
Plasma Lipidomic Remodeling in Behçet’s Disease Reveals Alterations Associated with Vascular Involvement

 ,
,  and
and
Highlights
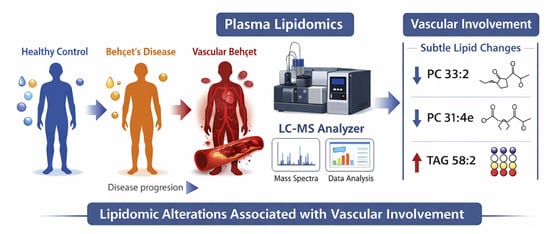
- Plasma lipidomics revealed coordinated remodeling of membrane, signaling, and energy-related lipids in Behçet’s disease.
- Vascular involvement was associated with subtle, lipid-specific alterations rather than global lipidomic changes.
- Lipidomic alterations suggest integrated changes in membrane organization, inflammatory signaling, and metabolic adaptation in BD
- These findings provide a basis for future mechanistic and targeted lipidomic studies in vascular involvement.
Abstract
1. Introduction
2. Materials and Methods
2.1. The Study Population
2.2. Plasma Lipidomics Analysis
2.3. Statistical Analysis
2.4. Class-Based Pathway Analysis
3. Results
3.1. Comparison of Lipid Profile Between BD and Healthy Control
3.2. Comparison of Lipid Profile Between BD and VBD Groups
4. Discussion
4.1. Differentiation of Lipid Profile Between BD and Healthy Control
4.1.1. Phosphatidylcholines (PCs)
4.1.2. Phosphatidylinositols (PIs)
4.1.3. Diacylglycerols (DAGs)
4.1.4. Sphingomyelins (SMs)
4.1.5. Triacylglycerols (TAGs)
4.1.6. Biological Implications of Altered Lipid Classes
4.1.7. Class-Based Pathway Analysis of Altered Lipids
4.2. Lipid-Based Differentiation with Vascular Involvement in BD
4.3. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alpsoy, E. Behçet’s Disease: A Comprehensive Review with a Focus on Epidemiology, Etiology and Clinical Features, and Management of Mucocutaneous Lesions. J. Dermatol. 2016, 43, 620–632. [Google Scholar] [CrossRef]
- Yazici, H.; Seyahi, E.; Hatemi, G.; Yazici, Y. Behçet Syndrome: A Contemporary View. Nat. Rev. Rheumatol. 2018, 14, 107–119. [Google Scholar] [CrossRef]
- Kural-Seyahi, E.; Fresko, I.; Seyahi, N.; Ozyazgan, Y.; Mat, C.; Hamuryudan, V.; Yurdakul, S.; Yazici, H. The Long-Term Mortality and Morbidity of Behçet Syndrome: A 2-Decade Outcome Survey of 387 Patients Followed at a Dedicated Center. Medicine 2003, 82, 60–76. [Google Scholar] [CrossRef] [PubMed]
- Tascilar, K.; Melikoglu, M.; Ugurlu, S.; Sut, N.; Caglar, E.; Yazici, H. Vascular Involvement in Behçet’s Syndrome: A Retrospective Analysis of Associations and the Time Course. Rheumatology 2014, 53, 2018–2022. [Google Scholar] [CrossRef] [PubMed]
- Koç, Y.; Güllü, I.; Akpek, G.; Akpolat, T.; Kansu, E.; Kiraz, S.; Batman, F.; Kansu, T.; Balkanci, F.; Akkaya, S. Vascular Involvement in Behçet’s Disease. J. Rheumatol. 1992, 19, 402–410. [Google Scholar] [PubMed]
- Seyahi, E.; Yurdakul, S. Behçet’s Syndrome and Thrombosis. Mediterr. J. Hematol. Infect. Dis. 2011, 3, e2011026. [Google Scholar] [CrossRef]
- Quehenberger, O.; Dennis, E.A. The Human Plasma Lipidome. N. Engl. J. Med. 2011, 365, 1812–1823. [Google Scholar] [CrossRef]
- Wenk, M.R. Lipidomics: New Tools and Applications. Cell 2010, 143, 888–895. [Google Scholar] [CrossRef]
- Ben-Fradj, M.K.; Naceur, I.; Talbi, E.; Wada, R.; Feki, O.; Smiti-Khanfir, M.; Feki, M. Altered Polyunsaturated Fatty Acids and Oxylipins Profile in Behçet’s Disease. Korean J. Intern. Med. 2025, 40, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Park, M.J.; Park, S.; Lee, E.-S.; Lee, D.Y. Integrative Metabolomics of Plasma and PBMCs Identifies Distinctive Metabolic Signatures in Behçet’s Disease. Arthritis Res. Ther. 2023, 25, 5. [Google Scholar] [CrossRef]
- Yücel, Ç.; Sertoğlu, E.; Omma, A.; Koçak, E.; Erdoğan Kablan, S.; Özgürtaş, T.; Nemutlu, E. Comparative Metabolomic Profiles of Vascular Involvement in Behçet’s Disease. Eur. J. Rheumatol. 2023, 10, 130–135. [Google Scholar] [CrossRef]
- International Study Group for Behçet’s Disease. Criteria for Diagnosis of Behcet’s Disease. Lancet 1990, 335, 1078–1080. [Google Scholar] [CrossRef]
- Hamuryudan, V.; Fresko, I.; Direskeneli, H.; Tenant, M.J.; Yurdakul, S.; Akoglu, T.; Yazici, H. Evaluation of the Turkish Translation of a Disease Activity Form for Behçet’s Syndrome. Rheumatology 1999, 38, 734–736. [Google Scholar] [CrossRef] [PubMed]
- Moral, K.; Koçak, E.; Kablan, S.E.; Nemutlu, E.; Şendur, H.N.; Cerit, M.; Yalçın, M.M.; Bilican, G.; Cindoruk, M.; Karakan, T.; et al. Hepatic and Metabolic Effects of SGLT2 Inhibitors in Type 2 Diabetes and MASLD Assessed by Metabolomic and Lipidomic Analysis. Sci. Rep. 2026. [Google Scholar] [CrossRef]
- Esim, O.; Adatepe, S.; Aksoy, O.A.; Goksel, B.A.; Yilmaz, T.; Baykal, B.; Yucel, Ç.; Kablan, S.E.; Eylem, C.C.; Kocak, E.; et al. Targeted Hepatic Delivery of Ezetimibe via Red Blood Cell-Coated Nanoparticles for the Treatment of Non-Alcoholic Fatty Liver Disease through Inflammation Modulation. Int. J. Pharm. 2025, 685, 126241. [Google Scholar] [CrossRef]
- van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane Lipids: Where They Are and How They Behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Wu, X.; Goudarzi, M.; Shi, J.; Song, W.; Li, C.; Liu, J.; Chen, H.; Zhang, X.; Zeng, X.; et al. Metabolomic Alterations Associated with Behçet’s Disease. Arthritis Res. Ther. 2018, 20, 214. [Google Scholar] [CrossRef] [PubMed]
- Braverman, N.E.; Moser, A.B. Functions of Plasmalogen Lipids in Health and Disease. Biochim. Biophys. Acta 2012, 1822, 1442–1452. [Google Scholar] [CrossRef]
- Wang, B.; Tontonoz, P. Phospholipid Remodeling in Physiology and Disease. Annu. Rev. Physiol. 2019, 81, 165–188. [Google Scholar] [CrossRef]
- Petkevicius, K.; Virtue, S.; Bidault, G.; Jenkins, B.; Çubuk, C.; Morgantini, C.; Aouadi, M.; Dopazo, J.; Serlie, M.; Koulman, A.; et al. Accelerated Phosphatidylcholine Turnover in Macrophages Promotes Adipose Tissue Inflammation in Obesity. eLife 2019, 8, e47990. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, P.; Li, F.; Xiao, X.; Zhang, Y.; Li, N.; Du, L.; Yang, P. Identification of the Immune-Related Biomarkers in Behcet’s Disease by Plasma Proteomic Analysis. Arthritis Res. Ther. 2023, 25, 92. [Google Scholar] [CrossRef]
- Carrasco, S.; Mérida, I. Diacylglycerol, When Simplicity Becomes Complex. Trends Biochem. Sci. 2007, 32, 27–36. [Google Scholar] [CrossRef]
- Mérida, I.; Ávila-Flores, A.; Merino, E. Diacylglycerol Kinases: At the Hub of Cell Signalling. Biochem. J. 2008, 409, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.G. Regulation of Phosphoinositide-Specific Phospholipase C. Annu. Rev. Biochem. 2001, 70, 281–312. [Google Scholar] [CrossRef] [PubMed]
- Thiele, M.; Suvitaival, T.; Trošt, K.; Kim, M.; de Zawadzki, A.; Kjaergaard, M.; Rasmussen, D.N.; Lindvig, K.P.; Israelsen, M.; Detlefsen, S.; et al. Sphingolipids Are Depleted in Alcohol-Related Liver Fibrosis. Gastroenterology 2023, 164, 1248–1260. [Google Scholar] [CrossRef] [PubMed]
- Chew, W.S.; Torta, F.; Ji, S.; Choi, H.; Begum, H.; Sim, X.; Khoo, C.M.; Khoo, E.Y.H.; Ong, W.-Y.; Van Dam, R.M.; et al. Large-Scale Lipidomics Identifies Associations between Plasma Sphingolipids and T2DM Incidence. JCI Insight 2019, 4, e126925. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Su, J.; Zhu, H.; Chen, Y.; Shukla, S.; Yu, L.; Wang, D.; Kang, D. The Protective Effect of Higher Serum TAG (51:4) Levels against Parkinson’s Disease. Br. J. Nutr. 2025, 133, 806–816. [Google Scholar] [CrossRef]
- Mendes-Frias, A.; Santos-Lima, B.; Furtado, D.Z.S.; Ruperez, F.J.; Assunção, N.A.; Matias, M.J.; Gomes, V.; Gaifem, J.; Barbas, C.; Castro, A.G.; et al. Dysregulation of Glycerophospholipid Metabolism during Behçet’s Disease Contributes to a pro-Inflammatory Phenotype of Circulating Monocytes. J. Transl. Autoimmun. 2020, 3, 100056. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Values |
|---|---|
| Age at diagnosis [years], mean ± SD | 32.3 ± 4.9 |
| Disease duration [months], median (IQR) | 94.8 (71.7) |
| Body mass index (BMI), mean ± SD | 23.3 ± 2.7 |
| Active disease, n (%) | 28 (58.3) |
| BDCAF, median (IQR) | 2.0 (0.0–3.0) |
| Disease symptoms in patient population, n (%) | |
| Oral ulcers | 48 (100) |
| Genital ulcers | 36 (75) |
| Erythema nodosum | 24 (50) |
| Papulopustular lesions | 28 (58.3) |
| Positive pathergy test | 12 (25) |
| Arthritis | 21 (43.7) |
| Uveitis | 24 (50) |
| Vascular involvement: | 18 (37.5) |
| Venous thrombosis | 16 (89) |
| Arterial aneurism | 1 (5.5) |
| Pulmonary aneurism | 1 (5.5) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Koçak, E.; Yücel, Ç.; Kablan, S.E.; Sertoğlu, E.; Özgürtaş, T.; Nemutlu, E.; Omma, A. Plasma Lipidomic Remodeling in Behçet’s Disease Reveals Alterations Associated with Vascular Involvement. Metabolites 2026, 16, 363. https://doi.org/10.3390/metabo16060363
Koçak E, Yücel Ç, Kablan SE, Sertoğlu E, Özgürtaş T, Nemutlu E, Omma A. Plasma Lipidomic Remodeling in Behçet’s Disease Reveals Alterations Associated with Vascular Involvement. Metabolites. 2026; 16(6):363. https://doi.org/10.3390/metabo16060363
Chicago/Turabian StyleKoçak, Engin, Çiğdem Yücel, Sevilay Erdoğan Kablan, Erdim Sertoğlu, Taner Özgürtaş, Emirhan Nemutlu, and Ahmet Omma. 2026. "Plasma Lipidomic Remodeling in Behçet’s Disease Reveals Alterations Associated with Vascular Involvement" Metabolites 16, no. 6: 363. https://doi.org/10.3390/metabo16060363
APA StyleKoçak, E., Yücel, Ç., Kablan, S. E., Sertoğlu, E., Özgürtaş, T., Nemutlu, E., & Omma, A. (2026). Plasma Lipidomic Remodeling in Behçet’s Disease Reveals Alterations Associated with Vascular Involvement. Metabolites, 16(6), 363. https://doi.org/10.3390/metabo16060363