

Short Working Memory Impairment Associated with Hippocampal Microglia Activation in Chronic Hepatic Encephalopathy

 , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
2.3. Induction of Type C HE
- -
- Group 1 (TAA): Male Wistar rats (n = 8, 162.48 ± 11.60 g), received repeated intraperitoneal injections (i.p) of TAA solution (100 mg TAA/kg. prepared in 0.9% NaCl), separated by a 24 h interval for 10 consecutive days. In order to minimize weight loss, hypoglycemia, dehydration and renal failure, which may arise from liver failure, a solution of dextrose (5%), NaCl (0.45%) and KCl20 mEq/L) was added to drinking water for 10 days;
- -
- Group 2 (Control): Male Wistar rats (n = 8, 162.03 g ± 10.28 g) received an equal volume of 0.9% sodium chloride solution (NaCl 0.9%).
2.4. Assessment of Liver Function
2.4.1. Biochemical Analysis
2.4.2. Liver Morphometric Analysis
2.4.3. Liver Histopathology
2.5. Short Working Memory Assessment
2.5.1. Novel Object Recognition Test
2.5.2. T-Maze Test
2.5.3. Y-Maze Test
2.6. Immunofluorescence
3. Statistical Analysis
4. Results
4.1. Assessment of Liver Function
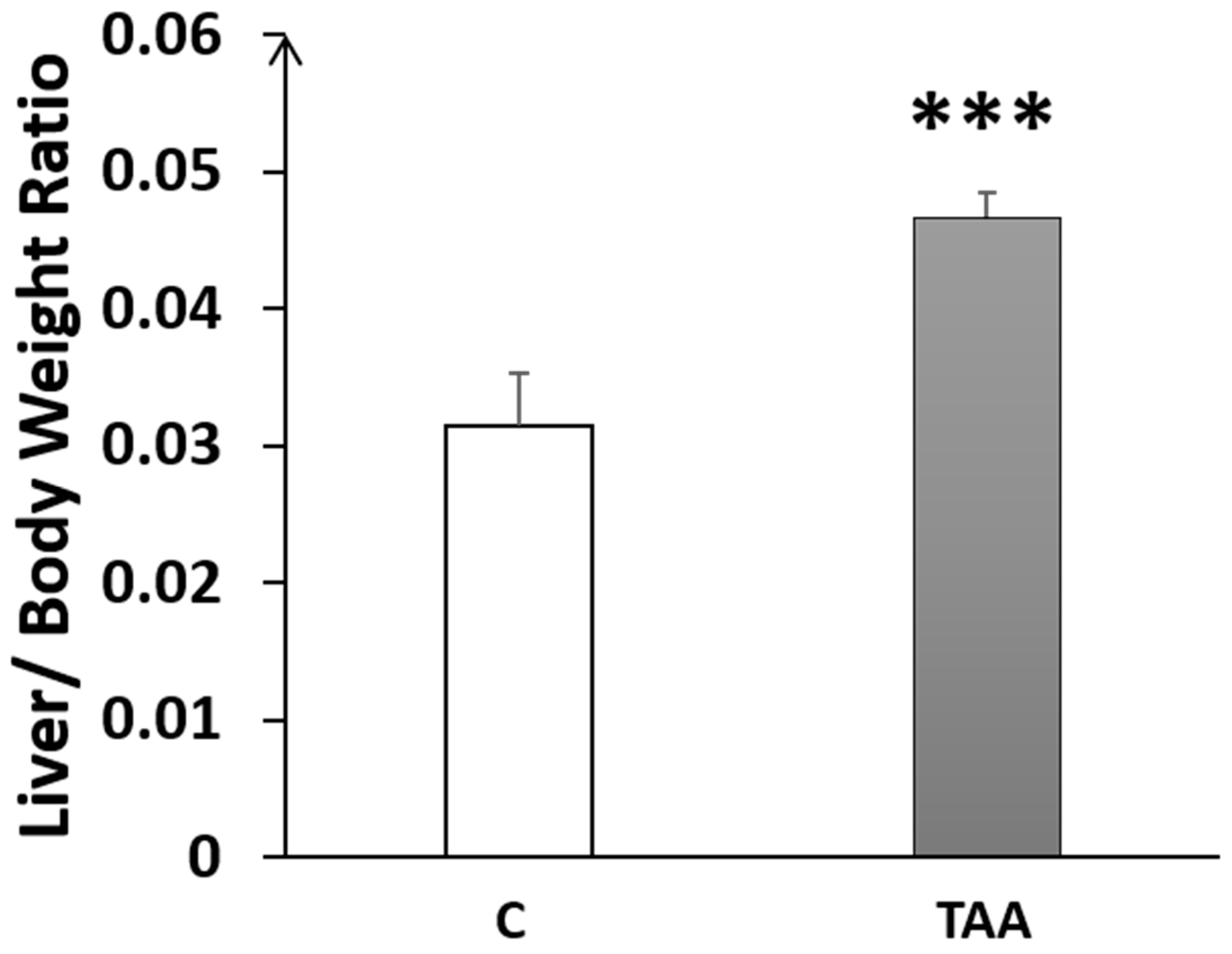
4.1.1. Liver Morphometry
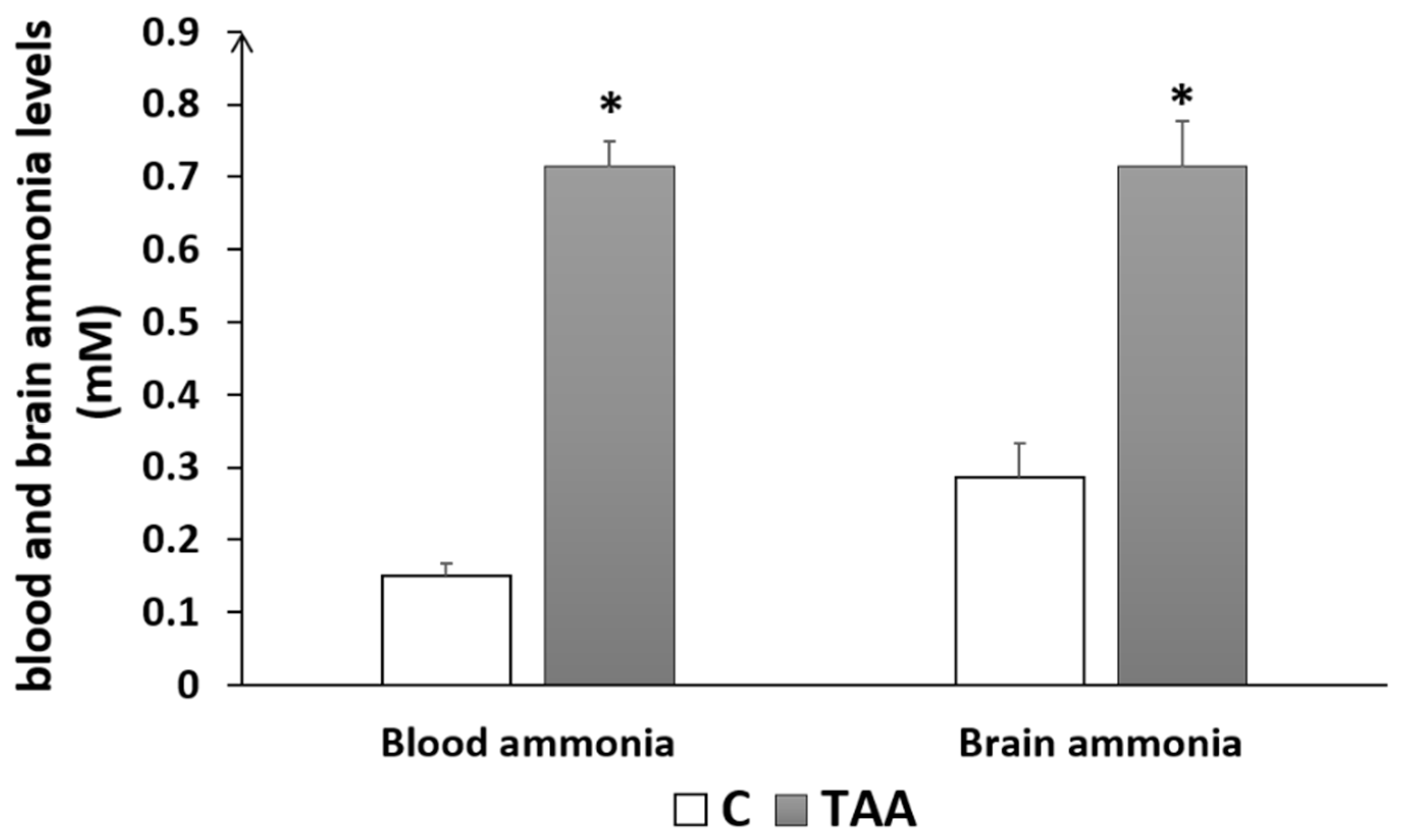
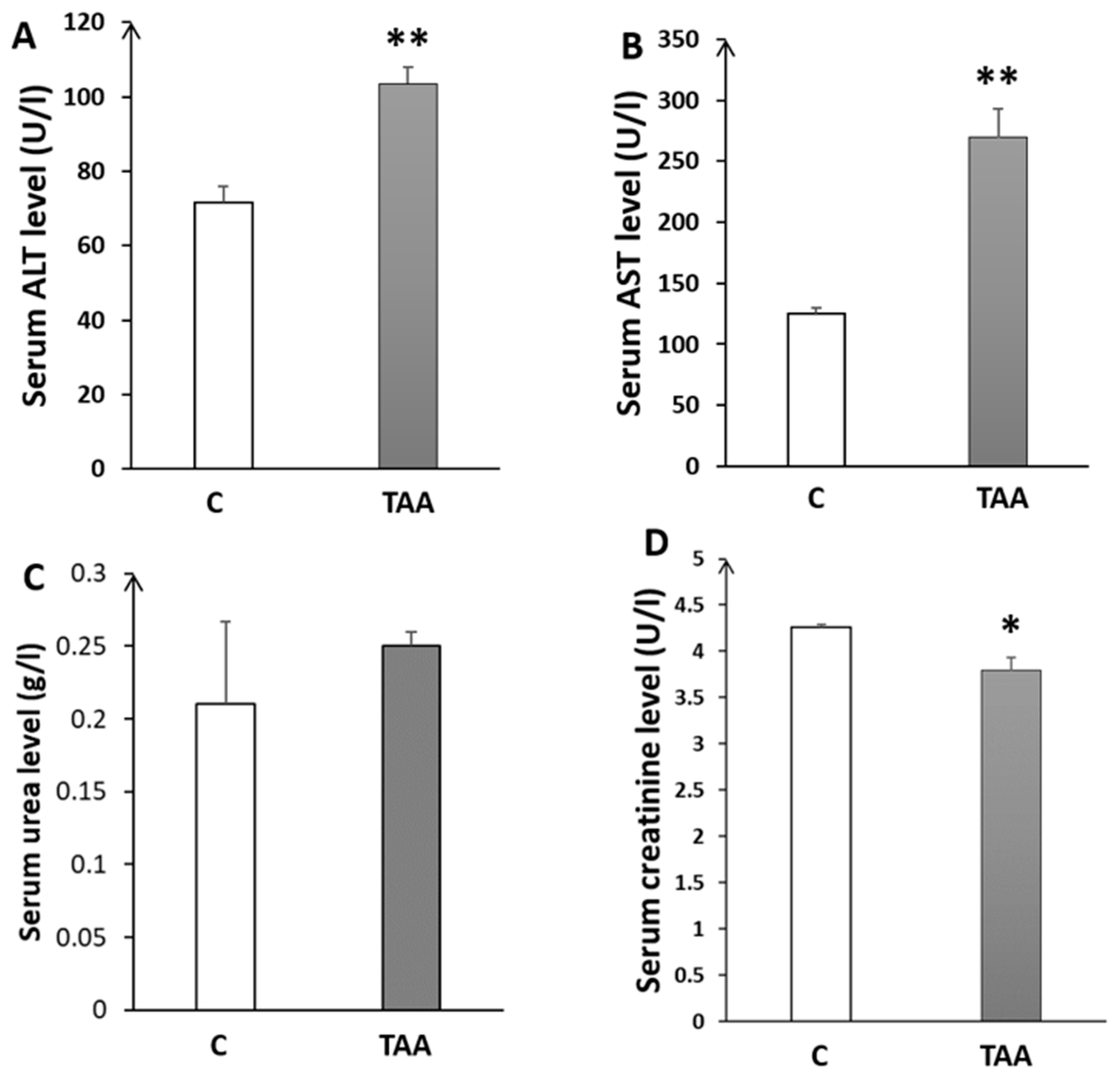
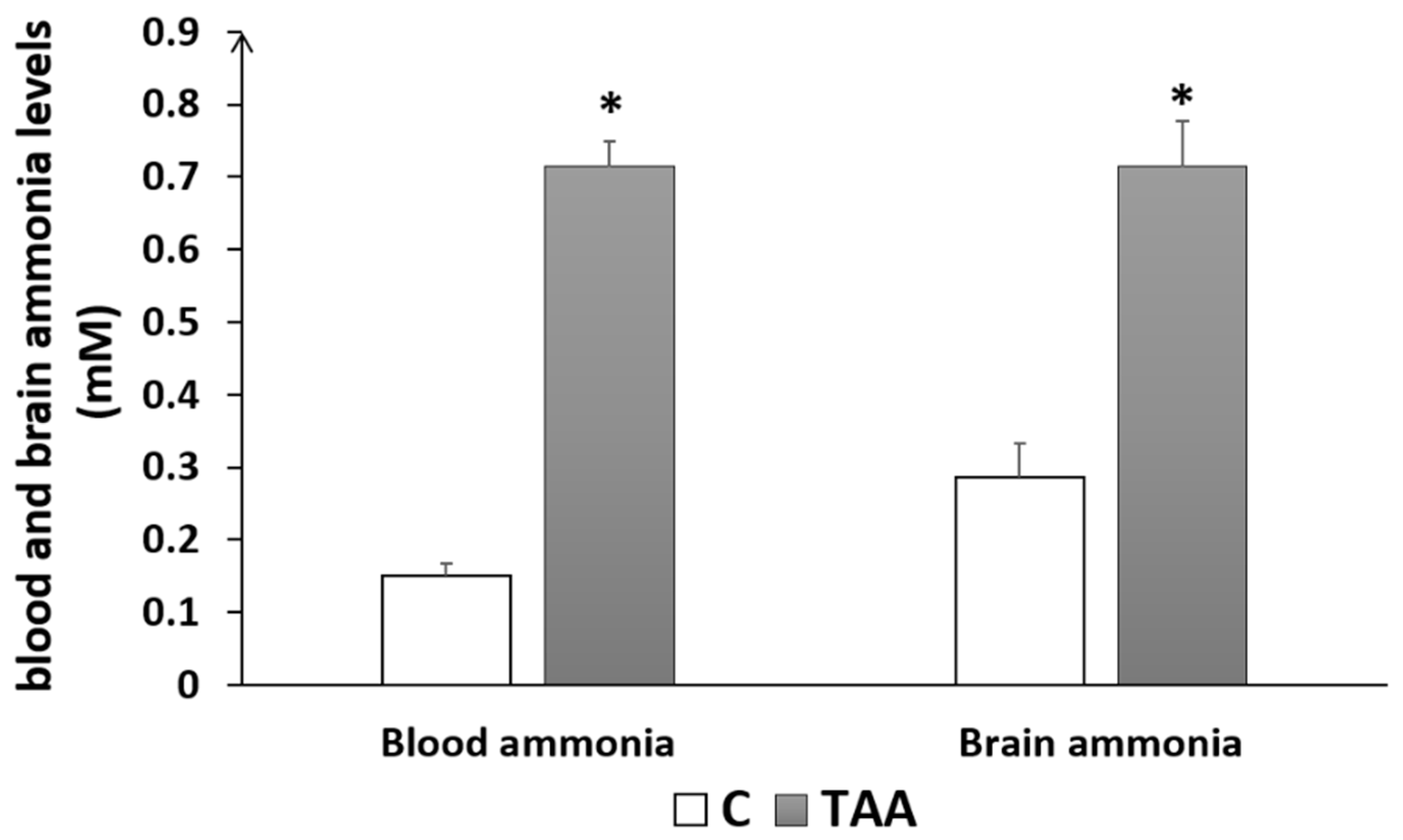
4.1.2. Biochemical Analysis
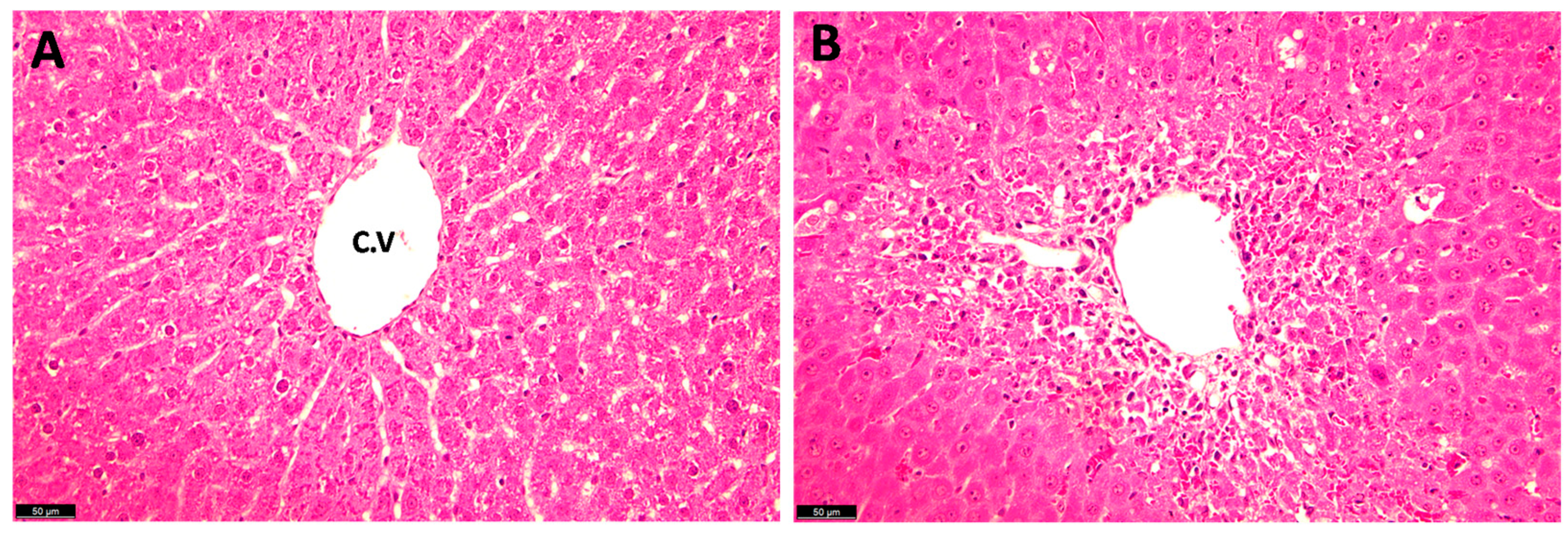
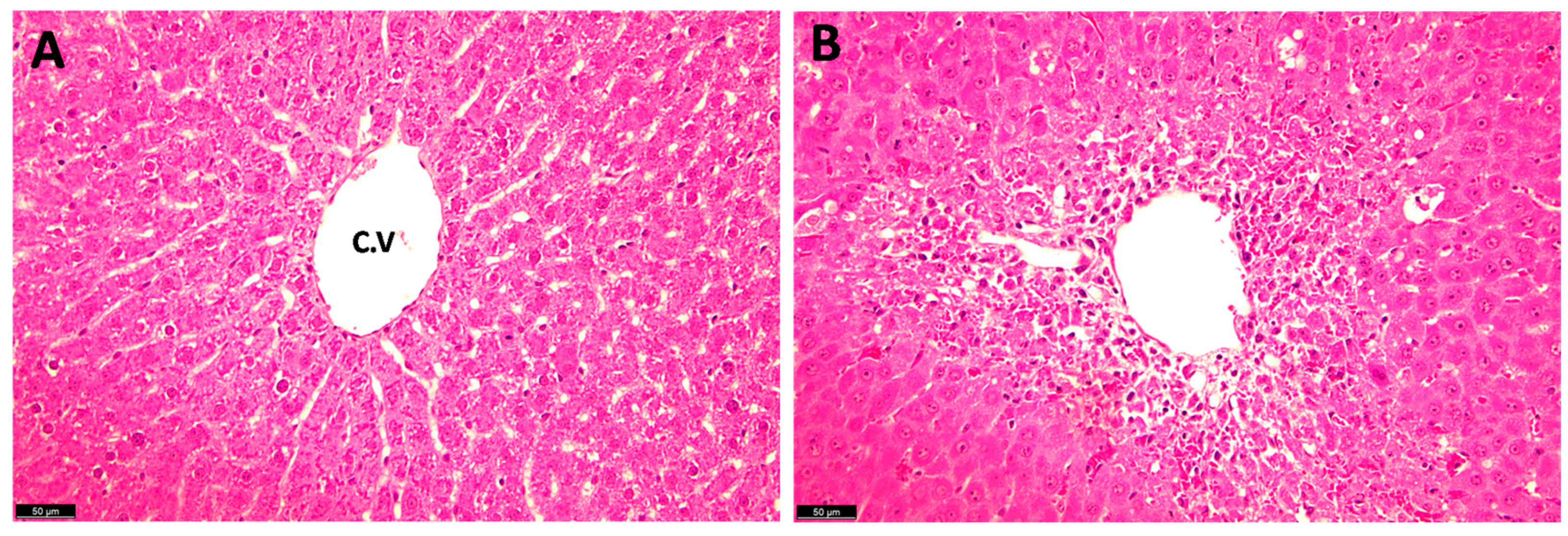
4.1.3. Histopathological Examination
4.2. Measurement of Working Memory Impairment
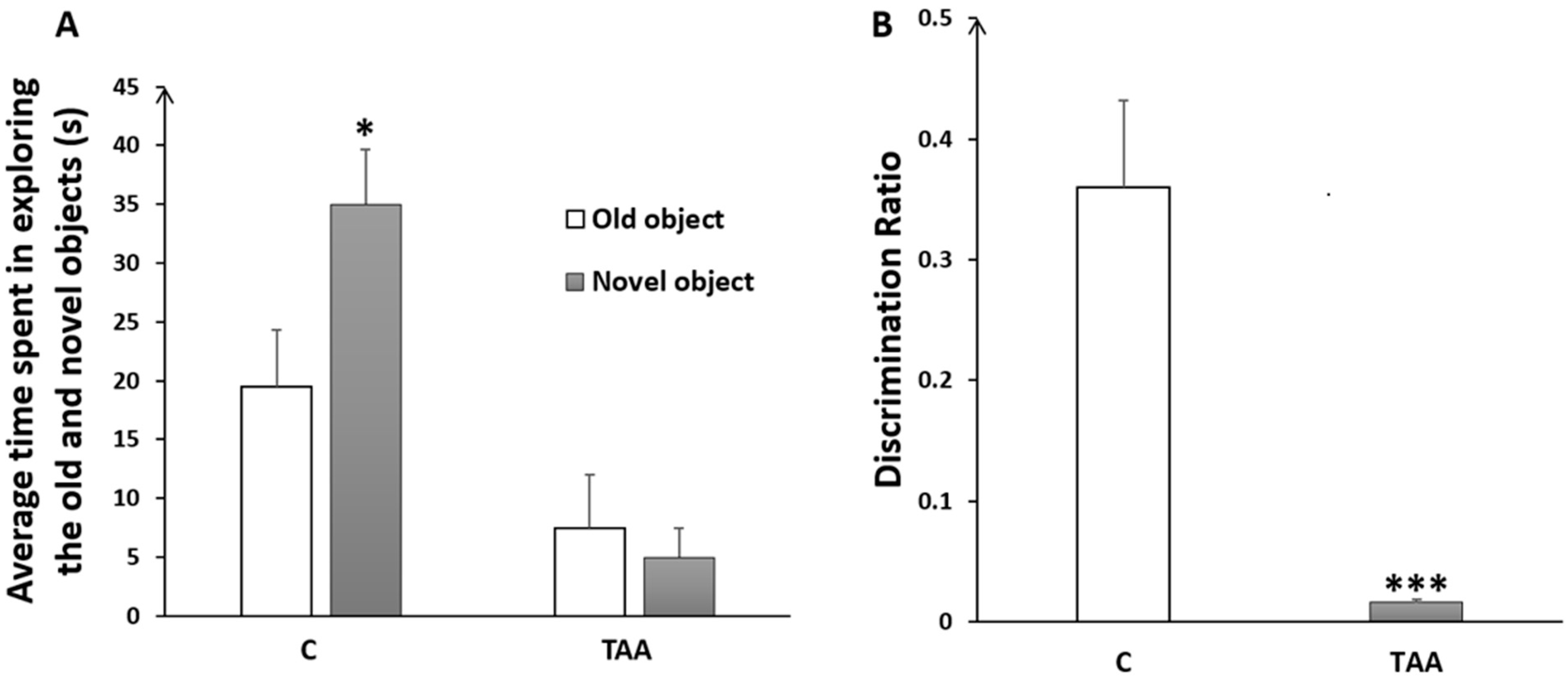
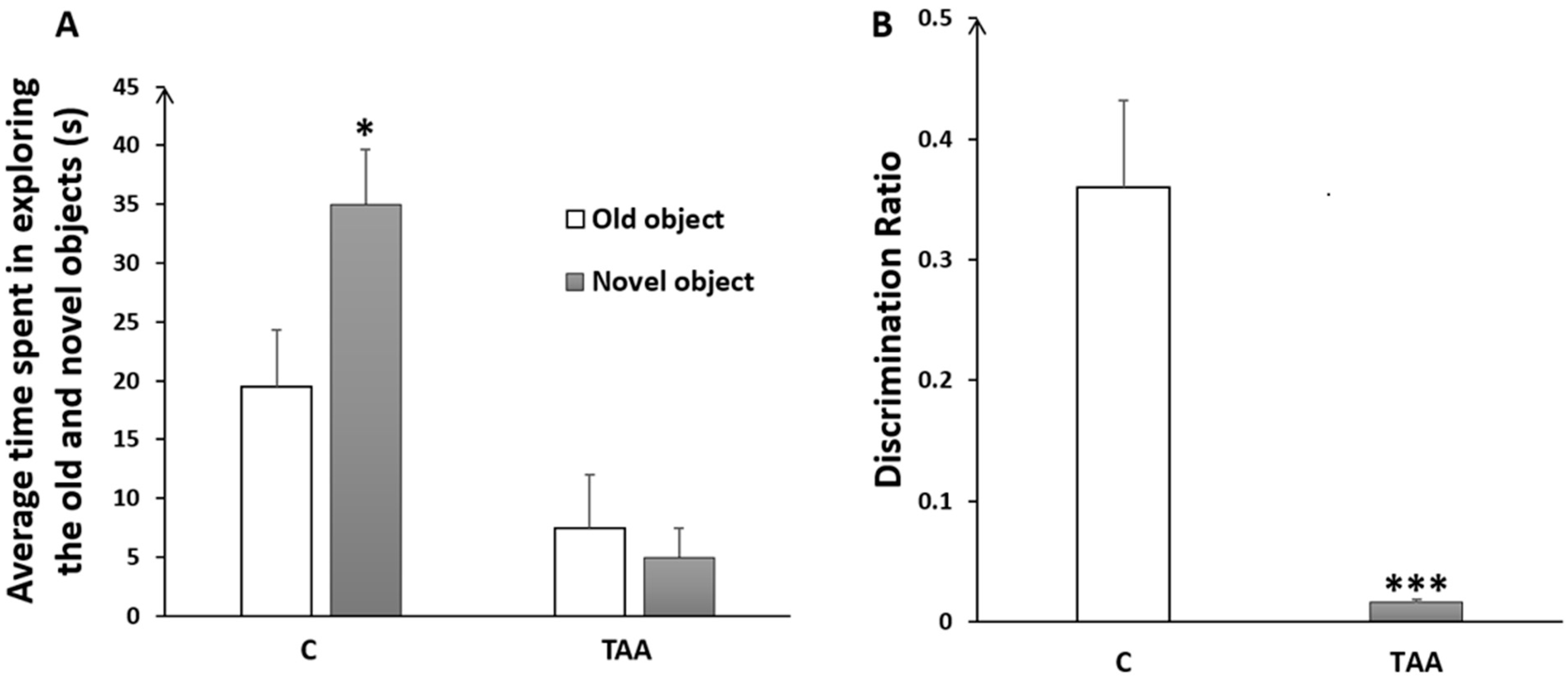
4.2.1. Memory Impairment as Measured by Novel Object Recognition Test
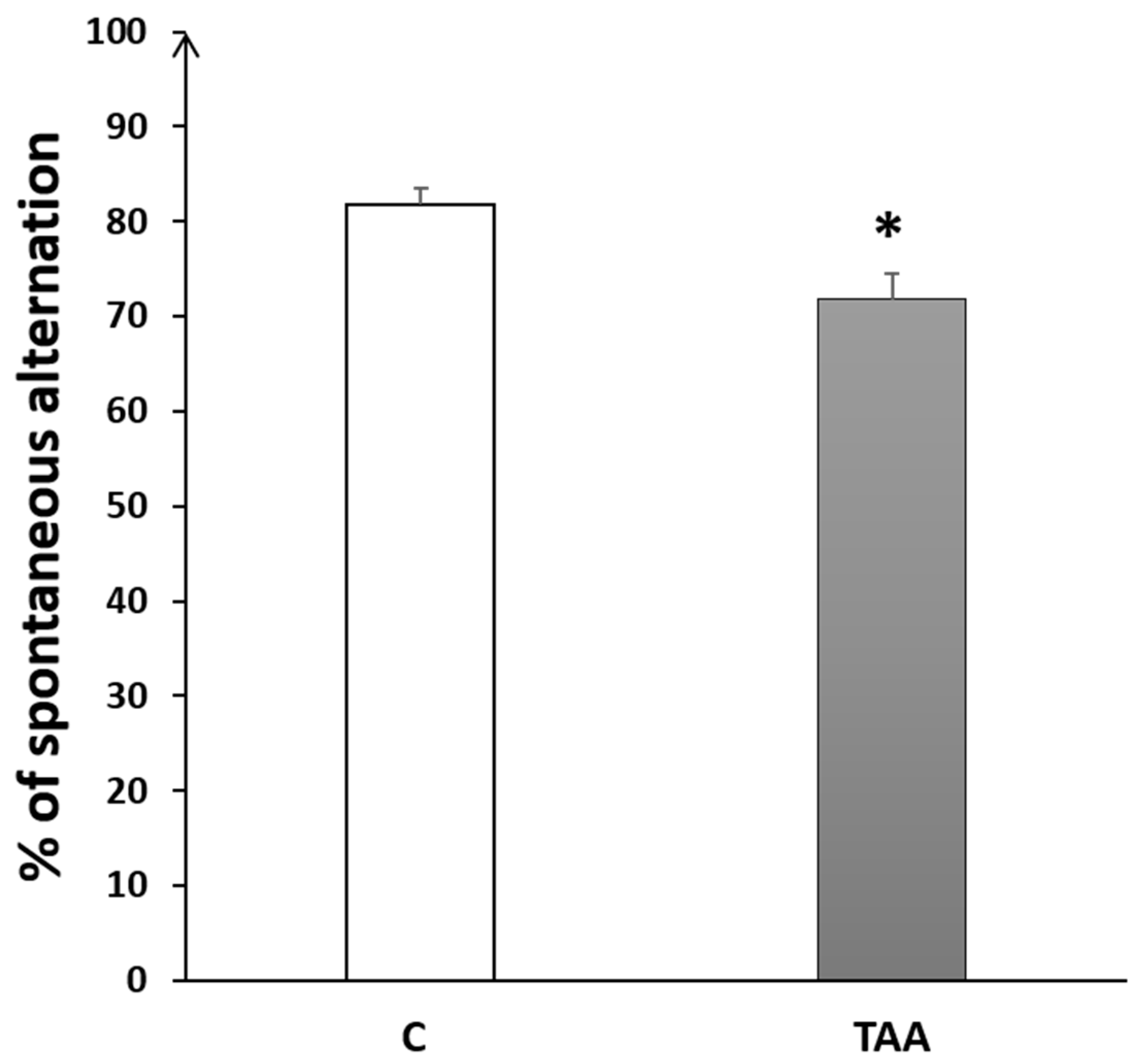
4.2.2. Memory Impairment as Measured by Y-Maze Test
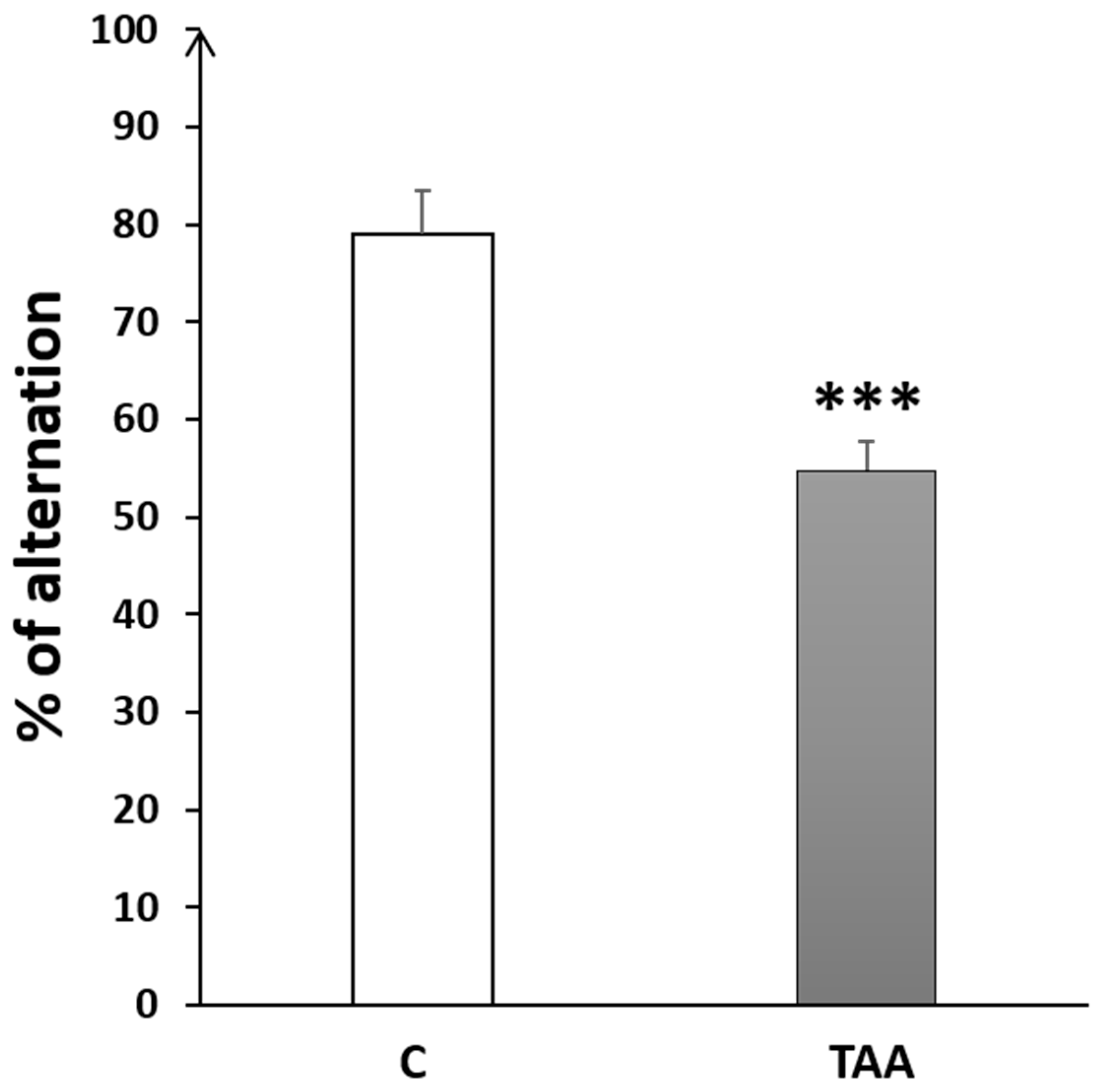
4.2.3. Memory Impairment as Measured by T-Maze Test
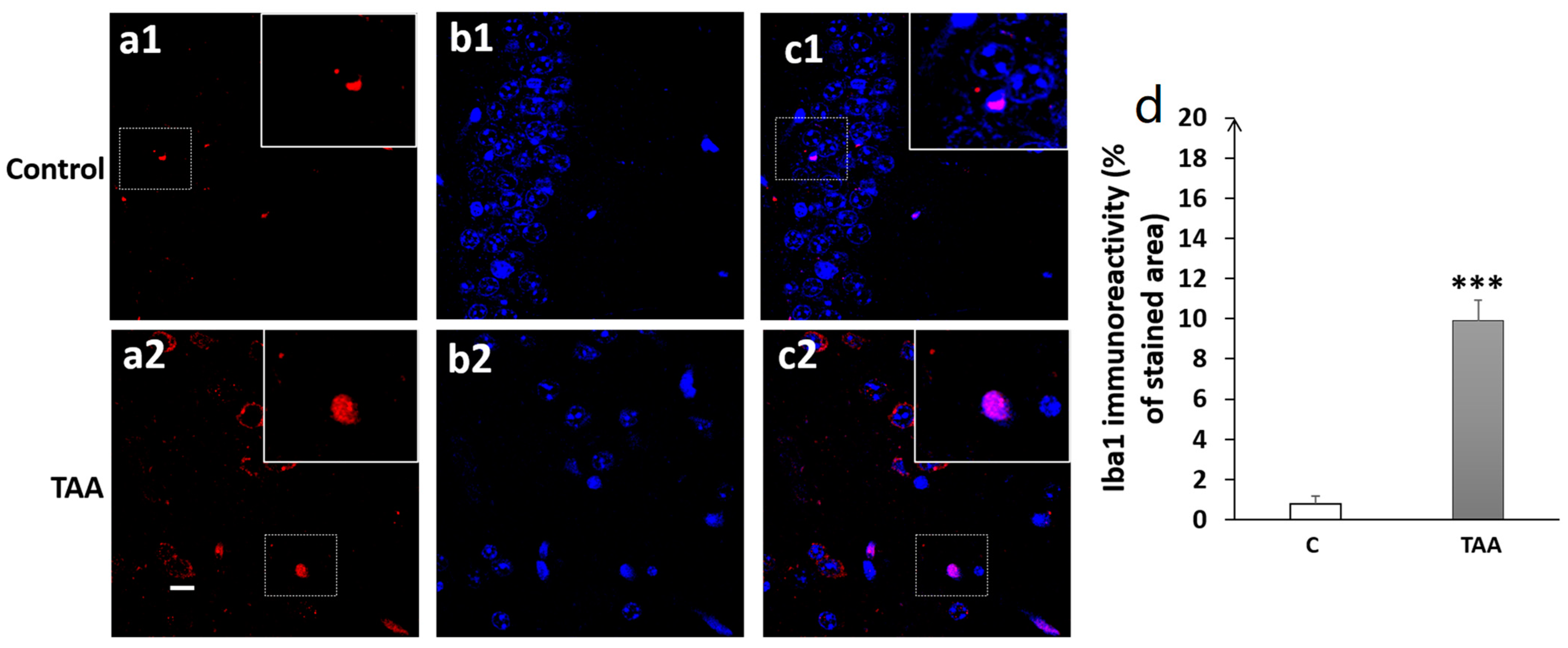
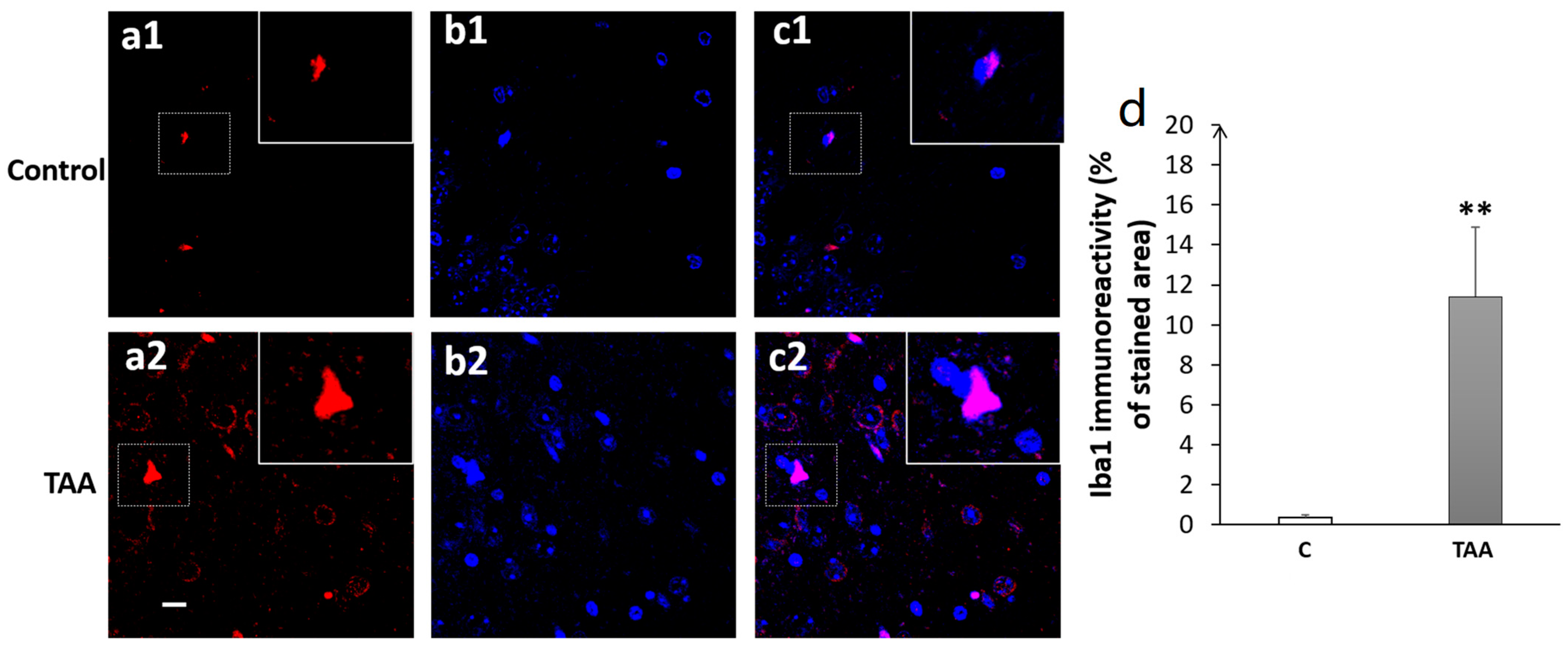
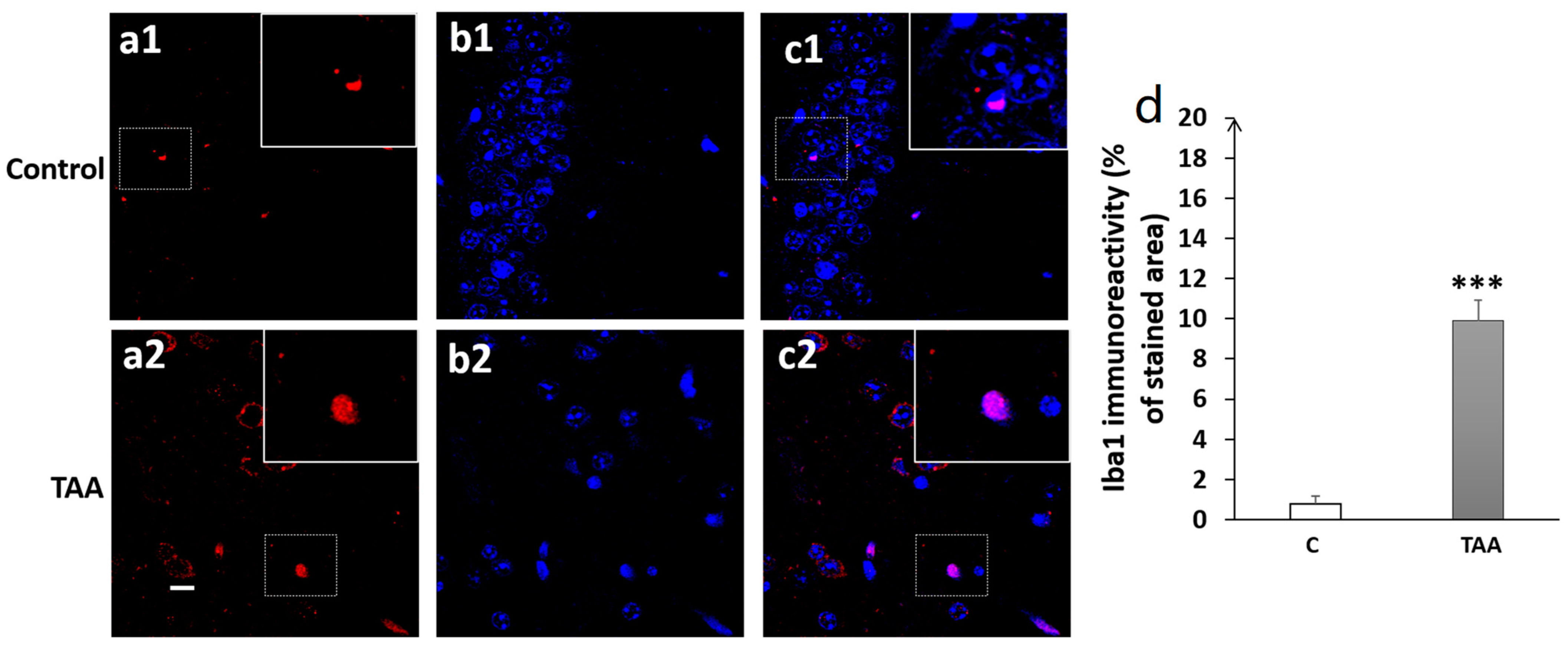
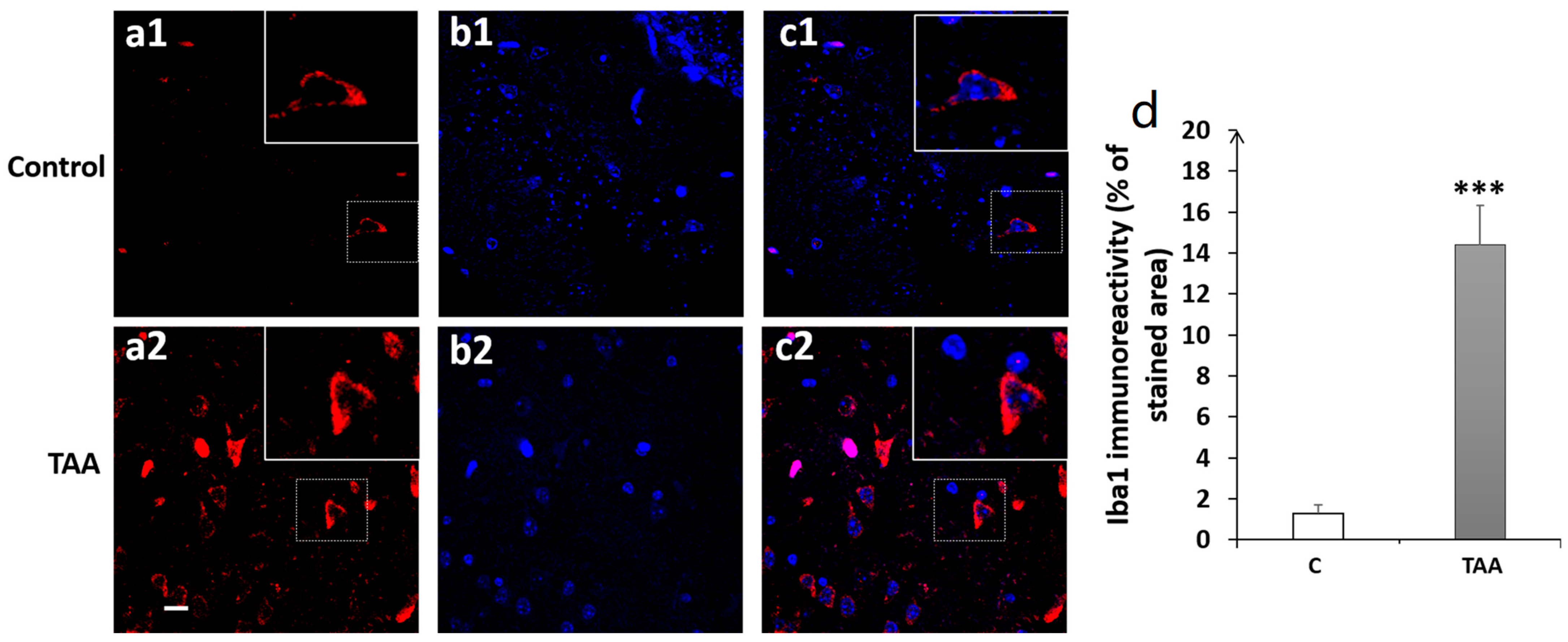
4.3. Immunofluorescence Study
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- El Hiba, O.; Elgot, A.; Ahboucha, S.; Gamrani, H. Differential regional responsiveness of astroglia in mild hepatic encephalopathy: An Immunohistochemical approach in bile duct ligated rat. Acta Histochem. 2016, 118, 338–346. [Google Scholar] [CrossRef] [PubMed]
- El Hiba, O.; Gamrani, H.; Ahboucha, S. Increased Reissner’s fiber material in the subcommissural organ and ventricular area in bile duct ligated rats. Acta Histochem. 2012, 114, 673–681. [Google Scholar] [CrossRef] [PubMed]
- El Hiba, O.; Gamrani, H.; Chatoui, H.; Ahboucha, S. Loss of tyrosine hydroxylase expression within the nigro-striato-cortical pathways in the cirrhotic rat: The possible restorative effect of the neurosteroid dehydroepiandrosterone sulfate. Acta Histochem. 2013, 115, 637–645. [Google Scholar] [CrossRef] [PubMed]
- El-Mansoury, B.; Smimih, K.; Aimrane, A.; Draoui, A.; Ferssiwi, A.; Bitar, A.; Jayakumar, A.R.; El Hiba, O. P8 Cognitive Impairment Associated With Cortical and Hippocampal Microglia Activation and Altered Synaptic Function in a Rat Model of Chronic Hepatic Encephalopathy. Off. J. Am. Coll. Gastroenterol. ACG 2023, 118, S7. [Google Scholar] [CrossRef]
- Méndez, M.; Méndez-Lopez, M.; Lopez, L.; Aller, M.A.; Arias, J.; Cimadevilla, J.M.; Arias, J.L. Spatial memory alterations in three models of hepatic encephalopathy. Behav. Brain Res. 2008, 188, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Coltart, I.; Tranah, T.H.; Shawcross, D.L. Inflammation and hepatic encephalopathy. Arch. Biochem. Biophys. 2013, 536, 189–196. [Google Scholar] [CrossRef]
- Jayakumar, A.R.; Rao, K.V.R.; Norenberg, M.D. Neuroinflammation in hepatic encephalopathy: Mechanistic aspects. J. Clin. Exp. Hepatol. 2015, 5, S21–S28. [Google Scholar] [CrossRef] [PubMed]
- Lier, J.; Winter, K.; Bleher, J.; Grammig, J.; Mueller, W.C.; Streit, W.; Bechmann, I. Loss of IBA1-Expression in brains from individuals with obesity and hepatic dysfunction. Brain Res. 2019, 1710, 220–229. [Google Scholar] [CrossRef]
- Thameem Dheen, S.; Kaur, C.; Ling, E.-A. Microglial Activation and its Implications in the Brain Diseases. Curr. Med. Chem. 2007, 14, 1189–1197. [Google Scholar] [CrossRef]
- Norden, D.M.; Trojanowski, P.J.; Villanueva, E.; Navarro, E.; Godbout, J.P. Sequential activation of microglia and astrocyte cytokine expression precedes increased Iba-1 or GFAP immunoreactivity following systemic immune challenge. Glia 2016, 64, 300–316. [Google Scholar] [CrossRef]
- Lull, M.E.; Block, M.L. Microglial activation and chronic neurodegeneration. Neurotherapeutics 2010, 7, 354–365. [Google Scholar] [CrossRef]
- Jiao, F.; Wang, Y.; Chen, Q.; Cao, P.; Shi, C.; Pei, M.; Wang, L.; Gong, Z. Role of SIRT1 in Hepatic Encephalopathy: In Vivo and In Vitro Studies Focusing on the NLRP3 Inflammasome. Oxid. Med. Cell. Longev. 2021, 2021, 5522708. [Google Scholar] [CrossRef] [PubMed]
- McMillin, M.; Grant, S.; Frampton, G.; Andry, S.; Brown, A.; DeMorrow, S. Fractalkine suppression during hepatic encephalopathy promotes neuroinflammation in mice. J. Neuroinflamm. 2016, 13, 198. [Google Scholar] [CrossRef] [PubMed]
- El-Mansoury, B.; El Hiba, O.; El Khiat, A.; Jayakumar, A.R. Hepatic Encephalopathy as a Gliopathy: The Mechanisms of Alzheimer Type II Astrocytosis. In Physiology and Function of Glial Cells in Health and Disease; IGI Global: Hershey, PA, USA, 2024; pp. 223–242. [Google Scholar]
- El Hiba, O.; El-Mansoury, B.; Norenberg, M.; Jayakumar, A.R. Astroglia and Acute Hepatic Encephalopathy: The Mechanisms of Cytotoxic Brain Edema. In Physiology and Function of Glial Cells in Health and Disease; IGI Global: Hershey, PA, USA, 2024; pp. 243–260. [Google Scholar]
- Cabrera-Pastor, A.; Llansola, M.; Montoliu, C.; Malaguarnera, M.; Balzano, T.; Taoro-Gonzalez, L.; García-García, R.; Mangas-Losada, A.; Izquierdo-Altarejos, P.; Arenas, Y.M. Peripheral inflammation induces neuroinflammation that alters neurotransmission and cognitive and motor function in hepatic encephalopathy: Underlying mechanisms and therapeutic implications. Acta Physiol. 2019, 226, e13270. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, R.; Cauli, O.; Gomez–Pinedo, U.; Agusti, A.; Hernandez–Rabaza, V.; Garcia–Verdugo, J.; Felipo, V. Hyperammonemia induces neuroinflammation that contributes to cognitive impairment in rats with hepatic encephalopathy. Gastroenterology 2010, 139, 675–684. [Google Scholar] [CrossRef]
- Butterworth, R.F. Hepatic encephalopathy: A central neuroinflammatory disorder? Hepatology 2011, 53, 1372–1376. [Google Scholar] [CrossRef] [PubMed]
- Dhanda, S.; Gupta, S.; Halder, A.; Sunkaria, A.; Sandhir, R. Systemic inflammation without gliosis mediates cognitive deficits through impaired BDNF expression in bile duct ligation model of hepatic encephalopathy. Brain Behav. Immun. 2018, 701, 214–232. [Google Scholar] [CrossRef]
- Jalan, R.; Shawcross, D.; Davies, N. The molecular pathogenesis of hepatic encephalopathy. Int. J. Biochem. Cell Biol. 2003, 35, 1175–1181. [Google Scholar]
- Chen, J.; Buchanan, J.B.; Sparkman, N.L.; Godbout, J.P.; Freund, G.G.; Johnson, R.W. Neuroinflammation and disruption in working memory in aged mice after acute stimulation of the peripheral innate immune system. Brain Behav. Immun. 2008, 22, 301–311. [Google Scholar] [CrossRef]
- Demorrow, S.; Cudalbu, C.; Davies, N.; Jayakumar, A.R.; Rose, C.F. 2021 ISHEN guidelines on animal models of hepatic encephalopathy. Liver Int. 2021, 41, 1474–1488. [Google Scholar] [CrossRef]
- Huang, X.; Yang, C.; Huang, M. Protective mechanism of the EZH2/microRNA-15a-5p/CXCL10 axis in rats with depressive-like behaviors. J. Chem. Neuroanat. 2023, 132, 102283. [Google Scholar] [CrossRef]
- El Khiat, A.; Tamegart, L.; Draoui, A.; El Fari, R.; Sellami, S.; Rais, H.; El Hiba, O.; Gamrani, H. Kinetic deterioration of short memory in rat with acute hepatic encephalopathy: Involvement of astroglial and neuronal dysfunctions. Behav. Brain Res. 2019, 367, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, R.; Hussain, H.; Ravelo, N.; Sriramajayam, K.; Di Gregorio, D.M.; Paulrasu, K.; Chen, P.; Young, K.; Masciarella, A.D.; Jayakumar, A.R. Kidney Damage in Long COVID: Studies in Experimental Mice. Biology 2023, 12, 1070. [Google Scholar] [CrossRef] [PubMed]
- Paidas, M.J.; Mohamed, A.B.; Norenberg, M.D.; Saad, A.; Barry, A.F.; Colon, C.; Kenyon, N.S.; Jayakumar, A.R. Multi-organ histopathological changes in a mouse hepatitis virus model of COVID-19. Viruses 2021, 13, 1703. [Google Scholar] [CrossRef] [PubMed]
- Bevins, R.A.; Besheer, J. Object recognition in rats and mice: A one-trial non-matching-to-sample learning task to study ‘recognition memory’. Nat. Protoc. 2006, 1, 1306–1311. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, V.S.; Santos, J.R.; Leão, A.H.F.F.; Medeiros, A.M.; Melo, T.G.; Izídio, G.S.; Cabral, A.; Ribeiro, R.A.; Abílio, V.C.; Ribeiro, A.M. Repeated treatment with a low dose of reserpine as a progressive model of Parkinson’s disease. Behav. Brain Res. 2012, 231, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Deacon, R.M.J.; Rawlins, J.N.P. T-maze alternation in the rodent. Nat. Protoc. 2006, 1, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Moreno, T.; Castillo, C.G.; Martínez-Serrano, A. Long term behavioral effects of functional dopaminergic neurons generated from human neural stem cells in the rat 6-OH-DA Parkinson’s disease model. Effects of the forced expression of BCL-XL. Behav. Brain Res. 2012, 232, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Bontempi, B.; Whelan, K.T.; Risbrough, V.B.; Lloyd, G.K.; Menzaghi, F. Cognitive enhancing properties and tolerability of cholinergic agents in mice: A comparative study of nicotine, donepezil, and SIB-1553A, a subtype-selective ligand for nicotinic acetylcholine receptors. Neuropsychopharmacology 2003, 28, 1235–1246. [Google Scholar] [CrossRef]
- Draoui, A.; El Hiba, O.; Abbaoui, A.; El Fari, R.; Aitihya, M.; Gamrani, H. Differential impairment of short working and spatial memories in a rat model of progressive Parkinson’s disease onset: A focus on the prodromal stage. Brain Res. Bull. 2019, 150, 307–316. [Google Scholar] [CrossRef]
- Fukuda, T.; Sato, Y.; Takiguchi, M.; Yamamoto, T.; Murasawa, H.; Pawlak, A.; Kobayashi, H.; Tamano, H.; Takeda, A. Dehydroeffusol Rescues Amyloid β25–35-Induced Spatial Working Memory Deficit. Plant Foods Hum. Nutr. 2020, 75, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Naolapo, O.J.O.; Naolapo, A.Y.O.; Josiah, T.; Osaku, M.; Akanji, O.O.; Abiodun, O.R. Elevated plus maze and Y maze behavioural effects of Monosodium glutamate in Swiss albinoo mice Elevated Plus Maze and Y-Maze Behavioral Effects of Subchronic, Oral Low Dose Monosodium Glutamate in Swiss Albino Mice. J. pharm. Biol. Sci. 2016, 3, 21–27. [Google Scholar]
- Jayakumar, A.R.; Tong, X.Y.; Shamaladevi, N.; Barcelona, S.; Gaidosh, G.; Agarwal, A.; Norenberg, M.D. Defective synthesis and release of astrocytic thrombospondin-1 mediates the neuronal TDP-43 proteinopathy, resulting in defects in neuronal integrity associated with chronic traumatic encephalopathy: In vitro studies. J. Neurochem. 2017, 140, 645–661. [Google Scholar] [CrossRef] [PubMed]
- Jayakumar, A.R.; Tong, X.Y.; Curtis, K.M.; Ruiz-Cordero, R.; Shamaladevi, N.; Abuzamel, M.; Johnstone, J.; Gaidosh, G.; Rama Rao, K.V.; Norenberg, M.D. Decreased astrocytic thrombospondin-1 secretion after chronic ammonia treatment reduces the level of synaptic proteins: In vitro and in vivo studies. J. Neurochem. 2014, 131, 333–347. [Google Scholar] [CrossRef]
- Paidas, M.J.; Cosio, D.S.; Ali, S.; Kenyon, N.S.; Jayakumar, A.R. Long-Term Sequelae of COVID-19 in Experimental Mice. Mol. Neurobiol. 2022, 59, 5970–5986. [Google Scholar] [CrossRef] [PubMed]
- Fede, G.; D’Amico, G.; Arvaniti, V.; Tsochatzis, E.; Germani, G.; Georgiadis, D.; Morabito, A.; Burroughs, A.K. Renal failure and cirrhosis: A systematic review of mortality and prognosis. J. Hepatol. 2012, 56, 810–818. [Google Scholar] [CrossRef]
- Schmandra, T.C.; Bauer, H.; Petrowsky, H.; Herrmann, G.; Encke, A.; Hanisch, E. Effect of fibrin glue occlusion of the hepatobiliary tract on thioacetamide-induced liver failure. Am. J. Surg. 2001, 182, 58–63. [Google Scholar] [CrossRef]
- Singh, S.; Trigun, S.K. Activation of neuronal nitric oxide synthase in cerebellum of chronic hepatic encephalopathy rats is associated with up-regulation of NADPH-producing pathway. Cerebellum 2010, 9, 384–397. [Google Scholar] [CrossRef]
- Abdelghffar, E.A.R.; El-Nashar, H.A.S.; Fayez, S.; Obaid, W.A.; Eldahshan, O.A. Ameliorative effect of oregano (Origanum vulgare) versus silymarin in experimentally induced hepatic encephalopathy. Sci. Rep. 2022, 12, 17854. [Google Scholar] [CrossRef]
- Dejong, C.H.C.; Deutz, N.E.P.; Soeters, P.B. Cerebral cortex ammonia and glutamine metabolism in two rat models of chronic liver insufficiency-induced hyperammonemia: Influence of pair-feeding. J. Neurochem. 1993, 60, 1047–1057. [Google Scholar] [CrossRef]
- Blei, A.T.; Olafsson, S.; Therrien, G.; Butterworth, R.F. Ammonia-induced brain edema and intracranial hypertension in rats after portacaval anastomosis. Hepatology 1994, 19, 1437–1444. [Google Scholar] [CrossRef]
- Butterworth, R.F.; Girard, G.; Giguère, J. Regional differences in the capacity for ammonia removal by brain following portocaval anastomosis. J. Neurochem. 1988, 51, 486–490. [Google Scholar] [CrossRef] [PubMed]
- Pinzani, M.; Rombouts, K. Liver fibrosis: From the bench to clinical targets. Dig. Liver Dis. 2004, 36, 231–242. [Google Scholar] [CrossRef]
- Méndez, M.; Méndez-López, M.; López, L.; Aller, M.A.; Arias, J.; Arias, J.L. Working memory impairment and reduced hippocampal and prefrontal cortex c-Fos expression in a rat model of cirrhosis. Physiol. Behav. 2008, 95, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Wesierska, M.; Klinowska, H.D.; Adamska, I.; Fresko, I.; Sadowska, J.; Albrecht, J. Cognitive flexibility but not cognitive coordination is affected in rats with toxic liver failure. Behav. Brain Res. 2006, 171, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tan, S.; Du, J.; Chen, Y.; Jia, J.; Feng, J.; Liu, K. International Immunopharmacology Dexmedetomidine alleviates neuroinflammation, restores sleep disorders and neurobehavioral abnormalities in rats with minimal hepatic encephalopathy. Int. Immunopharmacol. 2021, 96, 107795. [Google Scholar] [CrossRef]
- Leke, R.; Oliveira, D.L.; Forgiarini, L.F.; Escobar, T.D.C.; Hammes, T.O.; Meyer, F.S.; Keiding, S.; Silveira, T.R.; Schousboe, A. Impairment of short term memory in rats with hepatic encephalopathy due to bile duct ligation. Metab. Brain Dis. 2013, 28, 187–192. [Google Scholar] [CrossRef]
- Dhanda, S.; Sandhir, R. Role of dopaminergic and serotonergic neurotransmitters in behavioral alterations observed in rodent model of hepatic encephalopathy. Behav. Brain Res. 2015, 286, 222–235. [Google Scholar] [CrossRef]
- Chen, J.R.; Wang, B.N.; Tseng, G.F.; Wang, Y.J.; Huang, Y.S.; Wang, T.J. Morphological changes of cortical pyramidal neurons in hepatic encephalopathy. BMC Neurosci. 2014, 15, 15. [Google Scholar] [CrossRef]
- García-Ayllón, M.-S.; Cauli, O.; Silveyra, M.-X.; Rodrigo, R.; Candela, A.; Compañ, A.; Jover, R.; Pérez-Mateo, M.; Martínez, S.; Felipo, V. Brain cholinergic impairment in liver failure. Brain 2008, 131, 2946–2956. [Google Scholar] [CrossRef]
- Méndez, M.; Méndez-López, M.; López, L.; Aller, M.Á.; Arias, J.; Arias, J.L. Associative learning deficit in two experimental models of hepatic encephalopathy. Behav. Brain Res. 2009, 198, 346–351. [Google Scholar] [CrossRef]
- Cauli, O.; Rodrigo, R.; Piedrafita, B.; Boix, J.; Felipo, V. Inflammation and hepatic encephalopathy: Ibuprofen restores learning ability in rats with portacaval shunts. Hepatology 2007, 46, 514–519. [Google Scholar] [CrossRef] [PubMed]
- Agusti, A.; Cauli, O.; Rodrigo, R.; Llansola, M.; Hernández-Rabaza, V.; Felipo, V. p38 MAP kinase is a therapeutic target for hepatic encephalopathy in rats with portacaval shunts. Gut 2011, 60, 1572–1579. [Google Scholar] [CrossRef]
- Monfort, P.; Erceg, S.; Piedrafita, B.; Llansola, M.; Felipo, V. Chronic liver failure in rats impairs glutamatergic synaptic transmission and long-term potentiation in hippocampus and learning ability. Eur. J. Neurosci. 2007, 25, 2103–2111. [Google Scholar] [CrossRef] [PubMed]
- Zemtsova, I.; Görg, B.; Keitel, V.; Bidmon, H.-J.; Schrör, K.; Häussinger, D. Microglia activation in hepatic encephalopathy in rats and humans. Hepatology 2011, 54, 204–215. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.A.K.; Sharifi, Y.; Newman, T.A.; Davies, N.; Vairappan, B.; Perry, H.V.; Jalan, R. Characterisation of temporal microglia and astrocyte immune responses in bile duct-ligated rat models of cirrhosis. Liver Int. 2014, 34, 1184–1191. [Google Scholar] [CrossRef]
- Dadsetan, S.; Balzano, T.; Forteza, J.; Cabrera-Pastor, A.; Taoro-Gonzalez, L.; Hernandez-Rabaza, V.; Gil-Perotín, S.; Cubas-Núñez, L.; García-Verdugo, J.-M.; Agusti, A. Reducing peripheral inflammation with infliximab reduces neuroinflammation and improves cognition in rats with hepatic encephalopathy. Front. Mol. Neurosci. 2016, 9, 106. [Google Scholar] [CrossRef]
- Hernandez-Rabaza, V.; Agusti, A.; Cabrera-Pastor, A.; Fustero, S.; Delgado, O.; Taoro-Gonzalez, L.; Montoliu, C.; Llansola, M.; Felipo, V. Sildenafil reduces neuroinflammation and restores spatial learning in rats with hepatic encephalopathy: Underlying mechanisms. J. Neuroinflamm. 2015, 12, 195. [Google Scholar] [CrossRef]
- Hernández-Rabaza, V.; Cabrera-Pastor, A.; Taoro-González, L.; Malaguarnera, M.; Agustí, A.; Llansola, M.; Felipo, V. Hyperammonemia induces glial activation, neuroinflammation and alters neurotransmitter receptors in hippocampus, impairing spatial learning: Reversal by sulforaphane. J. Neuroinflamm. 2016, 13, 41. [Google Scholar] [CrossRef]
- Wilson, C.J.; Finch, C.E.; Cohen, H.J. Cytokines and cognition—The case for a head-to-toe inflammatory paradigm. J. Am. Geriatr. Soc. 2002, 50, 2041–2056. [Google Scholar] [CrossRef]
- Boitard, C.; Cavaroc, A.; Sauvant, J.; Aubert, A.; Castanon, N.; Layé, S.; Ferreira, G. Impairment of hippocampal-dependent memory induced by juvenile high-fat diet intake is associated with enhanced hippocampal inflammation in rats. Brain Behav. Immun. 2014, 40, 9–17. [Google Scholar] [CrossRef]
- Bellinger, F.P.; Madamba, S.; Siggins, G.R. Interleukin 1β inhibits synaptic strength and long-term potentiation in the rat CA1 hippocampus. Brain Res. 1993, 628, 227–234. [Google Scholar] [CrossRef]
- Cunningham, A.J.; Murray, C.A.; O’neill, L.A.J.; Lynch, M.A.; O’connor, J.J. Interleukin-1β (IL-1β) and tumour necrosis factor (TNF) inhibit long-term potentiation in the rat dentate gyrus in vitro. Neurosci. Lett. 1996, 203, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Tancredi, V.; D’Antuono, M.; Cafè, C.; Giovedì, S.; Buè, M.C.; D’Arcangelo, G.; Onofri, F.; Benfenati, F. The inhibitory effects of interleukin-6 on synaptic plasticity in the rat hippocampus are associated with an inhibition of mitogen-activated protein kinase ERK. J. Neurochem. 2000, 75, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Hellstrom, I.C.; Danik, M.; Luheshi, G.N.; Williams, S. Chronic LPS exposure produces changes in intrinsic membrane properties and a sustained IL-β-dependent increase in GABAergic inhibition in hippocampal CA1 pyramidal neurons. Hippocampus 2005, 15, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Shaw, K.N.; Commins, S.; O’Mara, S.M. Cyclooxygenase inhibition attenuates endotoxin-induced spatial learning deficits, but not an endotoxin-induced blockade of long-term potentiation. Brain Res. 2005, 1038, 231–237. [Google Scholar] [CrossRef]
- Perry, V.H.; Cunningham, C.; Holmes, C. Systemic infections and inflammation affect chronic neurodegeneration. Nat. Rev. Immunol. 2007, 7, 161–167. [Google Scholar] [CrossRef]
- Rosi, S.; Ramirez-Amaya, V.; Vazdarjanova, A.; Worley, P.F.; Barnes, C.A.; Wenk, G.L. Neuroinflammation alters the hippocampal pattern of behaviorally induced Arc expression. J. Neurosci. 2005, 25, 723–731. [Google Scholar] [CrossRef]
- Tanaka, S.; Ide, M.; Shibutani, T.; Ohtaki, H.; Numazawa, S.; Shioda, S.; Yoshida, T. Lipopolysaccharide-induced microglial activation induces learning and memory deficits without neuronal cell deathin rats. J. Neurosci. Res. 2006, 83, 557–566. [Google Scholar] [CrossRef]
- Palin, K.; Bluthé, R.-M.; Verrier, D.; Tridon, V.; Dantzer, R.; Lestage, J. Interleukin-1β mediates the memory impairment associated with a delayed type hypersensitivity response to bacillus Calmette–Guérin in the rat hippocampus. Brain Behav. Immun. 2004, 18, 223–230. [Google Scholar] [CrossRef]
- Leone, P.; Mincheva, G.; Balzano, T.; Malaguarnera, M.; Felipo, V.; Llansola, M. Rifaximin Improves Spatial Learning and Memory Impairment in Rats with Liver Damage-Associated Neuroinflammation. Biomedicines 2022, 10, 1263. [Google Scholar] [CrossRef]
- Taoro-González, L.; Cabrera-Pastor, A.; Sancho-Alonso, M.; Arenas, Y.M.; Meseguer-Estornell, F.; Balzano, T.; ElMlili, N.; Felipo, V. Differential role of interleukin-1β in neuroinflammation-induced impairment of spatial and nonspatial memory in hyperammonemic rats. FASEB J. 2019, 33, 9913–9928. [Google Scholar] [CrossRef] [PubMed]
- Balzano, T.; Dadsetan, S.; Forteza, J.; Cabrera-Pastor, A.; Taoro-Gonzalez, L.; Malaguarnera, M.; Gil-Perotin, S.; Cubas-Nuñez, L.; Casanova, B.; Castro-Quintas, A. Chronic hyperammonemia induces peripheral inflammation that leads to cognitive impairment in rats: Reversed by anti-TNF-α treatment. J. Hepatol. 2020, 73, 582–592. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | TAA | |
|---|---|---|
| ApoA1 | 4.2 ± 0.4 µg/mL | 0.25 ± 0.06 µg/mL *** |
| α-2-Macroglobulin | 14.2 ± 2.3 ng/mL | 56.3 ± 5.9 ng/mL *** |
| Hyaluronicacid | 41.8 ± 5.9 ng/mL | 142.2 ± 13.8 ng/mL *** |
| Procollagen III amino terminal peptide | 17.8 ± 4.1 ng/mL | 28.1 ± 2.6 ng/mL * |
| Tissue inhibitor of metalloproteinase 1 | 0.56 ± 0.08 ng/mL | 1.8 ± 0.2 ng/mL *** |
| Matrix metalloproteinase-3 | 162.7 ± 16.7 ng/mL | 96.4 ± 12.8 ng/mL ** |
| Matrix metalloproteinase-9 | 52.3 ± 6.9 ng/mL | 36.4 ± 2.1 ng/mL * |
| YKL40 | 1.2 ± 0.2 ng/mL | 2.7 ± 0.7 ng/mL * |
| CTGF | 98.2 ± 11.6 ng/mL | 178.5 ± 36.9 ng/mL ** |
| Paraoxonase-1 | 3.8 ± 0.5 µg/mL | 1.8 ± 0.4 µg/mL *** |
| TGF BETA | 4.1 ± 1.2 pg/mg protein | 11.2 ± 3.1 pg/mg protein ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Mansoury, B.; Smimih, K.; El Khiat, A.; Draoui, A.; Aimrane, A.; Chatoui, R.; Ferssiwi, A.; Bitar, A.; Gamrani, H.; Jayakumar, A.R.; et al. Short Working Memory Impairment Associated with Hippocampal Microglia Activation in Chronic Hepatic Encephalopathy. Metabolites 2024, 14, 193. https://doi.org/10.3390/metabo14040193
El-Mansoury B, Smimih K, El Khiat A, Draoui A, Aimrane A, Chatoui R, Ferssiwi A, Bitar A, Gamrani H, Jayakumar AR, et al. Short Working Memory Impairment Associated with Hippocampal Microglia Activation in Chronic Hepatic Encephalopathy. Metabolites. 2024; 14(4):193. https://doi.org/10.3390/metabo14040193
Chicago/Turabian StyleEl-Mansoury, Bilal, Kamal Smimih, Abdelaati El Khiat, Ahmed Draoui, Abdelmohcine Aimrane, Redouane Chatoui, Abdesslam Ferssiwi, Abdelali Bitar, Halima Gamrani, Arumugam R. Jayakumar, and et al. 2024. "Short Working Memory Impairment Associated with Hippocampal Microglia Activation in Chronic Hepatic Encephalopathy" Metabolites 14, no. 4: 193. https://doi.org/10.3390/metabo14040193
APA StyleEl-Mansoury, B., Smimih, K., El Khiat, A., Draoui, A., Aimrane, A., Chatoui, R., Ferssiwi, A., Bitar, A., Gamrani, H., Jayakumar, A. R., & El Hiba, O. (2024). Short Working Memory Impairment Associated with Hippocampal Microglia Activation in Chronic Hepatic Encephalopathy. Metabolites, 14(4), 193. https://doi.org/10.3390/metabo14040193