

Exogenous γ-Aminobutyric Acid (GABA) Enhanced Response to Abiotic Stress in Hypsizygus marmoreus by Improving Mycelial Growth and Antioxidant Capacity

 and
and
Abstract
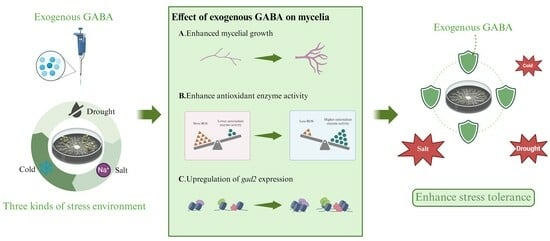
1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Strains
2.1.2. Culture Medium
2.2. Methods
2.2.1. Determination of Mycelial Growth Rate
2.2.2. Quality Inspection of the Fresh Aerial Mycelia
2.2.3. Microscopic Observation of the Mycelia
2.2.4. Determination of the GABA Content
2.2.5. Determination of Antioxidant Enzyme Activity
2.2.6. Determination of gad2 Expression
2.3. Data Analysis
3. Results and Analysis
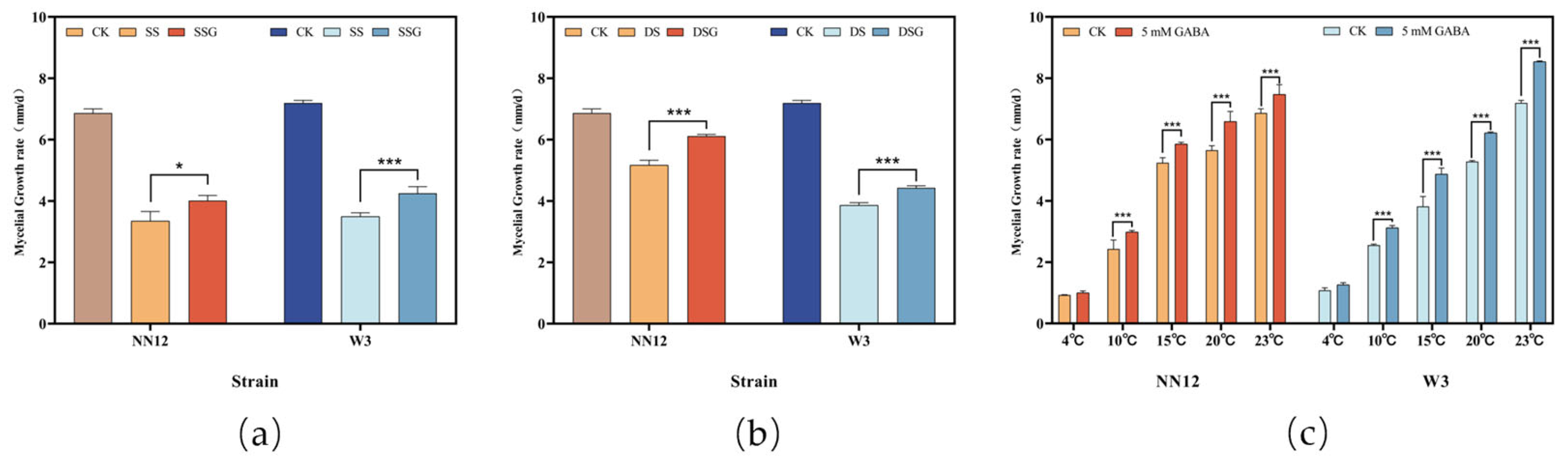
3.1. Effect of Exogenous GABA or Abiotic Stress Factors on Mycelial Growth under Standard Culture Conditions
3.2. Effect of Exogenous GABA on Mycelial Growth under Stress
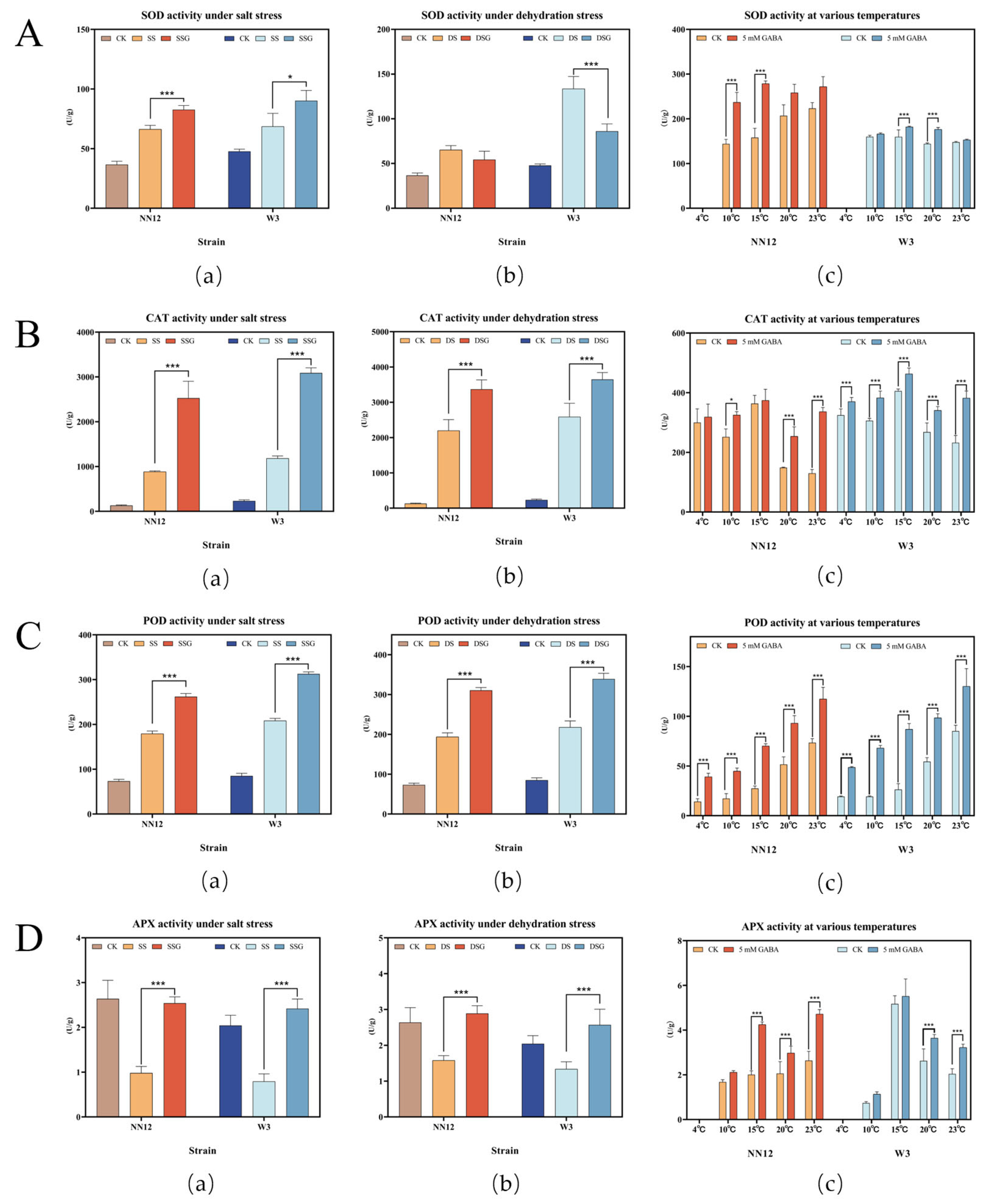
3.3. Effect of Exogenous GABA on Mycelial Antioxidant Capacity under Stress
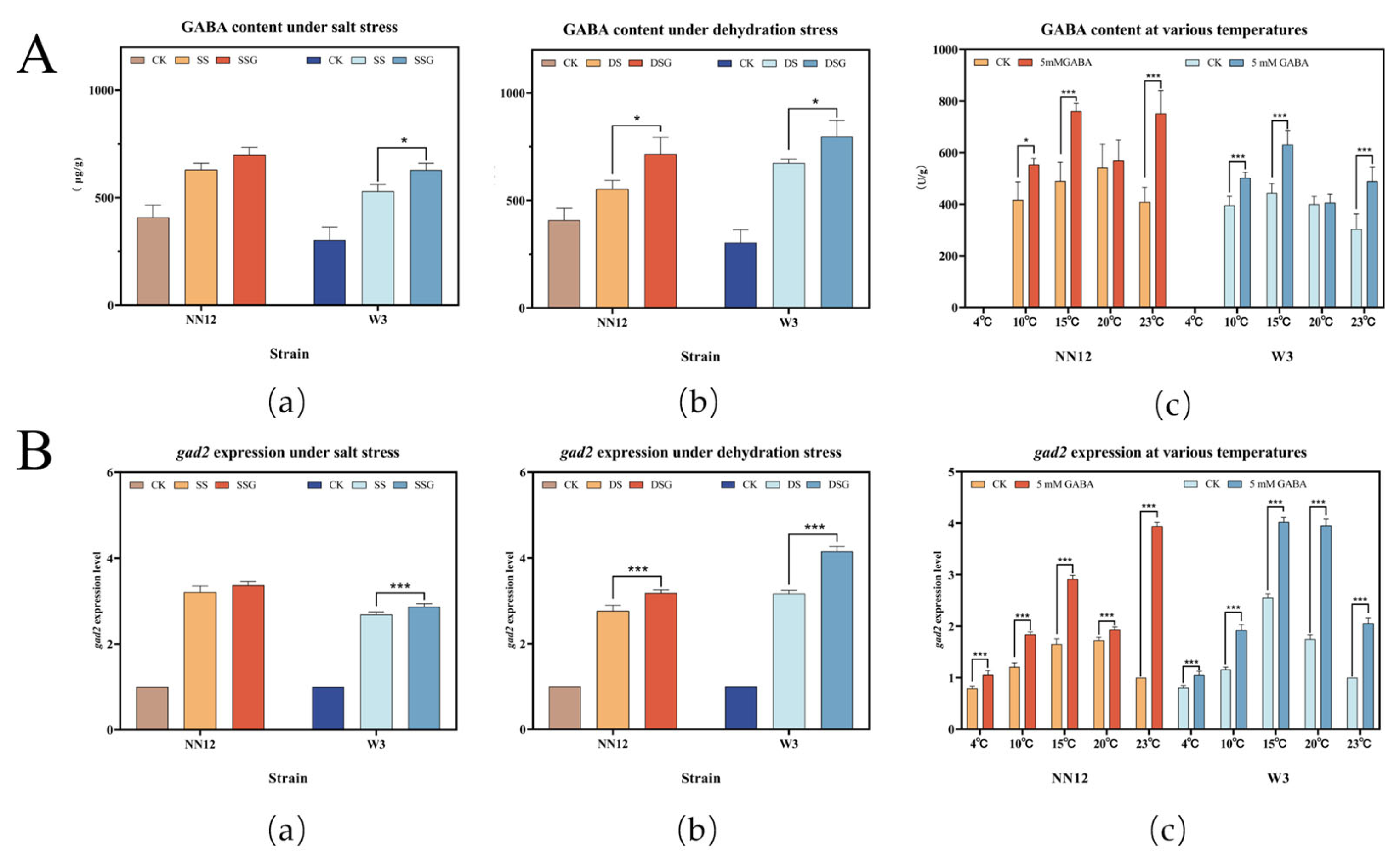
3.4. Effects of Exogenous GABA on Endogenous GABA Content and gad2 Expression Levels in Mycelia under Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Steward, F.C.; Thompson, J.F.; Dent, C.E. γ-Aminobutyric acid: A constituent of the potato tuber? Science 1949, 54, 447–451. [Google Scholar]
- Roberts, E.; Frankel, S. gamma-Aminobutyric acid in brain: Its formation from glutamic acid. J. Biol. Chem. 1950, 187, 55–63. [Google Scholar] [CrossRef]
- Awapara, J.; Landua, A.J.; Fuerst, R.; Seale, B. Free gamma-aminobutyric acid in brain. J. Biol. Chem. 1950, 187, 35–39. [Google Scholar] [CrossRef]
- Tafet, G.E.; Nemeroff, C.B. Pharmacological Treatment of Anxiety Disorders: The Role of the HPA Axis. Front. Psychiatry 2020, 11, 443. [Google Scholar] [CrossRef]
- Hayakawa, K.; Kimura, M.; Kamata, K. Mechanism underlying γ-aminobutyric acid-induced antihypertensive effect in spontaneously hypertensive rats. Eur. J. Pharmacol. 2002, 438, 107–113. [Google Scholar] [CrossRef]
- Yang, N.C.; Jhou, K.Y.; Tseng, C.Y. Antihypertensive effect of mulberry leaf aqueous extract containing γ-aminobutyric acid in spontaneously hypertensive rats. Food Chem. 2012, 132, 1796–1801. [Google Scholar] [CrossRef]
- Huang, D.; Wang, Y.; Thompson, J.W.; Yin, T.; Alexander, P.B.; Qin, D.; Mudgal, P.; Wu, H.; Liang, Y.; Tan, L.; et al. Cancer-cell-derived GABA promotes β-catenin-mediated tumour growth and immunosuppression. Nat. Cell Biol. 2022, 24, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Opolski, A.; Mazurkiewicz, M.; Wietrzyk, J.; Kleinrok, Z.; Radzikowski, C. The role of GABA-ergic system in human mammary gland pathology and in growth of transplantable murine mammary cancer. J. Exp. Clin. Cancer Res. 2000, 19, 383–390. [Google Scholar]
- Song, L.; Du, A.; Xiong, Y.; Jiang, J.; Zhang, Y.; Tian, Z.; Yan, H. γ-Aminobutyric acid inhibits the proliferation and increases oxaliplatin sensitivity in human colon cancer cells. Tumor Biol. 2016, 37, 14885–14894. [Google Scholar] [CrossRef] [PubMed]
- Fava, G.; Marucci, L.; Glaser, S.; Francis, H.; De Morrow, S.; Benedetti, A.; Alvaro, D.; Venter, J.; Meininger, C.; Patel, T.; et al. γ-Aminobutyric acid inhibits cholangiocarcinoma growth by cyclic AMP-dependent regulation of the protein kinase A/extracellular signal-regulated kinase 1/2 pathway. Cancer Res. 2005, 65, 11437–11446. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhao, M.; Han, Y.; Zhang, H. GABAergic Inhibitory Interneuron Deficits in Alzheimer’s Disease: Implications for Treatment. Front. Neurosci. 2020, 14, 660. [Google Scholar] [CrossRef]
- van van Hugte, E.J.H.; Schubert, D.; Kasri, N.N. Excitatory/inhibitory balance in epilepsies and neurodevelopmental disorders: Depolarizing γ-aminobutyric acid as a common mechanism. Epilepsia 2023, 64, 1975–1990. [Google Scholar] [CrossRef]
- Arnold, L.A.; Forkuo, G.S.; Nieman, A.N.; Yu, O.B.; Guthrie, M.L.; Yuan, N.Y.; Kodali, R.; Jahan, R.; Emala, C.W.; Cook, J.M.; et al. A New Pharmacological Approach for Asthma through Tissue-Specific Modulation of the GABA(A) Receptor. J. Allergy Clin. Immunol. 2016, 137, AB393. [Google Scholar] [CrossRef]
- Hagan, D.W.; Ferreira, S.M.; Santos, G.J.; Phelps, E.A. The role of GABA in islet function. Front. Endocrinol. 2022, 13, 972115. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, S.A.; Tyerman, S.D.; Gilliham, M.; Xu, B. γ-Aminobutyric acid (GABA) signalling in plants. Cell. Mol. Life Sci. 2017, 74, 1577–1603. [Google Scholar] [CrossRef]
- Breitkreuz, K.E.; Shelp, B.J.; Fischer, W.N.; Schwacke, R.; Rentsch, D. Identification and characterization of GABA, proline and quaternary ammonium compound transporters from Arabidopsis thaliana. FEBS Lett. 1999, 450, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Shi, S.Q.; Chen, W.; Xie, T.T.; Du, C.J.; Sun, J.C.; Shi, Z.; Gao, R.F.; Jiang, Z.P.; Xiao, W.F. Effects of exogenous γ-Aminobutyric acid on the regulation of respiration and protein expression in germinating seeds of mungbean (Vigna radiata) under salt conditions. Electron. J. Biotechnol. 2020, 47, 1–9. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Y.Z.; Ma, K.K.; Bai, M.M.; Liu, Y.Y.; Yu, X.J. Physiological changes associated with enhanced cold resistance during Medicago ruthenica germination and seedling growth in response to exogenous γ-aminobutyric acid. Grassl. Sci. 2023, 69, 120–131. [Google Scholar] [CrossRef]
- White, J.P.; Prell, J.; Ramachandran, V.K.; Poole, P.S. Characterization of a γ-Aminobutyric Acid Transport System of Rhizobium leguminosarum bv. viciae 3841. J. Bacteriol. 2009, 191, 1547–1555. [Google Scholar] [CrossRef]
- Hijaz, F.; Nehela, Y.; Killiny, N. Application of gamma-aminobutyric acid increased the level of phytohormones in Citrus sinensis. Planta 2018, 248, 909–918. [Google Scholar] [CrossRef]
- Mazzucotelli, E.; Tartari, A.; Cattivelli, L.; Forlani, G. Metabolism of γ-aminobutyric acid during cold acclimation and freezing and its relationship to frost tolerance in barley and wheat. J. Exp. Bot. 2006, 57, 3755–3766. [Google Scholar] [CrossRef]
- Li, Z.; Yu, J.; Peng, Y.; Huang, B. Metabolic pathways regulated by γ-aminobutyric acid (GABA) contributing to heat tolerance in creeping bentgrass (Agrostis stolonifera). Sci. Rep. 2016, 6, 30338. [Google Scholar] [CrossRef]
- Wang, Y.; Xiong, F.; Nong, S.; Liao, J.; Xing, A.; Shen, Q.; Ma, Y.; Fang, W.; Zhu, X. Effects of nitric oxide on the GABA, polyamines, and proline in tea (Camellia sinensis) roots under cold stress. Sci. Rep. 2020, 10, 12240. [Google Scholar] [CrossRef]
- Yang, R.; Guo, Q.; Gu, Z. GABA shunt and polyamine degradation pathway on γ-aminobutyric acid accumulation in germinating fava bean (Vicia faba L.) under hypoxia. Food Chem. 2013, 136, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Lu, X.Y.; Zong, H.; Zhuge, B. γ-aminobutyric acid accumulation enhances the cell growth of Candida glycerinogenes under hyperosmotic conditions. J. Gen. Appl. Microbiol. 2018, 64, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Feng, D.; Gao, Q.; Sun, X.; Ning, S.; Qi, N.; Hua, Z.; Tang, J. Effects of foliage-applied exogenous γ-aminobutyric acid on seedling growth of two rice varieties under salt stress. PLoS ONE 2023, 18, e0281846. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, N.; Liu, C.L.; Wang, H.T.; Li, Y.X.; Xie, Y.M.; Ma, F.W.; Liang, J.K.; Li, C.Y. Exogenous GABA improves the resistance of apple seedlings to long-term drought stress by enhancing GABA shunt and secondary cell wall biosynthesis. Tree Physiol. 2022, 42, 2563–2577. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, X.; Peng, C.; Shi, H.; Yang, J.; He, M.; Zhang, M.; Zhou, Y.; Duan, L. Exogenous Gamma-aminobutyric Acid Coordinates Active Oxygen and Amino Acid Homeostasis to Enhance Heat Tolerance in Wheat Seedlings. J. Plant Growth Regul. 2022, 41, 2787–2797. [Google Scholar] [CrossRef]
- Cheng, B.; Li, Z.; Liang, L.; Cao, Y.; Zeng, W.; Zhang, X.; Ma, X.; Huang, L.; Nie, G.; Liu, W.; et al. The γ-Aminobutyric Acid (GABA) Alleviates Salt Stress Damage during Seeds Germination of White Clover Associated with Na+/K+ Transportation, Dehydrins Accumulation, and Stress-Related Genes Expression in White Clover. Int. J. Mol. Sci. 2018, 19, 2520. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.; Wu, X.; Gong, B.; Huo, R.; Zhao, L.; Li, J.; Lue, G.; Gao, H. GABA Metabolism, Transport and Their Roles and Mechanisms in the Regulation of Abiotic Stress (Hypoxia, Salt, Drought) Resistance in Plants. Metabolites 2023, 13, 347. [Google Scholar] [CrossRef]
- Wu, X.L.; Jia, Q.Y.; Ji, S.X.; Gong, B.B.; Li, J.R.; Lü, G.Y.; Gao, H.B. Gamma-aminobutyric acid (GABA) alleviates salt damage in tomato by modulating Na+ uptake, the GAD gene, amino acid synthesis and reactive oxygen species metabolism. BMC Plant Biol. 2020, 20, 465. [Google Scholar] [CrossRef]
- Nayyar, H.; Kaur, R.; Kaur, S.; Singh, R. γ-Aminobutyric Acid (GABA) Imparts Partial Protection from Heat Stress Injury to Rice Seedlings by Improving Leaf Turgor and Upregulating Osmoprotectants and Antioxidants. J. Plant Growth Regul. 2014, 33, 408–419. [Google Scholar] [CrossRef]
- Li, D.H.; Zhang, D.P.; Zhang, Z.Z.; Xing, Y.M.; Sun, N.; Wang, S.; Cai, H. Exogenous Application of GABA Alleviates Alkali Damage in Alfalfa by Increasing the Activities of Antioxidant Enzymes. Agronomy 2022, 12, 1577. [Google Scholar] [CrossRef]
- Zhou, H.; Chen, H.; Bao, D.; Shin, T.Y.; Zhong, Y.; Zhang, X.; Wu, Y. Recent advances of γ-aminobutyric acid: Physiological and immunity function, enrichment, and metabolic pathway. Front. Nutr. 2022, 9, 1076223. [Google Scholar]
- Shelp, B.J.; Bozzo, G.G.; Trobacher, C.P.; Zarei, A.; Deyman, K.L.; Brikis, C.J. Hypothesis/review: Contribution of putrescine to 4-aminobutyrate (GABA) production in response to abiotic stress. Plant Sci. 2012, 193, 130–135. [Google Scholar] [CrossRef]
- Mei, X.; Chen, Y.; Zhang, L.; Fu, X.; Wei, Q.; Grierson, D.; Zhou, Y.; Huang, Y.; Dong, F.; Yang, Z. Dual mechanisms regulating glutamate decarboxylases and accumulation of gamma-aminobutyric acid in tea (Camellia sinensis) leaves exposed to multiple stresses. Sci. Rep. 2016, 6, 23685. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Hu, X.M.; Jin, L.F.; Shi, C.Y.; Liu, Y.Z.; Peng, S.A. Identification and transcript analysis of two glutamate decarboxylase genes, CsGAD1 and CsGAD2, reveal the strong relationship between CsGAD1 and citrate utilization in citrus fruit. Mol. Biol. Rep. 2014, 41, 6253–6262. [Google Scholar] [CrossRef] [PubMed]
- Takayama, M.; Koike, S.; Kusano, M.; Matsukura, C.; Saito, K.; Ariizumi, T.; Ezura, H. Tomato Glutamate Decarboxylase Genes SlGAD2 and SlGAD3 Play Key Roles in Regulating γ-Aminobutyric Acid Levels in Tomato (Solanum lycopersicum). Plant Cell Physiol. 2015, 56, 1533–1545. [Google Scholar] [CrossRef] [PubMed]
- Akcay, N.; Bor, M.; Karabudak, T.; Ozdemir, F.; Turkan, I. Contribution of Gamma amino butyric acid (GABA) to salt stress responses of Nicotiana sylvestris CMSII mutant and wild type plants. J. Plant Physiol. 2012, 169, 452–458. [Google Scholar] [CrossRef]
- Al-Quraan, N.A.; Sartawe, F.A.-B.; Qaryouti, M.M. Characterization of γ-aminobutyric acid metabolism and oxidative damage in wheat (Triticum aestivum L.) seedlings under salt and osmotic stress. J. Plant Physiol. 2013, 170, 1003–1009. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Xu, Z.; Xu, W.; Li, J.; Zhao, N.; Zhou, Y. Application of γ-aminobutyric acid demonstrates a protective role of polyamine and GABA metabolism in muskmelon seedlings under Ca(NO3)2 stress. Plant Physiol. Biochem. 2015, 92, 1–10. [Google Scholar] [CrossRef]
- Pannerchelvan, S.; Rios-Solis, L.; Wong, F.W.F.; Zaidan, U.H.; Wasoh, H.; Mohamed, M.S.; Tan, J.S.; Mohamad, R.; Halim, M. Strategies for improvement of gamma-aminobutyric acid (GABA) biosynthesis via lactic acid bacteria (LAB) fermentation. Food Funct. 2023, 14, 3929–3948. [Google Scholar] [CrossRef]
- Mleczek, M.; Siwulski, M.; Rzymski, P.; Budka, A.; Kalac, P.; Jasniska, A.; Gasecka, M.; Budzynska, S.; Niedzielski, P. Comparison of elemental composition of mushroom Hypsizygus marmoreus originating from commercial production and experimental cultivation. Sci. Hortic. 2018, 236, 30–35. [Google Scholar] [CrossRef]
- Liu, M.; Meng, G.; Zhang, J.; Zhao, H.; Jia, L. Antioxidant and Hepatoprotective Activities of Mycelia Selenium Polysaccharide by Hypsizigus marmoreus SK-02. Biol. Trace Elem. Res. 2016, 172, 437–448. [Google Scholar] [CrossRef]
- Yan, P.-S.; Cao, L.-X.; Zhang, B.-Z. Efficient Purification of Antiproliferative Polysaccharides from Hypsizigus marmoreus with Radial Flow Chromatography. Biotechnol. Prog. 2014, 30, 872–878. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.K.; Ng, T.B. Hypsin, a novel thermostable ribosome-inactivating protein with antifungal and antiproliferative activities from fruiting bodies of the edible mushroom Hypsizigus marmoreus. Biochem. Biophys. Res. Commun. 2001, 285, 1071–1075. [Google Scholar] [CrossRef] [PubMed]
- Hae, J.R.; Min, Y.U.; Ji, Y.A.; Chang, H.J.; Dam, H.; Tae, W.K.; Tae, Y.H. Anti-obesity Effect of Hypsizigus marmoreus in High Fat-fed Mice. J. Korean Soc. Food Sci. Nutr. 2011, 40, 1708–1714. [Google Scholar]
- Bao, H.; You, S. Molecular Characteristics of Water-Soluble Extracts from Hypsizigus marmoreus and Their in Vitro Growth Inhibition of Various Cancer Cell Lines and Immunomodulatory Function in Raw 264.7 Cells. Biosci. Biotechnol. Biochem. 2011, 75, 891–898. [Google Scholar] [CrossRef]
- Qi, Y.; Zhang, R.; Zhang, M.; Wen, Q.; Shen, J. Effects of exogenous ascorbic acid on the mycelia growth and primordia formation of Pleurotus ostreatus. J. Basic Microbiol. 2021, 61, 736–744. [Google Scholar] [CrossRef]
- You, H.; Wu, C.; Xu, Y.; Hang, J.; Yang, G.; Xu, J. Effects of exogenous GABA on physiological indexes of mycelial of lentinus edodes under different culture temperatures. North. Hortic. 2023, 04, 111–117. [Google Scholar]
- Hou, Z.Q.; Zhao, L.; Wang, Y.T.; Liao, X.J. Purification and Characterization of Superoxide Dismutases from Sea Buckthorn and Chestnut Rose. J. Food Sci. 2019, 84, 746–753. [Google Scholar] [CrossRef]
- Chagas, R.M.; Silveira, J.A.G.; Ribeiro, R.V.; Vitorello, V.A.; Carrer, H. Photochemical damage and comparative performance of superoxide dismutase and ascorbate peroxidase in sugarcane leaves exposed to paraquat-induced oxidative stress. Pestic. Biochem. Physiol. 2008, 90, 181–188. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, Y.; Jiang, Q.; Zhang, Y.; Ma, L.; Ma, S.; Wang, J.; Ma, Y.; Du, M.; Li, J.; et al. Study on CAT activity of tomato leaf cells under salt stress based on microhyperspectral imaging and transfer learning algorithm. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2023, 302, 123047. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, J.; Li, R.F.; Ge, Y.R.; Li, Y.F.; Li, R.L. Plants’ Response to Abiotic Stress: Mechanisms and Strategies. Int. J. Mol. Sci. 2023, 24, 10915. [Google Scholar] [CrossRef]
- Kumari, V.V.; Banerjee, P.; Verma, V.C.; Sukumaran, S.; Chandran, M.A.S.; Gopinath, K.A.; Venkatesh, G.; Yadav, S.K.; Singh, V.K.; Awasthi, N.K. Plant Nutrition: An Effective Way to Alleviate Abiotic Stress in Agricultural Crops. Int. J. Mol. Sci. 2022, 23, 8519. [Google Scholar] [CrossRef]
- Li, C.; Xu, S. Edible mushroom industry in China: Current state and perspectives. Appl. Microbiol. Biotechnol. 2022, 106, 3949–3955. [Google Scholar] [CrossRef] [PubMed]
- Du, X.-H.; Zhao, Q.; Yang, Z.L. A review on research advances, issues, and perspectives of morels. Mycology 2015, 6, 78–85. [Google Scholar] [CrossRef]
- Money, N.P. Hyphal and mycelial consciousness: The concept of the fungal mind. Fungal Biol. 2021, 125, 257–259. [Google Scholar] [CrossRef]
- Zhang, J.; Hao, H.; Chen, M.; Wang, H.; Feng, Z.; Chen, H. Hydrogen-rich water alleviates the toxicities of different stresses to mycelial growth in Hypsizygus marmoreus. AMB Express 2017, 7, 107. [Google Scholar] [CrossRef]
- Kinnersley, A.M.; Lin, F. Receptor modifiers indicate that 4-aminobutyric acid (GABA) is a potential modulator of ion transport in plants. Plant Growth Regul. 2000, 32, 65–76. [Google Scholar] [CrossRef]
- Hijaz, F.; Killiny, N. Exogenous GABA is quickly metabolized to succinic acid and fed into the plant TCA cycle. Plant Signal. Behav. 2019, 14, e1573096. [Google Scholar] [CrossRef]
- Rajput, V.D.; Harish; Singh, R.K.; Verma, K.K.; Sharma, L.; Quiroz-Figueroa, F.R.; Meena, M.; Gour, V.S.; Minkina, T.; Sushkova, S.; et al. Recent Developments in Enzymatic Antioxidant Defence Mechanism in Plants with Special Reference to Abiotic Stress. Biology 2021, 10, 267. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Gong, J.; Luo, S.; Zuo, Y.; Shen, Y. Role of Gamma-Aminobutyric Acid in Plant Defense Response. Metabolites 2023, 13, 741. [Google Scholar] [CrossRef]
- Lei, M.; Wu, X.; Huang, C.; Qiu, Z.; Wang, L.; Zhang, R.; Zhang, J. Trehalose induced by reactive oxygen species relieved the radial growth defects of Pleurotus ostreatus under heat stress. Appl. Microbiol. Biotechnol. 2019, 103, 5379–5390. [Google Scholar] [CrossRef]
- Tyagi, S.; Shumayla; Singh, S.P.; Upadhyay, S.K. Role of Superoxide Dismutases (SODs) in Stress Tolerance in Plants. In Molecular Approaches in Plant Biology and Environmental Challenges; Singh, S.P., Upadhyay, S.K., Pandey, A., Kumar, S., Eds.; Energy Environment and Sustainability; Springer: Berlin/Heidelberg, Germany, 2019; pp. 51–77. [Google Scholar]
- Pandey, S.; Fartyal, D.; Agarwal, A.; Shukla, T.; James, D.; Kaul, T.; Negi, Y.K.; Arora, S.; Reddy, M.K. Abiotic Stress Tolerance in Plants: Myriad Roles of Ascorbate Peroxidase. Front. Plant Sci. 2017, 8, 581. [Google Scholar] [CrossRef]
- Xing, S.G.; Jun, Y.B.; Hau, Z.W.; Liang, L.Y. Higher accumulation of γ-aminobutyric acid induced by salt stress through stimulating the activity of diarnine oxidases in Glycine max (L.) Merr. roots. Plant Physiol. Biochem. 2007, 45, 560–566. [Google Scholar] [CrossRef] [PubMed]
- Bao, H.; Chen, X.; Lv, S.; Jiang, P.; Feng, J.; Fan, P.; Nie, L.; Li, Y. Virus-induced gene silencing reveals control of reactive oxygen species accumulation and salt tolerance in tomato by γ-aminobutyric acid metabolic pathway. Plant Cell Environ. 2015, 38, 600–613. [Google Scholar] [CrossRef]
- Li, Y.; Liu, B.; Peng, Y.; Liu, C.; Zhang, X.; Zhang, Z.; Liang, W.; Ma, F.; Li, C. Exogenous GABA alleviates alkaline stress in Malus hupehensis by regulating the accumulation of organic acids. Sci. Hortic. 2020, 261, 108982. [Google Scholar] [CrossRef]
- Cheng, P.D.; Yue, Q.Y.; Zhang, Y.T.; Zhao, S.; Khan, A.; Yang, X.Y.; He, J.Q.; Wang, S.C.; Shen, W.Y.; Qian, Q.; et al. Application of γ-aminobutyric acid (GABA) improves fruit quality and rootstock drought tolerance in apple. J. Plant Physiol. 2023, 280, 153890. [Google Scholar] [CrossRef]
- Aydin, S.; Naghshiband Hassani, R.; Soleimani Aghdam, M. Exogenous application of GABA retards cap browning in Agaricus bisporus and its possible mechanism. Postharvest Biol. Technol. 2021, 174, 111434. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) |
|---|---|
| gad2-q-Forward | GGTCTCACCAGCACGAAT |
| gad2-q-Reverse | TTGGAAGAGTGTTGTAGCG |
| ACT-Forward | CCGAGCGGAAGTACTCTGTG |
| ACT-Reverse | ATGCTATCTTGCCTCCAGCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Z.; Chen, H.; Zhou, C.; Gong, M.; Li, Y.; Shao, Y.; Wu, Y.; Bao, D. Exogenous γ-Aminobutyric Acid (GABA) Enhanced Response to Abiotic Stress in Hypsizygus marmoreus by Improving Mycelial Growth and Antioxidant Capacity. Metabolites 2024, 14, 94. https://doi.org/10.3390/metabo14020094
Cao Z, Chen H, Zhou C, Gong M, Li Y, Shao Y, Wu Y, Bao D. Exogenous γ-Aminobutyric Acid (GABA) Enhanced Response to Abiotic Stress in Hypsizygus marmoreus by Improving Mycelial Growth and Antioxidant Capacity. Metabolites. 2024; 14(2):94. https://doi.org/10.3390/metabo14020094
Chicago/Turabian StyleCao, Zhi, Hongyu Chen, Chenli Zhou, Ming Gong, Yan Li, Youran Shao, Yingying Wu, and Dapeng Bao. 2024. "Exogenous γ-Aminobutyric Acid (GABA) Enhanced Response to Abiotic Stress in Hypsizygus marmoreus by Improving Mycelial Growth and Antioxidant Capacity" Metabolites 14, no. 2: 94. https://doi.org/10.3390/metabo14020094
APA StyleCao, Z., Chen, H., Zhou, C., Gong, M., Li, Y., Shao, Y., Wu, Y., & Bao, D. (2024). Exogenous γ-Aminobutyric Acid (GABA) Enhanced Response to Abiotic Stress in Hypsizygus marmoreus by Improving Mycelial Growth and Antioxidant Capacity. Metabolites, 14(2), 94. https://doi.org/10.3390/metabo14020094