using MS-FINDER Software. Anal. Chem. 2016, 88, 7946–7958. [Google Scholar] [CrossRef] [PubMed]
- Sumner, L.W.; Lei, Z.; Nikolau, B.J.; Saito, K.; Roessner, U.; Trengove, R. Proposed quantitative and alphanumeric metabolite identification metrics. Metabolomics 2014, 10, 1047–1049. [Google Scholar] [CrossRef]
- Ikuo, I.; Katsumi, I.; Michihiro, S. Absorption and transport of base moieties of phosphatidylcholine and phosphatidylethanolamine in rats. Biochim. Biophys. Acta (BBA)/Lipids Lipid Metab. 1987, 921, 245–253. [Google Scholar] [CrossRef]
- Andersen, P.; Morris, R.; Amaral, D.; Bliss, T.; O’Keefe, J. The Hippocampus Book; Oxford University Press: New York, NY, USA, 2007; ISBN 9780195100273. [Google Scholar]
- Basil, M.C.; Levy, B.D. Specialized pro-resolving mediators: Endogenous regulators of infection and inflammation. Nat. Rev. Immunol. 2016, 16, 51–67. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Puerta, E.; Cedazo-Minguez, A.; Hjorth, E.; Schultzberg, M. Insufficient Resolution response in the hippocampus of a senescence-accelerated mouse model—SAMP8. J. Mol. Neurosci. 2014, 55, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Hopperton, K.E.; Trépanier, M.O.; James, N.C.E.; Chouinard-Watkins, R.; Bazinet, R.P. Fish oil feeding attenuates neuroinflammatory gene expression without concomitant changes in brain eicosanoids and docosanoids in a mouse model of Alzheimer’s disease. Brain. Behav. Immun. 2018, 69, 74–90. [Google Scholar] [CrossRef] [PubMed]
- Trépanier, M.O.; Hopperton, K.E.; Giuliano, V.; Masoodi, M.; Bazinet, R.P. Increased brain docosahexaenoic acid has no effect on the resolution of neuroinflammation following intracerebroventricular lipopolysaccharide injection. Neurochem. Int. 2018, 118, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Orr, S.K.; Palumbo, S.; Bosetti, F.; Mount, H.T.; Kang, J.X.; Greenwood, C.E.; Ma, D.W.L.; Serhan, C.N.; Bazinet, R.P. Unesterified docosahexaenoic acid is protective in neuroinflammation. J. Neurochem. 2013, 127, 378–393. [Google Scholar] [CrossRef] [PubMed]
- Belayev, L.; Khoutorova, L.; Atkins, K.D.; Eady, T.N.; Hong, S.; Lu, Y.; Obenaus, A.; Bazan, N.G. Docosahexaenoic acid therapy of experimental ischemic stroke. Transl. Stroke Res. 2011, 2, 33–41. [Google Scholar] [CrossRef]
- Marcheselli, V.L.; Hong, S.; Lukiw, W.J.; Tian, X.H.; Gronert, K.; Musto, A.; Hardy, M.; Gimenez, J.M.; Chiang, N.; Serhan, C.N.; et al. Novel docosanoids inhibit brain ischemia-reperfusion-mediated leukocyte infiltration and pro-inflammatory gene expression. J. Biol. Chem. 2003, 278, 43807–43817. [Google Scholar] [CrossRef]
- Levy, B.D.; Clish, C.B.; Schmidt, B.; Gronert, K.; Serhan, C.N. Lipid mediator class switching during acute inflammation: Signals in resolution. Nat. Immunol. 2001, 2, 612–619. [Google Scholar] [CrossRef]
- Chavko, M.; Nemoto, E.M.; Melick, J.A. Regional lipid composition in the rat brain. Mol. Chem. Neuropathol. 1993, 18, 123–131. [Google Scholar] [CrossRef]
- Miranda, A.M.; Bravo, F.V.; Chan, R.B.; Sousa, N.; Di Paolo, G.; Oliveira, T.G. Differential lipid composition and regulation along the hippocampal longitudinal axis. Transl. Psychiatry 2019, 9, 144. [Google Scholar] [CrossRef]
- Mu, Y.; Gage, F.H. Adult hippocampal neurogenesis and its role in Alzheimer’s disease. Mol. Neurodegener. 2011, 6, 85. [Google Scholar] [CrossRef]
- Kuruba, R.; Hattiangady, B.; Shetty, A.K. Hippocampal neurogenesis and neural stem cells in temporal lobe epilepsy. Epilepsy Behav. 2009, 14, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Harrison, P.J. The hippocampus in schizophrenia: A review of the neuropathological evidence and its pathophysiological implications. Psychopharmacology 2004, 174, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Cunnane, S.C.; Schneider, J.A.; Tangney, C.; Tremblay-Mercier, J.; Fortier, M.; Bennett, D.A.; Morris, M.C. Plasma and brain fatty acid profiles in mild cognitive impairment and alzheimer’s disease. J. Alzheimer’s Dis. 2012, 29, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Manicke, N.E.; Wiseman, J.M.; Ifa, D.R.; Cooks, R.G. Desorption electrospray ionization (DESI) mass spectrometry and tandem mass spectrometry (MS/MS) of phospholipids and sphingolipids: Ionization, adduct formation, and fragmentation. J. Am. Soc. Mass Spectrom. 2008, 19, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Meng, H.M.; Hu, G.R.; Li, F.L. Biosynthesis of nervonic acid and perspectives for its production by microalgae and other microorganisms. Appl. Microbiol. Biotechnol. 2018, 102, 3027–3035. [Google Scholar] [CrossRef] [PubMed]
- Martínez, M.; Mougan, I. Fatty acid composition of human brain phospholipids during normal development. J. Neurochem. 2002, 71, 2528–2533. [Google Scholar] [CrossRef] [PubMed]
- Astarita, G.; Jung, K.M.; Vasilevko, V.; DiPatrizio, N.V.; Martin, S.K.; Cribbs, D.H.; Head, E.; Cotman, C.W.; Piomelli, D. Elevated stearoyl-CoA desaturase in brains of patients with Alzheimer’s disease. PLoS ONE 2011, 6, e24777. [Google Scholar] [CrossRef]
- Nasaruddin, M.L.; Hölscher, C.; Kehoe, P.; Graham, S.F.; Green, B.D. Wide-ranging alterations in the brain fatty acid complement of subjects with late Alzheimer’s disease as detected by GC-MS. Am. J. Transl. Res. 2016, 8, 154–165. [Google Scholar]
- Nasaruddin, M.L.; Pan, X.; McGuinness, B.; Passmore, P.; Kehoe, P.G.; Hölscher, C.; Graham, S.F.; Green, B.D. Evidence that parietal lobe fatty acids may be more profoundly affected in moderate alzheimer’s disease (AD) pathology than in severe AD pathology. Metabolites 2018, 8, 69. [Google Scholar] [CrossRef]
- Iuliano, L.; Pacelli, A.; Ciacciarelli, M.; Zerbinati, C.; Fagioli, S.; Piras, F.; Orfei, M.D.; Bossù, P.; Pazzelli, F.; Serviddio, G.; et al. Plasma fatty acid lipidomics in amnestic mild cognitive impairment and Alzheimer’s disease. J. Alzheimer’s Dis. 2013, 36, 545–553. [Google Scholar]
- Kageyama, Y.; Kasahara, T.; Nakamura, T.; Hattori, K.; Deguchi, Y.; Tani, M.; Kuroda, K.; Yoshida, S.; Goto, Y.I.; Inoue, K.; et al. Plasma nervonic acid is a potential biomarker for major depressive disorder: A pilot study. Int. J. Neuropsychopharmacol. 2018, 21, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Carmeli, C.; Donati, A.; Antille, V.; Viceic, D.; Ghika, J.; von Gunten, A.; Clarke, S.; Meuli, R.; Frackowiak, R.S.; Knyazeva, M.G. Demyelination in mild cognitive impairment suggests progression path to Alzheimer’s disease. PLoS ONE 2013, 8, e72759. [Google Scholar] [CrossRef] [PubMed]
- Kelly, L.; Grehan, B.; Della Chiesa, A.; O’Mara, S.M.; Downer, E.; Sahyoun, G.; Massey, K.A.; Nicolaou, A.; Lynch, M.A. The polyunsaturated fatty acids, EPA and DPA exert a protective effect in the hippocampus of the aged rat. Neurobiol. Aging 2011, 32, 2318-e1. [Google Scholar] [CrossRef] [PubMed]
- Minogue, A.M.; Lynch, A.M.; Loane, D.J.; Herron, C.E.; Lynch, M.A. Modulation of amyloid-β-induced and age-associated changes in rat hippocampus by eicosapentaenoic acid. J. Neurochem. 2007, 103, 914–926. [Google Scholar] [CrossRef] [PubMed]
- Taneo, J.; Adachi, T.; Yoshida, A.; Takayasu, K.; Takahara, K.; Inaba, K. Amyloid β oligomers induce interleukin-1β production in primary microglia in a cathepsin B- and reactive oxygen species-dependent manner. Biochem. Biophys. Res. Commun. 2015, 458, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.; Kaur, G.; Larsen, A.; Loh, S.P.; Linderborg, K.; Weisinger, H.S.; Turchini, G.M.; Cameron-Smith, D.; Sinclair, A.J. A short-term n-3 DPA supplementation study in humans. Eur. J. Nutr. 2013, 52, 895–904. [Google Scholar] [CrossRef]
- Cansev, M.; Wurtman, R.J. Chronic administration of docosahexaenoic acid or eicosapentaenoic acid, but not arachidonic acid, alone or in combination with uridine, increases brain phosphatide and synaptic protein levels in gerbils. Neuroscience 2007, 148, 421–431. [Google Scholar] [CrossRef]
- Akbar, M.; Calderon, F.; Wen, Z.; Kim, H.-Y. Docosahexaenoic acid: A positive modulator of Akt signaling in neuronal survival. Proc. Natl. Acad. Sci. USA 2005, 102, 10858–10863. [Google Scholar] [CrossRef]
- Ginsberg, L.; Rafique, S.; Xuereb, J.H.; Rapoport, S.I.; Gershfeld, N.L. Disease and anatomic specificity of ethanolamine plasmalogen deficiency in Alzheimer’s disease brain. Brain Res. 1995, 698, 223–226. [Google Scholar] [CrossRef]
- Wood, P.L.; Barnette, B.L.; Kaye, J.A.; Quinn, J.F.; Woltjer, R.L. Non-targeted lipidomics of CSF and frontal cortex grey and white matter in control, mild cognitive impairment, and Alzheimer’s disease subjects. Acta Neuropsychiatr. 2015, 27, 270–278. [Google Scholar] [CrossRef]
- Yamashita, S.; Kiko, T.; Fujiwara, H.; Hashimoto, M.; Nakagawa, K.; Kinoshita, M.; Furukawa, K.; Arai, H.; Miyazawa, T. Alterations in the levels of amyloid-β, phospholipid hydroperoxide, and plasmalogen in the blood of patients with Alzheimer’s disease: Possible interactions between amyloid-β and these lipids. J. Alzheimer’s Dis. 2016, 50, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Fujino, T.; Yamada, T.; Asada, T.; Tsuboi, Y.; Wakana, C.; Mawatari, S.; Kono, S. Efficacy and blood plasmalogen changes by oral administration of plasmalogen in patients with mild Alzheimer’s disease and mild cognitive impairment: A multicenter, randomized, double-blind, placebo-controlled trial. EBioMedicine 2017, 17, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Holtzman, D.M.; McKeel, D.W., Jr. Plasmalogen deficiency in early Alzheimer’s disease subjects and in animal models: Molecular characterization using electrospray ionization mass spectrometry. J. Neurochem. 2001, 77, 1168–1180. [Google Scholar] [CrossRef] [PubMed]
- Chan, R.B.; Oliveira, T.G.; Cortes, E.P.; Honig, L.S.; Duff, K.E.; Small, S.A.; Wenk, M.R.; Shui, G.; Di Paolo, G. Comparative lipidomic analysis of mouse and human brain with Alzheimer disease. J. Biol. Chem. 2012, 287, 2678–2688. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Precursor Ion m/z | Mass Error a ppm | Retention Time (min) | Precursor Ion Species | Product Ions m/z | P-Value | FDR (q-Value) | Identification Level |
|---|---|---|---|---|---|---|---|---|
| Nervonic Acid | 365.3419 | 0.13 | 11.18 | [M − H]− | - | 0.0006 | 0.005 | confirmed (level 1) |
| Eicosenoic Acid | 309.2799 | 1.93 | 8.67 | [M − H]− | - | 0.0027 | 0.01 | confirmed (level 1) |
| Linoleic Acid | 279.2325 | 0.36 | 6.13 | [M − H]− | - | 0.0185 | 0.04 | confirmed (level 1) |
| Docosenoic Acid | 337.3109 | 0.89 | 10.00 | [M − H]− | - | 0.0003 | 0.005 | confirmed (level 1) |
| Eicosatrienoic Acid Isomer | 305.2485 | 1.64 | 6.68 | [M − H]− | 303, 261 | 0.0254 | 0.04 | putative (level 2) |
| 22-tricosenoic Acid Isomer | 351.3267 | 1.14 | 10.60 | [M − H]− | - | 0.0006 | 0.005 | putative (level 3) |
| Tetracosatetraeonic Acid | 359.2951 | 0.28 | 8.49 | [M − H]− | - | 0.0044 | 0.01 | putative (level 3) |
| Lysophosphatidic Acid Isomer | 409.2358 | 0.73 | 5.78 | [M − H]− | - | 0.0285 | 0.04 | putative (level 3) |
| PG (22:0/18:1) | 669.5595 | 1.34 | 15.33 | [M-C6H10O5-H]− | - | 0.0053 | 0.01 | putative (level 3) |
| Unknown-1 | 720.6507 | - | 15.00 | [M − H]− | - | 0.0093 | 0.02 | unknown |
| Unknown-2 | 425.2095 | - | 5.78 | [M − H]− | - | 0.0129 | 0.03 | unknown |
| Unknown-3 | 477.2027 | - | 7.23 | [M − H]− | - | 0.0291 | 0.04 | unknown |
| Compound | Precursor Ion m/z | Mass Error a ppm | Retention Time (min) | Precursor Ion Species | Product Ions m/z | P-Value | FDR (q-Value) | Identification Level |
|---|---|---|---|---|---|---|---|---|
| Eicosapentanoic Acid | 301.2171 | 1.33 | 5.10 | [M − H]− | - | 1.52 × 10−6 | 0.0002 | confirmed (level 1) |
| Docosapentanoic Acid | 329.2484 | 1.21 | 6.33 | [M − H]− | - | 0.0003 | 0.01 | confirmed (level 1) |
| Eicosadienoic Acid | 307.2638 | 0.32 | 7.60 | [M − H]− | - | 0.0007 | 0.03 | confirmed (level 1) |
| PG 20:4/22:6 | 841.5026 | 0.83 | 10.00 | [M − H]− | 327; 303 | 0.0007 | 0.03 | putative (level 2) |
| PG 18:2/18:2 | 769.5010 | −1.16 | 10.32 | [M − H]− | 279 | 3.89 × 10−5 | 0.003 | putative (level 2) |
| PI 18:0/20:5 | 883.5329 | −0.79 | 11.39 | [M − H]− | 581; 419; 301; 283; 241 | 1.62 × 10−6 | 0.0002 | putative (level 2) |
| PE 22:6/22:5 | 836.5238 | 0.96 | 11.41 | [M − H]− | 329; 283 | 0.0015 | 0.04 | putative (level 2) |
| PC 16:0/20:5 | 838.5609 | 1.31 | 11.53 | [M+CH3COO]− | 764; 301; 255 | 3.98 × 10−5 | 0.003 | putative (level 2) |
| PC 16:0/18:3 | 814.5164 | −4.17 | 11.53 | [M+CH3COO]− | 740; 277; 255 | 0.0005 | 0.02 | putative (level 2) |
| PC 16:0/18:2 | 792.5384 | 9.33 | 12.20 | [M + Cl]− | 742; 279; 255 | 1.18 × 10−8 | 2.95 × 10−6 | putative (level 2) |
| PE 16:1/20:4 | 736.4922 | 0.68 | 11.70 | [M − H]− | 303; 253 | 3.46 × 10−12 | 1.73 × 10−9 | putative (level 2) |
| PE (P-16:0/20:5) | 720.4970 | 0.28 | 12.14 | [M − H]− | 436; 418; 301 | 5.42 × 10−5 | 0.003 | putative (level 2) |
| PE 16:0/22:5 | 764.5229 | −0.13 | 12.28 | [M − H]− | 329; 255 | 0.0004 | 0.02 | putative (level 2) |
| PE (P-18:0/22:6) | 774.5448 | 1.42 | 12.78 | [M − H]− | 464; 446; 327 | 0.0015 | 0.04 | putative (level 2) |
| PE 18:1/20:3 | 766.5376 | −1.30 | 12.85 | [M − H]− | 305; 281 | 0.0013 | 0.04 | putative (level 2) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bennouna, D.; Solano, M.; Orchard, T.S.; DeVries, A.C.; Lustberg, M.; Kopec, R.E. The Effects of Doxorubicin-based Chemotherapy and Omega-3 Supplementation on Mouse Brain Lipids. Metabolites 2019, 9, 208. https://doi.org/10.3390/metabo9100208
Bennouna D, Solano M, Orchard TS, DeVries AC, Lustberg M, Kopec RE. The Effects of Doxorubicin-based Chemotherapy and Omega-3 Supplementation on Mouse Brain Lipids. Metabolites. 2019; 9(10):208. https://doi.org/10.3390/metabo9100208
Chicago/Turabian StyleBennouna, Djawed, Melissa Solano, Tonya S. Orchard, A. Courtney DeVries, Maryam Lustberg, and Rachel E. Kopec. 2019. "The Effects of Doxorubicin-based Chemotherapy and Omega-3 Supplementation on Mouse Brain Lipids" Metabolites 9, no. 10: 208. https://doi.org/10.3390/metabo9100208
APA StyleBennouna, D., Solano, M., Orchard, T. S., DeVries, A. C., Lustberg, M., & Kopec, R. E. (2019). The Effects of Doxorubicin-based Chemotherapy and Omega-3 Supplementation on Mouse Brain Lipids. Metabolites, 9(10), 208. https://doi.org/10.3390/metabo9100208