
Donkey Colostrum and Milk: How Dietary Probiotics Can Affect Metabolomic Profile, Alkaline Sphingomyelinase and Alkaline Phosphatase Activity

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
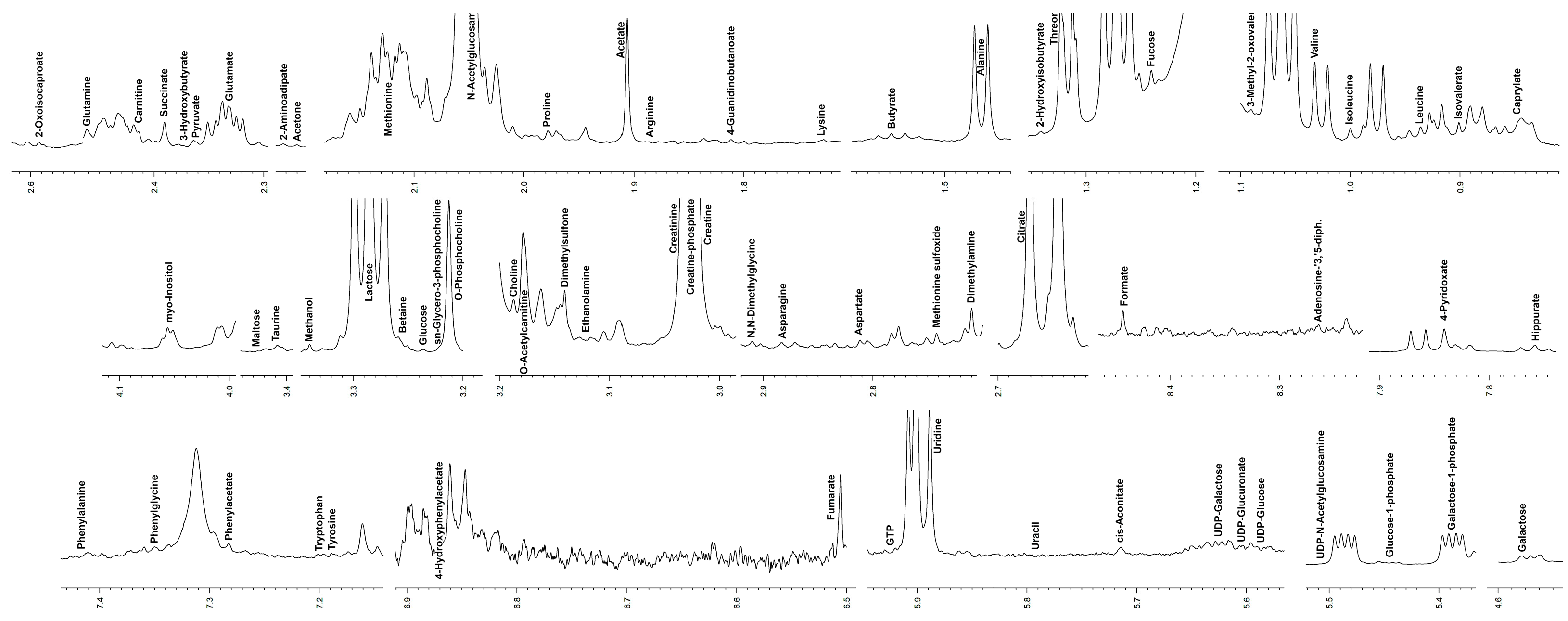
2.2. Metabolomic Analysis
2.3. Alkaline Sphingomyelinase Assay
2.4. Photometric Alkaline Phosphatase Assay
2.5. Statistical Analysis
3. Results
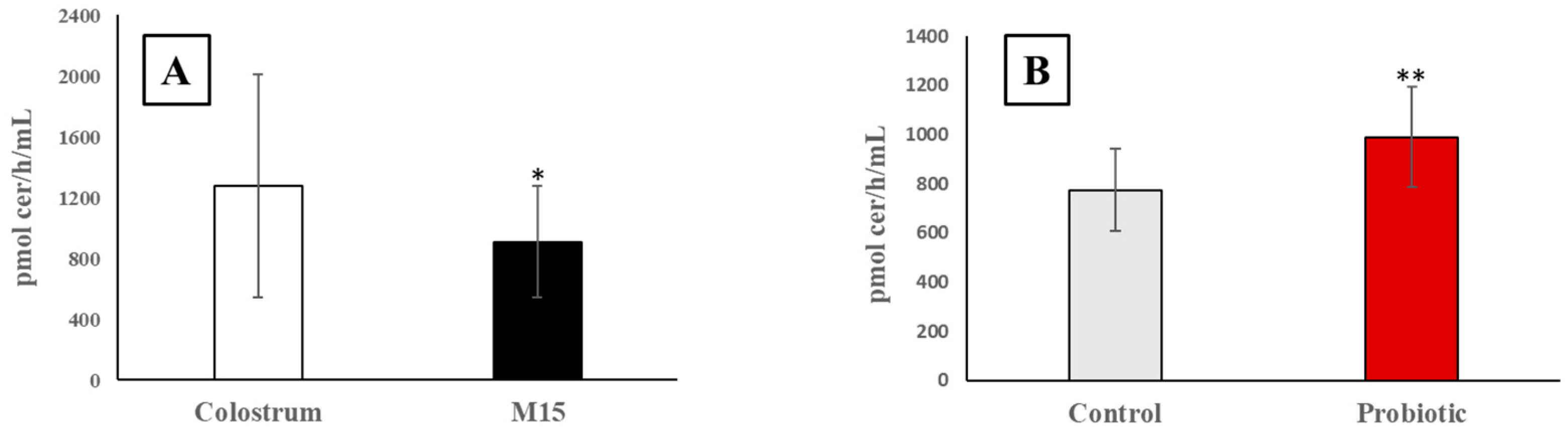
3.1. Alkaline Sphingomyelinase Activity in Colostrum and Milk
3.2. Alkaline Phosphate Activity in Colostrum and Milk
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Murgia, A.; Scano, P.; Contu, M.; Ibba, I.; Altea, M.; Bussu, M.; Demuru, M.; Porcu, A.; Caboni, P. Characterization of Donkey Milk and Metabolite Profile Comparison with Human Milk and Formula Milk. LWT 2016, 74, 427–433. [Google Scholar] [CrossRef]
- Bertino, E.; Agosti, M.; Peila, C.; Corridori, M.; Pintus, R.; Fanos, V. The Donkey Milk in Infant Nutrition. Nutrients 2022, 14, 403. [Google Scholar] [CrossRef] [PubMed]
- Martini, M.; Altomonte, I.; Licitra, R.; Salari, F. Nutritional and Nutraceutical Quality of Donkey Milk. J. Equine Vet. Sci. 2018, 65, 33–37. [Google Scholar] [CrossRef]
- Li, Y.; Ma, Q.-S.; Zhou, M.-M.; Zhang, Z.-W.; Zhan, Y.-D.; Liu, G.-Q.; Zhu, M.-X.; Wang, C.-F. A Metabolomics Comparison in Milk from Two Dezhou Donkey Strains. Eur. Food Res. Technol. 2022, 248, 1267–1275. [Google Scholar] [CrossRef]
- Di Salvo, E.; Conte, F.; Casciaro, M.; Gangemi, S.; Cicero, N. Bioactive Natural Products in Donkey and Camel Milk: A Perspective Review. Nat. Prod. Res. 2022, 1–15. [Google Scholar] [CrossRef]
- Živkov Baloš, M.; Ljubojević Pelić, D.; Jakšić, S.; Lazić, S. Donkey Milk: An Overview of Its Chemical Composition and Main Nutritional Properties or Human Health Benefit Properties. J. Equine Vet. Sci. 2023, 121, 104225. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ma, Q.; Liu, G.; Wang, C. Effects of Donkey Milk on Oxidative Stress and Inflammatory Response. J. Food Biochem. 2022, 46, e13935. [Google Scholar] [CrossRef] [PubMed]
- Garhwal, R.; Sangwan, K.; Mehra, R.; Kumar, N.; Bhardwaj, A.; Pal, Y.; Buttar, H.S.; Kumar, H. A Systematic Review of the Bioactive Components, Nutritional Qualities and Potential Therapeutic Applications of Donkey Milk. J. Equine Vet. Sci. 2022, 115. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Zhao, L.; Jiang, L.; Dong, M.L.; Ren, F.Z. The Antimicrobial Activity of Donkey Milk and Its Microflora Changes during Storage. Food Contro. 2008, 19, 1191–1195. [Google Scholar] [CrossRef]
- Mao, X.; Gu, J.; Sun, Y.; Xu, S.; Zhang, X.; Yang, H.; Ren, F. Anti-Proliferative and Anti-Tumour Effect of Active Components in Donkey Milk on A549 Human Lung Cancer Cells. Int. Dairy J. 2009, 19, 703–708. [Google Scholar] [CrossRef]
- Polidori, P.; Vincenzetti, S. Use of Donkey Milk in Children with Cow’s Milk Protein Allergy. Foods 2013, 2, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.Z.; Sun, H.Z.; Zhou, M.; Wang, O.; Chen, Y.; Liu, J.X.; Guan, L.L. Multi-Omics Reveals Functional Genomic and Metabolic Mechanisms of Milk Production and Quality in Dairy Cows. Bioinformatics 2020, 36, 2530–2537. [Google Scholar] [CrossRef] [PubMed]
- Laghi, L.; Picone, G.; Capozzi, F. Nuclear Magnetic Resonance for Foodomics beyond Food Analysis. Trends Anal. Chem. 2014, 59, 93–102. [Google Scholar] [CrossRef]
- Munekata, P.E.S.; Pateiro, M.; Rocchetti, G.; Domínguez, R.; Rocha, J.M.; Lorenzo, J.M. Application of Metabolomics to Decipher the Role of Bioactive Compounds in Plant and Animal Foods. Curr. Opin. Food Sci. 2022, 46. [Google Scholar] [CrossRef]
- Melzer, N.; Wittenburg, D.; Hartwig, S.; Jakubowski, S.; Kesting, U.; Willmitzer, L.; Lisec, J.; Reinsch, N.; Repsilber, D. Investigating Associations between Milk Metabolite Profiles and Milk Traits of Holstein Cows. J. Dairy Sci. 2013, 96, 1521–1534. [Google Scholar] [CrossRef]
- Lu, J.; Antunes Fernandes, E.; Páez Cano, A.E.; Vinitwatanakhun, J.; Boeren, S.; Van Hooijdonk, T.; Van Knegsel, A.; Vervoort, J.; Hettinga, K.A. Changes in Milk Proteome and Metabolome Associated with Dry Period Length, Energy Balance, and Lactation Stage in Postparturient Dairy Cows. J. Proteome Res. 2013, 12, 3288–3296. [Google Scholar] [CrossRef]
- O’Callaghan, T.F.; Vázquez-Fresno, R.; Serra-Cayuela, A.; Dong, E.; Mandal, R.; Hennessy, D.; McAuliffe, S.; Dillon, P.; Wishart, D.S.; Stanton, C.; et al. Pasture Feeding Changes the Bovine Rumen and Milk Metabolome. Metabolites 2018, 8, 27. [Google Scholar] [CrossRef]
- Scano, P.; Carta, P.; Ibba, I.; Manis, C.; Caboni, P. An Untargeted Metabolomic Comparison of Milk Composition from Sheep Kept Under Different Grazing Systems. Dairy 2020, 1, 30–41. [Google Scholar] [CrossRef]
- Manis, C.; Scano, P.; Nudda, A.; Carta, S.; Pulina, G.; Caboni, P. LC-QTOF/MS Untargeted Metabolomics of Sheep Milk under Cocoa Husks Enriched Diet. Dairy 2021, 2, 112–121. [Google Scholar] [CrossRef]
- Martini, M.; Altomonte, I.; Manica, E.; Salari, F. Changes in Donkey Milk Lipids in Relation to Season and Lactation. J. Food Compos. Anal. 2015, 41, 30–34. [Google Scholar] [CrossRef]
- Mecocci, S.; Gevi, F.; Pietrucci, D.; Cavinato, L.; Luly, F.R.; Pascucci, L.; Petrini, S.; Ascenzioni, F.; Zolla, L.; Chillemi, G.; et al. Anti-Inflammatory Potential of Cow, Donkey and Goat Milk Extracellular Vesicles as Revealed by Metabolomic Profile. Nutrients 2020, 12, 2908. [Google Scholar] [CrossRef]
- Britt, J.H.; Cushman, R.A.; Dechow, C.D.; Dobson, H.; Humblot, P.; Hutjens, M.F.; Jones, G.A.; Ruegg, P.S.; Sheldon, I.M.; Stevenson, J.S. Invited Review: Learning from the Future-A Vision for Dairy Farms and Cows in 2067. J. Dairy Sci. 2018, 101, 3722–3741. [Google Scholar] [CrossRef] [PubMed]
- Delia, E.; Tafaj, M.; Männer, K.; Delia, E.; Tafaj, M.; Männer, K. Efficiency of Probiotics in Farm Animals. Probiotic. Anim. 2012. [Google Scholar] [CrossRef]
- Pellegrino, M.S.; Frola, I.D.; Natanael, B.; Gobelli, D.; Nader-Macias, M.E.F.; Bogni, C.I. In Vitro Characterization of Lactic Acid Bacteria Isolated from Bovine Milk as Potential Probiotic Strains to Prevent Bovine Mastitis. Probiotics Antimicrob Proteins 2019, 11, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, R.S.; de Silva, L.C.S.; de Souza, M.R.; Reis, R.B.; Bicalho, A.F.; Nunes, J.P.S.; Dias, A.A.M.; Nicoli, J.R.; Neumann, E.; Nunes, C. Prospecting of Potentially Probiotic Lactic Acid Bacteria from Bovine Mammary Ecosystem: Imminent Partners from Bacteriotherapy against Bovine Mastitis. Int. Microbiol. 2022, 25, 189–206. [Google Scholar] [CrossRef] [PubMed]
- Moallem, U.; Lehrer, H.; Livshitz, L.; Zachut, M.; Yakoby, S. The Effects of Live Yeast Supplementation to Dairy Cows during the Hot Season on Production, Feed Efficiency, and Digestibility. J. Dairy Sci. 2009, 92, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Wang, J.Q.; Kang, H.Y.; Dong, S.H.; Sun, P.; Bu, D.P.; Zhou, L.Y. Effect of Feeding Bacillus Subtilis Natto Fermentation Product on Milk Production and Composition, Blood Metabolites and Rumen Fermentation in Early Lactation Dairy Cows. J. Anim. Physiol. Anim. Nutr. 2012, 96, 506–512. [Google Scholar] [CrossRef] [PubMed]
- So, S.; Wanapat, M.; Cherdthong, A. Effect of Sugarcane Bagasse as Industrial By-Products Treated with Lactobacillus Casei TH14, Cellulase and Molasses on Feed Utilization, Ruminal Ecology and Milk Production of Mid-Lactating Holstein Friesian Cows. J. Sci. Food Agric. 2021, 101, 4481–4489. [Google Scholar] [CrossRef]
- Laghi, L.; Zhu, C.; Campagna, G.; Rossi, G.; Bazzano, M.; Laus, F. Probiotic Supplementation in Trained Trotter Horses: Effect on Blood Clinical Pathology Data and Urine Metabolomic Assessed in Field. J. Appl. Physiol. 2018, 125, 654. [Google Scholar] [CrossRef]
- Schoster, A.; Weese, J.S.; Guardabassi, L. Probiotic Use in Horses—What Is the Evidence for Their Clinical Efficacy? J. Vet. Intern. Med. 2014, 28, 1640–1652. [Google Scholar] [CrossRef]
- Iqbal, J.; Walsh, M.T.; Hammad, S.M.; Hussain, M.M. Sphingolipids and Lipoproteins in Health and Metabolic Disorders. Trends Endocrinol. Metab. 2017, 28, 506. [Google Scholar] [CrossRef] [PubMed]
- Marzo, F.; Jauregui, P.; Barrenetxe, J.; Martínez-Peñuela, A.; Ibañez, F.C.; Milagro, F.I. Effect of a Diet Supplemented with Sphingomyelin and Probiotics on Colon Cancer Development in Mice. Probiotics Antimicrob. Proteins 2022, 14, 407. [Google Scholar] [CrossRef] [PubMed]
- Duan, R.D.; Bergman, T.; Xu, N.; Wu, J.; Cheng, Y.; Duan, J.; Nelander, S.; Palmberg, C.; Nilsson, Å. Identification of Human Intestinal Alkaline Sphingomyelinase as a Novel Ecto-Enzyme Related to the Nucleotide Phosphodiesterase Family. J. Biol. Chem. 2003, 278, 38528–38536. [Google Scholar] [CrossRef] [PubMed]
- Duan, R.D. Alkaline Sphingomyelinase: An Old Enzyme with Novel Implications. Biochim. Biophys. Acta 2006, 1761, 281–291. [Google Scholar] [CrossRef]
- Di Marzio, L.; Di Leo, A.; Cinque, B.; Fanini, D.; Agnifili, A.; Berloco, P.; Linsalata, M.; Lorusso, D.; Barone, M.; De Simone, C.; et al. Detection of Alkaline Sphingomyelinase Activity in Human Stool: Proposed Role as a New Diagnostic and Prognostic Marker of Colorectal Cancer. Cancer Epidemiol. Biomark. Prev. 2005, 14, 856–862. [Google Scholar] [CrossRef]
- Soo, I.; Madsen, K.L.; Tejpar, Q.; Sydora, B.C.; Sherbaniuk, R.; Cinque, B.; di Marzio, L.; Cifone, M.G.; Desimone, C.; Fedorak, R.N. VSL#3 Probiotic Upregulates Intestinal Mucosal Alkaline Sphingomyelinase and Reduces Inflammation. Can. J. Gastroenterol. 2008, 22, 237–242. [Google Scholar] [CrossRef]
- Duan, R.D.; Cheng, Y.; Jönsson, B.A.G.; Ohlsson, L.; Herbst, A.; Hellström-Westas, L.; Nilsson, Å. Human Meconium Contains Significant Amounts of Alkaline Sphingomyelinase, Neutral Ceramidase, and Sphingolipid Metabolites. Pediatr. Res. 2007, 61, 61–66. [Google Scholar] [CrossRef]
- Nilsson, Å.; Duan, R.D.; Ohlsson, L. Digestion and Absorption of Milk Phospholipids in Newborns and Adults. Front. Nutr. 2021, 8, 532. [Google Scholar] [CrossRef]
- Rankin, S.A.; Christiansen, A.; Lee, W.; Banavara, D.S.; Lopez-Hernandez, A. Invited Review: The Application of Alkaline Phosphatase Assays for the Validation of Milk Product Pasteurization. J. Dairy Sci. 2010, 93, 5538–5551. [Google Scholar] [CrossRef]
- Clawin-Rädecker, I.; De Block, J.; Egger, L.; Willis, C.; Da Silva Felicio, M.T.; Messens, W. The Use of Alkaline Phosphatase and Possible Alternative Testing to Verify Pasteurisation of Raw Milk, Colostrum, Dairy and Colostrum-Based Products. EFSA J 2021, 19, e06576. [Google Scholar] [CrossRef]
- Marchand, S.; Merchiers, M.; Messens, W.; Coudijzer, K.; De Block, J. Thermal Inactivation Kinetics of Alkaline Phosphatase in Equine Milk. Int. Dairy J. 2009, 19, 763–767. [Google Scholar] [CrossRef]
- Giacometti, F.; Bardasi, L.; Merialdi, G.; Morbarigazzi, M.; Federici, S.; Piva, S.; Serraino, A. Shelf Life of Donkey Milk Subjected to Different Treatment and Storage Conditions. J. Dairy Sci. 2016, 99, 4291–4299. [Google Scholar] [CrossRef] [PubMed]
- Evans, L.; Crane, M. The Clinical Companion of the Donkey, 1st ed.; Evans, L., Crane, M., Eds.; The Donkey Sanctuary: Sidmouth, UK, 2018; ISBN 978-1789013-900. [Google Scholar]
- Gur, M.; Zuckerman-Levin, N.; Masarweh, K.; Hanna, M.; Laghi, L.; Marazzato, M.; Levanon, S.; Hakim, F.; Bar–Yoseph, R.; Wilschanski, M.; et al. The Effect of Probiotic Administration on Metabolomics and Glucose Metabolism in CF Patients. Pediatr. Pulmonol. 2022, 57, 2335–2343. [Google Scholar] [CrossRef] [PubMed]
- Dieterle, F.; Ross, A.; Schlotterbeck, G.; Senn, H. Probabilistic Quotient Normalization as Robust Method to Account for Dilution of Complex Biological Mixtures. Application In1H NMR Metabonomics. Anal. Chem. 2006, 78, 4281–4290. [Google Scholar] [CrossRef] [PubMed]
- Loidl, A.; Claus, R.; Deigner, H.P.; Hermetter, A. High-Precision Fluorescence Assay for Sphingomyelinase Activity of Isolated Enzymes and Cell Lysates. J. Lipid Res. 2002, 43, 815–823. [Google Scholar] [CrossRef]
- Xia, Y.; Yu, J.; Miao, W.; Shuang, Q. A UPLC-Q-TOF-MS-Based Metabolomics Approach for the Evaluation of Fermented Mare’s Milk to Koumiss. Food Chem 2020, 320, 126619. [Google Scholar] [CrossRef]
- Li, M.; Kang, S.; Zheng, Y.; Shao, J.; Zhao, H.; An, Y.; Cao, G.; Li, Q.; Yue, X.; Yang, M. Comparative Metabolomics Analysis of Donkey Colostrum and Mature Milk Using Ultra-High-Performance Liquid Tandem Chromatography Quadrupole Time-of-Flight Mass Spectrometry. J. Dairy Sci. 2020, 103, 992–1001. [Google Scholar] [CrossRef]
- Altomonte, I.; Salari, F.; Licitra, R.; Martini, M. Donkey and Human Milk: Insights into Their Compositional Similarities. Int. Dairy J. 2019, 89, 111–118. [Google Scholar] [CrossRef]
- Medhammar, E.; Wijesinha-Bettoni, R.; Stadlmayr, B.; Nilsson, E.; Charrondiere, U.R.; Burlingame, B. Composition of Milk from Minor Dairy Animals and Buffalo Breeds: A Biodiversity Perspective. J. Sci. Food Agric. 2012, 92, 445–474. [Google Scholar] [CrossRef]
- Malacarne, M.; Criscione, A.; Franceschi, P.; Bordonaro, S.; Formaggioni, P.; Marletta, D.; Summer, A. New Insights into Chemical and Mineral Composition of Donkey Milk throughout Nine Months of Lactation. Animals 2019, 9, 1161. [Google Scholar] [CrossRef]
- O’Callaghan, T.F.; O’Donovan, M.; Murphy, J.P.; Sugrue, K.; Tobin, J.T.; McNamara, A.E.; Yin, X.; Sundaramoorthy, G.; Brennan, L. The Bovine Colostrum and Milk Metabolome at the Onset of Lactation as Determined by 1H-NMR. Int. Dairy J. 2021, 113. [Google Scholar] [CrossRef]
- Lin, Y.; Sun, X.; Hou, X.; Qu, B.; Gao, X.; Li, Q. Effects of Glucose on Lactose Synthesis in Mammary Epithelial Cells from Dairy Cow. BMC Vet. Res. 2016, 12, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Lopez-Villalobos, N.; Sneddon, N.W.; Shalloo, L.; Franzoi, M.; De Marchi, M.; Penasa, M. Invited Review: Milk Lactose-Current Status and Future Challenges in Dairy Cattle. J. Dairy Sci. 2019, 102, 5883–5898. [Google Scholar] [CrossRef]
- Arumugam, M.K.; Paal, M.C.; Donohue, T.M.; Ganesan, M.; Osna, N.A.; Kharbanda, K.K. Beneficial Effects of Betaine: A Comprehensive Review. Biology 2021, 10, 456. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.; Cabrera-Ghayouri, S.; Christie, L.-A.; Held, K.S.; Viswanath, V. Translational Preclinical Pharmacologic Disease Models for Ophthalmic Drug Development. Pharm. Res. 2019, 36, 58. [Google Scholar] [CrossRef] [PubMed]
- Holmes-McNary, M.Q.; Cheng, W.L.; Mar, M.H.; Fussell, S.; Zeisel, S.H. Choline and Choline Esters in Human and Rat Milk and in Infant Formulas. Am. J. Clin. Nutr 1996, 64, 572–576. [Google Scholar] [CrossRef]
- Kim, Y.H.; Kim, D.H.; Lim, H.; Baek, D.Y.; Shin, H.K.; Kim, J.K. The Anti-Inflammatory Effects of Methylsulfonylmethane on Lipopolysaccharide-Induced Inflammatory Responses in Murine Macrophages. Biol. Pharm. Bull 2009, 32, 651–656. [Google Scholar] [CrossRef]
- Branco Lopes, R.; Bernal-Cordoba, C.; Fausak, E.D.; Silva-Del-Rio, N. Effect of Prebiotics on Growth and Health of Dairy Calves: A Protocol for a Systematic Review and Meta-Analysis. PLoS ONE 2021, 16, e0253379. [Google Scholar] [CrossRef]
- Nalla, K.; Manda, N.K.; Dhillon, H.S.; Kanade, S.R.; Rokana, N.; Hess, M.; Puniya, A.K. Impact of Probiotics on Dairy Production Efficiency. Front. Microbiol 2022, 13, 1519. [Google Scholar] [CrossRef]
- Fox, P.F. Book: Chemistry And Biochemistry of Dairy. In Handbook of Seafood and Seafood Products Analysis; CRC Press: Boca Raton, FL, USA, 2015; p. 1. [Google Scholar]
- Guilloteau, P.; Martin, L.; Eeckhaut, V.; Ducatelle, R.; Zabielski, R.; Van Immerseel, F. From the Gut to the Peripheral Tissues: The Multiple Effects of Butyrate. Nutr. Res. Rev. 2010, 23, 366–384. [Google Scholar] [CrossRef]
- Meijer, K.; De Vos, P.; Priebe, M.G. Butyrate and Other Short-Chain Fatty Acids as Modulators of Immunity: What Relevance for Health? Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Elamin, E.E.; Masclee, A.A.; Dekker, J.; Pieters, H.J.; Jonkers, D.M. Short-Chain Fatty Acids Activate AMP-Activated Protein Kinase and Ameliorate Ethanol-Induced Intestinal Barrier Dysfunction in Caco-2 Cell Monolayers. J. Nutr. 2013, 143, 1872–1881. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, J.; He, T.; Becker, S.; Zhang, G.; Li, D.; Ma, X. Butyrate: A Double-Edged Sword for Health? Adv. Nutr. 2018, 9, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Cant, J.P.; Kim, J.J.M.; Cieslar, S.R.L.; Doelman, J. Symposium Review: Amino Acid Uptake by the Mammary Glands: Where Does the Control Lie? J. Dairy Sci. 2018, 101, 5655–5666. [Google Scholar] [CrossRef] [PubMed]
- Nemati, M.; Menatian, S.; Ghasemi, S.J.; Hooshmandfar, R.; Taheri, M.; Saifi, T. Effect of Protected-Glutamine Supplementation on Performance, Milk Composition and Some Blood Metabolites in Fresh Holstein Cows. Iran. J. Vet. Res. 2018, 19, 225. [Google Scholar]
- Newsholme, P.; Procopio, J.; Ramos Lima, M.M.; Pithon-Curi, T.C.; Curi, R. Glutamine and Glutamate-Their Central Role in Cell Metabolism and Function. Cell Biochem. Funct. 2003, 21, 1–9. [Google Scholar] [CrossRef]
- Phang, J.M.; Liu, W.; Zabirnyk, O. Proline Metabolism and Microenvironmental Stress. Annu. Rev. Nutr. 2010, 30, 441–463. [Google Scholar] [CrossRef]
- Wu, J.; Liu, F.; Nilsson, Å.; Duan, R.D. Pancreatic Trypsin Cleaves Intestinal Alkaline Sphingomyelinase from Mucosa and Enhances the Sphingomyelinase Activity. Am. J. Physiol. Gastrointest Liver Physiol. 2004, 287, G967–G973. [Google Scholar] [CrossRef]
- Allison, M.J.; Bryant, M.P.; Doetsch, R.N. Studies on the Metabolic Function of Branched-Chain Volatile Fatty Acids, Growth Factors for Ruminococci. I. Incorporation of Isovalerate into Leucine. J. Bacteriol. 1962, 83, 523–532. [Google Scholar] [CrossRef]
- Vranković, L.; Aladrović, J.; Octenjak, D.; Bijelić, D.; Cvetnić, L.; Stojević, Z. Milk Fatty Acid Composition as an Indicator of Energy Status in Holstein Dairy Cows. Arch. Anim. Breed. 2017, 60, 205–212. [Google Scholar] [CrossRef]
- Zhang, P.; Chen, Y.; Cheng, Y.; Hertervig, E.; Ohlsson, L.; Nilsson, Å.; Duan, R.D. Alkaline Sphingomyelinase (NPP7) Promotes Cholesterol Absorption by Affecting Sphingomyelin Levels in the Gut: A Study with NPP7 Knockout Mice. Am. J. Physiol. Gastrointest Liver Physiol. 2014, 306, 903–908. [Google Scholar] [CrossRef] [PubMed]
- Tsiamita, A.; Valiakos, G.; Natsaridis, N.; Fotiadou, S.; Manouras, A.; Malissiova, E. Preliminary Results on the Comparative Evaluation of Alkaline Phosphatase Commercial Tests Efficiency in Non-Cow Milk Pasteurization. BioTech 2022, 11, 39. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Bacterial Strain in SLAB51® Blend | Total Number of CFU for 18 g |
|---|---|
| Streptococcus thermophilus DSM 32245/CNCM I-5570 | 960 Billion |
| Lactobacillus brevis DSM 27961/CNCM I-5566 | 432 Billion |
| Bifidobacterium lactis DSM 32246/CNCM I-5571 | 300 Billion |
| Bifidobacterium lactis DSM 32247/CNCM I-5572 | 300 Billion |
| Lactobacillus plantarum DSM 32244/CNCM I-5569 | 192 Billion |
| Lactobacillus paracasei DSM 32243/CNCM I-5568 | 144 Billion |
| Lactobacillus acidophilus DSM 32241/CNCM I-5567 | 60 Billion |
| Lactobacillus helveticus DSM 32242/CNCM I-5573 | 12 Billion |
| Molecule (mmol/L) | Colostrum (C) | Milk (M15) | p Values |
|---|---|---|---|
| Formate | 0.017 ± 0.006 | 0.014 ± 0.003 | 0.102 |
| 4-Pyridoxate | 0.176 ± 0.101 | 0.189 ± 0.078 | 0.488 |
| Hippurate | 0.172 ± 0.107 | 0.172 ± 0.067 | 0.992 |
| Phenylglycine | 0.010 ± 0.011 | 0.048 ± 0.108 | 0.123 |
| Phenylacetate | 0.037 ± 0.020 | 0.035 ± 0.012 | 0.709 |
| Tyrosine | 0.026 ± 0.009 | 0.026 ± 0.005 | 0.906 |
| 4-Hydroxyphenylacetate | 0.028 ± 0.008 | 0.032 ± 0.009 | 0.181 |
| Fumarate | 0.011 ± 0.004 | 0.008 ± 0.002 | 0.002 |
| Uridine | 0.565 ± 0.322 | 0.187 ± 0.190 | 0.000 |
| cis-Aconitate | 0.034 ± 0.018 | 0.027 ± 0.005 | 0.163 |
| UDP-glucose | 0.084 ± 0.084 | 0.061 ± 0.011 | 0.241 |
| Glucose-1-phosphate | 0.128 ± 0.074 | 0.089 ± 0.094 | 0.111 |
| Galactose-1-phosphate | 1.635 ± 0.407 | 1.251 ± 0.385 | 0.007 |
| Galactose | 0.888 ± 0.096 | 0.801 ± 0.134 | 0.018 |
| myo-Inositol | 1.226 ± 0.518 | 1.780 ± 0.594 | 0.006 |
| Maltose | 0.130 ± 0.058 | 0.136 ± 0.038 | 0.653 |
| Taurine | 1.004 ± 0.237 | 1.026 ± 0.541 | 0.855 |
| Methanol | 0.047 ± 0.019 | 0.052 ± 0.041 | 0.617 |
| Lactose | 162.428 ± 33.321 | 165.190 ± 33.996 | 0.725 |
| Betaine | 0.032 ± 0.025 | 0.063 ± 0.066 | 0.035 |
| TMAO | 0.026 ± 0.016 | 0.020 ± 0.012 | 0.173 |
| Glucose | 0.689 ± 0.218 | 0.673 ± 0.219 | 0.802 |
| sn-Glycero-3-phosphocholine | 1.818 ± 0.546 | 1.450 ± 0.410 | 0.029 |
| O-Phosphocholine | 0.245 ± 0.183 | 0.148 ± 0.171 | 0.045 |
| Choline | 0.035 ± 0.051 | 0.032 ± 0.048 | 0.845 |
| O-Acetylcarnitine | 0.080 ± 0.037 | 0.056 ± 0.020 | 0.017 |
| Dimethyl sulfone | 0.027 ± 0.010 | 0.017 ± 0.007 | 0.003 |
| Malonate | 0.011 ± 0.005 | 0.013 ± 0.006 | 0.182 |
| Creatinine | 0.087 ± 0.071 | 0.066 ± 0.040 | 0.155 |
| Creatine-phosphate | 1.259 ± 0.437 | 0.920 ± 0.400 | 0.023 |
| Creatine | 0.637 ± 0.148 | 0.724 ± 0.215 | 0.030 |
| N.N-Dimethylglycine | 0.003 ± 0.001 | 0.003 ± 0.000 | 0.053 |
| Asparagine | 0.075 ± 0.026 | 0.089 ± 0.022 | 0.110 |
| Aspartate | 0.631 ± 1.027 | 0.262 ± 0.262 | 0.133 |
| Methionine sulfoxide | 0.007 ± 0.002 | 0.006 ± 0.001 | 0.056 |
| Dimethylamine | 0.007 ± 0.003 | 0.008 ± 0.003 | 0.239 |
| Citrate | 5.662 ± 2.117 | 5.284 ± 1.435 | 0.517 |
| 2-Oxoisocaproate | 0.039 ± 0.029 | 0.047 ± 0.022 | 0.272 |
| Glutamine | 0.804 ± 0.881 | 1.069 ± 0.792 | 0.347 |
| Carnitine | 0.154 ± 0.055 | 0.130 ± 0.066 | 0.238 |
| Succinate | 0.074 ± 0.024 | 0.066 ± 0.011 | 0.161 |
| Pyruvate | 0.016 ± 0.012 | 0.021 ± 0.019 | 0.334 |
| Glutamate | 0.757 ± 0.354 | 1.144 ± 0.339 | 0.000 |
| 2-Aminoadipate | 0.037 ± 0.020 | 0.037 ± 0.013 | 0.930 |
| Acetone | 0.004 ± 0.001 | 0.005 ± 0.001 | 0.005 |
| Methionine | 0.127 ± 0.108 | 0.153 ± 0.136 | 0.534 |
| N-Acetylglucosamine | 0.273 ± 0.488 | 0.208 ± 0.247 | 0.565 |
| Proline | 0.233 ± 0.115 | 0.226 ± 0.100 | 0.839 |
| Acetate | 0.134 ± 0.072 | 0.136 ± 0.078 | 0.924 |
| 4-Guanidinobutanoate | 0.039 ± 0.011 | 0.047 ± 0.016 | 0.079 |
| Butyrate | 0.051 ± 0.095 | 0.085 ± 0.077 | 0.182 |
| Alanine | 0.202 ± 0.072 | 0.263 ± 0.087 | 0.011 |
| 2-Hydroxyisobutyrate | 0.004 ± 0.002 | 0.003 ± 0.001 | 0.051 |
| Threonine | 0.383 ± 0.333 | 0.730 ± 1.062 | 0.173 |
| Lactate | 0.775 ± 1.544 | 1.672 ± 4.603 | 0.410 |
| Fucose | 0.081 ± 0.030 | 0.086 ± 0.055 | 0.736 |
| Ethanol | 286.591 ± 46.471 | 261.203 ± 43.007 | 0.042 |
| Valine | 0.110 ± 0.063 | 0.148 ± 0.050 | 0.033 |
| Isoleucine | 0.028 ± 0.028 | 0.043 ± 0.022 | 0.079 |
| Leucine | 0.035 ± 0.028 | 0.041 ± 0.023 | 0.491 |
| Isovalerate | 0.038 ± 0.025 | 0.039 ± 0.019 | 0.965 |
| Caprylate | 0.271 ± 0.185 | 0.635 ± 0.484 | 0.004 |
| TSP | 6.460 ± 2.342 | 6.088 ± 1.282 | 0.506 |
| Molecule (mmol/L) | Group B (Supplemented) | Group A (Control) | p Values |
|---|---|---|---|
| Formate | 0.016 ± 0.004 | 0.014 ± 0.002 | 0.075 |
| 4-Pyridoxate | 0.183 ± 0.071 | 0.267 ± 0.032 | 0.001 |
| Hippurate | 0.164 ± 0.043 | 0.161 ± 0.102 | 0.465 |
| Phenylglycine | 0.017 ± 0.017 | 0.009 ± 0.008 | 0.092 |
| Phenylacetate | 0.036 ± 0.008 | 0.033 ± 0.007 | 0.182 |
| Tyrosine | 0.026 ± 0.003 | 0.030 ± 0.009 | 0.118 |
| 4-Hydroxyphenylacetate | 0.033 ± 0.007 | 0.024 ± 0.009 | 0.010 |
| Fumarate | 0.009 ± 0.002 | 0.008 ± 0.002 | 0.101 |
| Uridine | 0.145 ± 0.097 | 0.139 ± 0.222 | 0.465 |
| cis-Aconitate | 0.032 ± 0.006 | 0.030 ± 0.006 | 0.190 |
| UDP-glucose | 0.070 ± 0.009 | 0.065 ± 0.018 | 0.173 |
| Glucose-1-phosphate | 0.082 ± 0.064 | 0.036 ± 0.025 | 0.022 |
| Galactose-1-phosphate | 1.339 ± 0.150 | 1.178 ± 0.305 | 0.067 |
| Galactose | 0.888 ± 0.170 | 0.927 ± 0.142 | 0.288 |
| myo-Inositol | 1.870 ± 0.509 | 2.048 ± 0.124 | 0.148 |
| Maltose | 0.159 ± 0.046 | 0.136 ± 0.062 | 0.176 |
| Taurine | 0.794 ± 0.255 | 0.661 ± 0.180 | 0.093 |
| Methanol | 0.036 ± 0.009 | 0.047 ± 0.022 | 0.078 |
| Lactose | 177.382 ± 13.310 | 193.809 ± 9.391 | 0.002 |
| Betaine | 0.042 ± 0.018 | 0.051 ± 0.008 | 0.073 |
| TMAO | 0.011 ± 0.006 | 0.016 ± 0.012 | 0.107 |
| Glucose | 0.714 ± 0.170 | 0.551 ± 0.197 | 0.028 |
| sn-Glycero-3-phosphocholine | 1.345 ± 0.160 | 1.557 ± 0.329 | 0.036 |
| O-Phosphocholine | 0.068 ± 0.050 | 0.172 ± 0.150 | 0.022 |
| Choline | 0.014 ± 0.006 | 0.022 ± 0.015 | 0.056 |
| O-Acetylcarnitine | 0.055 ± 0.018 | 0.050 ± 0.015 | 0.232 |
| Dimethyl sulfone | 0.015 ± 0.006 | 0.019 ± 0.008 | 0.078 |
| Malonate | 0.011 ± 0.005 | 0.012 ± 0.003 | 0.491 |
| Creatinine | 0.050 ± 0.018 | 0.061 ± 0.018 | 0.089 |
| Creatine-phosphate | 0.892 ± 0.159 | 0.925 ± 0.218 | 0.347 |
| Creatine | 0.821 ± 0.172 | 0.766 ± 0.133 | 0.210 |
| N.N-Dimethylglycine | 0.003 ± 0.000 | 0.003 ± 0.001 | 0.114 |
| Asparagine | 0.093 ± 0.024 | 0.076 ± 0.030 | 0.081 |
| Aspartate | 0.233 ± 0.213 | 1.061 ± 2.710 | 0.162 |
| Methionine sulfoxide | 0.006 ± 0.000 | 0.006 ± 0.001 | 0.411 |
| Dimethylamine | 0.009 ± 0.003 | 0.007 ± 0.003 | 0.054 |
| Citrate | 4.284 ± 1.180 | 4.589 ± 1.378 | 0.296 |
| 2-Oxoisocaproate | 0.051 ± 0.014 | 0.030 ± 0.010 | 0.000 |
| Glutamine | 1.255 ± 0.308 | 0.841 ± 0.541 | 0.021 |
| Carnitine | 0.152 ± 0.119 | 0.110 ± 0.046 | 0.154 |
| Succinate | 0.061 ± 0.012 | 0.058 ± 0.014 | 0.328 |
| Pyruvate | 0.014 ± 0.005 | 0.014 ± 0.002 | 0.449 |
| Glutamate | 1.257 ± 0.207 | 1.191 ± 0.213 | 0.243 |
| 2-Aminoadipate | 0.041 ± 0.015 | 0.035 ± 0.009 | 0.143 |
| Acetone | 0.005 ± 0.001 | 0.004 ± 0.001 | 0.152 |
| Methionine | 0.097 ± 0.032 | 0.110 ± 0.111 | 0.351 |
| N-Acetylglucosamine | 0.119 ± 0.049 | 0.137 ± 0.119 | 0.328 |
| Proline | 0.162 ± 0.057 | 0.206 ± 0.095 | 0.105 |
| Acetate | 0.134 ± 0.057 | 0.117 ± 0.058 | 0.263 |
| 4-Guanidinobutanoate | 0.044 ± 0.010 | 0.041 ± 0.011 | 0.241 |
| Butyrate | 0.201 ± 0.190 | 0.070 ± 0.046 | 0.024 |
| Alanine | 0.277 ± 0.071 | 0.228 ± 0.079 | 0.076 |
| 2-Hydroxyisobutyrate | 0.004 ± 0.002 | 0.003 ± 0.001 | 0.107 |
| Threonine | 0.336 ± 0.111 | 0.393 ± 0.128 | 0.146 |
| Lactate | 0.242 ± 0.083 | 0.287 ± 0.124 | 0.167 |
| Fucose | 0.114 ± 0.091 | 0.098 ± 0.034 | 0.308 |
| Ethanol | 268.419 ± 20.585 | 317.981 ± 20.994 | 0.000 |
| Valine | 0.140 ± 0.029 | 0.123 ± 0.037 | 0.131 |
| Isoleucine | 0.032 ± 0.012 | 0.032 ± 0.009 | 0.482 |
| Leucine | 0.030 ± 0.010 | 0.034 ± 0.011 | 0.210 |
| Isovalerate | 0.045 ± 0.016 | 0.020 ± 0.011 | 0.000 |
| Caprylate | 0.728 ± 0.342 | 0.408 ± 0.164 | 0.007 |
| TSP | 6.789 ± 0.468 | 7.438 ± 0.623 | 0.007 |
| Sample | Colostrum | M15 | Milk at 45 Days (Control Group) | Milk at 45 Days (Probiotic Group) |
|---|---|---|---|---|
| ALP (U/mL) | N.D. | N.D. | N.D. | Range 0.97–11.73 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laus, F.; Laghi, L.; Bazzano, M.; Cifone, M.G.; Cinque, B.; Yang, Y.; Marchegiani, A. Donkey Colostrum and Milk: How Dietary Probiotics Can Affect Metabolomic Profile, Alkaline Sphingomyelinase and Alkaline Phosphatase Activity. Metabolites 2023, 13, 622. https://doi.org/10.3390/metabo13050622
Laus F, Laghi L, Bazzano M, Cifone MG, Cinque B, Yang Y, Marchegiani A. Donkey Colostrum and Milk: How Dietary Probiotics Can Affect Metabolomic Profile, Alkaline Sphingomyelinase and Alkaline Phosphatase Activity. Metabolites. 2023; 13(5):622. https://doi.org/10.3390/metabo13050622
Chicago/Turabian StyleLaus, Fulvio, Luca Laghi, Marilena Bazzano, Maria Grazia Cifone, Benedetta Cinque, Yaosen Yang, and Andrea Marchegiani. 2023. "Donkey Colostrum and Milk: How Dietary Probiotics Can Affect Metabolomic Profile, Alkaline Sphingomyelinase and Alkaline Phosphatase Activity" Metabolites 13, no. 5: 622. https://doi.org/10.3390/metabo13050622
APA StyleLaus, F., Laghi, L., Bazzano, M., Cifone, M. G., Cinque, B., Yang, Y., & Marchegiani, A. (2023). Donkey Colostrum and Milk: How Dietary Probiotics Can Affect Metabolomic Profile, Alkaline Sphingomyelinase and Alkaline Phosphatase Activity. Metabolites, 13(5), 622. https://doi.org/10.3390/metabo13050622