
Non/Low-Caloric Artificial Sweeteners and Gut Microbiome: From Perturbed Species to Mechanisms

 , ,
, ,
Abstract

1. Introduction
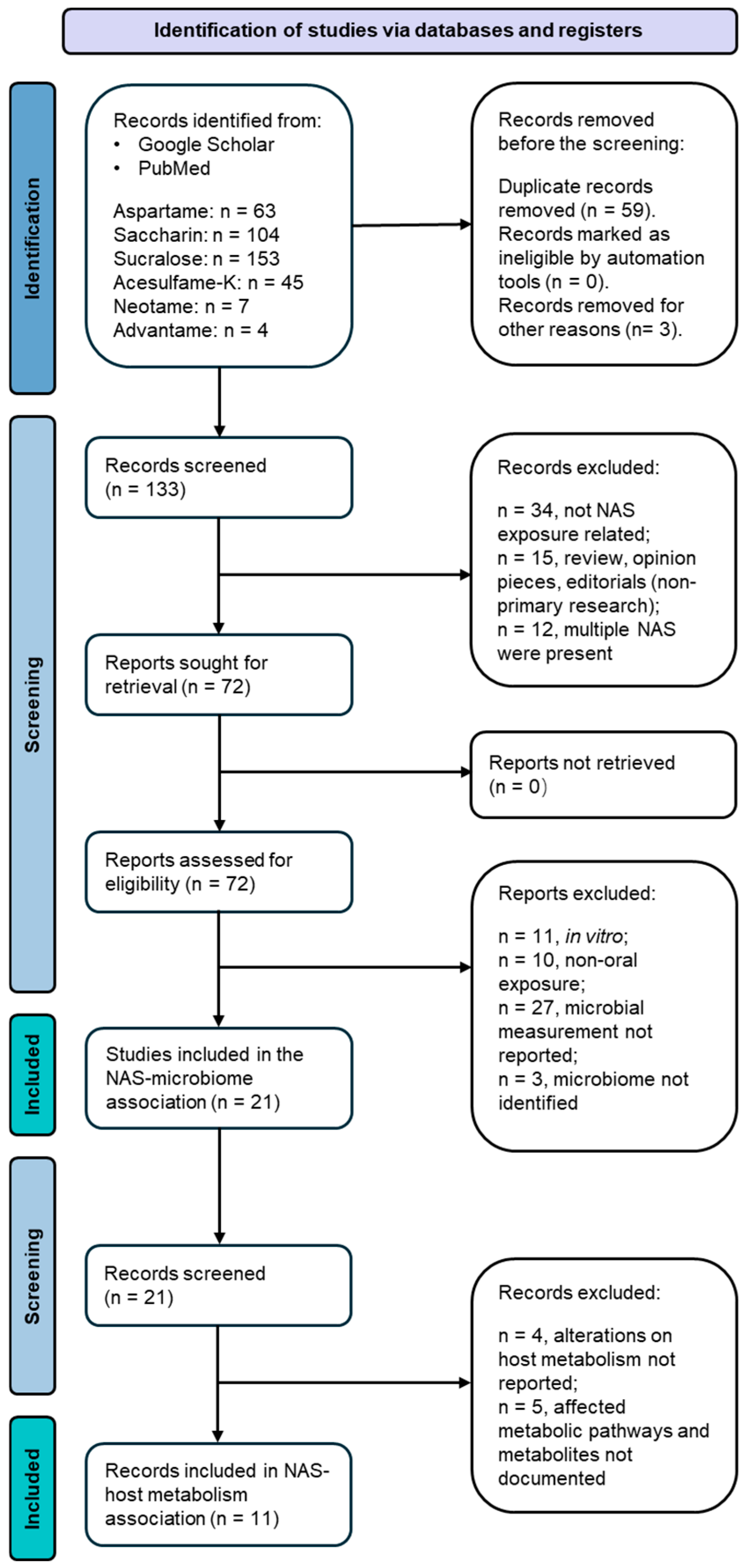
2. Materials and Methods
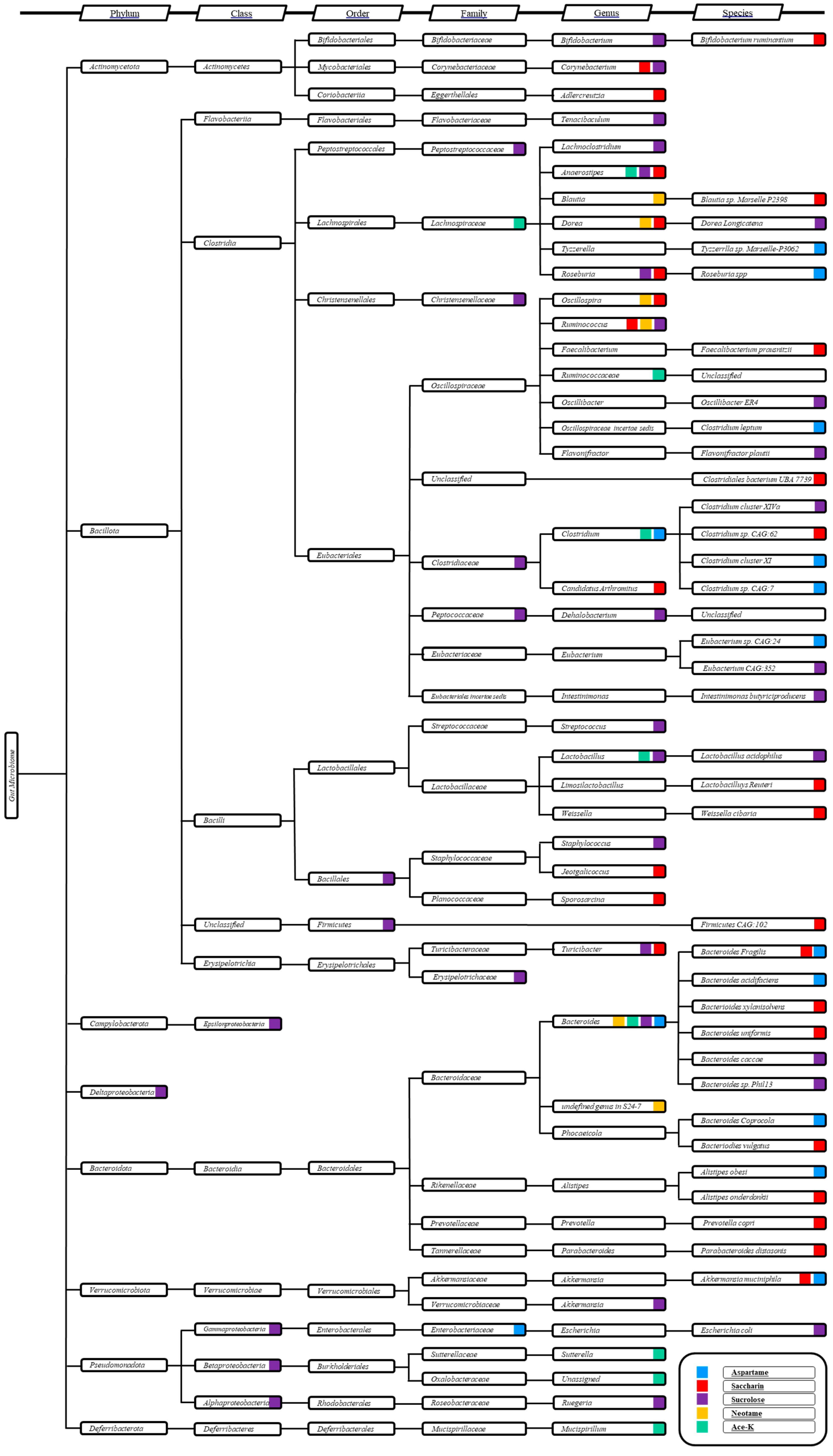
3. Gut Microbiota Species Modulated by NAS Exposure
3.1. Aspartame
3.2. Saccharin
3.3. Sucralose
3.4. Neotame
3.5. Acesulfame Potassium
4. Alterations of Metabolism in NAS–Microbiome–Host Interactions
4.1. Aspartame
4.2. Saccharin
4.3. Sucralose
4.4. Neotame
4.5. Acesulfame Potassium
5. Challenges in Deciphering Underlying NAS–Gut Microbiome Mechanisms
6. Limitation
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Abarca-Gómez, L.; Abdeen, Z.A.; Hamid, Z.A.; Abu-Rmeileh, N.M.; Acosta-Cazares, B.; Acuin, C.; Adams, R.J.; Aekplakorn, W.; Afsana, K.; Aguilar-Salinas, C.A.; et al. Worldwide Trends in Body-Mass Index, Underweight, Overweight, and Obesity from 1975 to 2016: A Pooled Analysis of 2416 Population-Based Measurement Studies in 128·9 Million Children, Adolescents, and Adults. Lancet 2017, 390, 2627–2642. [Google Scholar] [CrossRef] [PubMed]
- Malik, V.S.; Hu, F.B. The Role of Sugar-Sweetened Beverages in the Global Epidemics of Obesity and Chronic Diseases. Nat. Rev. Endocrinol. 2022, 18, 205–218. [Google Scholar] [CrossRef]
- Yu, L.; Zhou, H.; Zheng, F.; Song, J.; Lu, Y.; Yu, X.; Zhao, C. Sugar Is the Key Cause of Overweight/Obesity in Sugar-Sweetened Beverages (SSB). Front. Nutr. 2022, 9, 885704. [Google Scholar] [CrossRef]
- AlFaris, N.A.; Alshwaiyat, N.M.; Alkhalidy, H.; AlTamimi, J.Z.; Alagal, R.I.; Alsaikan, R.A.; Alsemari, M.A.; BinMowyna, M.N.; AlKehayez, N.M. Sugar-Sweetened Beverages Consumption in a Multi-Ethnic Population of Middle-Aged Men and Association with Sociodemographic Variables and Obesity. Front. Nutr. 2022, 9, 987048. [Google Scholar] [CrossRef]
- Lin, W.-T.; Kao, Y.-H.; Li, M.S.; Luo, T.; Lin, H.-Y.; Lee, C.-H.; Seal, D.W.; Hu, C.; Chen, L.-S.; Tseng, T.-S. Sugar-Sweetened Beverages Intake, Abdominal Obesity, and Inflammation among US Adults without and with Prediabetes—An NHANES Study. Int. J. Environ. Res. Public Health 2022, 20, 681. [Google Scholar] [CrossRef] [PubMed]
- Arshad, S.; Rehman, T.; Saif, S.; Rajoka, M.S.R.; Ranjha, M.M.; Hassoun, A.; Cropotova, J.; Trif, M.; Younas, A.; Aadil, R.M. Replacement of Refined Sugar by Natural Sweeteners: Focus on Potential Health Benefits. SSRN J. 2022. [Google Scholar] [CrossRef]
- Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; Israeli, D.; Zmora, N.; Gilad, S.; Weinberger, A.; et al. Artificial Sweeteners Induce Glucose Intolerance by Altering the Gut Microbiota. Nature 2014, 514, 181–186. [Google Scholar] [CrossRef]
- Pino-Seguel, P.; Moya, O.; Borquez, J.C.; Pino-de La Fuente, F.; Díaz-Castro, F.; Donoso-Barraza, C.; Llanos, M.; Troncoso, R.; Bravo-Sagua, R. Sucralose Consumption Ameliorates High-Fat Diet-Induced Glucose Intolerance and Liver Weight Gain in Mice. Front. Nutr. 2022, 9, 979624. [Google Scholar] [CrossRef]
- Guru, S.K.; Li, Y.; Savinova, O.V.; Zhang, Y. Long-Term Consumption of Artificial Sweeteners Does Not Affect Cardiovascular Health and Survival in Rats. PeerJ 2022, 10, e13071. [Google Scholar] [CrossRef]
- Ayada, I.; Li, J.; Pan, Q. Sugar-Sweetened Beverages and Risk of Liver Disease. JAMA 2023, 330, 2217. [Google Scholar] [CrossRef]
- Jones, G.S.; Graubard, B.I.; Ramirez, Y.; Liao, L.M.; Huang, W.-Y.; Alvarez, C.S.; Yang, W.; Zhang, X.; Petrick, J.L.; McGlynn, K.A. Sweetened Beverage Consumption and Risk of Liver Cancer by Diabetes Status: A Pooled Analysis. Cancer Epidemiol. 2022, 79, 102201. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, K. Sugar-Sweetened Beverage Intake Linked to Early-Onset Colorectal Cancer Risk. AJN Am. J. Nurs. 2021, 121, 52. [Google Scholar] [CrossRef] [PubMed]
- Ringel, N.E.; Hovey, K.M.; Andrews, C.A.; Mossavar-Rahmani, Y.; Shadyab, A.H.; Snetselaar, L.G.; Howard, B.V.; Iglesia, C.B. Association of Artificially Sweetened Beverage Consumption and Urinary Tract Cancers in the Women’s Health Initiative Observational Study. Eur. Urol. Open Sci. 2023, 47, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Yang, S.; Ye, Z.; Zhang, Y.; Zhang, Y.; He, P.; Zhou, C.; Hou, F.F.; Qin, X. Tea Consumption and New-Onset Acute Kidney Injury: The Effects of Milk or Sweeteners Addition and Caffeine/Coffee. Nutrients 2023, 15, 2201. [Google Scholar] [CrossRef] [PubMed]
- Debras, C.; Chazelas, E.; Sellem, L.; Julia, C.; Kesse-Guyot, E.; Allès, B.; Deschasaux-Tanguy, M.; Huybrechts, I.; Srour, B.; Touvier, M. Artificial Sweeteners and Risk of Cardiovascular Diseases in the Prospective NutriNet-Santé Cohort. Eur. J. Public Health 2022, 32, ckac129.013. [Google Scholar] [CrossRef]
- Gomez-Delgado, F.; Torres-Peña, J.D.; Gutierrez-Lara, G.; Romero-Cabrera, J.L.; Perez-Martinez, P. Artificial Sweeteners and Cardiovascular Risk. Curr. Opin. Cardiol. 2023, 38, 344–351. [Google Scholar] [CrossRef]
- Singh, S.; Kohli, A.; Trivedi, S.; Kanagala, S.G.; Anamika, F.N.U.; Garg, N.; Patel, M.A.; Munjal, R.S.; Jain, R. The Contentious Relationship between Artificial Sweeteners and Cardiovascular Health. Egypt. J. Intern. Med. 2023, 35, 43. [Google Scholar] [CrossRef]
- Yang, B.; Glenn, A.J.; Liu, Q.; Madsen, T.; Allison, M.A.; Shikany, J.M.; Manson, J.E.; Chan, K.H.K.; Wu, W.-C.; Li, J.; et al. Added Sugar, Sugar-Sweetened Beverages, and Artificially Sweetened Beverages and Risk of Cardiovascular Disease: Findings from the Women’s Health Initiative and a Network Meta-Analysis of Prospective Studies. Nutrients 2022, 14, 4226. [Google Scholar] [CrossRef]
- Hosseini, A.; Barlow, G.M.; Leite, G.; Rashid, M.; Parodi, G.; Wang, J.; Morales, W.; Weitsman, S.; Rezaie, A.; Pimentel, M.; et al. Consuming Artificial Sweeteners May Alter the Structure and Function of Duodenal Microbial Communities. iScience 2023, 26, 108530. [Google Scholar] [CrossRef]
- AL-Ishaq, R.K.; Kubatka, P.; Büsselberg, D. Sweeteners and the Gut Microbiome: Effects on Gastrointestinal Cancers. Nutrients 2023, 15, 3675. [Google Scholar] [CrossRef]
- Panthee, B.; Gyawali, S.; Panthee, P.; Techato, K. Environmental and Human Microbiome for Health. Life 2022, 12, 456. [Google Scholar] [CrossRef]
- Aggarwal, N.; Kitano, S.; Puah, G.R.Y.; Kittelmann, S.; Hwang, I.Y.; Chang, M.W. Microbiome and Human Health: Current Understanding, Engineering, and Enabling Technologies. Chem. Rev. 2023, 123, 31–72. [Google Scholar] [CrossRef]
- Shi, D.; Turroni, S.; Gong, L.; Wu, W.; Yim, H.C.H. Editorial: Manipulation of Gut Microbiota as a Key Target to Intervene on the Onset and Progression of Digestive System Diseases. Front. Med. 2022, 9, 999005. [Google Scholar] [CrossRef]
- McCallum, G.; Tropini, C. The Gut Microbiota and Its Biogeography. Nat. Rev. Microbiol. 2024, 22, 105–118. [Google Scholar] [CrossRef]
- Li, Q.; Li, N.; Cai, W.; Xiao, M.; Liu, B.; Zeng, F. Fermented Natural Product Targeting Gut Microbiota Regulate Immunity and Anti-Inflammatory Activity: A Possible Way to Prevent COVID-19 in Daily Diet. J. Funct. Foods 2022, 97, 105229. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Liu, Y.; Li, H.; Zhao, Y.; Wright, A.-D.G.; Cai, J.; Tian, G.; Mao, X. Differential Effect of Dietary Fibers in Intestinal Health of Growing Pigs: Outcomes in the Gut Microbiota and Immune-Related Indexes. Front. Microbiol. 2022, 13, 843045. [Google Scholar] [CrossRef]
- Campbell, C.; Kandalgaonkar, M.R.; Golonka, R.M.; Yeoh, B.S.; Vijay-Kumar, M.; Saha, P. Crosstalk between Gut Microbiota and Host Immunity: Impact on Inflammation and Immunotherapy. Biomedicines 2023, 11, 294. [Google Scholar] [CrossRef]
- Lu, Y.; Yuan, X.; Wang, M.; He, Z.; Li, H.; Wang, J.; Li, Q. Gut Microbiota Influence Immunotherapy Responses: Mechanisms and Therapeutic Strategies. J. Hematol. Oncol. 2022, 15, 47. [Google Scholar] [CrossRef]
- Kim, D.-H. Gut Microbiota-Mediated Drug-Drug Interactions. Drug Metab. Pharmacokinet. 2017, 32, S18–S19. [Google Scholar] [CrossRef]
- Dhurjad, P.; Dhavaliker, C.; Gupta, K.; Sonti, R. Exploring Drug Metabolism by the Gut Microbiota: Modes of Metabolism and Experimental Approaches. Drug. Metab. Dispos. 2022, 50, 224–234. [Google Scholar] [CrossRef]
- Mishima, E.; Abe, T. Role of the Microbiota in Hypertension and Antihypertensive Drug Metabolism. Hypertens. Res. 2022, 45, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Farhat, E.K.; Sher, E.K.; Džidić-Krivić, A.; Banjari, I.; Sher, F. Functional Biotransformation of Phytoestrogens by Gut Microbiota with Impact on Cancer Treatment. J. Nutr. Biochem. 2023, 118, 109368. [Google Scholar] [CrossRef] [PubMed]
- Rooks, M.G.; Garrett, W.S. Gut Microbiota, Metabolites and Host Immunity. Nat. Rev. Immunol. 2016, 16, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.; Renwick, A.G.; Sims, J.; Snodin, D.J. Sucralose Metabolism and Pharmacokinetics in Man. Food Chem. Toxicol. 2000, 38, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.G.; John, B.A.; Hawkins, D.R. The Pharmacokinetics and Metabolism of Sucralose in the Dog. Food Chem. Toxicol. 2000, 38, 99–106. [Google Scholar] [CrossRef]
- John, B.A.; Wood, S.G.; Hawkins, D.R. The Pharmacokinetics and Metabolism of Sucralose in the Mouse. Food Chem. Toxicol. 2000, 38, 107–110. [Google Scholar] [CrossRef]
- Sims, J.; Roberts, A.; Daniel, J.W.; Renwick, A.G. The Metabolic Fate of Sucralose in Rats. Food Chem. Toxicol. 2000, 38, 115–121. [Google Scholar] [CrossRef]
- Sylvetsky, A.C.; Bauman, V.; Blau, J.E.; Garraffo, H.M.; Walter, P.J.; Rother, K.I. Plasma Concentrations of Sucralose in Children and Adults. Toxicol. Environ. Chem. 2017, 99, 535–542. [Google Scholar] [CrossRef]
- Palmnäs, M.S.A.; Cowan, T.E.; Bomhof, M.R.; Su, J.; Reimer, R.A.; Vogel, H.J.; Hittel, D.S.; Shearer, J. Low-Dose Aspartame Consumption Differentially Affects Gut Microbiota-Host Metabolic Interactions in the Diet-Induced Obese Rat. PLoS ONE 2014, 9, e109841. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, B.; Zhang, Y.; Wei, H.; Lei, Y.; Zhao, L. Structural Shifts of Mucosa-Associated Lactobacilli and Clostridium leptum Subgroup in Patients with Ulcerative Colitis. J. Clin. Microbiol. 2007, 45, 496–500. [Google Scholar] [CrossRef]
- Kabeerdoss, J.; Sankaran, V.; Pugazhendhi, S.; Ramakrishna, B.S. Clostridium Leptum Group Bacteria Abundance and Diversity in the Fecal Microbiota of Patients with Inflammatory Bowel Disease: A Case–Control Study in India. BMC Gastroenterol. 2013, 13, 20. [Google Scholar] [CrossRef] [PubMed]
- Kedia, S.; Rampal, R.; Paul, J.; Ahuja, V. Gut Microbiome Diversity in Acute Infective and Chronic Inflammatory Gastrointestinal Diseases in North India. J. Gastroenterol. 2016, 51, 660–671. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.-C.; Chien, Y.-W.; Yang, S.-C. The Alteration of Gut Microbiota in Newly Diagnosed Type 2 Diabetic Patients. Nutrition 2019, 63–64, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Santacruz, A.; Marcos, A.; Wärnberg, J.; Martí, A.; Martin-Matillas, M.; Campoy, C.; Moreno, L.A.; Veiga, O.; Redondo-Figuero, C.; Garagorri, J.M.; et al. Interplay Between Weight Loss and Gut Microbiota Composition in Overweight Adolescents. Obesity 2009, 17, 1906–1915. [Google Scholar] [CrossRef] [PubMed]
- Koleva, P.T.; Valcheva, R.S.; Sun, X.; Gänzle, M.G.; Dieleman, L.A. Inulin and Fructo-Oligosaccharides Have Divergent Effects on Colitis and Commensal Microbiota in HLA-B27 Transgenic Rats. Br. J. Nutr. 2012, 108, 1633–1643. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Adachi, K.; Sugiyama, T.; Shimozato, A.; Ebi, M.; Ogasawara, N.; Funaki, Y.; Goto, C.; Sasaki, M.; Kasugai, K. Association of Intestinal Microbiota with Metabolic Markers and Dietary Habits in Patients with Type 2 Diabetes. Digestion 2016, 94, 66–72. [Google Scholar] [CrossRef]
- Nie, K.; Ma, K.; Luo, W.; Shen, Z.; Yang, Z.; Xiao, M.; Tong, T.; Yang, Y.; Wang, X. Roseburia Intestinalis: A Beneficial Gut Organism from the Discoveries in Genus and Species. Front. Cell. Infect. Microbiol. 2021, 11, 757718. [Google Scholar] [CrossRef]
- Suez, J.; Cohen, Y.; Valdés-Mas, R.; Mor, U.; Dori-Bachash, M.; Federici, S.; Zmora, N.; Leshem, A.; Heinemann, M.; Linevsky, R.; et al. Personalized Microbiome-Driven Effects of Non-Nutritive Sweeteners on Human Glucose Tolerance. Cell 2022, 185, 3307–3328.e19. [Google Scholar] [CrossRef]
- Scott, N.; Whittle, E.; Jeraldo, P.; Chia, N. A Systemic Review of the Role of Enterotoxic Bacteroides Fragilis in Colorectal Cancer. Neoplasia 2022, 29, 100797. [Google Scholar] [CrossRef]
- Yang, J.-Y.; Lee, Y.-S.; Kim, Y.; Lee, S.-H.; Ryu, S.; Fukuda, S.; Hase, K.; Yang, C.-S.; Lim, H.S.; Kim, M.-S.; et al. Gut Commensal Bacteroides Acidifaciens Prevents Obesity and Improves Insulin Sensitivity in Mice. Mucosal Immunol. 2017, 10, 104–116. [Google Scholar] [CrossRef]
- Chen, K.; Nakasone, Y.; Yi, S.; Ibrahim, H.R.; Sakao, K.; Hossain, M.A.; Hou, D.-X. Natural Garlic Organosulfur Compounds Prevent Metabolic Disorder of Lipid and Glucose by Increasing Gut Commensal Bacteroides acidifaciens. J. Agric. Food Chem. 2022, 70, 5829–5837. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, G.; Zhang, Q.; Liu, Z.; Jiang, X.; Xin, Y. Function of Akkermansia Muciniphila in Type 2 Diabetes and Related Diseases. Front. Microbiol. 2023, 14, 1172400. [Google Scholar] [CrossRef] [PubMed]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia Muciniphila in Overweight and Obese Human Volunteers: A Proof-of-Concept Exploratory Study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Perraudeau, F.; McMurdie, P.; Bullard, J.; Cheng, A.; Cutcliffe, C.; Deo, A.; Eid, J.; Gines, J.; Iyer, M.; Justice, N.; et al. Improvements to Postprandial Glucose Control in Subjects with Type 2 Diabetes: A Multicenter, Double Blind, Randomized Placebo-Controlled Trial of a Novel Probiotic Formulation. BMJ Open Diab. Res. Care 2020, 8, e001319. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Xiong, Q.; Tian, C.; Li, L.; Zhao, J.; Lin, X.; Guo, X.; He, Y.; Liang, W.; Zuo, X.; et al. Inulin-Type Prebiotics Reduce Serum Uric Acid Levels via Gut Microbiota Modulation: A Randomized, Controlled Crossover Trial in Peritoneal Dialysis Patients. Eur. J. Nutr. 2022, 61, 665–677. [Google Scholar] [CrossRef]
- Olaisen, M.; Flatberg, A.; Granlund, A.V.B.; Røyset, E.S.; Martinsen, T.C.; Sandvik, A.K.; Fossmark, R. Bacterial Mucosa-Associated Microbiome in Inflamed and Proximal Noninflamed Ileum of Patients with Crohn’s Disease. Inflamm. Bowel Dis. 2021, 27, 12–24. [Google Scholar] [CrossRef]
- Péan, N.; Le Lay, A.; Brial, F.; Wasserscheid, J.; Rouch, C.; Vincent, M.; Myridakis, A.; Hedjazi, L.; Dumas, M.-E.; Grundberg, E.; et al. Dominant Gut Prevotella Copri in Gastrectomised Non-Obese Diabetic Goto–Kakizaki Rats Improves Glucose Homeostasis through Enhanced FXR Signalling. Diabetologia 2020, 63, 1223–1235. [Google Scholar] [CrossRef]
- Mirande, C.; Kadlecikova, E.; Matulova, M.; Capek, P.; Bernalier-Donadille, A.; Forano, E.; Béra-Maillet, C. Dietary Fibre Degradation and Fermentation by Two Xylanolytic Bacteria Bacteroides xylanisolvens XB1A T and Roseburia intestinalis XB6B4 from the Human Intestine. J. Appl. Microbiol. 2010, 109, 451–460. [Google Scholar] [CrossRef]
- Hajjar, R.; Gonzalez, E.; Fragoso, G.; Oliero, M.; Alaoui, A.A.; Calvé, A.; Vennin Rendos, H.; Djediai, S.; Cuisiniere, T.; Laplante, P.; et al. Gut Microbiota Influence Anastomotic Healing in Colorectal Cancer Surgery through Modulation of Mucosal Proinflammatory Cytokines. Gut 2023, 72, 1143–1154. [Google Scholar] [CrossRef]
- Yang, J.; Zheng, P.; Li, Y.; Wu, J.; Tan, X.; Zhou, J.; Sun, Z.; Chen, X.; Zhang, G.; Zhang, H.; et al. Landscapes of Bacterial and Metabolic Signatures and Their Interaction in Major Depressive Disorders. Sci. Adv. 2020, 6, eaba8555. [Google Scholar] [CrossRef]
- Gauffin Cano, P.; Santacruz, A.; Moya, Á.; Sanz, Y. Bacteroides Uniformis CECT 7771 Ameliorates Metabolic and Immunological Dysfunction in Mice with High-Fat-Diet Induced Obesity. PLoS ONE 2012, 7, e41079. [Google Scholar] [CrossRef] [PubMed]
- López-Almela, I.; Romaní-Pérez, M.; Bullich-Vilarrubias, C.; Benítez-Páez, A.; Gómez Del Pulgar, E.M.; Francés, R.; Liebisch, G.; Sanz, Y. Bacteroides uniformis Combined with Fiber Amplifies Metabolic and Immune Benefits in Obese Mice. Gut Microbes 2021, 13, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Mobini, R.; Tremaroli, V.; Ståhlman, M.; Karlsson, F.; Levin, M.; Ljungberg, M.; Sohlin, M.; Bertéus Forslund, H.; Perkins, R.; Bäckhed, F.; et al. Metabolic Effects of Lactobacillus reuteri DSM 17938 in People with Type 2 Diabetes: A Randomized Controlled Trial. Diabetes Obes. Metab. 2017, 19, 579–589. [Google Scholar] [CrossRef]
- Remely, M.; Hippe, B.; Zanner, J.; Aumueller, E.; Brath, H.; Haslberger, A.G. Gut Microbiota of Obese, Type 2 Diabetic Individuals Is Enriched in Faecalibacterium Prausnitzii, Akkermansia Muciniphila and Peptostreptococcus Anaerobius after Weight Loss. Endocr. Metab. Immune Disord.-Drug Targets 2016, 16, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Abou-Donia, M.B.; El-Masry, E.M.; Abdel-Rahman, A.A.; McLendon, R.E.; Schiffman, S.S. Splenda Alters Gut Microflora and Increases Intestinal P-Glycoprotein and Cytochrome P-450 in Male Rats. J. Toxicol. Environ. Health Part A 2008, 71, 1415–1429. [Google Scholar] [CrossRef]
- Bian, X.; Chi, L.; Gao, B.; Tu, P.; Ru, H.; Lu, K. Gut Microbiome Response to Sucralose and Its Potential Role in Inducing Liver Inflammation in Mice. Front. Physiol. 2017, 8, 487. [Google Scholar] [CrossRef]
- Chi, L.; Bian, X.; Gao, B.; Tu, P.; Lai, Y.; Ru, H.; Lu, K. Effects of the Artificial Sweetener Neotame on the Gut Microbiome and Fecal Metabolites in Mice. Molecules 2018, 23, 367. [Google Scholar] [CrossRef]
- Bian, X.; Chi, L.; Gao, B.; Tu, P.; Ru, H.; Lu, K. The Artificial Sweetener Acesulfame Potassium Affects the Gut Microbiome and Body Weight Gain in CD-1 Mice. PLoS ONE 2017, 12, e0178426. [Google Scholar] [CrossRef]
- Olivier-Van Stichelen, S.; Rother, K.I.; Hanover, J.A. Maternal Exposure to Non-Nutritive Sweeteners Impacts Progeny’s Metabolism and Microbiome. Front. Microbiol. 2019, 10, 1360. [Google Scholar] [CrossRef]
- Frankenfeld, C.L.; Sikaroodi, M.; Lamb, E.; Shoemaker, S.; Gillevet, P.M. High-Intensity Sweetener Consumption and Gut Microbiome Content and Predicted Gene Function in a Cross-Sectional Study of Adults in the United States. Ann. Epidemiol. 2015, 25, 736–742.e4. [Google Scholar] [CrossRef]
- Ahmad, S.Y.; Friel, J.K.; MacKay, D.S. The Effect of the Artificial Sweeteners on Glucose Metabolism in Healthy Adults: A Randomized, Double-Blinded, Crossover Clinical Trial. Appl. Physiol. Nutr. Metab. 2020, 45, 606–612. [Google Scholar] [CrossRef]
- Bian, X.; Tu, P.; Chi, L.; Gao, B.; Ru, H.; Lu, K. Saccharin Induced Liver Inflammation in Mice by Altering the Gut Microbiota and Its Metabolic Functions. Food Chem. Toxicol. 2017, 107, 530–539. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, J.P.D.S.; He, F.; Mangian, H.F.; Oba, P.M.; De Godoy, M.R.C. Dietary Supplementation of a Fiber-Prebiotic and Saccharin-Eugenol Blend in Extruded Diets Fed to Dogs. J. Anim. Sci. 2019, 97, 4519–4531. [Google Scholar] [CrossRef] [PubMed]
- Serrano, J.; Smith, K.R.; Crouch, A.L.; Sharma, V.; Yi, F.; Vargova, V.; LaMoia, T.E.; Dupont, L.M.; Serna, V.; Tang, F.; et al. High-Dose Saccharin Supplementation Does Not Induce Gut Microbiota Changes or Glucose Intolerance in Healthy Humans and Mice. Microbiome 2021, 9, 11. [Google Scholar] [CrossRef] [PubMed]
- Murali, A.; Giri, V.; Cameron, H.J.; Sperber, S.; Zickgraf, F.M.; Haake, V.; Driemert, P.; Walk, T.; Kamp, H.; Rietjens, I.M.; et al. Investigating the Gut Microbiome and Metabolome Following Treatment with Artificial Sweeteners Acesulfame Potassium and Saccharin in Young Adult Wistar Rats. Food Chem. Toxicol. 2022, 165, 113123. [Google Scholar] [CrossRef] [PubMed]
- Uebanso, T.; Ohnishi, A.; Kitayama, R.; Yoshimoto, A.; Nakahashi, M.; Shimohata, T.; Mawatari, K.; Takahashi, A. Effects of Low-Dose Non-Caloric Sweetener Consumption on Gut Microbiota in Mice. Nutrients 2017, 9, 560. [Google Scholar] [CrossRef]
- Rodriguez-Palacios, A.; Harding, A.; Menghini, P.; Himmelman, C.; Retuerto, M.; Nickerson, K.P.; Lam, M.; Croniger, C.M.; McLean, M.H.; Durum, S.K.; et al. The Artificial Sweetener Splenda Promotes Gut Proteobacteria, Dysbiosis, and Myeloperoxidase Reactivity in Crohn’s Disease–Like Ileitis. Inflamm. Bowel Dis. 2018, 24, 1005–1020. [Google Scholar] [CrossRef]
- Wang, Q.-P.; Browman, D.; Herzog, H.; Neely, G.G. Non-Nutritive Sweeteners Possess a Bacteriostatic Effect and Alter Gut Microbiota in Mice. PLoS ONE 2018, 13, e0199080. [Google Scholar] [CrossRef]
- Zheng, Z.; Xiao, Y.; Ma, L.; Lyu, W.; Peng, H.; Wang, X.; Ren, Y.; Li, J. Low Dose of Sucralose Alter Gut Microbiome in Mice. Front. Nutr. 2022, 9, 848392. [Google Scholar] [CrossRef]
- Thomson, P.; Santibañez, R.; Aguirre, C.; Galgani, J.E.; Garrido, D. Short-Term Impact of Sucralose Consumption on the Metabolic Response and Gut Microbiome of Healthy Adults. Br. J. Nutr. 2019, 122, 856–862. [Google Scholar] [CrossRef]
- Méndez-García, L.A.; Bueno-Hernández, N.; Cid-Soto, M.A.; De León, K.L.; Mendoza-Martínez, V.M.; Espinosa-Flores, A.J.; Carrero-Aguirre, M.; Esquivel-Velázquez, M.; León-Hernández, M.; Viurcos-Sanabria, R.; et al. Ten-Week Sucralose Consumption Induces Gut Dysbiosis and Altered Glucose and Insulin Levels in Healthy Young Adults. Microorganisms 2022, 10, 434. [Google Scholar] [CrossRef] [PubMed]
- Chi, L.; Yang, Y.; Bian, X.; Gao, B.; Tu, P.; Ru, H.; Lu, K. Chronic Sucralose Consumption Inhibits Farnesoid X Receptor Signaling and Perturbs Lipid and Cholesterol Homeostasis in the Mouse Livers, Potentially by Altering Gut Microbiota Functions. Sci. Total Environ. 2024, 919, 169603. [Google Scholar] [CrossRef] [PubMed]
- Hanawa, Y.; Higashiyama, M.; Kurihara, C.; Tanemoto, R.; Ito, S.; Mizoguchi, A.; Nishii, S.; Wada, A.; Inaba, K.; Sugihara, N.; et al. Acesulfame Potassium Induces Dysbiosis and Intestinal Injury with Enhanced Lymphocyte Migration to Intestinal Mucosa. J. Gastroenterol. Hepatol. 2021, 36, 3140–3148. [Google Scholar] [CrossRef] [PubMed]
- Baldelli, V.; Scaldaferri, F.; Putignani, L.; Del Chierico, F. The Role of Enterobacteriaceae in Gut Microbiota Dysbiosis in Inflammatory Bowel Diseases. Microorganisms 2021, 9, 697. [Google Scholar] [CrossRef] [PubMed]
- Morgan, X.C.; Tickle, T.L.; Sokol, H.; Gevers, D.; Devaney, K.L.; Ward, D.V.; Reyes, J.A.; Shah, S.A.; LeLeiko, N.; Snapper, S.B.; et al. Dysfunction of the Intestinal Microbiome in Inflammatory Bowel Disease and Treatment. Genome Biol. 2012, 13, R79. [Google Scholar] [CrossRef] [PubMed]
- Lupp, C.; Robertson, M.L.; Wickham, M.E.; Sekirov, I.; Champion, O.L.; Gaynor, E.C.; Finlay, B.B. Host-Mediated Inflammation Disrupts the Intestinal Microbiota and Promotes the Overgrowth of Enterobacteriaceae. Cell Host Microbe 2007, 2, 204. [Google Scholar] [CrossRef]
- Gophna, U.; Sommerfeld, K.; Gophna, S.; Doolittle, W.F.; Veldhuyzen Van Zanten, S.J.O. Differences between Tissue-Associated Intestinal Microfloras of Patients with Crohn’s Disease and Ulcerative Colitis. J. Clin. Microbiol. 2006, 44, 4136–4141. [Google Scholar] [CrossRef]
- Martinez-Medina, M.; Aldeguer, X.; Gonzalez-Huix, F.; Acero, D.; Garcia-Gil, J.L. Abnormal Microbiota Composition in the Ileocolonic Mucosa of Crohn’s Disease Patients as Revealed by Polymerase Chain Reaction-Denaturing Gradient Gel Electrophoresis: Inflamm. Bowel Dis. 2006, 12, 1136–1145. [Google Scholar] [CrossRef]
- Wang, M.; Molin, G.; Ahrné, S.; Adawi, D.; Jeppsson, B. High Proportions of Proinflammatory Bacteria on the Colonic Mucosa in a Young Patient with Ulcerative Colitis as Revealed by Cloning and Sequencing of 16S rRNA Genes. Dig. Dis. Sci. 2007, 52, 620–627. [Google Scholar] [CrossRef]
- Carvalho, F.A.; Barnich, N.; Sauvanet, P.; Darcha, C.; Gelot, A.; Darfeuille-Michaud, A. Crohn’s Disease-Associated Escherichia Coli LF82 Aggravates Colitis in Injured Mouse Colon via Signaling by Flagellin: Inflamm. Bowel Dis. 2008, 14, 1051–1060. [Google Scholar] [CrossRef]
- Mukhopadhya, I.; Hansen, R.; El-Omar, E.M.; Hold, G.L. IBD—What Role Do Proteobacteria Play? Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 219–230. [Google Scholar] [CrossRef]
- Lavelle, A.; Lennon, G.; O’Sullivan, O.; Docherty, N.; Balfe, A.; Maguire, A.; Mulcahy, H.E.; Doherty, G.; O’Donoghue, D.; Hyland, J.; et al. Spatial Variation of the Colonic Microbiota in Patients with Ulcerative Colitis and Control Volunteers. Gut 2015, 64, 1553–1561. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Liu, W.; Wu, J.; Wang, W.; Wang, Z.; Yu, X.; Zhang, H.; Zhu, L.; Zhan, X. Metabolic Profiles of Oligosaccharides Derived from Four Microbial Polysaccharides by Faecal Inocula from Type 2 Diabetes Patients. Int. J. Food Sci. Nutr. 2021, 72, 1083–1094. [Google Scholar] [CrossRef]
- Sarbini, S.R.; Kolida, S.; Gibson, G.R.; Rastall, R.A. In Vitro Fermentation of Commercial α-Gluco-Oligosaccharide by Faecal Microbiota from Lean and Obese Human Subjects. Br. J. Nutr. 2012, 109, 1980–1989. [Google Scholar] [CrossRef] [PubMed]
- Bornemann, V.; Werness, S.C.; Buslinger, L.; Schiffman, S.S. Intestinal Metabolism and Bioaccumulation of Sucralose in Adipose Tissue In The Rat. J. Toxicol. Environ. Health Part A 2018, 81, 913–923. [Google Scholar] [CrossRef]
- Magnuson, B.A.; Carakostas, M.C.; Moore, N.H.; Poulos, S.P.; Renwick, A.G. Biological Fate of Low-Calorie Sweeteners. Nutr. Rev. 2016, 74, 670–689. [Google Scholar] [CrossRef]
- Matthews, H.B.; Fields, M.; Fishbein, L. Saccharin. Distribution and Excretion of a Limited Dose in the Rat. J. Agric. Food Chem. 1973, 21, 916–919. [Google Scholar] [CrossRef]
- Renwick, A.G. The Metabolism of Intense Sweeteners. Xenobiotica 1986, 16, 1057–1071. [Google Scholar] [CrossRef]
- Byard, J.L.; McChesney, E.W.; Golberg, L.; Coulston, F. Excretion and Metabolism of Saccharin in Man. II. Studies with 14C-Labelled and Unlabelled Saccharin. Food Cosmet. Toxicol. 1974, 12, 175–184. [Google Scholar] [CrossRef]
- McChesney, E.W.; Golberg, L. The Excretion and Metabolism of Saccharin in Man. I. Methods of Investigation and Preliminary Results. Food Cosmet. Toxicol. 1973, 11, 403–414. [Google Scholar] [CrossRef]
- Lethco, E.J.; Wallace, W.C. The Metabolism of Saccharin in Animals. Toxicology 1975, 3, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Sweatman, T.W.; Renwick, A.G. The Tissue Distribution and Pharmacokinetics of Saccharin in the Rat. Toxicol. Appl. Pharmacol. 1980, 55, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Stegink, L.D.; Filer, L.J. Aspartame: Physiology and Biochemistry, 1st ed.; CRC Press: Boca Raton, FL, USA, 2020; ISBN 978-1-00-014652-3. [Google Scholar]
- Stegink, L.D. The Aspartame Story: A Model for the Clinical Testing of a Food Additive. Am. J. Clin. Nutr. 1987, 46, 204–215. [Google Scholar] [CrossRef] [PubMed]
- Butchko, H.H.; Stargel, W.W.; Comer, C.P.; Mayhew, D.A.; Benninger, C.; Blackburn, G.L.; De Sonneville, L.M.J.; Geha, R.S.; Hertelendy, Z.; Koestner, A.; et al. Aspartame: Review of Safety. Regul. Toxicol. Pharmacol. 2002, 35, S1–S93. [Google Scholar] [CrossRef]
- Hooper, N.M.; Hesp, R.J.; Tieku, S. Metabolism of Aspartame by Human and Pig Intestinal Microvillar Peptidases. Biochem. J. 1994, 298, 635–639. [Google Scholar] [CrossRef]
- Ranney, R.E.; Oppermann, J.A.; Muldoon, E.; McMahon, F.G. Comparative Metabolism of Aspartame in Experimental Animals and Humans. J. Toxicol. Environ. Health 1976, 2, 441–451. [Google Scholar] [CrossRef]
- O’donnell, K. Aspartame, Neotame and Advantame. In Sweeteners and Sugar Alternatives in Food Technology; O’Donnell, K., Kearsley, M.W., Eds.; Wiley: Hoboken, NJ, USA, 2012; pp. 117–136. ISBN 978-0-470-65968-7. [Google Scholar]
- Nettleton, J.E.; Cho, N.A.; Klancic, T.; Nicolucci, A.C.; Shearer, J.; Borgland, S.L.; Johnston, L.A.; Ramay, H.R.; Noye Tuplin, E.; Chleilat, F.; et al. Maternal Low-Dose Aspartame and Stevia Consumption with an Obesogenic Diet Alters Metabolism, Gut Microbiota and Mesolimbic Reward System in Rat Dams and Their Offspring. Gut 2020, 69, 1807–1817. [Google Scholar] [CrossRef]
- Fons, F.; Gomez, A.; Karjalainen, T. Mechanisms of Colonisation and Colonisation Resistance of the Digestive Tract Part 2: Bacteria/Bacteria Interactions. Microb. Ecol. Health Dis. 2000, 12, 240–246. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, F.; Ding, X.; Wu, G.; Lam, Y.Y.; Wang, X.; Fu, H.; Xue, X.; Lu, C.; Ma, J.; et al. Gut Bacteria Selectively Promoted by Dietary Fibers Alleviate Type 2 Diabetes. Science 2018, 359, 1151–1156. [Google Scholar] [CrossRef]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-Chain Fatty Acids in Control of Body Weight and Insulin Sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef]
- Chambers, E.S.; Preston, T.; Frost, G.; Morrison, D.J. Role of Gut Microbiota-Generated Short-Chain Fatty Acids in Metabolic and Cardiovascular Health. Curr. Nutr. Rep. 2018, 7, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The Role of Short-Chain Fatty Acids in Health and Disease. In Advances in Immunology; Elsevier: Amsterdam, The Netherlands, 2014; Volume 121, pp. 91–119. ISBN 978-0-12-800100-4. [Google Scholar]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed]
- Byrne, C.S.; Chambers, E.S.; Morrison, D.J.; Frost, G. The Role of Short Chain Fatty Acids in Appetite Regulation and Energy Homeostasis. Int. J. Obes. 2015, 39, 1331–1338. [Google Scholar] [CrossRef] [PubMed]
- Bach Knudsen, K.E. Microbial Degradation of Whole-Grain Complex Carbohydrates and Impact on Short-Chain Fatty Acids and Health. Adv. Nutr. 2015, 6, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Parada Venegas, D.; De La Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef]
- Hamer, H.M.; Jonkers, D.; Venema, K.; Vanhoutvin, S.; Troost, F.J.; Brummer, R.-J. Review Article: The Role of Butyrate on Colonic Function. Aliment. Pharmacol. Ther. 2008, 27, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yin, A.; Li, H.; Wang, R.; Wu, G.; Shen, J.; Zhang, M.; Wang, L.; Hou, Y.; Ouyang, H.; et al. Dietary Modulation of Gut Microbiota Contributes to Alleviation of Both Genetic and Simple Obesity in Children. EBioMedicine 2015, 2, 968–984. [Google Scholar] [CrossRef]
- Knowles, S.R.; Nelson, E.A.; Palombo, E.A. Investigating the Role of Perceived Stress on Bacterial Flora Activity and Salivary Cortisol Secretion: A Possible Mechanism Underlying Susceptibility to Illness. Biol. Psychol. 2008, 77, 132–137. [Google Scholar] [CrossRef]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment Dominates over Host Genetics in Shaping Human Gut Microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef]
- Milani, C.; Duranti, S.; Bottacini, F.; Casey, E.; Turroni, F.; Mahony, J.; Belzer, C.; Delgado Palacio, S.; Arboleya Montes, S.; Mancabelli, L.; et al. The First Microbial Colonizers of the Human Gut: Composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol. Mol. Biol. Rev. 2017, 81, e00036-17. [Google Scholar] [CrossRef]
- Ruiz-Ojeda, F.J.; Plaza-Díaz, J.; Sáez-Lara, M.J.; Gil, A. Effects of Sweeteners on the Gut Microbiota: A Review of Experimental Studies and Clinical Trials. Adv. Nutr. 2019, 10, S31–S48. [Google Scholar] [CrossRef] [PubMed]
- Moya-Pérez, A.; Neef, A.; Sanz, Y. Bifidobacterium Pseudocatenulatum CECT 7765 Reduces Obesity-Associated Inflammation by Restoring the Lymphocyte-Macrophage Balance and Gut Microbiota Structure in High-Fat Diet-Fed Mice. PLoS ONE 2015, 10, e0126976. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| NAS | Exposure | Altered Gut Microbiome * | Analytical Methods | Reference |
|---|---|---|---|---|
| Aspartame ADI: 50 mg/kg Sucrose equivalence: 200× 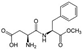 | Rats, SD Normal rats (N = 10–12), 5 mg/kg/d, 8 weeks Obese rats (N = 10–12), 7 mg/kg/d, 8 weeks | Normal rats: Clostridium leptum ↑ Obese rats: Clostridium cluster XI ↓ Enterobacteriaceae ↑ C. leptum ↑ and Roseburia spp. ↑ | Fecal DNA extraction + 16S rRNA sequencing + qRT-PCR analysis | (Palmnäs et al., 2014) [39] |
| Humans, Exposed (N = 20), 0.24 g and 5.76 g glucose, 4 weeks | Top glycemic responders: Bacteroides Fragilis and Bacteroides acidifaciens ↑ Bacteroides Coprocola ↓ Bottom glycemic responders: Akkenmensia muciniphila ↑ Top compared to bottom responders: Clostridium sp. CAG:7 ↑ Tyzzerrlla sp. Marseille-P3062 ↑ Alistipes obesi and Eubacterium sp. CAG:24 ↓ | 16S rRNA sequencing Illumina NextSeq platform | (Suez et al., 2022) [48] | |
| Humans, Exposed (N = 7), 1.7–33.2 mg/d, based on daily food records in four days Non-exposed (N = 24) | No significant changes | 16S rRNA sequencing Length heterogeneity polymerase chain reaction (PCR) fingerprinting | (Frankenfeld et al., 2015) [70] | |
| Humans, Exposed (N = 15), 0.425 g/day, 14 days | No significant changes | 16S rRNA sequencing Illumina MiSeq | (Ahmad et al., 2020) [71] | |
| Saccharin ADI: 5 mg/kg Sucrose equivalence: 300× 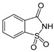 | Humans, Exposed (N = 20), 0.18 g and 5.82 g glucose, 28 days | Top glycemic responders: Prevotella copri ↑ Bacterioides xylanisolvens ↓ Alistipes onderdonkii ↑ Firmicutes CAG:102 ↓ Top compared to bottom responders: Blautia sp. Marselle P2398 ↑ Clostridium sp. CAG:62 ↑ Bifidobacterium ruminantium ↓ Clostridiales bacterium UBA 7739 ↓ Faecalibacterium prausnitzii ↓ Parabacteroides distasonis ↓ | 16S rRNA sequencing Illumina NextSeq platform | (Suez et al., 2022) [48] |
| Mice, C57BL/6 Exposed (N = 20), 3333 mg/kg/d, 11 weeks Control (N = 20) | Bacteroides uniformis ↑ Lactobacilluys reuteri ↓ Bacteriodies vulgatus ↑ Akkermansia muciniphila ↓ | 16S rRNA sequencing | (Suez et al., 2014) [7] | |
| Humans, (5 males and 2 females, aged 28–36) Exposed, 5 mg per kg of body weight for 5 days | Bacteroides fragilis ↑ Weissella cibaria ↑ Candidatus arthromitus ↓ | 16S rRNA sequencing | (Suez et al., 2014) [7] | |
| Mice, male, C57BL/6J (8 weeks old) 0.3 mg/mL in water for six months | After three-month consumption: Anaerostipes ↓ Ruminococcus ↓ Sporosarcina ↑ Jeotgalicoccus ↑ Akkermansia ↑ Scillospira and Corynebacterium ↑ After six-month consumption: Ruminococcus ↓ Adlercreutzia ↓ and Dorea ↓ Corynebacterium ↑, Roseburia ↑ and Turicibacter ↑. | 16S rRNA gene sequencing | (Bian et al., 2017) [72] | |
| Dogs, female beagles 0.02% saccharin and eugenol, or 5% fiber blend plus 0.02% saccharin and eugenol for 10 days (N = 8) | No shifts in fecal microbial richness and diversity | 16S rRNA gene sequencing | (Nogueira et al., 2019) [73] | |
| Mice, C57Bl\6J and Whole body T1R2-deficient mice, (eight-week-old) 250 mg/kg for 10 weeks | No alterations in microbial diversity or composition at any taxonomic level. | 16S rRNA gene sequencing | (Serrano et al., 2021) [74] | |
| Humans, 18–45 years old (1) pulp filler/placebo (1000 mg/day 1) sodium saccharin (400 mg/day), (3) lactisole (670 mg/day), or (4) sodium saccharin (400 mg/day) + lactisole (670 mg/day) twice daily for 2 weeks | No alterations in microbial diversity or composition at any taxonomic level | 16S rRNA gene sequencing | (Serrano et al., 2021) [74] | |
| Wistar rats, Exposed, 20 and 100 mg/kg body weight/day for 28 days | No effects on microbiome changes | 16S rRNA gene sequencing | (Murali et al., 2022) [75] | |
| Sucralose ADI: 15 mg/kg Sucrose equivalence: 600× 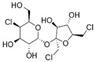 | Mice, C57BL/6 J Exposed (N = 10), 9–22 mg/kg/d, 6 months Control (N = 10) | Turicibacteraceae Turicibacter ↑ Lachnospiraceae ruminococcus ↑ Ruminococcaceae ruminococcus ↑ Verrucomicrobiaceae akkermansia ↑ Unclassified members in Family Clostridiaceae↑Christensenellaceae ↑ Staphylococcaceae Staphylococcus ↓ Streptococcaceae streptococcus ↓ Dehalobacteriaceae dehalobacterium ↓ Lachnospiraceae anaerostipes ↓ Lachnospiraceae roseburia ↓ Unassigned Peptostreptococcaceae ↓ Erysipelotrichaceae ↓ Bacillales ↓ | 16S rRNA sequencing | (Bian, 2017) [66] |
| Humans, Exposed (N = 20), 0.18 g and 5.82 g glucose, 28 days | Top glycemic responders: Eubacterium CAG:352 ↑ Dorea longicatena ↑ Oscillibacter ER4 ↓ Top compared to bottom responders: Bacteroides caccae ↑ Bacteroides sp. Phil13 ↑ Flavonifractor plautii ↑ Intestinimonas butriciproducens ↓ | 16S rRNA sequencing Illumina NextSeq platform | (Suez et al., 2022) [48] | |
| Rats, SD Exposed (N = 10/group), Splenda 1.1, 3.3, 5.5 or 11 mg/kg/d, 12 weeks Control (N = 10) | Bifidobacterial ↓ Lactobacilli ↓ Bacteroides ↓ | Culturing plates | (Abou-Donia et al., 2008) [65] | |
| Mice, C57Bl/6J mice (4 weeks old) Exposed (N = 8/group), sucralose 1.4 ± 0.1 mg/kg BW/day and 14.2 ± 2.2 mg/kg BW/day Control (N = 8) | Clostridium cluster XIVa↓ | 16S rRNA sequencing | (Uebanso et al., 2017) [76] | |
| Mice, SAMP1/YitFc (SAMP) Exposed(N = 5–7/group), 6-week supplementation of Splenda; ingredients: sucralose/maltodextrin, 1:99, w/w), 1.08 mg/mL; 3.5 mg/mL; 35 mg/mL | Five classes in Proteobacteria phylum ↑ (Alphaproteobacteria, Betaproteobacteria, Gammaproteobacteria, Epsilonproteobacteria, Deltaproteobacteria) Escherichia coli ↑ | Culturing plates + 16S rRNA sequencing | (Rodriguez-Palacios et al., 2018) [77] | |
| Mice, C57BL/6 (5 weeks old) Exposed (N = 8/group), 8 weeks, sucralose (2.5%, w/v) | In chow-only mice: Firmicutes ↓, Bacteroidetes ↓, Bifidobacterium ↑ In high-fat-diet mice: Firmicutes ↑, Bacteroidetes ↓ | 16S rDNA sequencing | (Wang et al., 2018) [78] | |
| Mice, Pathogen-free (SPF) C57BL/6J, male, (28 days) Exposed (N = 8/group), 0.0003 g/mL, 0.003 mg/mL, 0.03 mg/mL, 0.3 mg/mL per day for 16 weeks | In jejunum: Tenacibaculum ↑, Ruegeria ↑ In ileum: Staphylococcus ↑, Corynebacterium ↑ In cecum: Lachnoclostridium ↓, Lachnospiraceae UCG-006 ↓ | 16S rDNA sequencing | (Zheng et al., 2022) [79] | |
| Humans, 18–50 years old Exposed (N = 16), 780 mg sucralose/day for 7 days Control (N = 14) | No significant changes | 16S rDNA sequencing | (Thomson et al., 2019) [80] | |
| Humans, 18–35 years old Exposed (N = 20/group), 48 mg Splenda/day for 10 weeks | Lactobacillus acidophilus ↓ Blautia coccoides ↑ | 16S rRNA sequencing Quantitative polymerase chain reaction (qPCR) | (Méndez-García et al., 2022) [81] | |
| Mice, C57BL/6J, 8 weeks old Exposed (N = 10), 0.1 mg/mL for 6 months | Lactobacillus ↓ Ruminococcus ↓ | 16S rDNA sequencing | (Chi et al., 2024) [82] | |
| Acesulfame potassium ADI: 15 mg/kg Sucrose equivalence: 200× 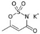 | Mice, CD-1 Exposed (N = 5), 37.5 mg/kg/d, 4 weeks Control (N = 5) | Males: Bacteroides ↑; Anaerostipes ↑; Sutterella ↑ Females: Mucispirillum ↑, Lactobacillus ↓, Clostridium ↓, an unassigned Ruminococcaceae genus and an unassigned Oxalobacteraceae genus ↓ | 16S rRNA sequencing | (Bian, et al., 2017) [68] |
| Mice, C57Bl/6J mice (4 weeks old) Exposed (N = 9/group), 15 mg/kg BW/day Control (N = 8) | No significant changes | 16S rRNA sequencing | Uebanso et al., 2017) [76] | |
| Mice, C57BL/6J, (8 weeks old) Exposed 150 mg/kg b.w./day for 8 weeks | Clostridiaceae ↓ Lachnospiraceae ↓ Ruminococcacea ↓ | 16S rRNA sequencing | (Hanawa et al., 2021) [83] | |
| Humans, Exposed (N = 7), 1.7–33.2 mg/d, based on daily food records in four days Non-exposed (N = 24) | No significant changes | 16S rRNA sequencing | (Frankenfeld et al., 2015) [70] | |
| Neotame ADI: 18 mg/kg Sucrose equivalence: 7000–13,000× 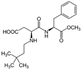 | Mice, CD-1 Exposed (N = 5), 0.75 mg/kg/d, 4 weeks Control (N = 5) | Bacteroidetes including Bacteroides and one undefined genus in S24-7 ↑ Three genera in the family Ruminococcaceae, consisting of Oscillospira, Ruminococcus, and one undefined genus, and five genera in family Lachnospiraceae, which contained Blautia, Dorea, Ruminococcus, and two undefined genera. ↓ | 16S rRNA sequencing | (Chi et al., 2018) [67] |
| NAS | Altered Host Metabolism Pathways * | Key Metabolites Changes in Host ** | Analytical Methods | Source |
|---|---|---|---|---|
| Aspartame ADI: 50 mg/kg Sucrose equivalence: 200× 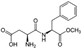 | Fasting hyperglycemia and impaired insulin tolerance Pathways: Gluconeogenesis (?) | Acetate (?), butyrate ↓, propionate ↑ | Serum metabolomic proton nuclear magnetic resonance spectroscopy (1H NMR) | (Palmnäs et al., 2014) [39] |
| Altered glycemic response Pathways: Fatty acid degradation ↓ L-methionine biosynthesis ↓ Peptidoglycan biosynthesis ↓ Top compared to bottom responders: Pyrimidine nucleobases salvage ↑ L-omithine biosynthesis ↑ Heme biosynthesis ↑ Urea Cycle ↑ Phosphonate and phosphinate metabolism ↓ Flavin biosynthesis ↓ Pyridoxal 5′-phosphate biosynthesis and salvage ↓ L-histidine degradation ↓ L-proline biosynthesis ↓ | Kynurenine ↑, terephthalic acid ↓, indole-3-acetate ↑, benzoate ↑ | Serum metabolomic UPLC + Q-ToF mass spectrometry | (Suez et al., 2022) [48] | |
| Saccharin ADI: Sucrose equivalence: 400× 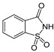 | Altered glycemic response UMP biosynthesis ↑ glycolysis and glycan degradation ↓ homolactic fermentation ↓ glycolysis I from glucose 6-phosphate ↓ glycolysis II from fructose 6-phosphate ↓ glycerol degradation to butanol ↓ hexitol degradation ↓ Neu5Ac degradation ↓ | 4-hydroxybenzoate ↑, benzoate ↑, indoxyl sulfate ↑, hexadecanedioic acid ↓ | Serum metabolomic UPLC + Q-ToF mass spectrometry | (Suez et al., 2022) [48] |
| Induced glucose intolerance glycolysis and glycan degradation ↑ | Propionate ↑ acetate ↑ | Fecal metabolomic HPLC | (Suez et al., 2014) [7] | |
| Increased inflammation in the host by increasing the abundance of bacterial genes involved in elevating the pro-inflammatory mediators LPS synthesis ↑ Bacterial toxins ↑ Flagellar assembly protein ↑ Fimbrial protein ↑ Drug resistance ↑ | Daidzein ↑ dihydrodaidzein ↑ O-desmethylangolensin ↑ Equol ↓ linoleoyl ethanolamide ↓ palmitoleoyl ethanolamide ↓ N,N-Dimethylsphingosine ↓ quinolinic acid ↑ | Fecal metabolite analysis HPLC-Q-ToF | (Bian et al., 2017) [72] | |
| - | Acetate ↑, propionate ↑, butyrate ↑ | Fecal metabolomic HPLC | (Serrano et al., 2021) [74] | |
| Altered amino acids, lipids, energy metabolism and bile acids in the plasma | - | Targeted MS-based metabolome profiling | (Murali et al., 2022) [75] | |
| Sucralose ADI: 15 mg/kg Sucrose equivalence: 600× 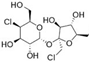 | LPS synthesis ↑ Flagella protein synthesis ↑ Fimbriae synthesis ↑ Bacterial toxins and drug resistance genes ↑ Quorum sensing signals ↓ Amino acids and derivatives ↓ Bile acids (?) | N-butanoyl-l-homoserine lactone ↓, N-(3-oxo-hexanoyl)-homoserine lactone ↓, N-tetradecanoyl-L-homoserine lactone ↓, and N-pentadecanoyl-L-homoserine lactone ↓ L-tryptophan (Trp) ↑, quinolinic acid ↑, kynurenic acid ↓, and 2-aminomuconic acid ↑ L-tyrosine ↑, p-hydroxyphenylacetic acid ↓, and cinnamic acid ↓ 3-Oxo-4,6-choladienoic acid ↑, 3β,7α-dihydroxy-5-cholestenoate ↓, 3α,7β,12α-trihydroxyoxocholanyl-glycine ↓, and lithocholic acid/isoallolithocholic acid/allolithocholic acid/isolithocholic acid ↓ | Fecal metabolomic HPLC-Q-ToF | (Bian, et al., 2017) [66] |
| Arginine biosynthesis ↑ Mixed acid fermentation ↓ TCA cycle ↓ Urate biosynthesis/inosine 5′-phosphate degradation ↓ Adenosine deoxyribonucleotide de novo biosynthesis ↓ Guanosine nucleotide de novo biosynthesis ↓ | Isocitrate ↑, trans-aconitate↑, serine ↑, N-acetylalanine ↑, aspartate ↑, quinolinate ↑, 2-C-methyl-D-erythritol 4-phosphate ↑, galactarate ↑, psicose ↑, pseudouridine ↓, uric acid ↓, and sebacic acid ↓ | Serum metabolomic UPLC + Q-ToF mass spectrometry | (Suez et al., 2022) [48] | |
| Cholesterol–bile acid metabolism | Hepatic cholesterol ↑ cholic acid ↑, ratio of secondary bile acids (dehydrocholic acids (DCA) and lithocholic acid (LCA)) to primary bile acids (CA and CDCA) ↑ | Liver/cecal metabolomic LC-MS | (Uebanso et al., 2017) [76] | |
| 1. Richness of bile salt hydrolase gene (choloylglycine hydrolase) ↓, secondary bile acid synthesis pathway ↓ 2. Bile acid compositions and Farnesoid X Receptor (FXR) activation: (1)Ratios of various free bile acids and taurine-conjugated bile acids, including αMCA/TαMCA, ωMCA/TωMCA, CDCA/TCDCA and DCA/TDCA ↓, moderately for βMCA/TβMCA (p < 0.07) and CA/TCA (p < 0.06) ↓ (2)Expression of genes of Farnesoid X Receptor (FXR) signaling in livers, including Shp, Cyp7a1, Cyp27a1, and Ntcp ↓ 3. Altered hepatic cholesterol homeostasis Expressions of genes encoding three major cholesterol efflux transporters, including Abca1, Abcg5, and Abcg8 ↑ Expression of genes associated with reverse cholesterol transport (RCT), including Ldlr and Scarb1 ↓ 4. Disrupted hepatic lipid homeostasis expression of two nuclear receptors, Srebp1c and Chrebp ↑ Acc1 gene and Cd36 gene ↑ | Hepatic lipid ↑, ceramide ↑, hosphatidylethanolamines ↑, phosphatidylserines (PS) ↑, phosphatidylcholines (PC) ↑↓ | Metabolomics and hepatic lipidomic UHPLC-ESI-TSQ Quantis triple quadrupole mass spectrometer | (Chi et al., 2024) [82] | |
| Acesulfame potassium ADI: 15 mg/kg Sucrose equivalence: 200× 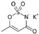 | Female: Carbohydrate metabolism ↓ Lipopolysaccharide synthesis ↑ Male: Carbohydrate adsorption and metabolism ↑ Lipopolysaccharide synthesis ↑ | Female: lactic acid ↓, succinic acid ↓, 2-Oleoylglycerol ↓ Male: pyruvic acid ↑, cholic acid ↑, deoxycholic acid ↓ | Fecal metabolomic LC-MS | (Bian et al., 2017) [68] |
| Neotame ADI: 18 mg/kg Sucrose equivalence: 7000–13,000× 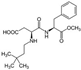 | Streptomycin biosynthesis ↑ Amino acid metabolism ↑ Folate biosynthesis ↑ Lipopolysaccharide biosynthesis ↑ Fatty acid metabolism ↓ Sporulation ↓ Benzoate degradation ↓ Carbohydrate metabolism ↓ Lipid metabolism ↓ Bacterial chemotaxis ↓ ABC transporters ↓ Butyrate fermentation pathways | Malic acid ↓, mannose-6-phosphate ↓, 5-aminovaleric acid ↓and glyceric acid ↓, 1,3-dipalmitate ↑, 1-monopalmitin ↑, linoleic acid↑, stearic acid ↑, cholesterol ↑, campesterol ↑, stigmastanol ↑ | Fecal metabolomic GC-MS | (Chi et al., 2018) [67] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, J.; Peng, J.; Hsiao, Y.-C.; Liu, C.-W.; Yang, Y.; Zhao, H.; Teitelbaum, T.; Wang, X.; Lu, K. Non/Low-Caloric Artificial Sweeteners and Gut Microbiome: From Perturbed Species to Mechanisms. Metabolites 2024, 14, 544. https://doi.org/10.3390/metabo14100544
Feng J, Peng J, Hsiao Y-C, Liu C-W, Yang Y, Zhao H, Teitelbaum T, Wang X, Lu K. Non/Low-Caloric Artificial Sweeteners and Gut Microbiome: From Perturbed Species to Mechanisms. Metabolites. 2024; 14(10):544. https://doi.org/10.3390/metabo14100544
Chicago/Turabian StyleFeng, Jiahao, Jingya Peng, Yun-Chung Hsiao, Chih-Wei Liu, Yifei Yang, Haoduo Zhao, Taylor Teitelbaum, Xueying Wang, and Kun Lu. 2024. "Non/Low-Caloric Artificial Sweeteners and Gut Microbiome: From Perturbed Species to Mechanisms" Metabolites 14, no. 10: 544. https://doi.org/10.3390/metabo14100544
APA StyleFeng, J., Peng, J., Hsiao, Y.-C., Liu, C.-W., Yang, Y., Zhao, H., Teitelbaum, T., Wang, X., & Lu, K. (2024). Non/Low-Caloric Artificial Sweeteners and Gut Microbiome: From Perturbed Species to Mechanisms. Metabolites, 14(10), 544. https://doi.org/10.3390/metabo14100544