
Sexual Dimorphism in the Response of Mercurialis annua to Stress


 ,
,  ,
,
Abstract
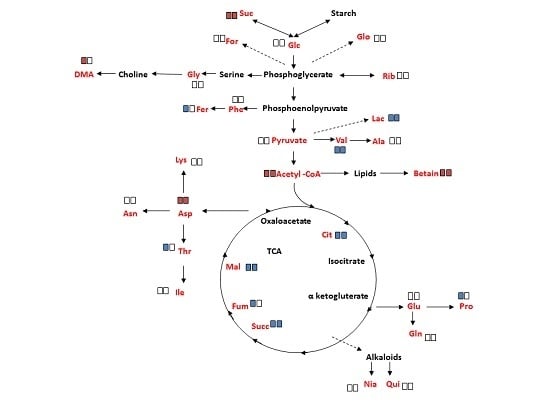
:
1. Introduction
2. Results
2.1. Development of M. annua Plants under Control and in Response to Salinity
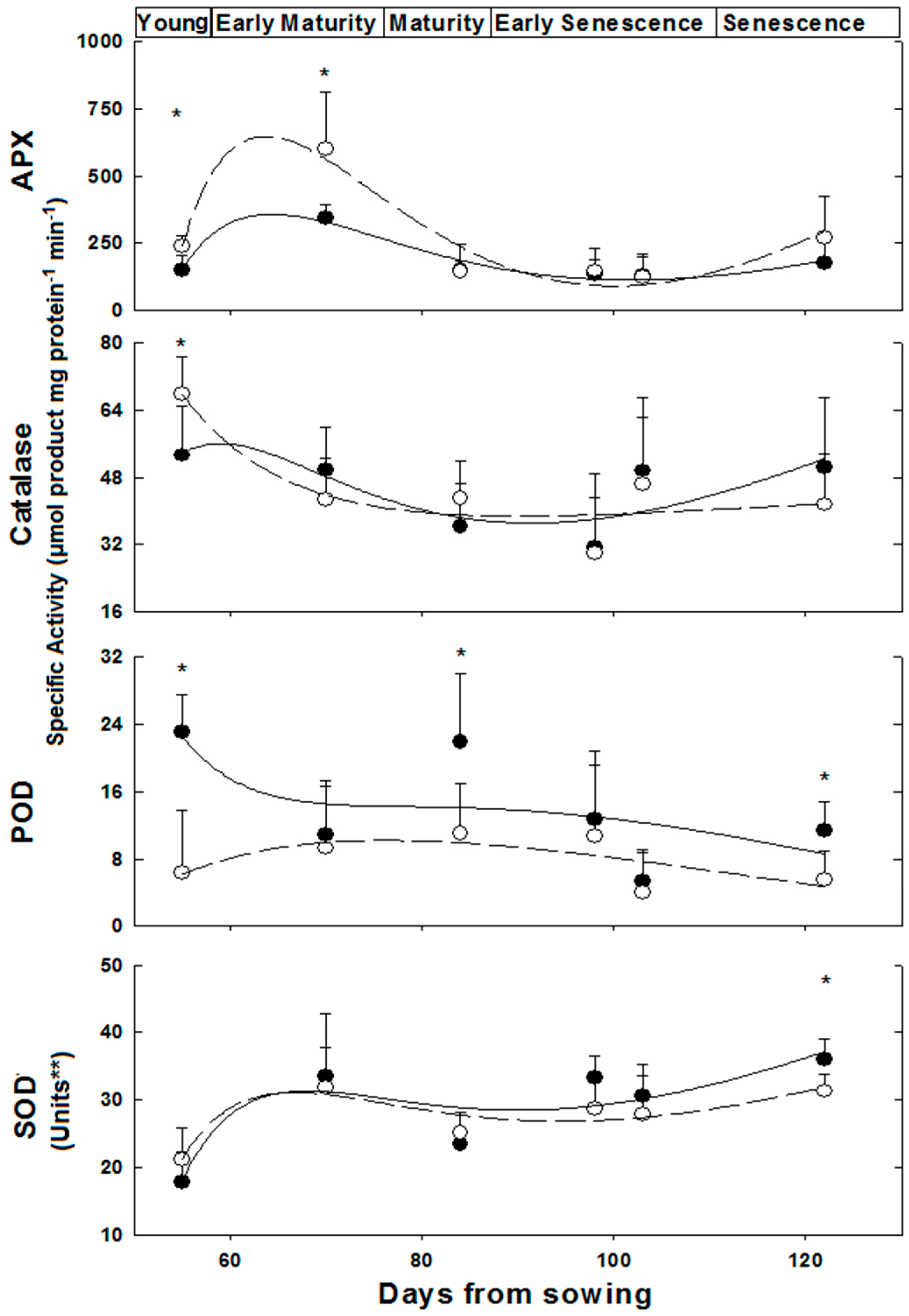
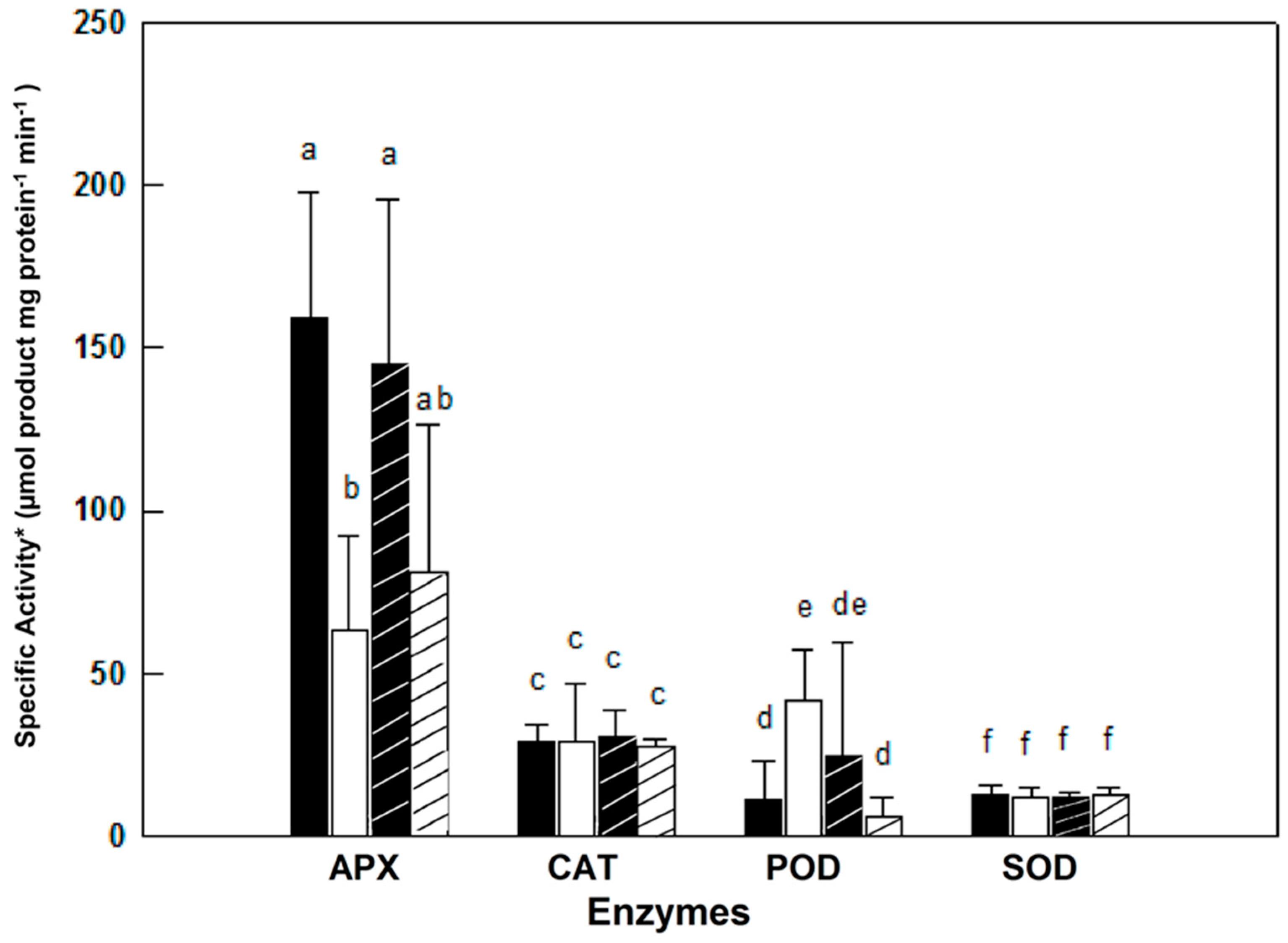
2.2. Anti-Oxidative and Detoxification Enzymes under Control and in Response to Salinity
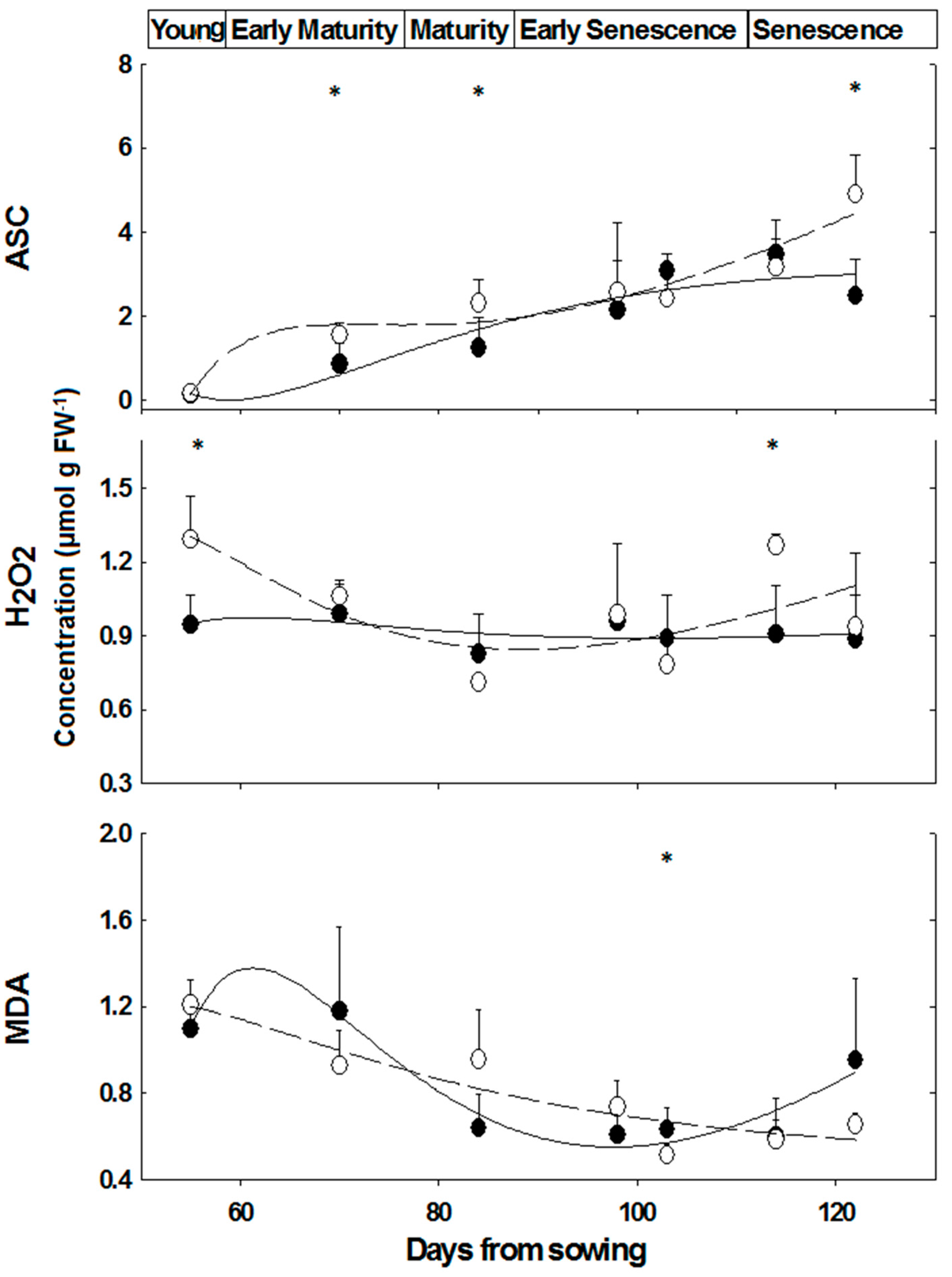
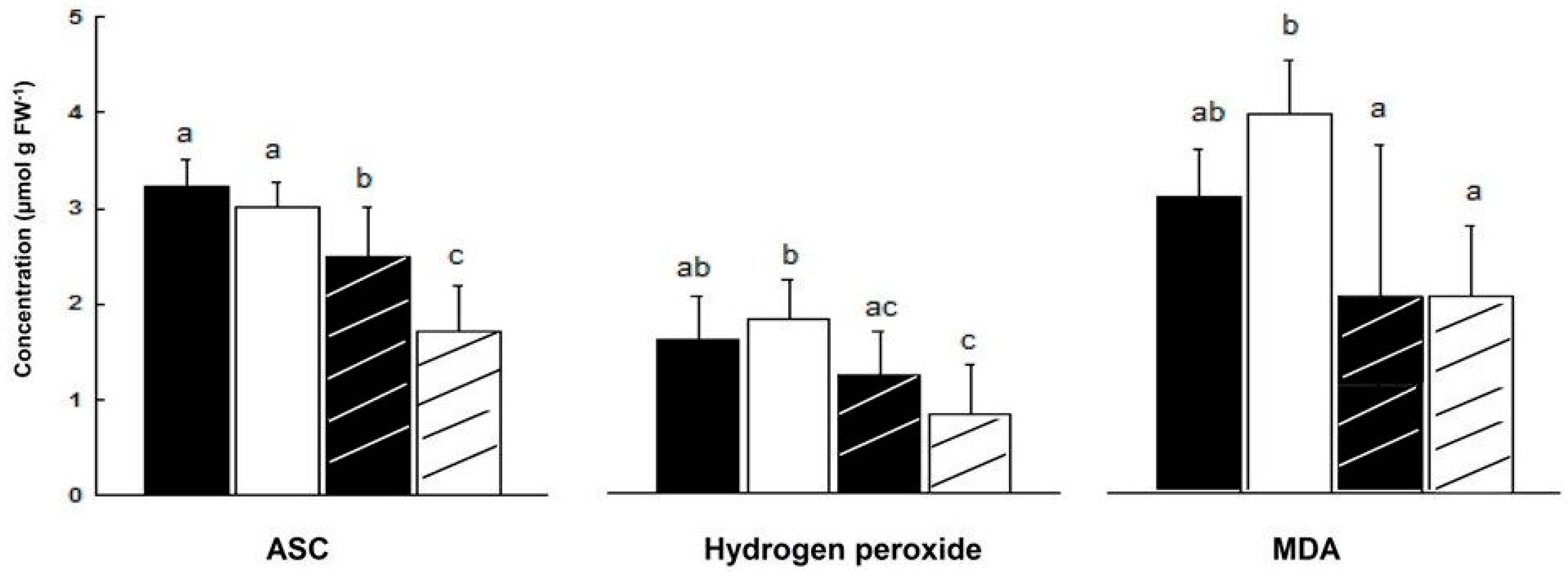
2.3. Content of Oxidative/Anti-Oxidative Substrates
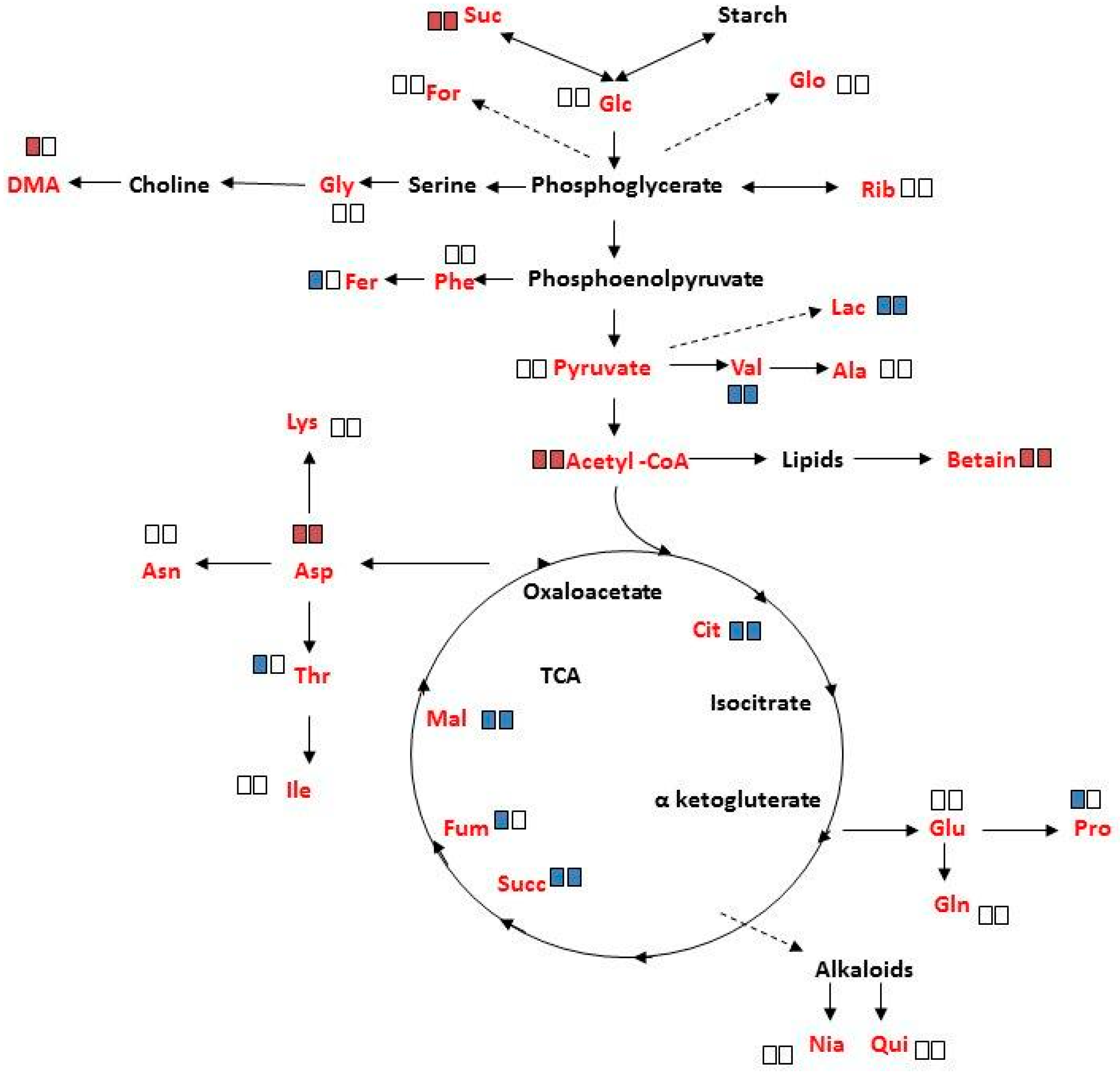
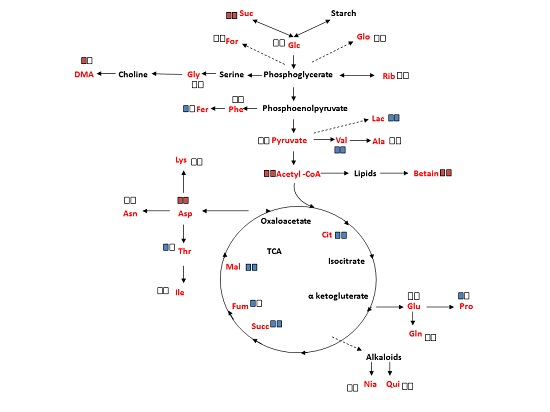
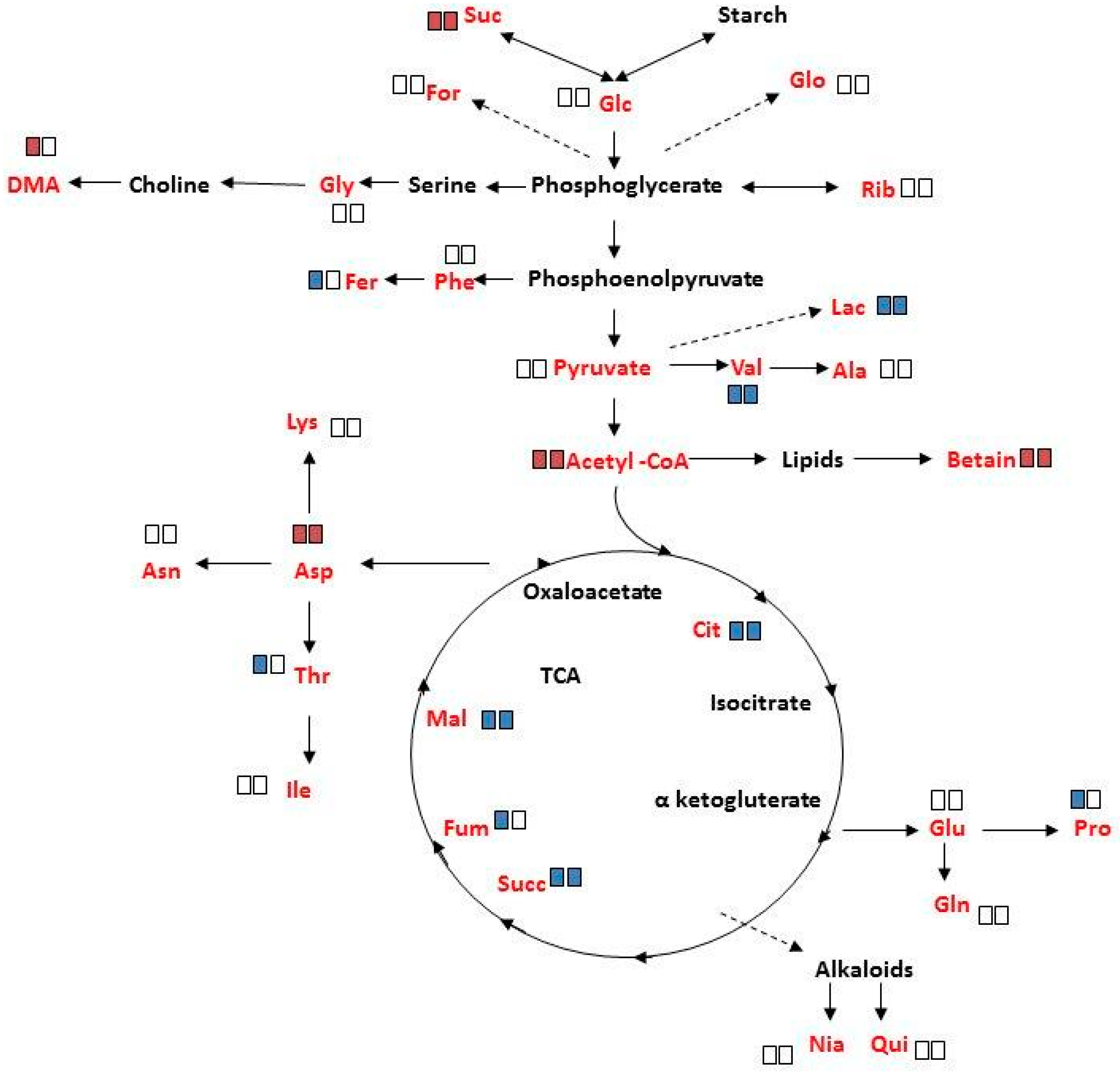
2.4. Overall Metabolite Profiles of M. annua under Salinity
3. Discussion
3.1. Dioecious Plant Development
3.2. Enzymes and Substrates during Development and Salinity
3.3. Metabolite Changes in M. annua under Salinity
4. Experimental Section
4.1. Plant Material and Growth Conditions
4.2. Experimental Design
4.3. CO2/H2O Gas Exchange
4.4. Protein Extraction
4.5. Enzyme Assays
4.6. Anti-Oxidative Metabolites Extraction and Assay
4.7. Metabolite Profiling and NMR Identification
5. Statistical Analysis
Supplementary Materials
Acknowledgments
Author contributions
Conflicts of Interest
Abbreviations
| APX | Ascorbate peroxidase |
| ASC | Ascorbate |
| CAT | Catalase |
| DMA | dimethylamine |
| GST | Glutathione S-transferase |
| MDA | Malondialdehyde |
| NBT | Nitro-blue-tetrazolium |
| POD | Guaiacol Peroxidase |
| ROS | Reactive Oxygen Species |
| SOD | Superoxide dismutase |
| TCA | Tricarboxylic Acid Cycle |
References
- Bawa, K.S. Evolution of dioecy in flowering plants. Annu. Rev. Ecol. Syst. 1980, 11, 15–39. [Google Scholar] [CrossRef]
- Juvany, M.; Munné-Bosch, S. Sex-related differences in stress tolerance in dioecious plants: A critical appraisal in a physiological context. J. Exp. Bot. 2015. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.; Nobuhiro Suzukis, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Redox regulation in photosynthetic organisms: Signaling, acclimation, and practical implications. Antioxid. Redox Signal. 2009, 11, 862–905. [Google Scholar] [CrossRef] [PubMed]
- Dionisio-Sese, M.L.; Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Plant Sci. 1998, 135, 1–9. [Google Scholar] [CrossRef]
- Shalata, A.; Mittova, V.; Volokita, M.; Guy, M.; Tal, M. Response of the cultivated tomato and its wild salt-tolerant relative Lycopersicon pennellii to salt-dependent oxidative stress: The root anti-oxidative system. Physiol. Plant. 2001, 112, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Mittova, V.; Tal, M.; Volokita, M.; Guy, M. Upregulation of the leaf mitochondrial and peroxisomal antioxidative systems in response to salt-induced oxidative stress in the wild salt-tolerant tomato species Lycopersicon pennellii. Plant Cell Environ. 2003, 26, 845–856. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef] [PubMed]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K.V. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Schauer, N.; Fernie, A.R. Plant metabolomics: Towards biological function and mechanism. Trends Plant Sci. 2006, 11, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Verpoorte, R.; Choi, Y.H.; Kim, H.K. NMR-based metabolomics at work in phytochemistry. Phytochem. Rev. 2007, 6, 3–14. [Google Scholar] [CrossRef]
- Eisenreich, W.; Bacher, A. Advances of high-resolution NMR techniques in the structural and metabolic analysis of plant biochemistry. Phytochemistry 2007, 68, 2799–2815. [Google Scholar] [CrossRef] [PubMed]
- Rozas, V.; DeSoto, L.; Olano, J.M. Sex-specific, age-dependent sensitivity of tree-ring growth to climate in the dioecious tree Juniperus thurifera. New Phytol. 2009, 182, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.Q.; Zheng, Q.S.; Liu, Z.P.; Xu, W.J.; Liu, L.; Zhao, G.M.; Long, X.H. Overexpression of Arabidopsis thaliana Na+/H+ antiporter gene enhanced salt resistance in transgenic poplar (Populus x euramericana ‘Neva’). Trees Struct. Funct. 2012, 26, 685–694. [Google Scholar] [CrossRef]
- Li, J.Y.; Zhao, C.Y.; Li, J.; Yan, Y.Y.; Yu, B.; Han, M. Growth and leaf gas exchange in Populus euphratica across soil water and salinity gradients. Photosynthetica 2013, 51, 321–329. [Google Scholar] [CrossRef]
- Durand, R.; Durand, B. Sex determination and reproductive organ differentiation in Mercurialis. Plant Sci. 1991, 80, 49–65. [Google Scholar] [CrossRef]
- Pannell, J.R.; Dorken, M.E.; Pujol, B.; Berjano, R. Gender variation and transitions between sexual systems in Mercurialis annua (Euphorbiaceae). Int. J. Plant Sci. 2008, 169, 129–139. [Google Scholar] [CrossRef]
- Pannell, J.R. Mixed genetic and environmental sex determination in an androdioecious population of Mercurialis annua. Heredity 1997, 78, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; El Aidi, J.; Ait-Ali, T.; Augur, C.; Teller, G.; Schoentgen, F.; Durand, R.; Durand, B. Sex-specific marker and trans-zeatin ribosidase in female annual Mercury. Plant Sci. 1998, 139, 93–103. [Google Scholar] [CrossRef]
- Khadka, D.K.; Nejidat, A.; Tal, M.; Golan-Goldhirsh, A. DNA markers for sex: Molecular evidence for gender dimorphism in dioecious Mercurialis annua L. Mol. Breed. 2002, 9, 251–257. [Google Scholar] [CrossRef]
- Khadka, D.K.; Nejidat, A.; Tal, M.; Golan-Goldhirsh, A. Molecular characterization of a gender-linked DNA marker and a related gene in Mercurialis annua L. Planta 2005, 222, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Vilas, J.; Pannell, J.R. Sexual dimorphism in resource acquisition and deployment: Both size and timing matter. Ann. Bot. 2011, 107, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Vilas, J.; Pannell, J.R. Do plants adjust their sex allocation and secondary sexual morphology in response to their neighbors? Ann. Bot. 2012, 110, 1471–1478. [Google Scholar] [CrossRef] [PubMed]
- Sánchez Vilas, J.; Pannell, J.R. Sex-differential herbivory in androdioecious Mercurialis annua. PLoS ONE 2011, 6, e22083. [Google Scholar] [CrossRef] [PubMed]
- Hesse, E.; Pannell, J.R. Sexual dimorphism in a dioecious population of the wind-pollinated herb Mercurialis annua: The interactive effects of resource availability and competition. Ann. Bot. 2011, 107, 1039–1045. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, P.; Zentgraf, U. The correlation between oxidative stress and leaf senescence during plant development. Cell. Mol. Biol. Lett. 2005, 10, 515–534. [Google Scholar] [PubMed]
- Renner, S.S.; Ricklefs, R.E. Dioecy and its correlates in the flowering plants. Am. J. Bot. 1995, 82, 596–606. [Google Scholar] [CrossRef]
- Jiang, H.; Peng, S.; Zhang, S.; Li, X.; Kropelainen, H.; Li, C. Transcriptional analysis in Populus yunnanensis provides insights into molecular mechanisms of sexual differences in salinity tolerance. J. Exp. Bot. 2012, 63, 3709–3726. [Google Scholar] [CrossRef] [PubMed]
- Juvany, M.; Müller, M.; Pintó-Marijuan, M.; Munné-Bosch, S. Sex-related differences in lipid peroxidation and photoprotection in Pistacia lentiscus. J. Exp. Bot. 2014, 65, 1039–1049. [Google Scholar] [CrossRef] [PubMed]
- Decker, K.L.; Pilson, D. Biased sex ratios in the dioecious annual Croton texensis (Euphorbiaceae) are not due to environmental sex determination. Am. J. Bot. 2000, 87, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Bram, M.R.; Quinn, J.A. Sex expression, sex-specific traits, and the effect of salinity on growth and reproduction of Amaranthus cannabinus (Amaranthaceae), a dioecious annual. Am. J. Bot. 2000, 87, 1609–1618. [Google Scholar] [CrossRef] [PubMed]
- Zluvova, J.; Zak, J.; Janousek, B.; Vyskot, B. Dioecious Silene latifolia plants show sexual dimorphism in the vegetative stage. BMC Plant Biol. 2010, 10, 208. [Google Scholar] [CrossRef] [PubMed]
- Conn, J.S. Phenological differentiation between the sexes of Rumex hastalulus: Niche partitioning or different optimal reproductive strategies? Bull. Torrey Bot. Club 1981, 108, 374–378. [Google Scholar] [CrossRef]
- Conn, J.S.; Blum, U. Differentiation between the sexes of Rumex hastatulus in net energy allocation, flowering and height. Bull. Torrey Bot. Club 1981, 108, 446–455. [Google Scholar] [CrossRef]
- Doust, J.L.; O’Brien, G.; Doust, L.L. Effect of density on secondary sex characteristics and sex ratio in Silene alba (Caryophyllaceae). Am. J. Bot. 1987, 74, 40–46. [Google Scholar] [CrossRef]
- Case, A.L.; Barrett, S.C.H. Environmental stress and the evolution of dioecy: Wurmbea dioica (Colchicaceae) in Western Australia. Evol. Ecol. 2004, 18, 145–164. [Google Scholar] [CrossRef]
- Gehring, J.L.; Linhart, Y.B. Sexual dimorphisms and response to low resources in the dioecious plant Silene latifolia (Caryophyllaceae). Int. J. Plant Sci. 1993, 154, 152–162. [Google Scholar] [CrossRef]
- Lloyd, A.; Webb, C.J. Secondary sex characteristics in plants. Bot. Rev. 1977, 43, 177–216. [Google Scholar] [CrossRef]
- Baker, H.G.; Cox, P.A. Further thoughts on dioecism and islands. Ann. Mo. Bot. Gard. 1984, 71, 244–253. [Google Scholar] [CrossRef]
- Anderson, G.J.; Stebbins, G.L. Dioecy versus gametophytic self-incompatibility: A test. Am. Nat. 1984, 124, 423–428. [Google Scholar] [CrossRef]
- Correns, C. Bestimmung, Vererbung und Verteilung des Geschlechtes bei den höheren Pflanzen. In Handbuch der Vererbungswissenschaft; Baur, E., Hartmann, M., Eds.; Gebrüder Borntraeger: Berlin, Germany, 1928; Volume 2, pp. 1–128. [Google Scholar]
- Zhao, H.; Li, Y.; Duan, B.; Korpelainen, H.; Li, C. Sex-related adaptive responses of Populus cathayana to photoperiod transitions. Plant Cell Environ. 2009, 32, 1401–1411. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Wei, X.D.; Chen, G.X.; Xu, Y.L. Changes in Photosynthetic Characteristics and Antioxidative Protection in Male and Female Ginkgo during Natural Senescence. J. Am. Soc. Hortic. Sci. 2012, 137, 349–360. [Google Scholar]
- Dhindsa, R.S.; Plumbhindsa, P.; Thorpe, T.A. Leaf Senescence: Correlated with increased levels of membrane permeability and lipid-peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Asada, K. Ascorbate peroxidase—A hydrogen peroxide scavenging enzyme in plants. Physiol. Plant. 1992, 85, 235–241. [Google Scholar] [CrossRef]
- Jiménez, A.; Hernández, J.A.; Pastori, G.; del Río, L.A.; Sevilla, F. Role of the ascorbate-glutathione cycle of mitochondria and peroxisomes in the senescence of pea leaves. Plant Physiol. 1998, 118, 1327–1335. [Google Scholar] [CrossRef] [PubMed]
- Lim, P.O.; Kim, H.J.; Nam, H.G. Leaf senescence. Annu. Rev. Plant Biol. 2007, 58, 115–136. [Google Scholar] [CrossRef] [PubMed]
- Sekmen, A.H.; Türkan, I.; Takio, S. Differential responses of antioxidative enzymes and lipid peroxidation to salt stress in salt-tolerant Plantago maritima and salt-sensitive Plantago media. Physiol. Plant. 2007, 131, 399–411. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Ghanashyam, C.; Bhattacharjee, A. Comprehensive expression analysis suggests overlapping and specific roles of rice glutathione S-transferase genes during development and stress responses. BMC Genom. 2010, 11, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Hirao, J.; Nishimura, M.; Arakawa, S.; Niino, N.; Mori, K.; Furukawa, T.; Sanbuissho, A.; Manabe, S.; Nishihara, M.; Mori, Y. Sex and circadian modulatory effects on rat liver as assessed by transcriptome analyses. J. Toxicol. Sci. 2011, 36, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Li, Y.; Hong, H.; Chang, C.-W.; Guo, L.-W.; Lyn-Cook, B.; Shi, L.; Ning, B. Sex Differences in the Expression of Drug-Metabolizing and Transporter Genes in Human Liver. J. Drug Metab. Toxicol. 2012, 3, 1–9. [Google Scholar]
- Dixon, D.P.; McEwen, A.G.; Lapthorn, A.J.; Edwards, R. Forced Evolution of a Herbicide Detoxifying Glutathione Transferase. J. Biol. Chem. 2003, 278, 23930–23935. [Google Scholar] [CrossRef] [PubMed]
- Cramer, G.R.; Ergül, A.; Grimplet, J.; Tillett, R.L.; Tattersall, E.A.R.; Bohlman, M.C.; Vincent, D.; Sonderegger, J.; Evans, J.; Osborne, C.; et al. Water and salinity stress in grapevines: Early and late changes in transcript and metabolite profiles. Funct. Integr. Genom. 2007, 7, 111–134. [Google Scholar] [CrossRef] [PubMed]
- Fumagalli, E.; Baldoni, E.; Abbruscato, P.; Piffanelli, P.; Genga, A.; Lamanna, R.; Consonni, R. NMR Techniques Coupled with Multivariate Statistical Analysis: Tools to Analyse Oryza sativa Metabolic Content under Stress Conditions. J. Agron. Crop Sci. 2009, 195, 77–88. [Google Scholar] [CrossRef]
- Bailey, N.J.C.; Oven, M.; Holmes, E.; Nicholson, J.K.; Zenk, M.H. Metabolomic analysis of the consequences of cadmium exposure in Silene cucubalus cell cultures via 1H NMR spectroscopy and chemometrics. Phytochemistry 2003, 62, 851–858. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Du, Y.; Chen, S.; Tang, H. Dynamic metabolomic responses of Tobacco (Nicotiana tabacum) plants to salt stress. J. Proteome Res. 2011, 10, 1904–1914. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, P.; Duckstein, S.; Conrad, J.; Knödler, M.; Meyer, U.; Stintzing, F.C. An approach to the chemotaxonomic differentiation of two European Dog’s Mercury Species: Mercurialis annua L. and M. perennis L. Chem. Biodivers. 2012, 9, 282–297. [Google Scholar] [CrossRef] [PubMed]
- Mittova, V.; Volokita, M.; Guy, M.; Tal, M. Activities of SOD and the ascorbate-glutathione cycle enzymes in subcellular compartments in leaves and roots of the cultivated tomato and its wild salt tolerant relative Lycopersicon pennellii. Physiol. Plant. 2000, 110, 42–51. [Google Scholar] [CrossRef]
- Schröder, P.; Maier, H.; Debus, R. Detoxification of herbicides in Phragmites australis. Z. Naturforschung Sect. C J. Biosci. 2005, 60, 317–324. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Jiménez, A.; Hernández, J.A.; del Rio, L.A.; Sevilla, F. Evidence for the presence of the ascorbate-glutathione cycle in mitochondria and peroxisomes of pea leaves. Plant Physiol. 1997, 114, 275–284. [Google Scholar] [PubMed]
- Zelitch, I. Physiological investigations of a tobacco mutant with O2-resistant photosynthesis and enhanced catalase activity. Plant Physiol. 1990, 93, 1521–1524. [Google Scholar] [CrossRef] [PubMed]
- Beyer, W.F.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Ann. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Kuroda, M.; Ozawa, T.; Imagawa, H. Changes in chloroplast peroxidase activities in relation to chlorophyll loss in barley leaf segments. Physiol. Plant. 1990, 80, 555–560. [Google Scholar] [CrossRef]
- Scalla, R.; Roulet, A. Cloning and characterization of a glutathione S-transferase induced by a herbicide safener in barley (Hordeum vulgare). Physiol. Plant. 2002, 116, 336–344. [Google Scholar] [CrossRef]
- Law, M.Y.; Charles, S.A.; Halliwell, B. Glutathione and ascorbic acid in spinach (Spinacia oleracea) chloroplasts. Biochem. J. 1983, 210, 899–903. [Google Scholar] [CrossRef] [PubMed]
- Draper, H.H.; Hadley, M. Malondialdehyde determination as index of lipid peroxidation. Methods Enzymol. 1990, 186, 421–431. [Google Scholar] [PubMed]
- Wolff, S. Ferrous ion oxidation in presence of ferric ion indicator xylenol orange for measurement of hydroperoxides. Methods Enzymol. 1994, 233, 182–189. [Google Scholar]
- Bouzier, A.K.; Thiaudiere, E.; Biran, M.; Rouland, R.; Canioni, P.; Merle, M. The metabolism of [3-(13)C]lactate in the rat brain is specific of a pyruvate carboxylase-deprived compartment. J. Neurochem. 2000, 75, 480–486. [Google Scholar] [CrossRef] [PubMed]

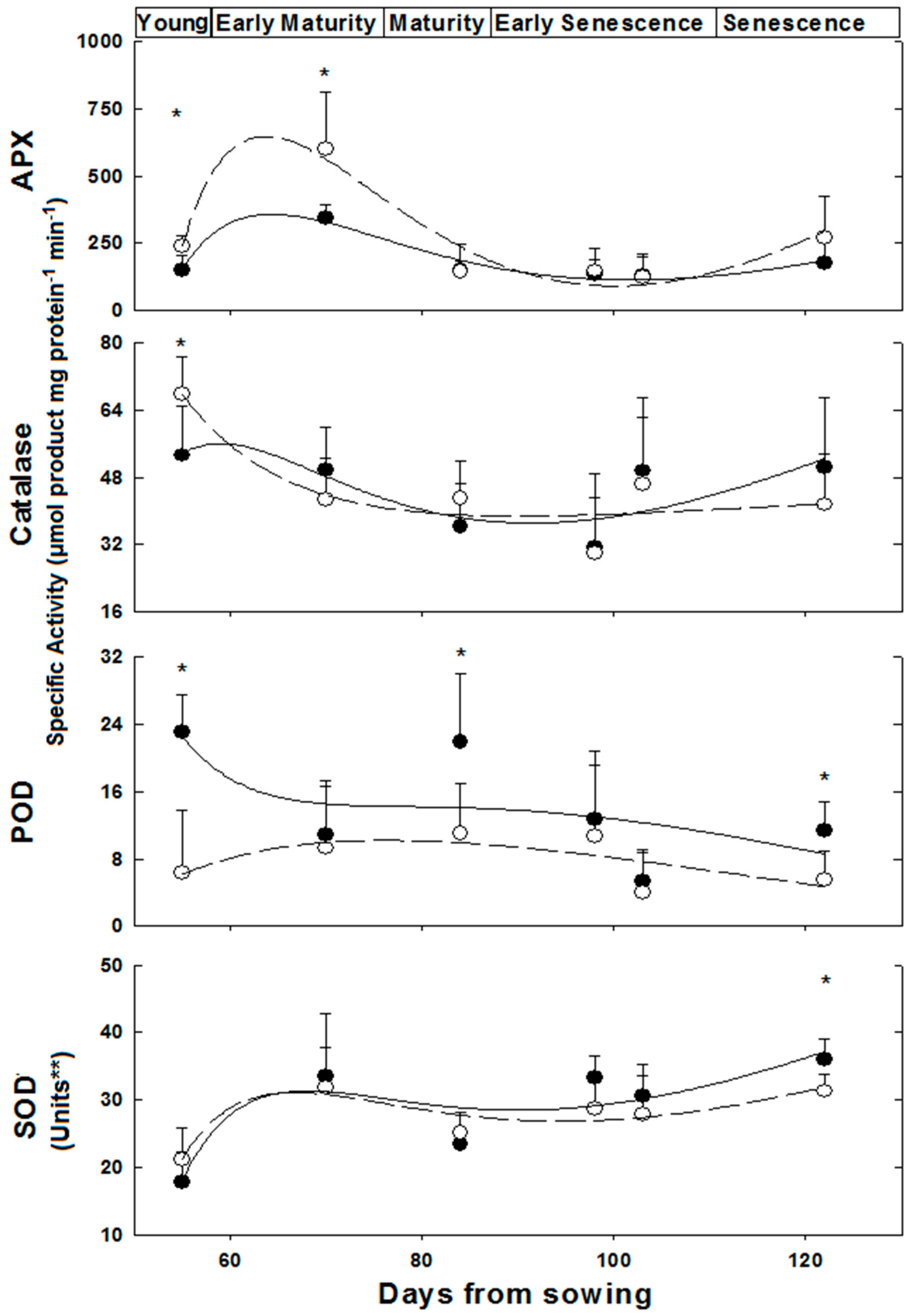
 —female control;
—female control;  —male control;
—male control;  —female salinity;
—female salinity;  —male salinity.
—female control; —male control; —female salinity; —male salinity.
—male salinity.
—female control; —male control; —female salinity; —male salinity.
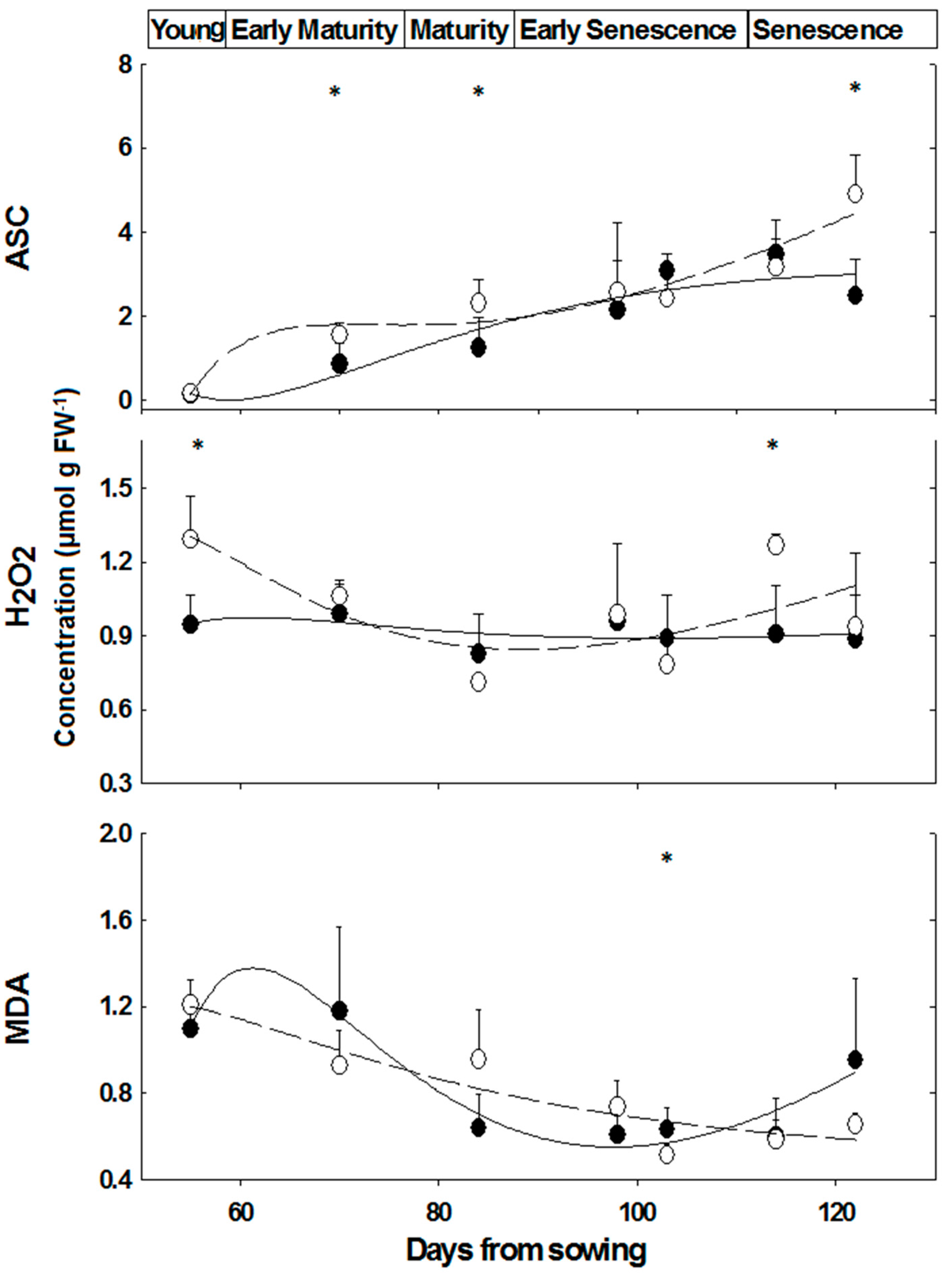 —female control; —male control; —female salinity; —male salinity.
—female control; —male control; —female salinity; —male salinity.
—female control; —male control; —female salinity; —male salinity.
—female control; —male control; —female salinity; —male salinity.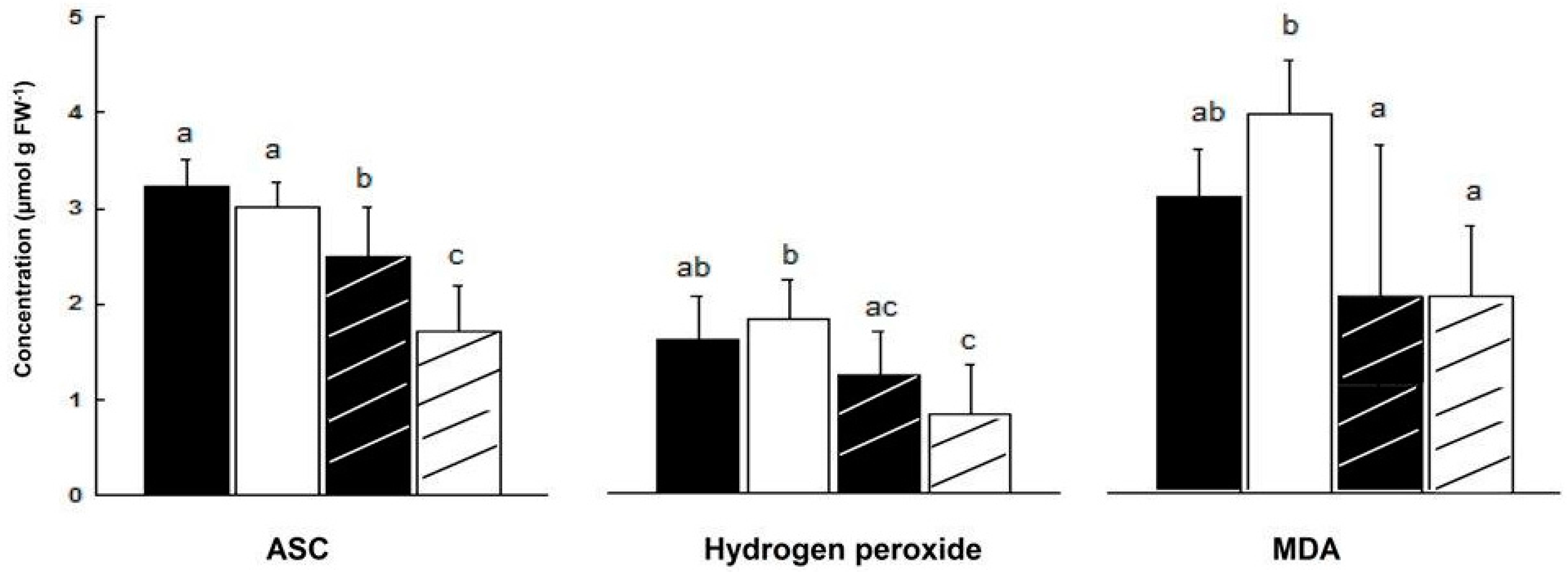

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (Days *) | Plant Height (cm) | Root Length (cm) | Reproductive Node (No.) | |||
|---|---|---|---|---|---|---|
| male | female | male | female | male | female | |
| 55 (‘young’) | 25 ± 5 a | 25 ± 2 a | 5 ± 1 a | 5 ± 1 a | 5 ± 0 a | 5 ± 0 a |
| 70 (‘early maturity’) | 40 ± 4 b | 38 ± 2 b | 6 ± 4 a,b | 6 ± 1 a,b | 7 ± 1 b | 7 ± 1 b |
| 84 (‘mature stage‘) | 48 ± 0 c | 48 ± 3 c | 8 ± 0 b | 7 ± 1 b | 7 ± 1 b | 6 ± 1 a,b |
| 98 (‘early senescence’) | 60 ± 3 d | 61 ± 3 d | 8 ± 1 b | 8 ± 1 b | 6 ± 1 a,b | 7 ± 1 b |
| 114 (‘senescence‘) | 61 ± 5 d | 54 ± 5 d | 12 ± 5 c | 13 ± 1 c | 6 ± 1 a,b | 12 ± 1 c |
| 122 (‘late senescence’) | 77 ± 16 d | 58 ± 11 d | 14 ± 5 b,c | 9 ± 3 b,c | 8 ± 1 b | 11 ± 1 c |
| Treatment | Gender | Fresh Weight (gr) | Plant Height (cm) | Root Length (cm) | Reproductive Node (No.) |
|---|---|---|---|---|---|
| Control | Male | 20 ± 7 a | 63 ± 7 b | 13 ± 7 c | 9 ± 1 d |
| Salinity | Female | 23 ± 11 a | 56 ± 10 b | 17 ± 5 c | 9 ± 1 d |
| Male | 18 ± 6 a | 59 ± 9 b | 13 ± 3 c | 7 ± 2 e | |
| Female | 20 ± 8 a | 54 ± 7 b | 14 ± 4 c | 12 ± 1 f |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orlofsky, E.M.; Kozhoridze, G.; Lyubenova, L.; Ostrozhenkova, E.; Winkler, J.B.; Schröder, P.; Bacher, A.; Eisenreich, W.; Guy, M.; Golan-Goldhirsh, A. Sexual Dimorphism in the Response of Mercurialis annua to Stress. Metabolites 2016, 6, 13. https://doi.org/10.3390/metabo6020013
Orlofsky EM, Kozhoridze G, Lyubenova L, Ostrozhenkova E, Winkler JB, Schröder P, Bacher A, Eisenreich W, Guy M, Golan-Goldhirsh A. Sexual Dimorphism in the Response of Mercurialis annua to Stress. Metabolites. 2016; 6(2):13. https://doi.org/10.3390/metabo6020013
Chicago/Turabian StyleOrlofsky, Ezra M., Giorgi Kozhoridze, Lyudmila Lyubenova, Elena Ostrozhenkova, J. Barbro Winkler, Peter Schröder, Adelbert Bacher, Wolfgang Eisenreich, Micha Guy, and Avi Golan-Goldhirsh. 2016. "Sexual Dimorphism in the Response of Mercurialis annua to Stress" Metabolites 6, no. 2: 13. https://doi.org/10.3390/metabo6020013
APA StyleOrlofsky, E. M., Kozhoridze, G., Lyubenova, L., Ostrozhenkova, E., Winkler, J. B., Schröder, P., Bacher, A., Eisenreich, W., Guy, M., & Golan-Goldhirsh, A. (2016). Sexual Dimorphism in the Response of Mercurialis annua to Stress. Metabolites, 6(2), 13. https://doi.org/10.3390/metabo6020013