Metabolomics of Two Pecan Varieties Provides Insights into Scab Resistance

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. General
2.2. UHPLC-QTOF-MS
2.3. Data Processing and Statistical Analysis
3. Results and Discussions
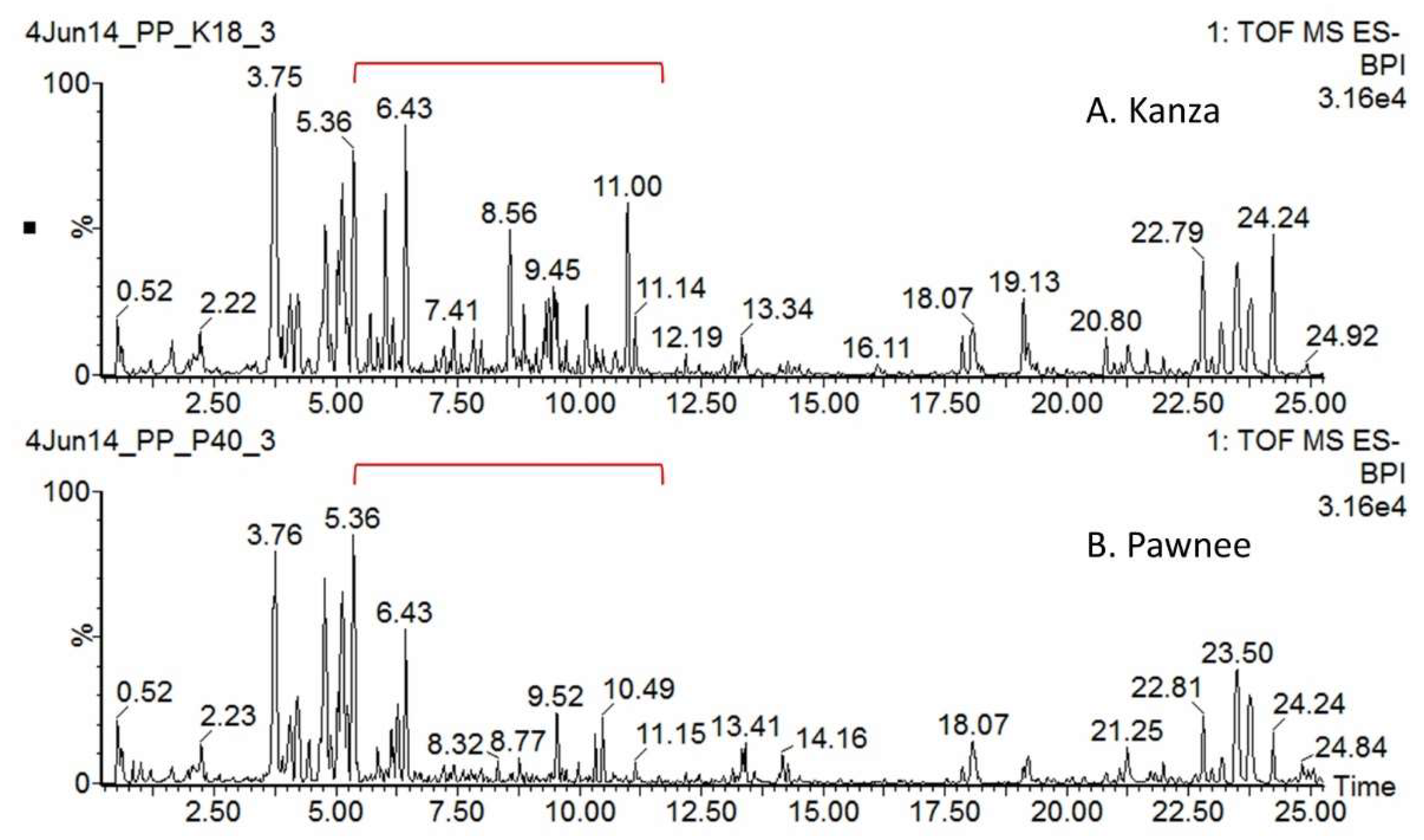
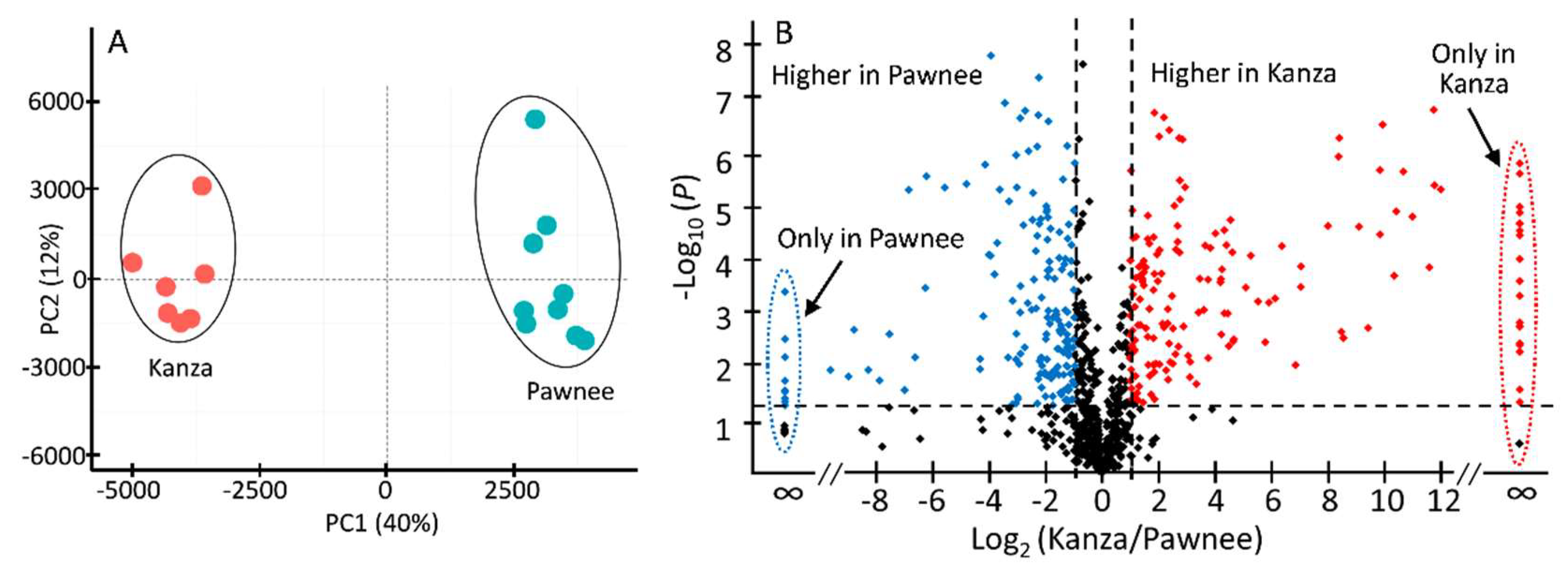
3.1. Kanza and Pawnee Had Distinct Metabolic Profiles
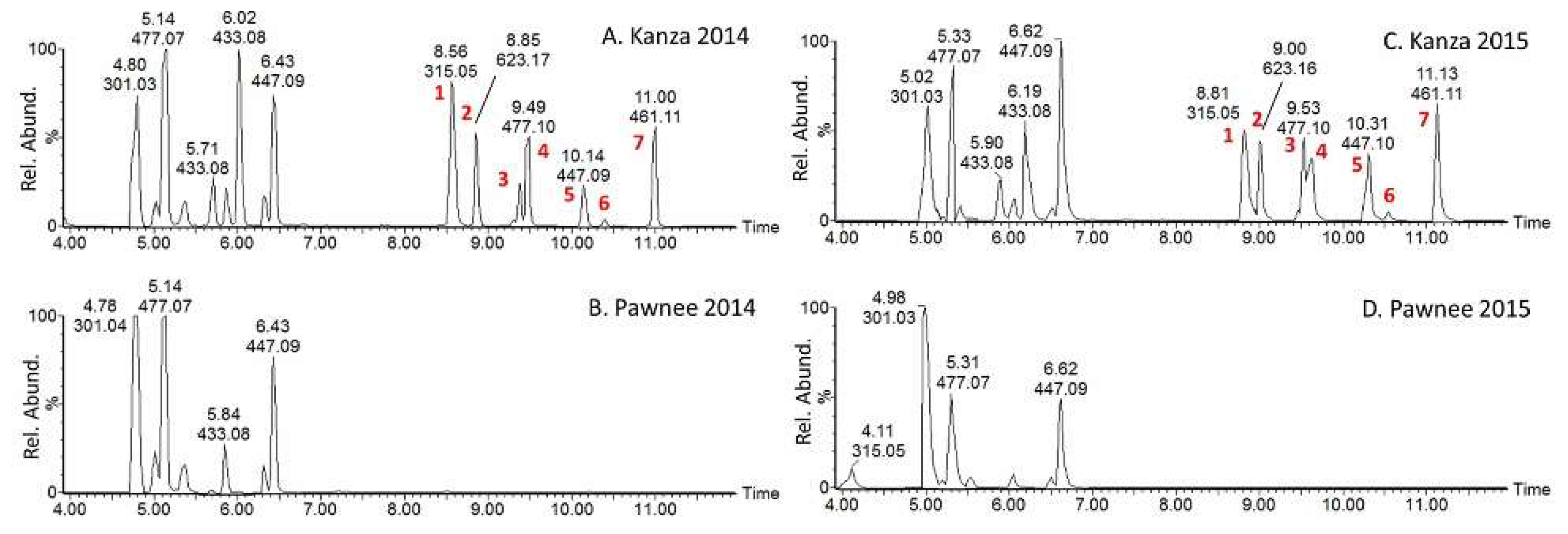
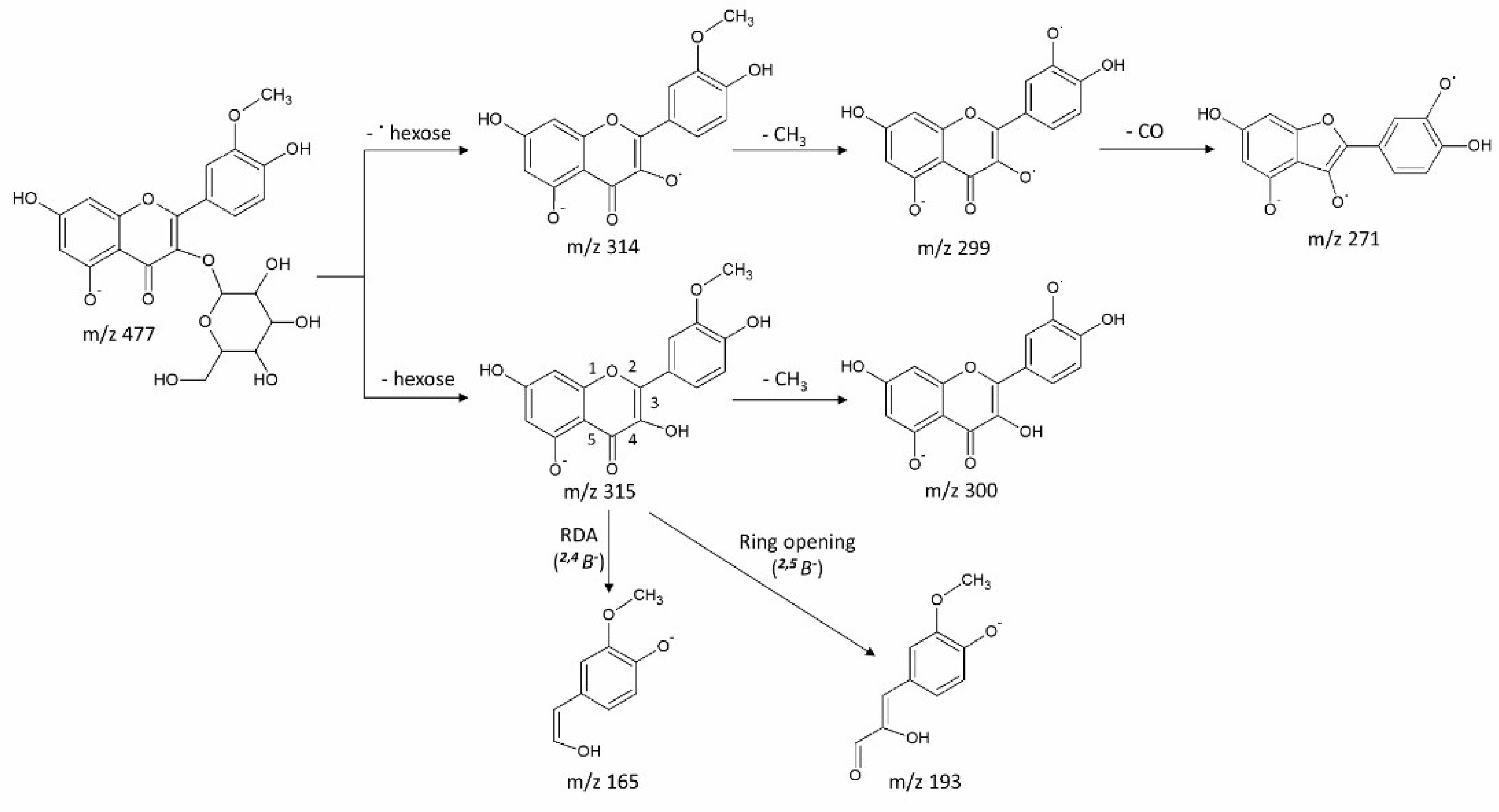
3.2. Putative Metabolite Identification via MS/MS
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- USDA-NASS. Noncitrus Fruits and Nuts 2017 Summary; USDA National Agricultural Statistics Service Survey; USDA-NASS: Washington, DC, USA, 2018.
- Blomhoff, R.; Carlsen, M.H.; Andersen, L.F.; Jacobs, D.R., Jr. Health benefits of nuts: Potential role of antioxidants. Br. J. Nutr. 2006, 96, S52–S60. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.B.; Stampfer, M.J. Nut consumption and risk of coronary heart disease: A review of epidemiologic evidence. Curr. Atheroscler. Rep. 1999, 1, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Hudthagosol, C.; Haddad, E.H.; McCarthy, K.; Wang, P.; Oda, K.; Sabate, J. Pecans acutely increase plasma postprandial antioxidant capacity and catechins and decrease LDL oxidation in humans. J. Nutr. 2011, 141, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Kris-Etherton, P.M.; Zhao, G.; Binkoski, A.E.; Coval, S.M.; Etherton, T.D. The effects of nuts on coronary heart disease risk. Nutr. Rev. 2001, 59, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Sabate, J.; Ros, E.; Salas-Salvado, J. Nuts: Nutrition and health outcomes. Preface. Br. J. Nutr. 2006, 96, S1–S2. [Google Scholar] [CrossRef] [PubMed]
- Ellsworth, J.L.; Kushi, L.H.; Folsom, A.R. Frequent nut intake and risk of death from coronary heart disease and all causes in postmenopausal women: The Iowa Women’s Health Study. Nutr. Metab. Cardiovasc. Dis. 2001, 11, 372–377. [Google Scholar] [PubMed]
- Bao, Y.; Han, J.; Hu, F.B.; Giovannucci, E.L.; Stampfer, M.J.; Willett, W.C.; Fuchs, C.S. Association of nut consumption with total and cause-specific mortality. N. Engl. J. Med. 2013, 369, 2001–2011. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Beecher, G.R.; Holden, J.M.; Haytowitz, D.B.; Gebhardt, S.E.; Prior, R.L. Lipophilic and hydrophilic antioxidant capacities of common foods in the United States. J. Agric. Food Chem. 2004, 52, 4026–4037. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Wong, J.; Ames, B.N. Gamma-tocopherol induces apoptosis in androgen-responsive LNCaP prostate cancer cells via caspase-dependent and independent mechanisms. Ann. N. Y. Acad. Sci. 2004, 1031, 399–400. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Wong, J.; Fyrst, H.; Saba, J.D.; Ames, B.N. gamma-Tocopherol or combinations of vitamin E forms induce cell death in human prostate cancer cells by interrupting sphingolipid synthesis. Proc. Natl. Acad. Sci. USA 2004, 101, 17825–17830. [Google Scholar] [CrossRef] [PubMed]
- Wahle, K.W.; Brown, I.; Rotondo, D.; Heys, S.D. Plant phenolics in the prevention and treatment of cancer. Adv. Exp. Med. Biol. 2010, 698, 36–51. [Google Scholar] [PubMed]
- Weaver, T. Routing Pecan Scab—Protecting a Popular Nut. Agric. Res. 1998, 46, 8. [Google Scholar]
- Littrell, R.H.; Bertrand, P.F. Management of Pecan Fruit and Foliar Diseases with Fungicides. Plant Dis. 1981, 65, 769–774. [Google Scholar] [CrossRef]
- Stevenson, K.; Bertrand, P.; Brennamin, T. Evidence for reduced sensitivity to propiconazole in the pecan scab fungus in Georgia. Phytopathology 2004, 94, S99. [Google Scholar]
- Thompson, T.E.; Grauke, L.J.; Reid, W.; Smith, M.W.; Winter, S.R. ‘Kanza’ pecan. Hortscience 1997, 32, 139–140. [Google Scholar]
- Lei, Z.; Jing, L.; Qiu, F.; Zhang, H.; Huhman, D.; Zhou, Z.; Sumner, L.W. Construction of an Ultrahigh Pressure Liquid Chromatography-Tandem Mass Spectral Library of Plant Natural Products and Comparative Spectral Analyses. Anal. Chem. 2015, 87, 7373–7381. [Google Scholar] [CrossRef] [PubMed]
- Horai, H.; Arita, M.; Kanaya, S.; Nihei, Y.; Ikeda, T.; Suwa, K.; Ojima, Y.; Tanaka, K.; Tanaka, S.; Aoshima, K.; et al. MassBank: A public repository for sharing mass spectral data for life sciences. J. Mass Spectrom. 2010, 45, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Wishart, D.S. Using MetaboAnalyst 3.0 for Comprehensive Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2016, 55. [Google Scholar] [CrossRef] [PubMed]
- VanEtten, H.D.; Mansfield, J.W.; Bailey, J.A.; Farmer, E.E. Two Classes of Plant Antibiotics: Phytoalexins versus “Phytoanticipins”. Plant Cell 1994, 6, 1191–1192. [Google Scholar] [CrossRef] [PubMed]
- Ekanayaka, E.A.; Celiz, M.D.; Jones, A.D. Relative mass defect filtering of mass spectra: A path to discovery of plant specialized metabolites. Plant Physiol. 2015, 167, 1221–1232. [Google Scholar] [CrossRef] [PubMed]
- Sumner, L.W.; Lei, Z.; Nikolau, B.J.; Saito, K. Modern plant metabolomics: Advanced natural product gene discoveries, improved technologies, and future prospects. Nat. Prod. Rep. 2015, 32, 212–229. [Google Scholar] [CrossRef] [PubMed]
- Kanwal, Q.; Hussain, I.; Latif Siddiqui, H.; Javaid, A. Antifungal activity of flavonoids isolated from mango (Mangifera indica L.) leaves. Nat. Prod. Res. 2010, 24, 1907–1914. [Google Scholar] [CrossRef] [PubMed]
- Slatnar, A.; Mikulic Petkovsek, M.; Halbwirth, H.; Stampar, F.; Stich, K.; Veberic, R. Polyphenol metabolism of developing apple skin of a scab resistant and a susceptible apple cultivar. Trees 2012, 26, 109–119. [Google Scholar] [CrossRef]
- Ullah, C.; Unsicker, S.B.; Fellenberg, C.; Constabel, C.P.; Schmidt, A.; Gershenzon, J.; Hammerbacher, A. Flavan-3-ols Are an Effective Chemical Defense against Rust Infection. Plant Physiol. 2017, 175, 1560–1578. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | tR min | [M-H]− | Formula | Error ppm | MS/MS, m/z (intensity %) | Identification |
|---|---|---|---|---|---|---|
| 1 | 8.56 | 315.0505 | C16H12O7 | 0.06 | 315 (70), 300 (41), 299 (14), 271 (40), 243 (12), 165 (100), 121 (50) | rhamnetin (7-Methoxyquercetin) |
| 2 | 8.85 | 623.1634 | C28H32O16 | 3.5 | 623 (100), 314 (85), 299 (20), 191 (20) | Isorhamnetin-3-galactoside-6′′-rhamnoside |
| 3 | 9.38 | 477.1039 | C22H22O12 | 1.3 | 477 (58), 315 (70), 314 (100), 300 (12), 299 (32), 271 (10), 193 (9), 165 (12) | Isorhamnetin-3-O-glucoside |
| 4 | 9.49 | 477.1034 | C22H22O12 | 0.2 | 477 (58), 315 (70), 314 (100), 300 (12), 299 (32), 271 (10), 193 (9), 165 (12) | Isorhamnetin-3-O-glucoside |
| 5 | 10.14 | 447.0925 | C21H20O11 | 0.4 | 447 (100), 445 (80), 432 (30), 316 (40), 315 (40), 161 (40), 101(60) | Rhamnetin-O-pentoside |
| 6 | 10.39 | 447.0963 | C21H20O11 | 8 | 447 (35), 315 (100), 314 (80), 299 (60), 271 (20), 165 (15) | Isorhamnetin-3-O-glucoside |
| 7 | 11.02 | 461.1071 | C22H22O11 | 2.6 | 461 (71), 315 (100), 314 (80), 299 (70), 271 (30), 165 (28) | Methylated isorhamnetin-3-O-rhamnoside |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, Z.; Kranawetter, C.; Sumner, B.W.; Huhman, D.; Wherritt, D.J.; Thomas, A.L.; Rohla, C.; Sumner, L.W. Metabolomics of Two Pecan Varieties Provides Insights into Scab Resistance. Metabolites 2018, 8, 56. https://doi.org/10.3390/metabo8040056
Lei Z, Kranawetter C, Sumner BW, Huhman D, Wherritt DJ, Thomas AL, Rohla C, Sumner LW. Metabolomics of Two Pecan Varieties Provides Insights into Scab Resistance. Metabolites. 2018; 8(4):56. https://doi.org/10.3390/metabo8040056
Chicago/Turabian StyleLei, Zhentian, Clayton Kranawetter, Barbara W. Sumner, David Huhman, Daniel J. Wherritt, Andrew L. Thomas, Charles Rohla, and Lloyd W. Sumner. 2018. "Metabolomics of Two Pecan Varieties Provides Insights into Scab Resistance" Metabolites 8, no. 4: 56. https://doi.org/10.3390/metabo8040056
APA StyleLei, Z., Kranawetter, C., Sumner, B. W., Huhman, D., Wherritt, D. J., Thomas, A. L., Rohla, C., & Sumner, L. W. (2018). Metabolomics of Two Pecan Varieties Provides Insights into Scab Resistance. Metabolites, 8(4), 56. https://doi.org/10.3390/metabo8040056