Specificities of Human Hepatocellular Carcinoma Developed on Non-Alcoholic Fatty Liver Disease in Absence of Cirrhosis Revealed by Tissue Extracts 1H-NMR Spectroscopy


 ,
,
Abstract
:1. Introduction
2. Results
2.1. Patients
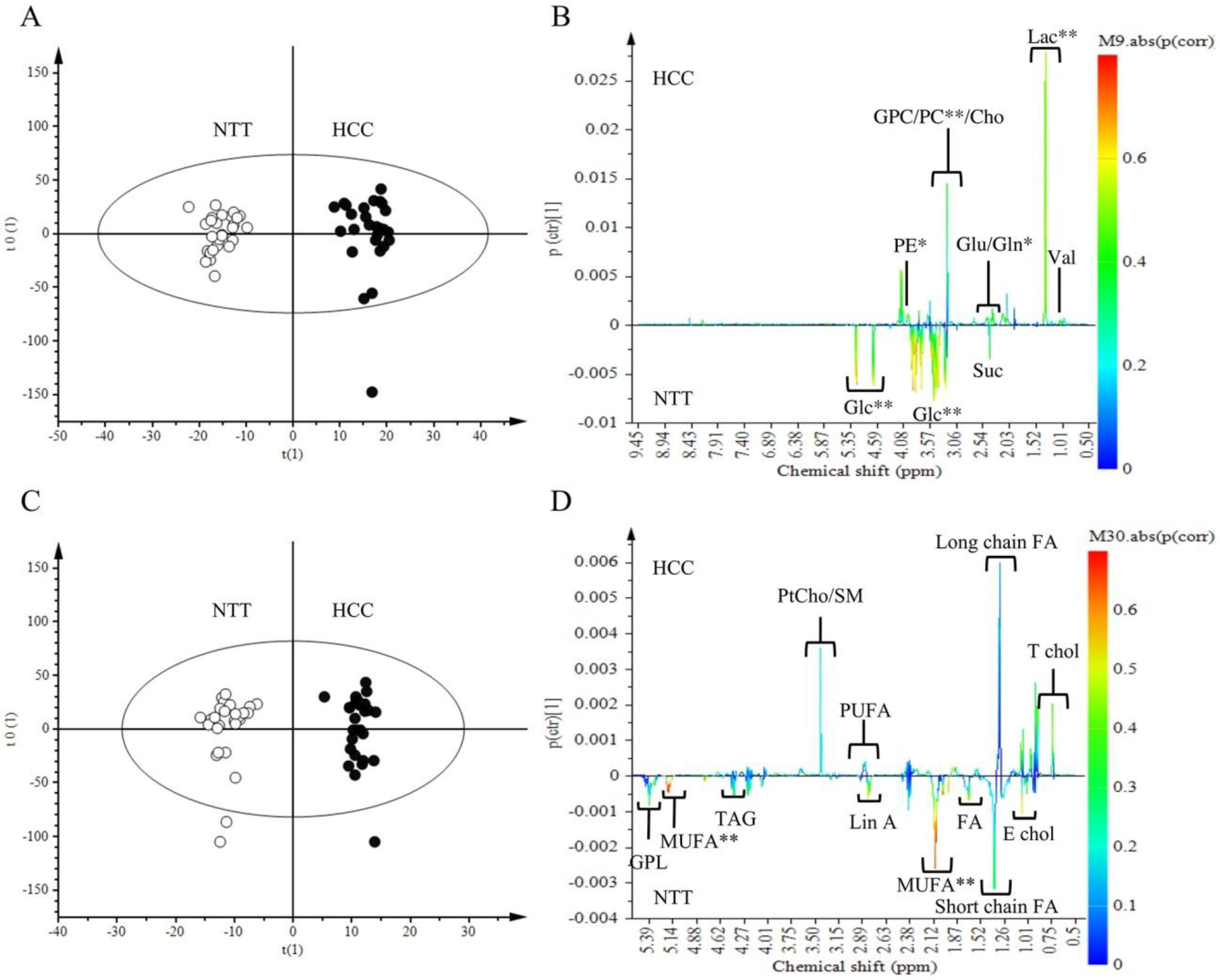
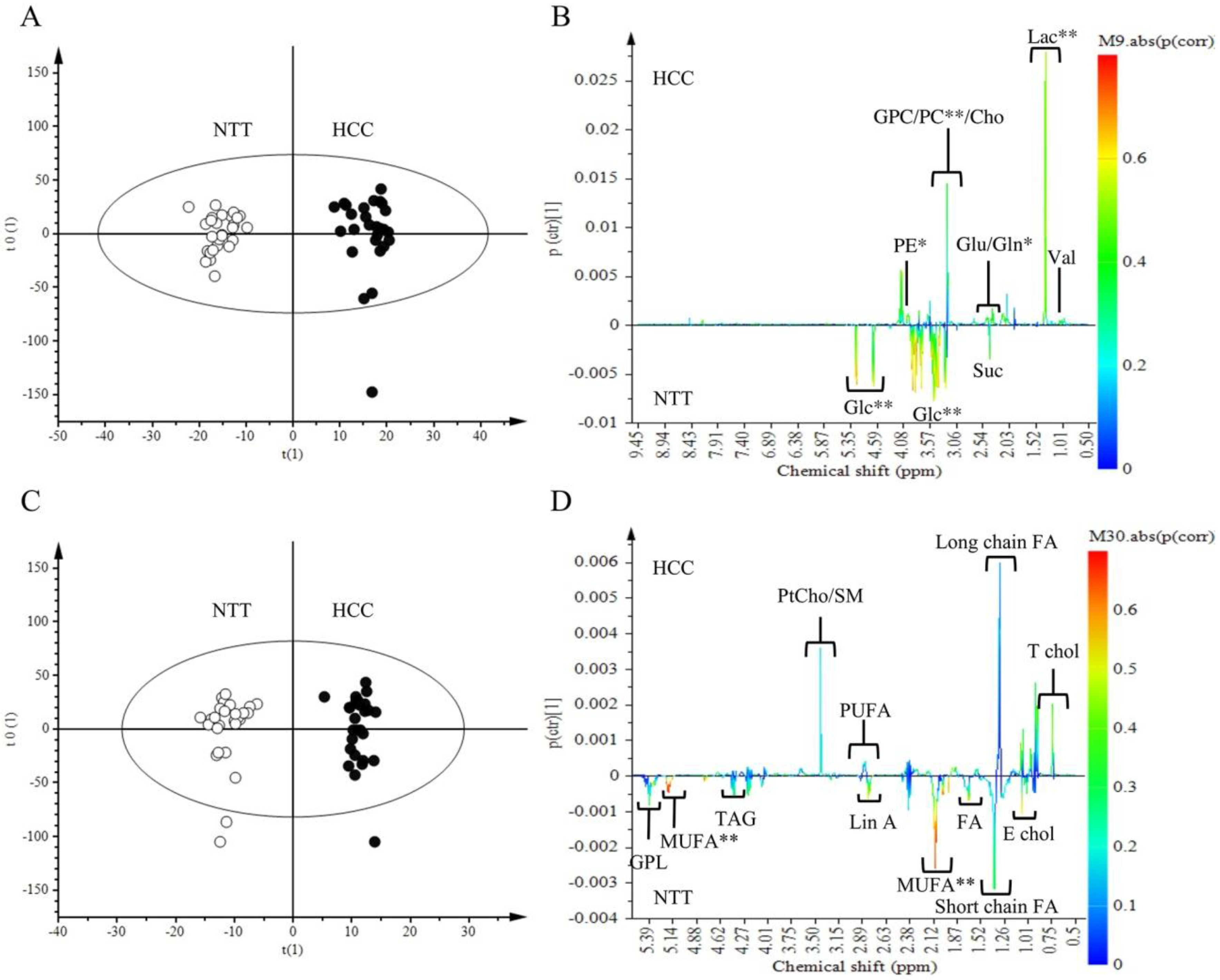
2.2. Metabolomics Comparison of HCC to NTT
2.3. Metabolites Quantification
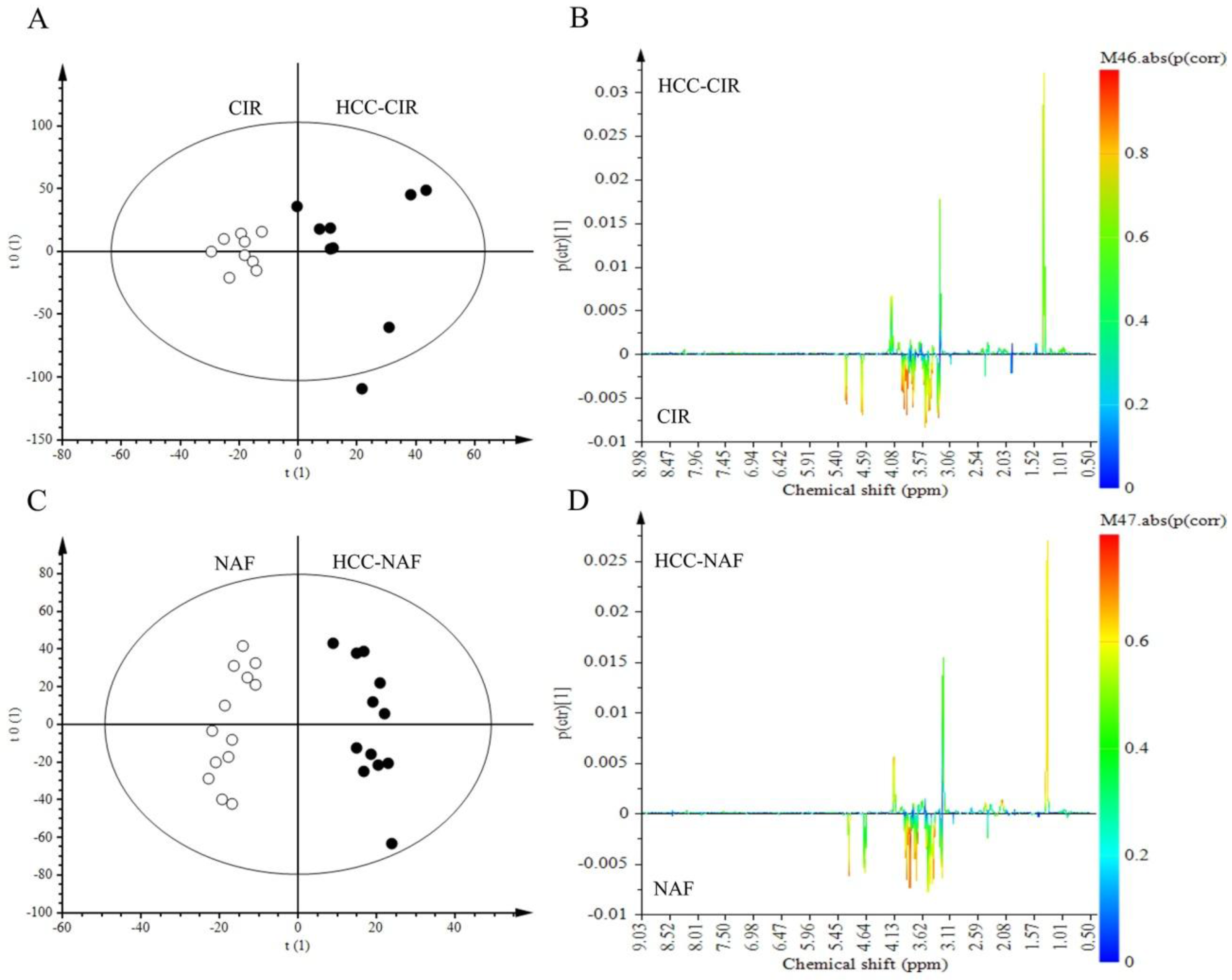
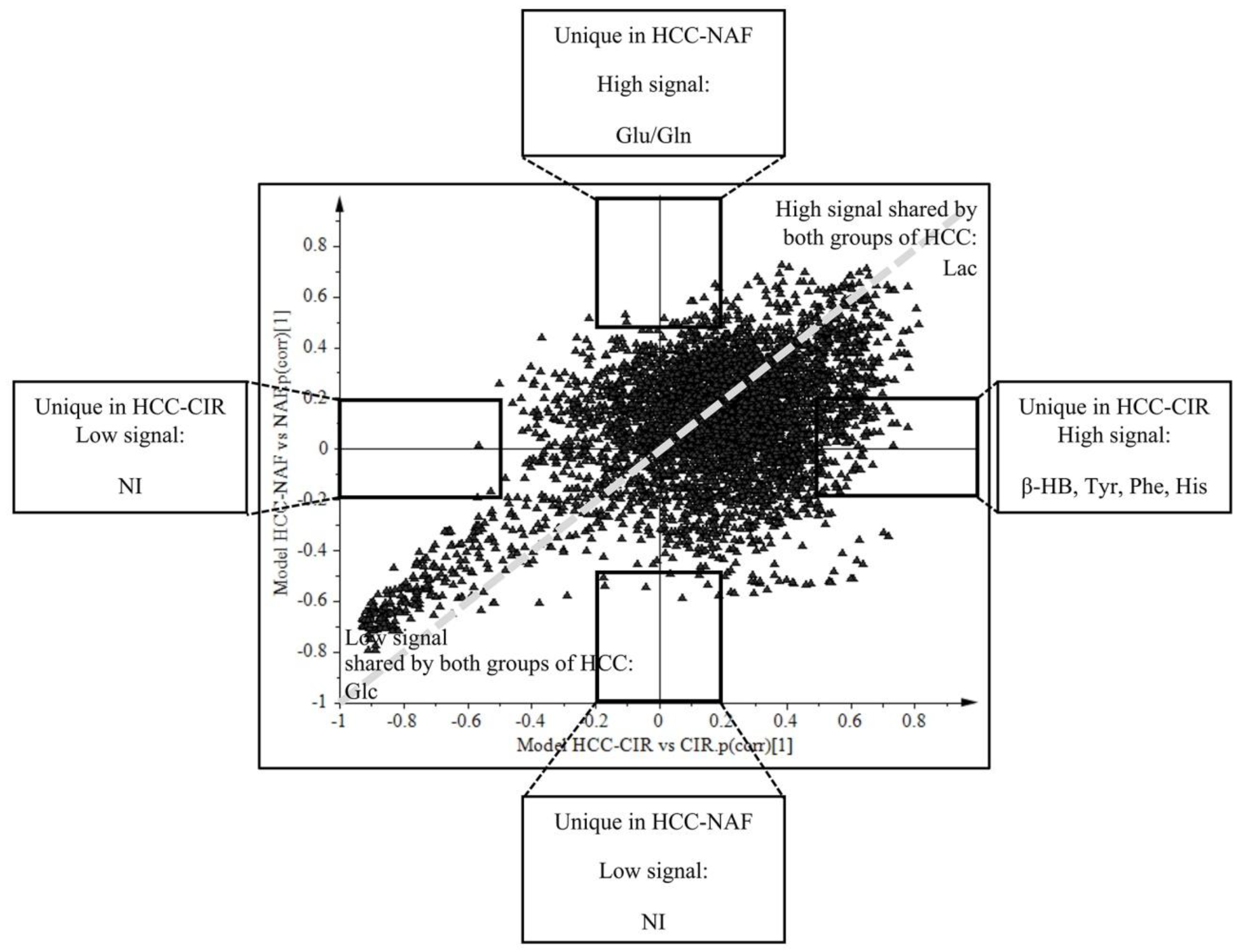
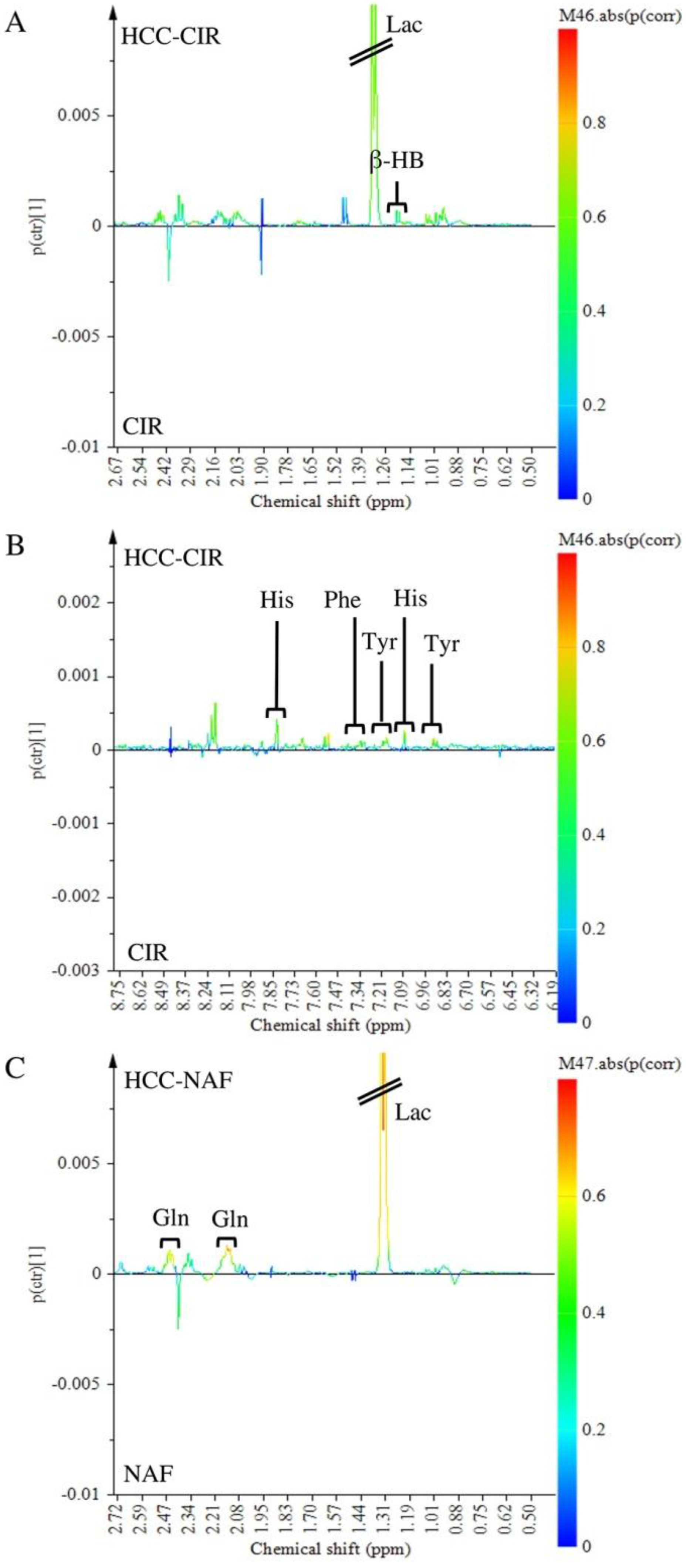
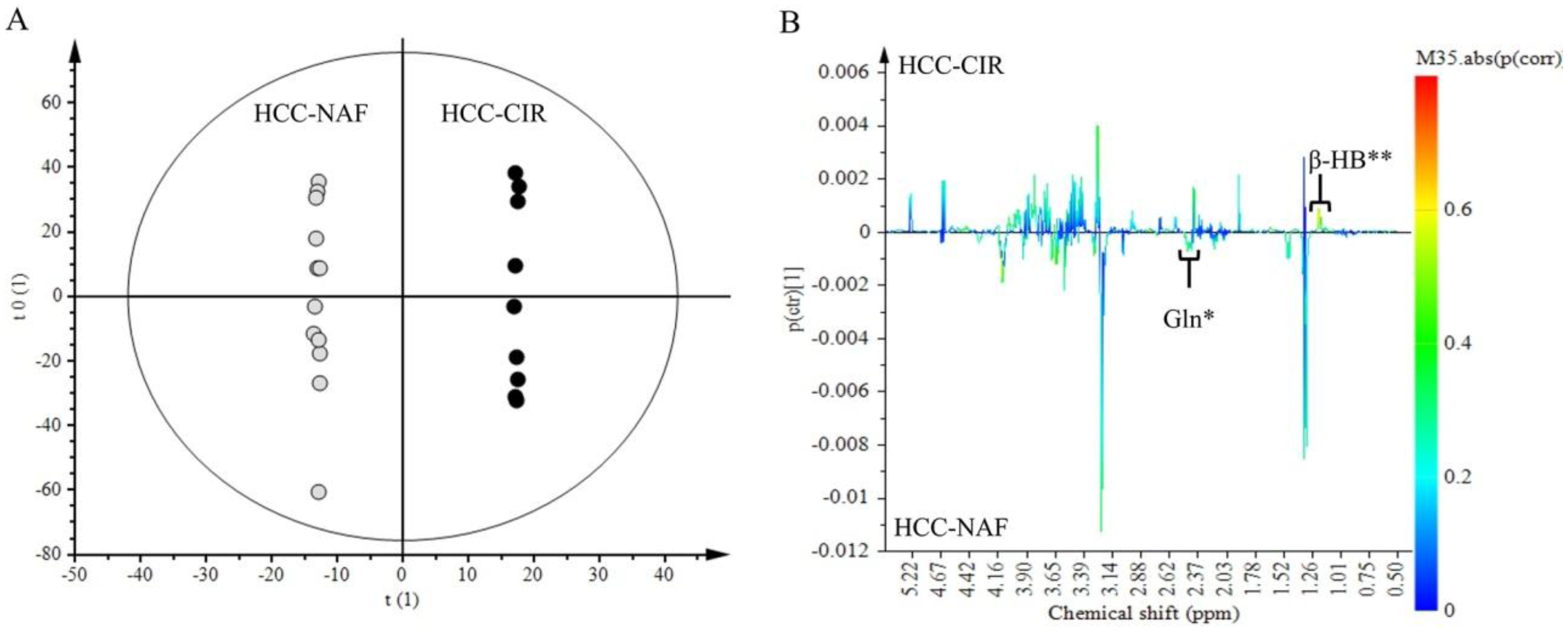
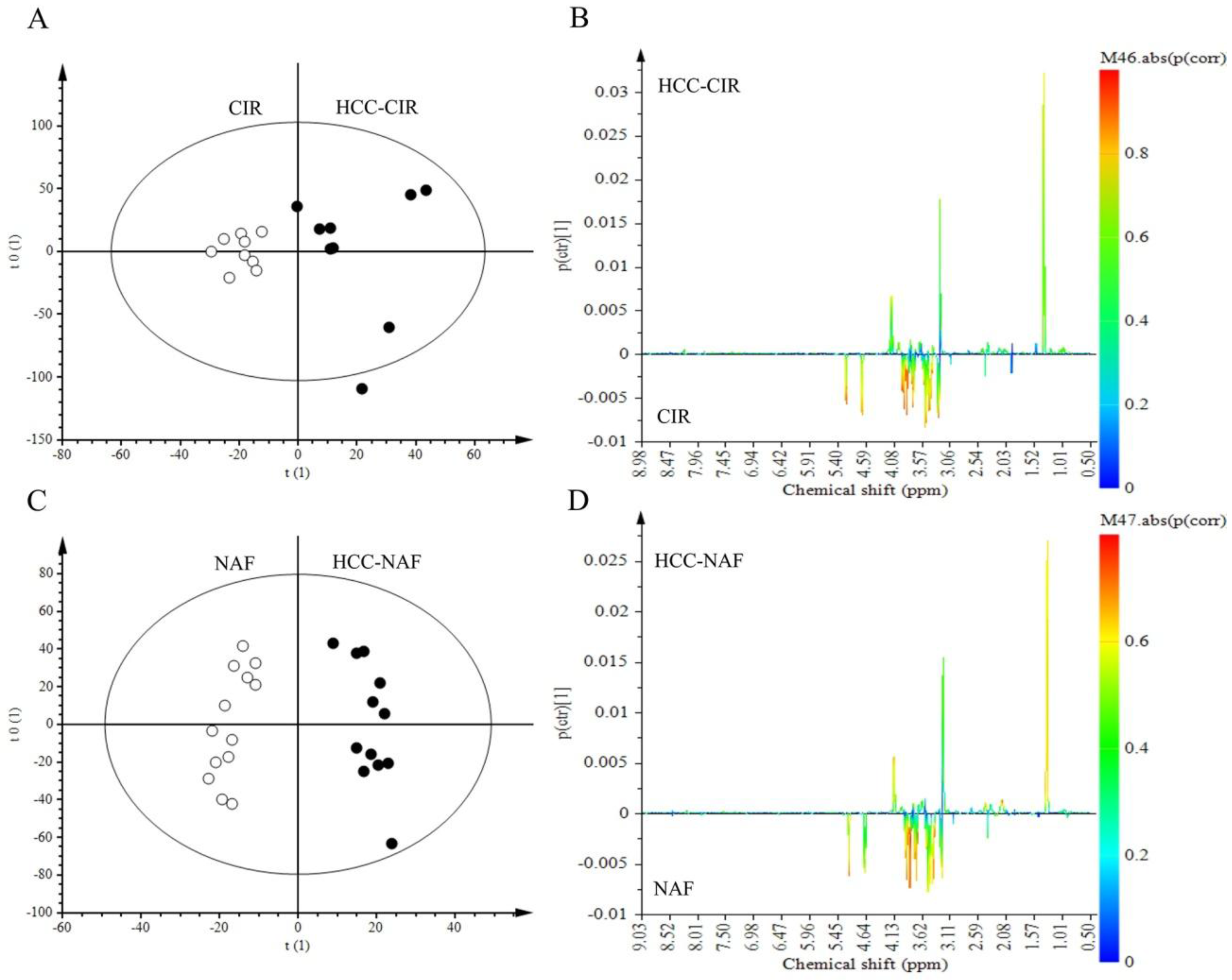
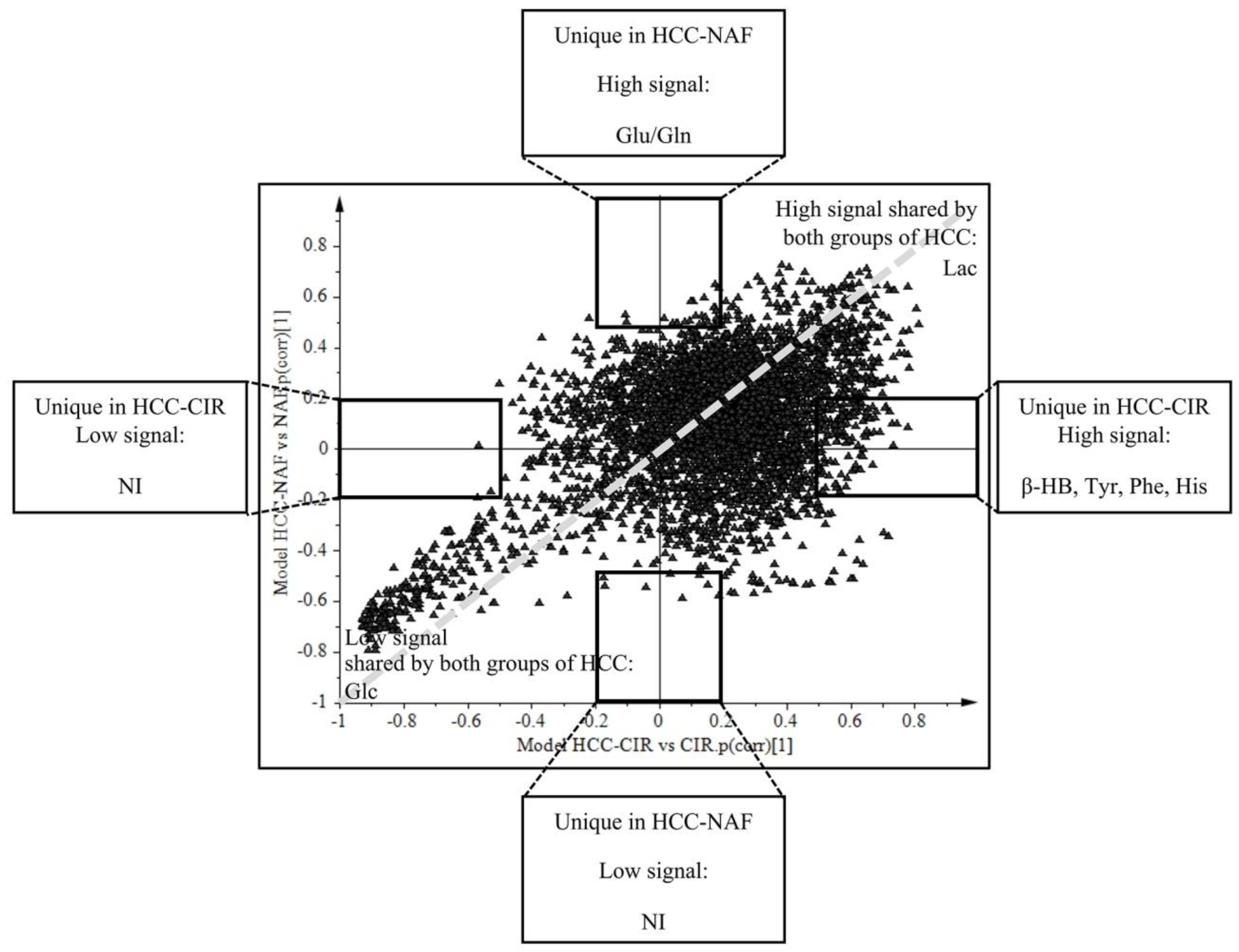
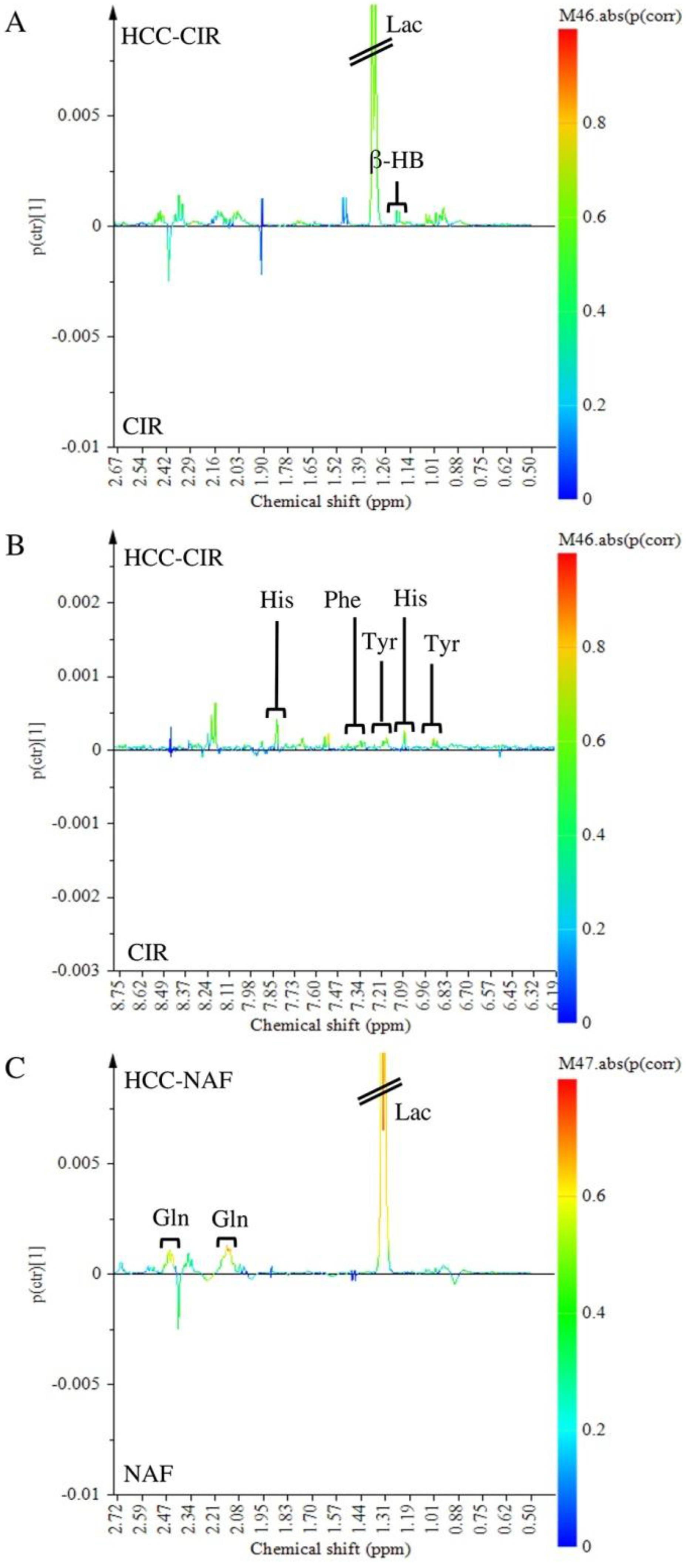
2.4. Comparison of HCC-NAFLD to HCC-Cirrhosis
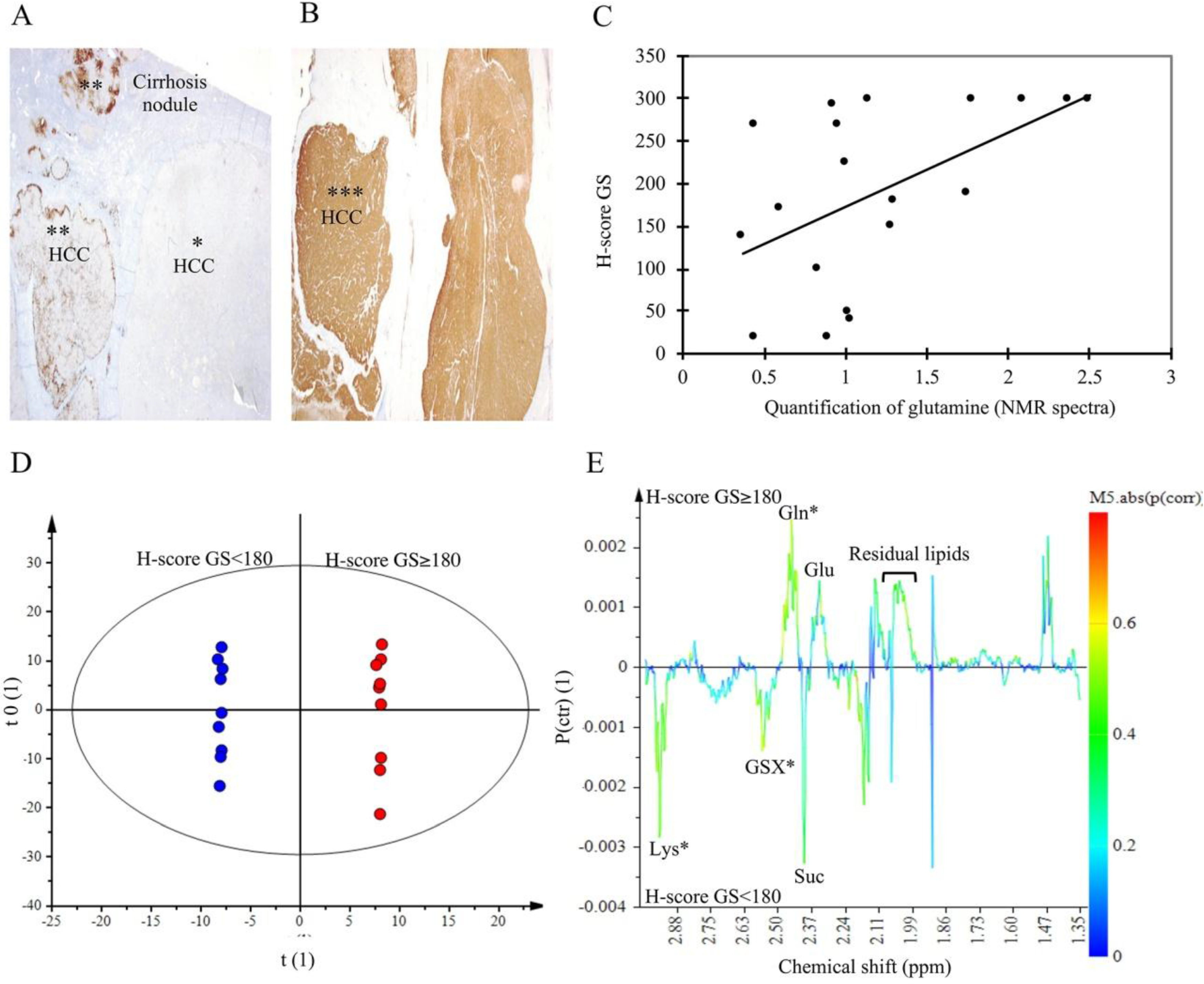
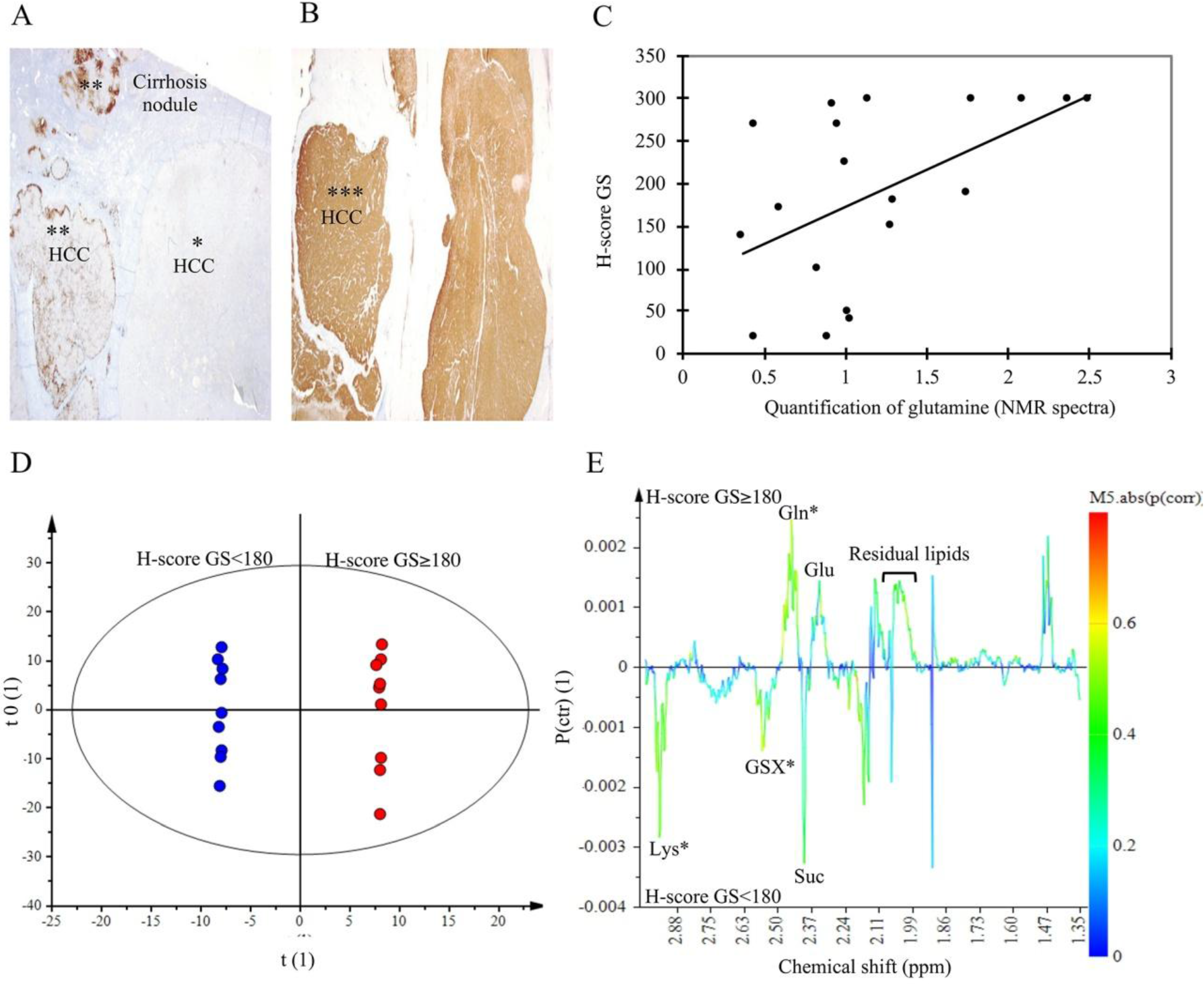
2.5. Glutamine Synthetase Immunostaining
3. Discussion
3.1. Advantage and Limitation of Using Tissue Extract Instead of Intact Tissue in this NMR Metabolomics Study
3.2. HCC Metabolic Profile Involves High Glycolysis and Impaired Phospholipids Metabolism
3.3. Metabolomics Shows Differences between HCC-NAFLD and HCC-Cirrhosis
4. Patients and Methods
4.1. Patients and Collection of Specimens
4.2. Histology
4.3. Sample Preparation for NMR-Spectroscopy
4.4. 1H-NMR Spectroscopy
4.5. Data Processing
4.6. Metabolites Quantification
4.7. Multivariate Data Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| Ace | Acetate |
| AFP | Alpha-Foeto-Protein |
| Ara A | Arachidonic Acid |
| Asc A | Ascorbic acid |
| BCAAs | Branched-Chain Amino Acids |
| BMI | Body Mass Index |
| Beta-Glc | β-glucose |
| Cho | Choline |
| Creat | Creatine |
| E Chol | Esterified Cholesterol |
| FID | Free Induction Decay |
| For | Formate |
| Fum | Fumarate |
| Gln | Glutamine |
| Glu | Glutamate |
| Gly | Glycine |
| GPC | Glycerophosphocholine |
| GS | Glutamine Synthetase |
| HBV | Hepatitis B Virus |
| HCC | Hepatocellular Carcinoma |
| HCV | Hepatitis C Virus |
| HDL | High Density Lipoprotein |
| His | Histidine |
| HMDB | Human Metabolome Database |
| H-NMR | Hydrogen-Nuclear Magnetic Resonance |
| HR-MAS MR | High Resolution Magic Angle Spinning Magnetic Resonance |
| Lac | Lactate |
| Leu | Leucine |
| Lin A | Linoleic Acid |
| MUFA | Monounsaturated Fatty Acid |
| NAFLD | Non-Alcoholic Fatty Liver Disease |
| NASH | Non-Alcoholic Steato-Hepatitis |
| NTT | Non Tumoral Tissue |
| OPLS-DA | Orthogonal Partial Least Square-Discriminant Analysis |
| PC | Phosphocholine |
| PCA | Principal Components Analysis |
| PE | Phosphoethanolamine |
| Phe | Phenylalanine |
| PLS | Partial Least Square |
| ppm | part per million |
| Ptcho | phosphatidylcholine |
| PUFA | Polyunsaturated Fatty Acid |
| Sar | Sarcosine |
| SM | Sphingomyelin |
| Suc | Succinate |
| SUS-Plot | Shared and Unique Structures-Plot |
| T Chol | Total Cholesterol |
| T FA | Total Fatty Acid |
| TAG | Triacylglycerol |
| TCA cycle | Citric Acid Cycle |
| TNF | tumor necrosis factor |
| TT | Tumor Tissue |
| Tyr | Tyrosine |
| Val | Valine |
| VIP | Variable Influence on the Projection |
| WHO | World Health Organisation |
| β-HB | β-hydroxybutyrate |
References
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, A.; Poklepovic, A.; Moyneur, E.; Barghout, V. Population-based risk factors and resource utilization for HCC: US perspective. Curr. Med. Res. Opin. 2010, 26, 2183–2191. [Google Scholar] [CrossRef] [PubMed]
- Welzel, T.M.; Graubard, B.I.; Zeuzem, S.; El-Serag, H.B.; Davila, J.A.; McGlynn, K.A. Metabolic syndrome increases the risk of primary liver cancer in the United States: A study in the SEER-Medicare database. Hepatology 2011, 54, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.H.; Younossi, Z.M. Nonalcoholic fatty liver disease: A manifestation of the metabolic syndrome. Cleve. Clin. J. Med. 2008, 75, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Paradis, V.; Zalinski, S.; Chelbi, E.; Guedj, N.; Degos, F.; Vilgrain, V.; Bedossa, P.; Belghiti, J. Hepatocellular carcinomas in patients with metabolic syndrome often develop without significant liver fibrosis: A pathological analysis. Hepatology 2009, 49, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Yasui, K.; Hashimoto, E.; Komorizono, Y.; Koike, K.; Arii, S.; Imai, Y.; Shima, T.; Kanbara, Y.; Saibara, T.; Mori, T.; et al. Characteristics of patients with nonalcoholic steatohepatitis who develop hepatocellular carcinoma. Clin. Gastroenterol. Hepatol. 2011, 9, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Piscaglia, F.; Svegliati-Baroni, G.; Barchetti, A.; Pecorelli, A.; Marinelli, S.; Tiribelli, C.; Bellentani, S. HCC-NAFLD Italian study group clinical patterns of hepatocellular carcinoma in nonalcoholic fatty liver disease: A multicenter prospective study. Hepatology 2016, 63, 827–838. [Google Scholar] [CrossRef] [PubMed]
- Baffy, G.; Brunt, E.M.; Caldwell, S.H. Hepatocellular carcinoma in non-alcoholic fatty liver disease: An emerging menace. J. Hepatol. 2012, 56, 1384–1391. [Google Scholar] [CrossRef] [PubMed]
- Stickel, F.; Hellerbrand, C. Non-alcoholic fatty liver disease as a risk factor for hepatocellular carcinoma: Mechanisms and implications. Gut 2010, 59, 1303–1307. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Lu, Q.; Liu, X.; Cong, H.; Zhao, L.; Wang, H.; Lin, D. Application of 1H-NMR-based metabonomics in the study of metabolic profiling of human hepatocellular carcinoma and liver cirrhosis. Cancer Sci. 2009, 100, 782–785. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Ding, L.; Yin, P.; Lu, X.; Wang, X.; Niu, J.; Gao, P.; Xu, G. Serum metabolic profiling study of hepatocellular carcinoma infected with hepatitis B or hepatitis C virus by using liquid chromatography-mass spectrometry. J. Proteome Res. 2012, 11, 5433–5442. [Google Scholar] [CrossRef] [PubMed]
- Nahon, P.; Amathieu, R.; Triba, M.N.; Bouchemal, N.; Nault, J.-C.; Ziol, M.; Seror, O.; Dhonneur, G.; Trinchet, J.-C.; Beaugrand, M.; et al. Identification of serum proton NMR metabolomic fingerprints associated with hepatocellular carcinoma in patients with alcoholic cirrhosis. Clin. Cancer Res. 2012, 18, 6714–6722. [Google Scholar] [CrossRef] [PubMed]
- Morvan, D.; Demidem, A. Metabolomics by proton nuclear magnetic resonance spectroscopy of the response to chloroethylnitrosourea reveals drug efficacy and tumor adaptive metabolic pathways. Cancer Res. 2007, 67, 2150–2159. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, C.; Nie, X.; Feng, X.; Chen, W.; Yue, Y.; Tang, H.; Deng, F. Metabonomic studies of human hepatocellular carcinoma using high-resolution magic-angle spinning 1H-NMR spectroscopy in conjunction with multivariate data analysis. J. Proteome Res. 2007, 6, 2605–2614. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Granados, B.; Morales, J.M.; Rodrigo, J.M.; Del Olmo, J.; Serra, M.A.; Ferrández, A.; Celda, B.; Monleón, D. Metabolic profile of chronic liver disease by NMR spectroscopy of human biopsies. Int. J. Mol. Med. 2011, 27, 111–117. [Google Scholar] [PubMed]
- Huang, Q.; Tan, Y.; Yin, P.; Ye, G.; Gao, P.; Lu, X.; Wang, H.; Xu, G. Metabolic characterization of hepatocellular carcinoma using nontargeted tissue metabolomics. Cancer Res. 2013, 73, 4992–5002. [Google Scholar] [CrossRef] [PubMed]
- Beyoğlu, D.; Imbeaud, S.; Maurhofer, O.; Bioulac-Sage, P.; Zucman-Rossi, J.; Dufour, J.-F.; Idle, J.R. Tissue metabolomics of hepatocellular carcinoma: Tumor energy metabolism and the role of transcriptomic classification. Hepatol. 2013, 58, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Budhu, A.; Roessler, S.; Zhao, X.; Yu, Z.; Forgues, M.; Ji, J.; Karoly, E.; Qin, L.-X.; Ye, Q.-H.; Jia, H.-L.; et al. Integrated metabolite and gene expression profiles identify lipid biomarkers associated with progression of hepatocellular carcinoma and patient outcomes. Gastroenterology 2013, 144, 1066–1075. [Google Scholar] [CrossRef] [PubMed]
- Paris, D.; Melck, D.; Stocchero, M.; D’Apolito, O.; Calemma, R.; Castello, G.; Izzo, F.; Palmieri, G.; Corso, G.; Motta, A. Monitoring liver alterations during hepatic tumorigenesis by NMR profiling and pattern recognition. Metabolomics 2010, 6, 405–416. [Google Scholar] [CrossRef]
- Darpolor, M.M.; Yen, Y.-F.; Chua, M.-S.; Xing, L.; Clarke-Katzenberg, R.H.; Shi, W.; Mayer, D.; Josan, S.; Hurd, R.E.; Pfefferbaum, A.; et al. In vivo MRSI of hyperpolarized [1-13C]pyruvate metabolism in rat hepatocellular carcinoma. NMR Biomed. 2011, 24, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Koppenol, W.H.; Bounds, P.L.; Dang, C.V. Otto Warburg’s contributions to current concepts of cancer metabolism. Nat. Rev. Cancer 2011, 11, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, S.; Li, Z.; Yang, J.; Huang, C.; Liang, R.; Liu, Z.; Zhou, R. 1H-NMR-based metabolomics of tumor tissue for the metabolic characterization of rat hepatocellular carcinoma formation and metastasis. Tumour Biol. J. 2011, 32, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.Y.; Milsom, J.P.; Sherlock, S. Plasma ratio of valine, leucine and isoleucine to phenylalanine and tyrosine in liver disease. Gut 1978, 19, 1068–1073. [Google Scholar] [CrossRef] [PubMed]
- Stepien, M.; Duarte-Salles, T.; Fedirko, V.; Floegel, A.; Barupal, D.K.; Rinaldi, S.; Achaintre, D.; Assi, N.; Tjønneland, A.; Overvad, K.; et al. Alteration of amino acid and biogenic amine metabolism in hepatobiliary cancers: Findings from a prospective cohort study. Int. J. Cancer 2016, 138, 348–360. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.; Torbenson, M.; Wu, T.-T.; Yeh, M.M. Non-alcoholic fatty liver disease contributes to hepatocarcinogenesis in non-cirrhotic liver: A clinical and pathological study. J. Gastroenterol. Hepatol. 2013, 28, 848–854. [Google Scholar] [CrossRef] [PubMed]
- DeBerardinis, R.J.; Cheng, T. Q’s next: The diverse functions of glutamine in metabolism, cell biology and cancer. Oncogene 2009, 29, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Di Tommaso, L.; Roncalli, M. Tissue biomarkers in hepatocellular tumors: Which, when, and how. Front. Med. 2017, 4, 10. [Google Scholar] [CrossRef] [PubMed]
- Long, J.; Wang, H.; Lang, Z.; Wang, T.; Long, M.; Wang, B. Expression level of glutamine synthetase is increased in hepatocellular carcinoma and liver tissue with cirrhosis and chronic hepatitis B. Hepatol. Int. 2011, 5, 698–706. [Google Scholar] [CrossRef] [PubMed]
- Wasfy, R.E.; Shams Eldeen, A.A. Roles of combined glypican-3 and glutamine synthetase in differential diagnosis of hepatocellular lesions. Asian Pac. J. Cancer Prev. APJCP 2015, 16, 4769–4775. [Google Scholar] [CrossRef] [PubMed]
- Cadoret, A.; Ovejero, C.; Terris, B.; Souil, E.; Lévy, L.; Lamers, W.H.; Kitajewski, J.; Kahn, A.; Perret, C. New targets of beta-catenin signaling in the liver are involved in the glutamine metabolism. Oncogene 2002, 21, 8293–8301. [Google Scholar] [CrossRef] [PubMed]
- Zucman-Rossi, J.; Benhamouche, S.; Godard, C.; Boyault, S.; Grimber, G.; Balabaud, C.; Cunha, A.S.; Bioulac-Sage, P.; Perret, C. Differential effects of inactivated Axin1 and activated beta-catenin mutations in human hepatocellular carcinomas. Oncogene 2007, 26, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Ando, S.; Shibahara, J.; Hayashi, A.; Fukayama, M. β-catenin alteration is rare in hepatocellular carcinoma with steatohepatitic features: Immunohistochemical and mutational study. Virchows Arch. Int. J. Pathol. 2015, 467, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Calderaro, J.; Couchy, G.; Imbeaud, S.; Amaddeo, G.; Letouzé, E.; Blanc, J.-F.; Laurent, C.; Hajji, Y.; Azoulay, D.; Bioulac-Sage, P.; et al. Histological subtypes of hepatocellular carcinoma are related to gene mutations and molecular tumour classification. J. Hepatol. 2017, 64, 727–738. [Google Scholar] [CrossRef] [PubMed]
- Bruix, J.; Sherman, M.; Llovet, J.M.; Beaugrand, M.; Lencioni, R.; Burroughs, A.K.; Christensen, E.; Pagliaro, L.; Colombo, M.; Rodés, J. Clinical management of hepatocellular carcinoma. Conclusions of the Barcelona-2000 EASL conference. European Association for the Study of the Liver. J. Hepatol. 2001, 35, 421–430. [Google Scholar] [CrossRef]
- Fan, T.W.-M. Metabolite profiling by one- and two-dimensional NMR analysis of complex mixtures. Prog. Nucl. Magn. Reson. Spectrosc. 1996, 28, 161–219. [Google Scholar] [CrossRef]
- Wiklund, S.; Johansson, E.; Sjöström, L.; Mellerowicz, E.J.; Edlund, U.; Shockcor, J.P.; Gottfries, J.; Moritz, T.; Trygg, J. Visualization of GC/TOF-MS-based metabolomics data for identification of biochemically interesting compounds using OPLS class models. Anal. Chem. 2008, 80, 115–122. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | HCC-Cirrhosis | HCC-NAFLD | p-Value | |
|---|---|---|---|---|
| Number of patients | - | 9 | 13 | - |
| Sex | Male | 8 (89%) | 11 (85%) | 0.77 |
| Sex | Female | 1 (11%) | 2 (15%) | 0.77 |
| Median [IQR] | 68 [64; 69] | 67 [65; 71] | 0.84 | |
| Hepatitis B and C Viruses | 2 (22%) | - | - | |
| Etiology | Alcohol | 0 (0%) | - | - |
| Etiology | Metabolic Syndrome | 3 (33%) | - | - |
| Metabolic Syndrome + Alcohol | 4 (45%) | - | - | |
| Yes | 7 (78%) | 10 (77%) | 0.96 | |
| Waist Circumference > 94 cm (M) > 80 cm (F) | 8 (89%) | 9 (69%) | 0.28 | |
| Metabolic Syndrome * (IDF/AHA/NHLBI 2009) | Triglycerides > 1.5 g/L or treatment | 2 (22%) | 3 (23%) | 0.96 |
| Metabolic Syndrome * (IDF/AHA/NHLBI 2009) | HDL-cholesterol < 0.4 g/L (M) < 0.5 g/L(F) | 5 (56%) | 7 (54%) | 0.94 |
| Blood Pressure > 135/80 mmHg or hypertension treatment | 5 (56%) | 10 (77%) | 0.29 | |
| Fasting Glucose > 1 g/L or diabetes treatment | 8 (89%) | 9 (69%) | 0.28 | |
| 18.5–24.9 | 1 (11%) | 1 (8%) | 0.78 | |
| 25–29.9 | 5 (56%) | 7 (54%) | 0.94 | |
| 30–34.9 | 3 (33%) | 5 (38%) | 0.81 | |
| Body Mass Index | 35–40 | 0 (0%) | 0 (0%) | - |
| Body Mass Index | <20 ng/mL | 4 (45%) | 11 (85%) | 0.047 |
| 20–200 ng/mL | 3 (33%) | 2 (15%) | 0.96 | |
| >200 ng/mL | 2 (22%) | 0 | 0.075 | |
| Well | 2 (22%) | 5 (38%) | 0.42 | |
| Serum AFP level | Moderately | 7 (78%) | 7 (54%) | 0.25 |
| Serum AFP level | Poorly | 0 (0%) | 1 (8%) | 0.39 |
| Differentiation (WHO Classification) | - | - | - | - |
| Metabolic Subset | Range of Integrated Signal (ppm) | Abbreviation | Metabolite | Median Change [IQR] HCC/NTT % | p-Value | |
|---|---|---|---|---|---|---|
| AQUEOUS PHASE | Glycolysis/TCA Cycle Derivatives | 1.30–1.35 | Lac | Lactate | 49 [19; 113] | <0.0001 |
| 1.43–1.49 | Ala | Alanine | 11 [−18; 50] | 0.49 | ||
| 1.90–1.93 | Ace | Acetate | 26 [−7; 135] | 0.025 | ||
| 2.38–2.41 | Suc | Succinic Acid | −29 [−63; −6] | 0.001 | ||
| 4.61–4.67 | Alpha-Glc | α-glucose | −48 [−71; −25] | 0.001 | ||
| 5.20–5.28 | Beta-Glc | β-glucose | −58 [−77; −23] | <0.0001 | ||
| 5.38–5.43 | Glycogen | Glycogen | −23 [−39; 41] | 0.47 | ||
| 6.49–6.53 | Fum | Fumarate | −11 [−37; 78] | 0.99 | ||
| Ketone bodies | 1.18–1.21 | β-HB | β-hydroxybutyrate | 8 [−19; 158] | 0.2 | |
| Glutamine Derivatives | 2.28–2.39 | Glu | Glutamate | 19 [−13; 170] | 0.019 | |
| 2.41–2.51 | Gln | Glutamine | 70 [9; 112] | 0.0001 | ||
| Amino Acids | 0.93–0.97 | Leu | Leucine | 46 [15; 92] | 0.007 | |
| 1.02–1.04 | Val | Valine | 52 [19; 102] | 0.002 | ||
| 7.16–7.20 | Tyr | Tyrosine | 50 [−2; 145] | 0.007 | ||
| 7.39–7.44 | Phe | Phenylalanine | 43 [−16; 85] | 0.009 | ||
| 7.07–7.11 | His | Histidine | 21 [−27; 155] | 0.09 | ||
| Amino Acid Derivatives | 2.98–3.07 | Creat | Creatine | −18 [−51; 8] | 0.052 | |
| 2.70–2.73 | Sar | Sarcosine | 27 [−2; 163] | 0.011 | ||
| 2.50–2.54 | GSX | Total Glutathione | −17 [−54; 86] | 0.8 | ||
| 3.52–3.56 | Gly | Glycine | −44 [−65; −18] | 0.08 | ||
| 8.43–8.46 | For | Formate | 27 [−12; 139] | 0.071 | ||
| Nucleotides | 5.73–5.83 | Uracil | Uracil | 138 [24; 381] | <0.0001 | |
| Vitamines and Co-factors | 4.49–4.53 | Asc A | Ascorbic acid | 16 [−16; 43] | 0.047 | |
| 8.40–8.43 | NAD | NAD | 50 [−3; 86] | 0.024 | ||
| Phospholipid Derivatives | 3.18–3.20 | Cho | Choline | 36 [−9; 110] | 0.013 | |
| 3.21–3.22 | PC | Phosphocholine | 59 [−12; 96] | 0.002 | ||
| 3.22–3.23 | GPC | Glycerophosphocholine | 0 [−26; 24] | 0.598 | ||
| 3.99–4.02 | PE | Phosphoethanolamine | 88 [−25; 284] | 0.01 | ||
| LIPID PHASE | Phospholipid Derivatives | 3.06–3.09 | MethylPtEth | Methyl-phosphatidylethanolamine | 0 [0; 100] | 0.19 |
| 3.12–3.18 | PtEth | Phosphatidylethanolamine | 3 [−21; 33] | 1 | ||
| 3.22–3.29 | PtCho+SM | Phosphatidylcholine + Sphingomyeline | 13 [−4; 46] | 0.06 | ||
| Cholesterol | 0.70–0.75 | T Chol CH3 | Total cholesterol CH3 | 29 [−8; 116] | 0.025 | |
| 1.03–1.05 | E Chol | Esterified cholesterol | −14 [−42;15] | 0.07 | ||
| Fatty Acids | 0.86–0.96 | T FA | Total fatty acids (terminal CH3) | 1 [−3; 6] | 0.2 | |
| 1.59–1.70 | β-CH2 FA | Fatty acids (β-CH2) | −7 [−15; 2] | 0.038 | ||
| 2.32–2.41 | α-CH2 FA | Fatty acids (α-CH2) | −3 [−11; 10] | 0.82 | ||
| Saturated Fatty Acids | 1.24–1.44 | Sat FA | Saturated FA -(CH2)n- | −1 [−4; 4] | 0.9 | |
| Unsaturated Fatty Acids | 2.02–2.12 | MUFA | Allylic -CH2-CH=CH Mono-Unsaturated fatty acids | −13 [−27; −1] | 0.002 | |
| 5.23–5.53 | UFA | Olefinic -CH=CH- FA | −6 [−15; 6] | 0.2 | ||
| 1.70–1.74 | Ara A | CH2 arachidonic acid and eicosapentaenoic acid | −66 [−76; −35] | 0.0001 | ||
| 2.77–2.83 | Lin A | CH2 linoleic acid | −19 [−36; −7] | 0.0001 | ||
| 2.85–2.95 | PUFA | Diallylic CH2 Polyunsaturated fatty acids =CH-CH2-CH= | 4 [−17; 21] | 0.71 | ||
| TAG | 4.15–4.25 | TAG | Triacylglycerol | −6 [−14; 6] | 0.17 | |
| Glycero PL | 3.99–4.10 | GPL | Glycerophospholipid backbone CH2-OP | −2 [−24; 30] | 0.62 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teilhet, C.; Morvan, D.; Joubert-Zakeyh, J.; Biesse, A.-S.; Pereira, B.; Massoulier, S.; Dechelotte, P.; Pezet, D.; Buc, E.; Lamblin, G.; et al. Specificities of Human Hepatocellular Carcinoma Developed on Non-Alcoholic Fatty Liver Disease in Absence of Cirrhosis Revealed by Tissue Extracts 1H-NMR Spectroscopy. Metabolites 2017, 7, 49. https://doi.org/10.3390/metabo7040049
Teilhet C, Morvan D, Joubert-Zakeyh J, Biesse A-S, Pereira B, Massoulier S, Dechelotte P, Pezet D, Buc E, Lamblin G, et al. Specificities of Human Hepatocellular Carcinoma Developed on Non-Alcoholic Fatty Liver Disease in Absence of Cirrhosis Revealed by Tissue Extracts 1H-NMR Spectroscopy. Metabolites. 2017; 7(4):49. https://doi.org/10.3390/metabo7040049
Chicago/Turabian StyleTeilhet, Camille, Daniel Morvan, Juliette Joubert-Zakeyh, Anne-Sophie Biesse, Bruno Pereira, Sylvie Massoulier, Pierre Dechelotte, Denis Pezet, Emmanuel Buc, Géraldine Lamblin, and et al. 2017. "Specificities of Human Hepatocellular Carcinoma Developed on Non-Alcoholic Fatty Liver Disease in Absence of Cirrhosis Revealed by Tissue Extracts 1H-NMR Spectroscopy" Metabolites 7, no. 4: 49. https://doi.org/10.3390/metabo7040049
APA StyleTeilhet, C., Morvan, D., Joubert-Zakeyh, J., Biesse, A.-S., Pereira, B., Massoulier, S., Dechelotte, P., Pezet, D., Buc, E., Lamblin, G., Peoc’h, M., Porcheron, J., Vasson, M.-P., Abergel, A., & Demidem, A. (2017). Specificities of Human Hepatocellular Carcinoma Developed on Non-Alcoholic Fatty Liver Disease in Absence of Cirrhosis Revealed by Tissue Extracts 1H-NMR Spectroscopy. Metabolites, 7(4), 49. https://doi.org/10.3390/metabo7040049