


Network Pharmacology and Experimental Verification to Unveil the Mechanism of N-Methyl-D-Aspartic Acid Rescue Humantenirine-Induced Excitotoxicity

Abstract
1. Introduction
2. Materials and Methods
2.1. Collection and Screening of Gelsemium Alkaloids
2.2. Screening of Targets Corresponding to the Alkaloid Components
2.3. Identification of Targets Related to Excitotoxicity
2.4. Prediction of Targets of Alkaloid Components Associated with Excitotoxicity
2.5. Protein-Protein Interaction (PPI) Network Construction and Analysis
2.6. Gene Ontology (GO) and Kyoto Encyclopedia Genes Genomes (KEGG) Pathway Enrichment Analysis
2.7. Construction of the Alkaloid–Target–Pathway Network
2.8. Molecular Docking
2.9. Animal Experiments
2.9.1. The LD50 of Acute Humantenirine Poisoning
2.9.2. The Antidotic Effect of NMDA on Humantenirine Poisoning
2.9.3. Drug Treatment and Sample Collection
Measurement of ATP Content in the Hippocampal Tissue of Mice
Determination of Mitochondrial Membrane Potential
Detection of the Expression of Key Protein in Mice Hippocampus by Western Blotting
3. Results
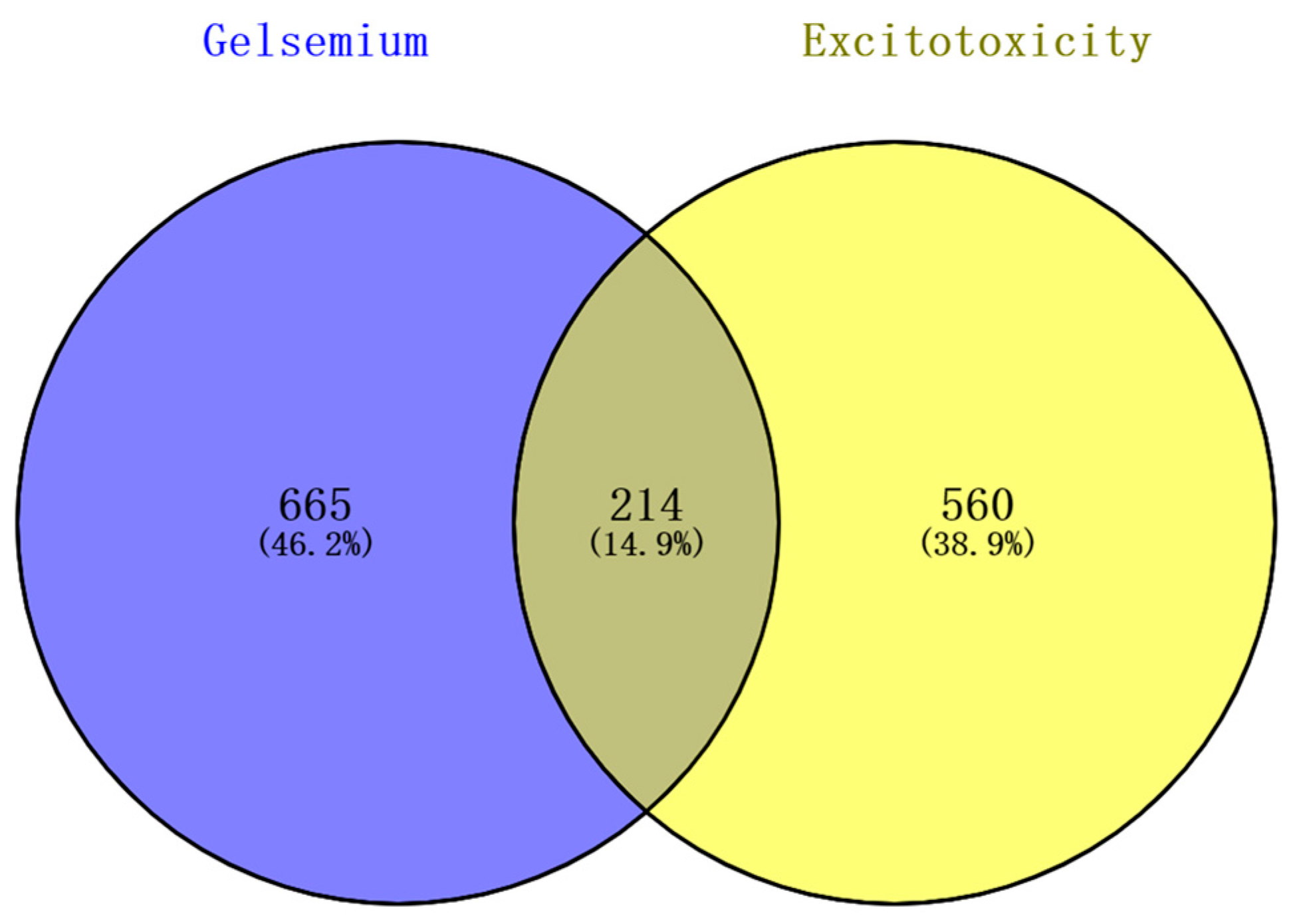
3.1. Putative Targets of Gelsemium Alkaloids Associated with Excitotoxicity
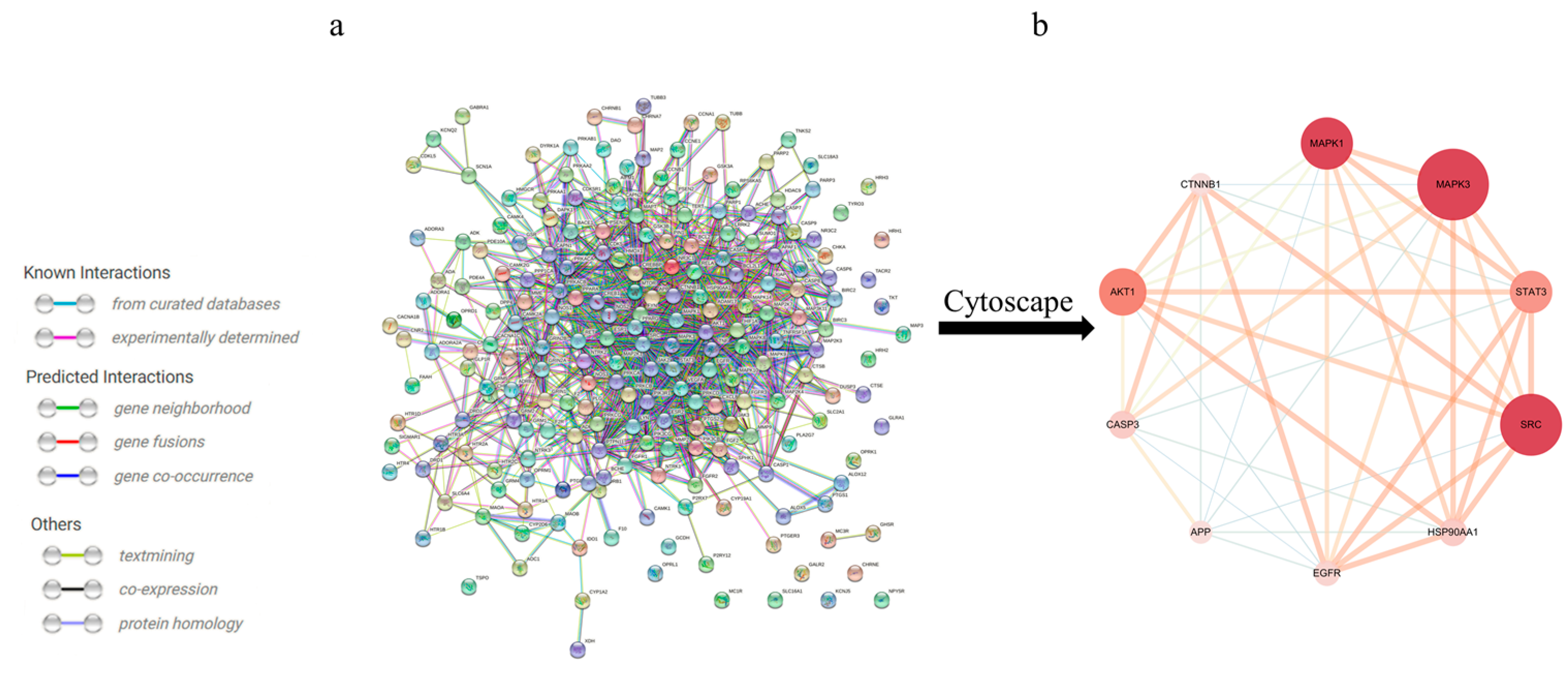
3.2. Construction of a PPI Network of Alkaloid-Excitotoxicity Intersection Targets
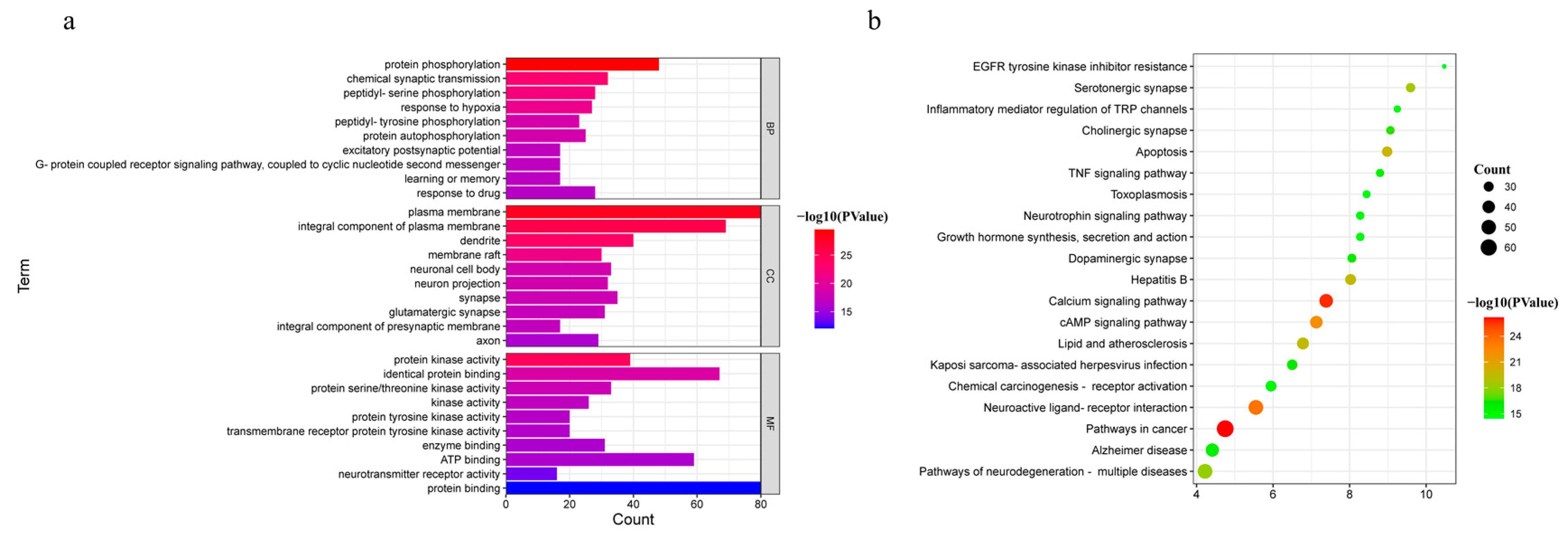
3.3. GO and KEGG Pathway Enrichment Analysis
3.4. Alkaloid–Target and Target–Pathway Network Analysis
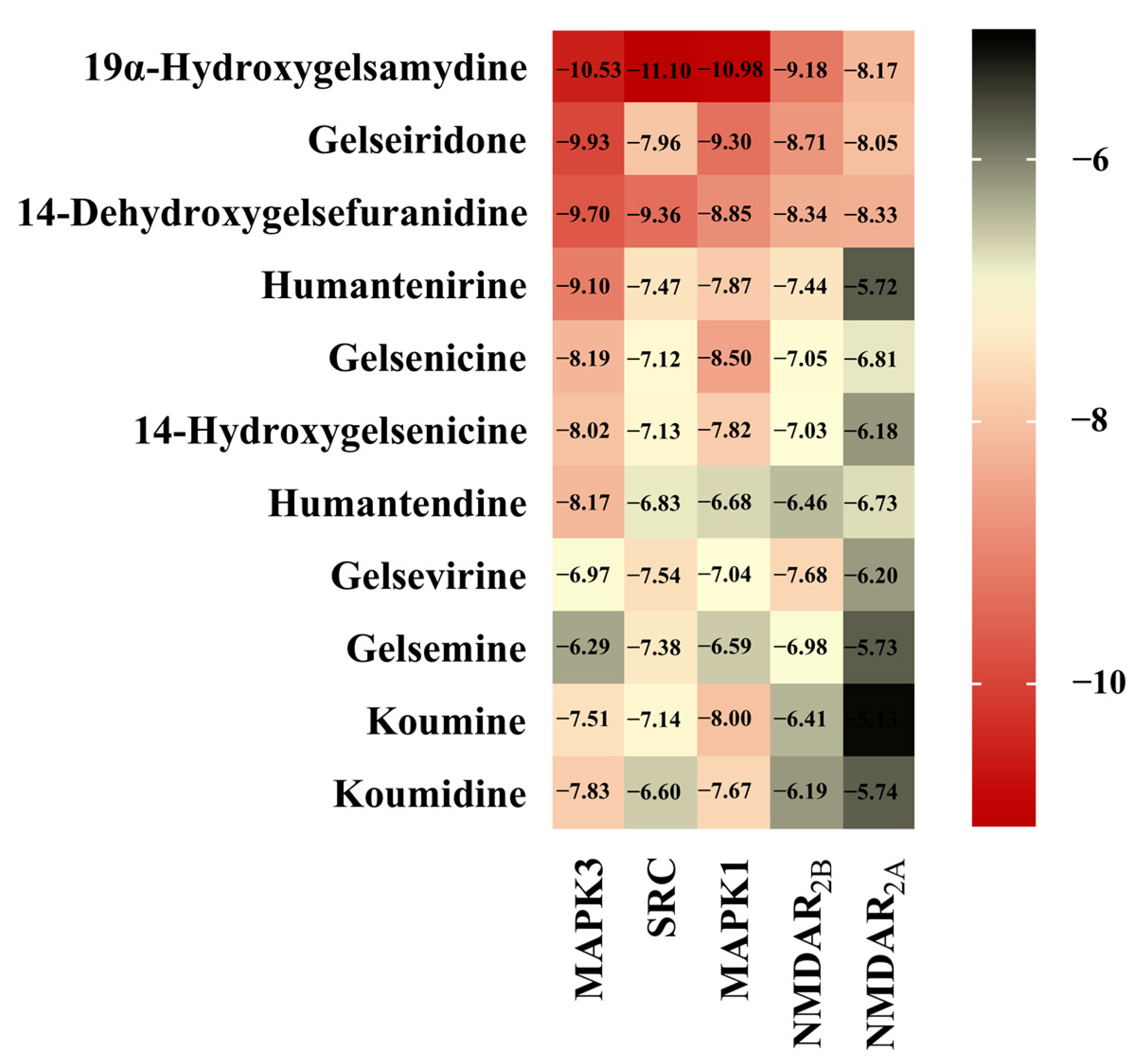
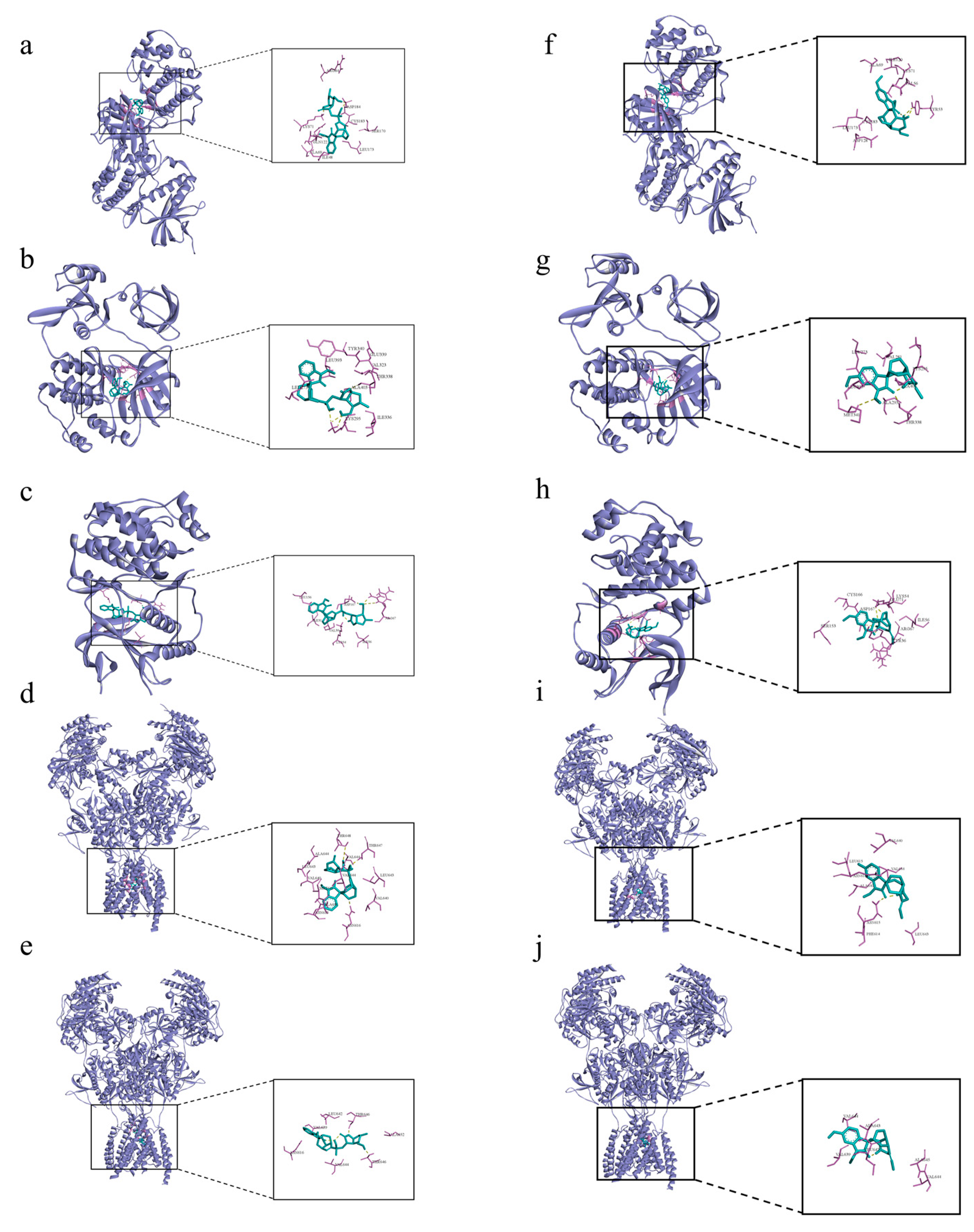
3.5. Molecular Docking
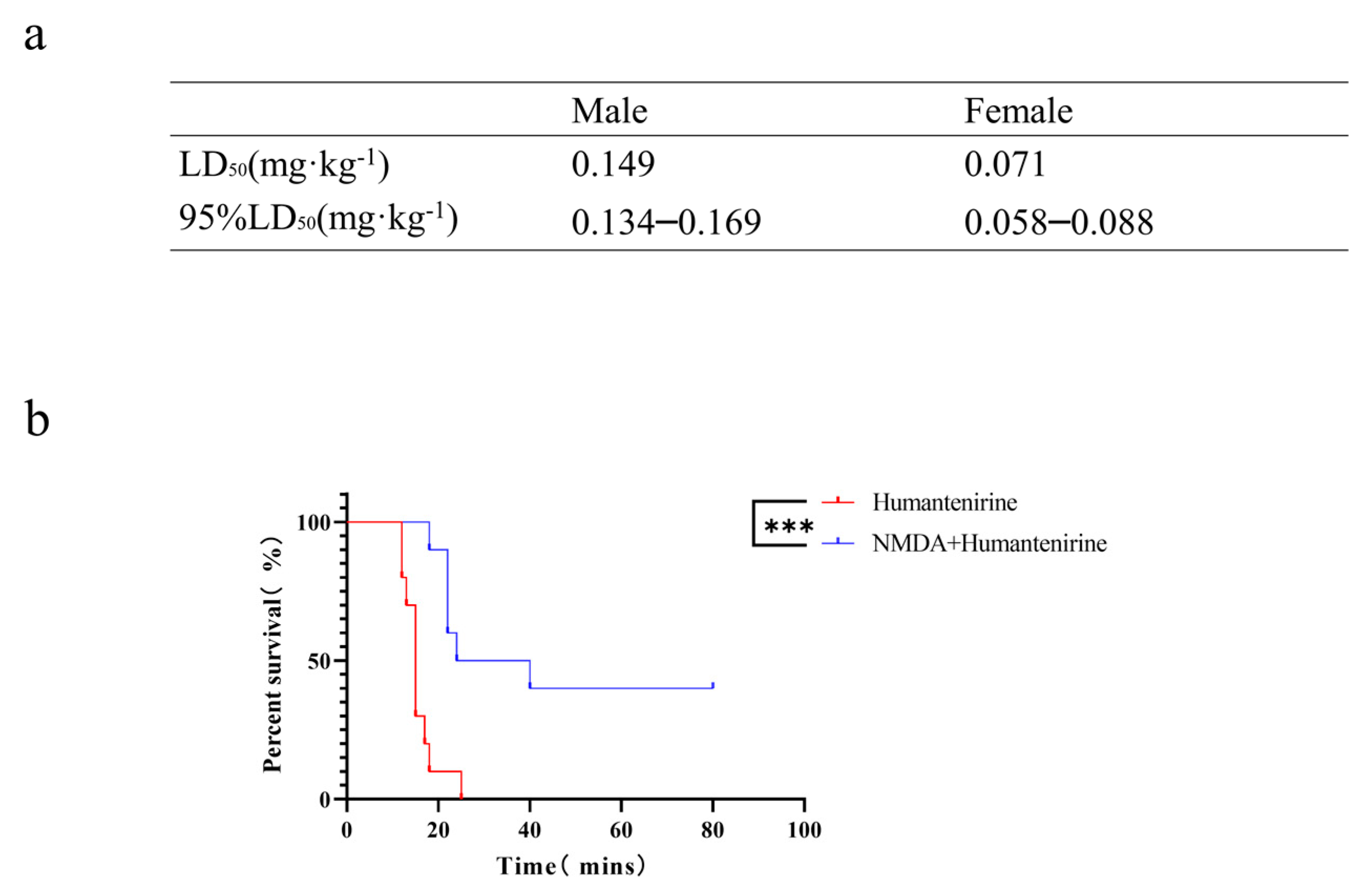
3.6. The LD50 of Acute Humantenirine Poisoning
3.7. The Antidotic Effect of NMDA on Humantenirine Poisoning
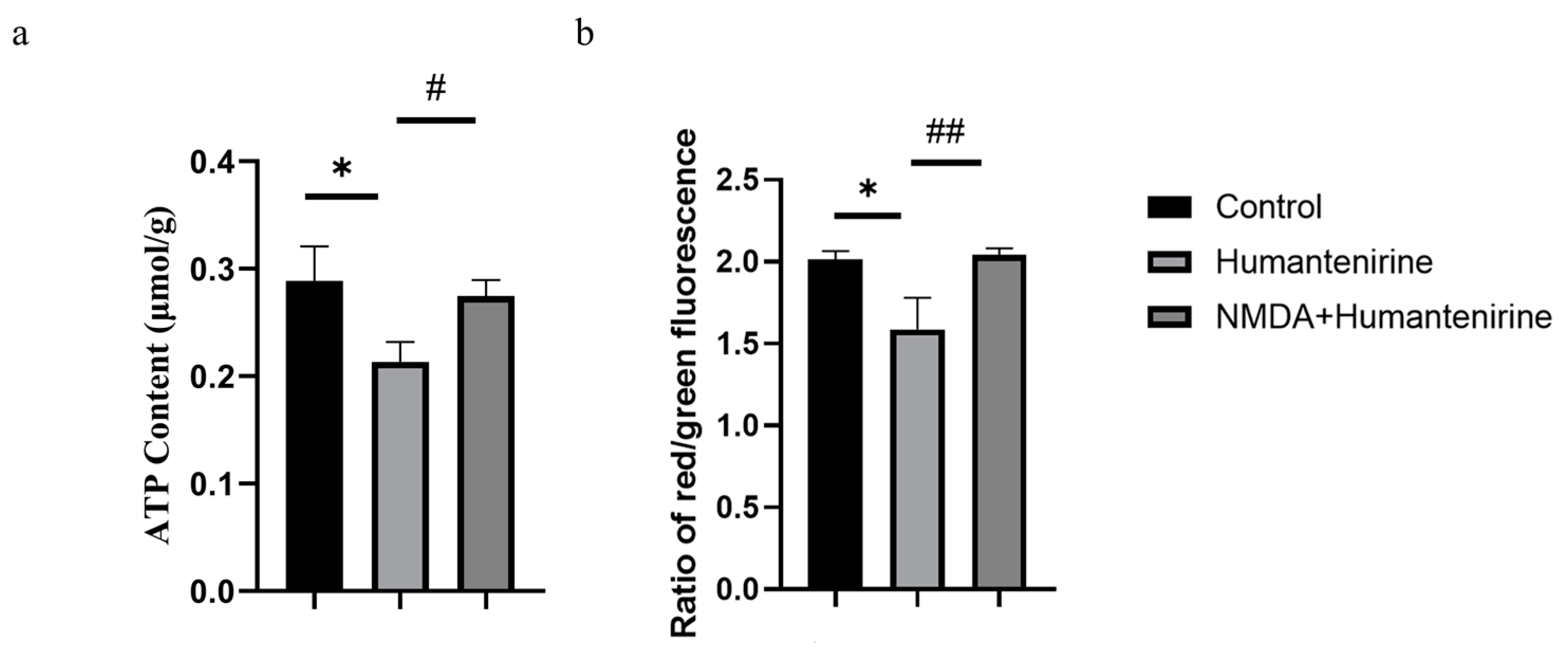
3.8. Effect of Humantenirine on ATP
3.9. Changes in Mitochondrial Membrane Potential
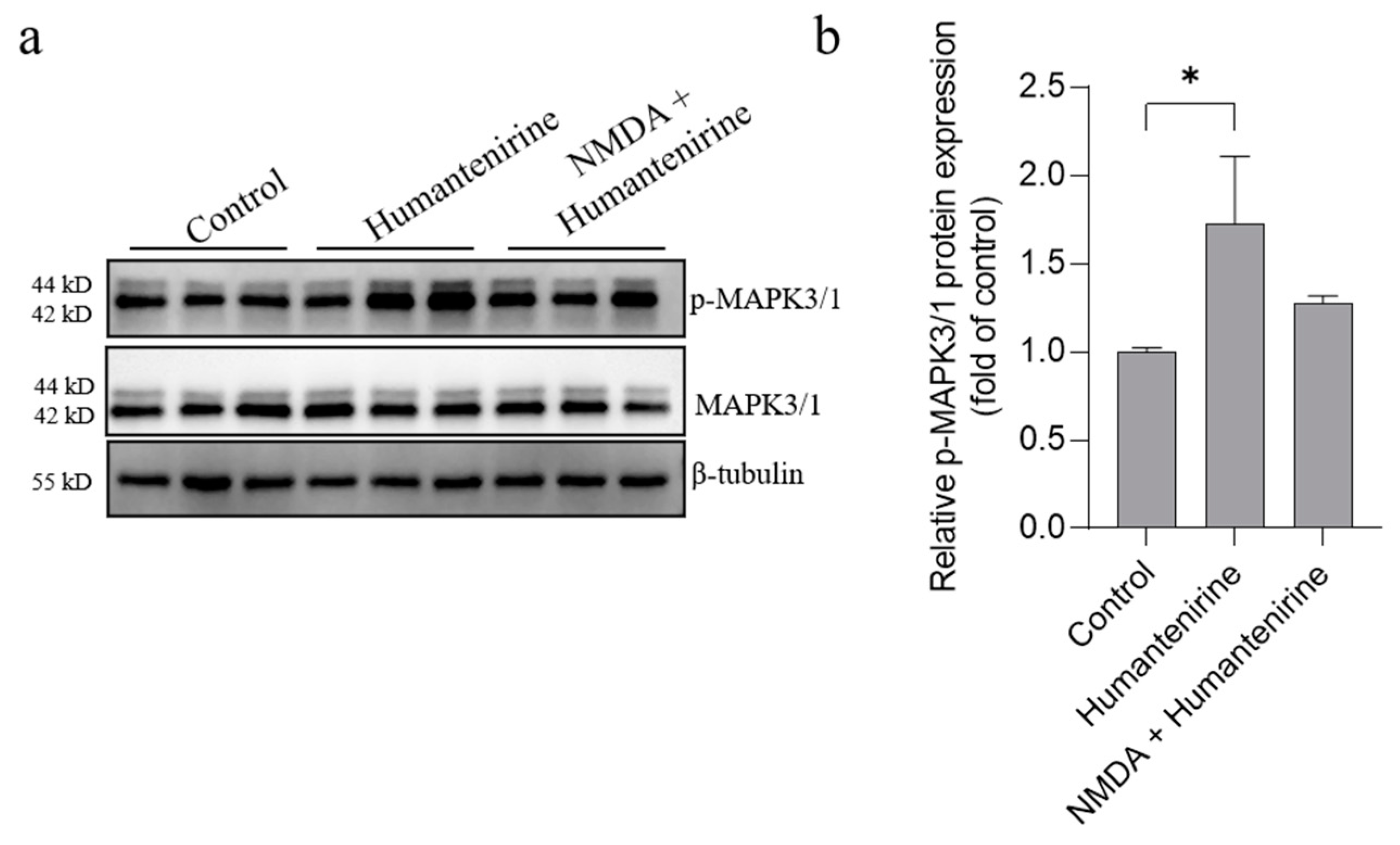
3.10. Humantenirine Induced Excitotoxicity by Upregulating Key Target Protein Expression in the Hippocampal Tissue of Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dutt, V.; Thakur, S.; Dhar, V.; Sharma, A. The genus Gelsemium: An update. Pharmacogn. Rev. 2010, 4, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.X.; Xu, Q.Q.; Meng, W.Q.; Cen, J.F.; Mao, G.C.; Pei, Z.P.; Xiao, K. Research progress on pharmacological and toxicological mechanism of Gelsemium elegans. J. Toxicol. 2020, 34, 336–341. [Google Scholar] [CrossRef]
- Zhao, Y.T.; Wu, S.P.; Hu, C.L.; Kang, J.; Zhao, M. Reviews on chemical compositions and pharmacological effect of Gelsemium elegans. Chin. J. Exp. Tradit. Med. Form. 2019, 25, 200–210. [Google Scholar]
- Bellavite, P.; Magnani, P.; Zanolin, E.; Conforti, A. Homeopathic Doses of Gelsemium sempervirens Improve the Behavior of Mice in Response to Novel Environments. Evidence-Based Complement. Altern. Med. 2011, 2011, 362517. [Google Scholar] [CrossRef]
- Jin, G.-L.; Su, Y.-P.; Liu, M.; Xu, Y.; Yang, J.; Liao, K.-J.; Yu, C.-X. Medicinal plants of the genus Gelsemium (Gelsemiaceae, Gentianales)—A review of their phytochemistry, pharmacology, toxicology and traditional use. J. Ethnopharmacol. 2014, 152, 33–52. [Google Scholar] [CrossRef] [PubMed]
- Du, X.B.; Dai, Y.H.; Zhang, C.L.; Lu, S.L.; Liu, Z.G. Studies on the Gelsemium Alkaloids—Ⅰ. The structure of Gelsenicine. Acta Chim. Sin. 1982, 40, 1137–1141. [Google Scholar]
- Zhou, Y.P.; Xu, W.; Chen, X.Y. Toxicity and respiratory inhibition of humantemine. Chin. J. Pharmacol. Toxicol. 1995, 9, 69–72. [Google Scholar]
- Chen, C.J.; He, J.L.; Wei, J.Y.; Chen, Y.X.; Qin, N.F.; He, J.; Yang, H.M. Literature analysis of 1034 cases of poisoning events induced by Gelsemium. J. Wuzhou Univ. 2020, 30, 11–19. [Google Scholar]
- Zhong, Y.X.; Xie, Y.H.; Jiang, Y.Y.; Liu, Y.P.; Shi, M.M.; Yang, W.M. Analysis of Gelsemium elegans alkalosis poisoning events in Guangxi Zhuang Autonomous Region from 2015 to 2017. Chin. J. Food Hyg. 2019, 31, 81–83. [Google Scholar] [CrossRef]
- Yang, S.; Liu, Y.; Sun, F.; Zhang, J.; Jin, Y.; Li, Y.; Zhou, J.; Li, Y.; Zhu, K. Gelsedine-type alkaloids: Discovery of natural neurotoxins presented in toxic honey. J. Hazard. Mater. 2019, 381, 120999. [Google Scholar] [CrossRef] [PubMed]
- Rujjanawate, C.; Kanjanapothi, D.; Panthong, A. Pharmacological effect and toxicity of alkaloids from Gelsemium elegans Benth. J. Ethnopharmacol. 2003, 89, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Lara, C.; Murath, P.; Muñoz, B.; Marileo, A.M.; Martín, L.S.; Martín, V.P.S.; Burgos, C.F.; A Mariqueo, T.; Aguayo, L.G.; Fuentealba, J.; et al. Functional modulation of glycine receptors by the alkaloid gelsemine. Br. J. Pharmacol. 2016, 173, 2263–2277. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-J.; Zuo, M.-T.; Qi, X.-J.; Huang, C.-Y.; Liu, Z.-Y. Phosphoproteomics reveals NMDA receptor-mediated excitotoxicity as a key signaling pathway in the toxicity of gelsenicine. Food Chem. Toxicol. 2021, 156, 112507. [Google Scholar] [CrossRef]
- Hopkins, A.L. Network pharmacology. Nat. Biotechnol. 2007, 25, 1110–1111. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.-G.; Wu, H.-B.; Chen, J.-S.; Li, X.; Qiu, Z.-K. Exploring the Potential Antidepressant Mechanisms of Pinellia by Using the Network Pharmacology and Molecular Docking. Metab. Brain Dis. 2022, 37, 1071–1094. [Google Scholar] [CrossRef]
- Khanal, P.; Zargari, F.; Far, B.F.; Kumar, D.; R, M.; Mahdi, Y.K.; Jubair, N.K.; Saraf, S.K.; Bansal, P.; Singh, R.; et al. Integration of System Biology Tools to Investigate Huperzine A as an Anti-Alzheimer Agent. Front. Pharmacol. 2021, 12. [Google Scholar] [CrossRef]
- Hu, J.; Zhao, L.; Li, N.; Yang, Y.; Qu, T.; Ren, H.; Cui, X.; Tao, H.; Chen, Z.; Peng, Y. Investigation of the active ingredients and pharmacological mechanisms of Porana sinensis Hemsl. Against rheumatoid arthritis using network pharmacology and experimental validation. PLoS ONE 2022, 17, e0264786. [Google Scholar] [CrossRef]
- Xue, R.; Fang, Z.; Zhang, M.; Yi, Z.; Wen, C.; Shi, T. TCMID: Traditional Chinese medicine integrative database for herb molecular mechanism analysis. Nucleic Acids Res. 2012, 41, D1089–D1095. [Google Scholar] [CrossRef]
- Chen, C.Y.-C. TCM Database@Taiwan: The world’s largest traditional Chinese medicine database for drug screening in silico. PLoS ONE 2011, 6, e15939. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef]
- Xia, C.Y.; Xu, J.K.; Li, L.; Lian, W.W.; Yan, Y.; Ma, B.Z.; He, J.; Zhang, W.K. Identifying the mechanism underlying antidepressant-like effects of loganin by network pharmacology in combination with experimental validation. J. Ethnopharmacol. 2021, 281, 114526. [Google Scholar] [CrossRef]
- Oliveros, J.C. Venny. An Interactive Tool for Comparing Lists with Venn’s Diagrams. Available online: https://bioinfogp.cnb.csic.es/tools/venny/index.html.2007–2015 (accessed on 2 August 2022). [CrossRef]
- Xu, Y.; Chen, J.; Yang, Z.; Xu, L. Identification of RNA Expression Profiles in Thyroid Cancer to Construct a Competing Endogenous RNA (ceRNA) Network of mRNAs, Long Noncoding RNAs (lncRNAs), and microRNAs (miRNAs). Experiment 2019, 25, 1140–1154. [Google Scholar] [CrossRef]
- Lu, J.; Yan, J.; Yan, J.; Zhang, L.; Chen, M.; Chen, Q.; Cheng, L.; Li, P. Network pharmacology based research into the effect and mechanism of Xijiao Dihuang decoction against sepsis. Biomed. Pharmacother. 2019, 122, 109777. [Google Scholar] [CrossRef]
- Xu, G.; Lv, X.; Feng, Y.; Li, H.; Chen, C.; Lin, H.; Li, H.; Wang, C.; Chen, J.; Sun, J. Study on the effect of active components of Schisandra chinensis on liver injury and its mechanisms in mice based on network pharmacology. Eur. J. Pharmacol. 2021, 910, 174442. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Lin, T.-Y.; Chen, I.-Y.; Lee, M.-Y.; Lu, C.-W.; Chiu, K.-M.; Wang, S.-J. Inhibition of Glutamate Release from Rat Cortical Nerve Terminals by Dehydrocorydaline, an Alkaloid from Corydalis yanhusuo. Molecules 2022, 27, 960. [Google Scholar] [CrossRef]
- Lu, C.-W.; Lin, T.-Y.; Chiu, K.-M.; Lee, M.-Y.; Huang, J.-H.; Wang, S.-J. Silymarin Inhibits Glutamate Release and Prevents against Kainic Acid-Induced Excitotoxic Injury in Rats. Biomedicines 2020, 8, 486. [Google Scholar] [CrossRef]
- Lin, T.Y.; Lu, C.W.; Huang, S.K.; Wang, S.J. Curcumin Inhibits Glutamate Release from Rat Prefrontal Nerve Endings by Affecting Vesicle Mobilization. Int. J. Mol. Sci. 2012, 13, 9097–9109. [Google Scholar] [CrossRef]
- Nateri, A.S.; Raivich, G.; Gebhardt, C.; Da Costa, C.; Naumann, H.; Vreugdenhil, M.; Makwana, M.; Brandner, S.; Adams, R.H.; Jefferys, J.G.R.; et al. ERK activation causes epilepsy by stimulating NMDA receptor activity. EMBO J. 2007, 26, 4891–4901. [Google Scholar] [CrossRef]
- Takagi, N.; Shinno, K.; Teves, L.; Bissoon, N.; Wallace, M.C.; Gurd, J.W. Transient ischemia differentially increases tyrosine phosphorylation of NMDA receptor subunits 2A and 2B. J. Neurochem. 2002, 69, 1060–1065. [Google Scholar] [CrossRef] [PubMed]
- Cheung, H.H.; Takagi, N.; Teves, L.; Logan, R.; Wallace, M.C.; Gurd, J.W. Altered Association of Protein Tyrosine Kinases with Postsynaptic Densities after Transient Cerebral Ischemia in the Rat Brain. J. Cereb. Blood Flow Metab. 2000, 20, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Gan, C.-L.; Zou, Y.; Chen, D.; Shui, X.; Hu, L.; Li, R.; Zhang, T.; Wang, J.; Mei, Y.; Wang, L.; et al. Blocking ERK-DAPK1 Axis Attenuates Glutamate Excitotoxicity in Epilepsy. Int. J. Mol. Sci. 2022, 23, 6370. [Google Scholar] [CrossRef]
- Wang, Y.T.; Salter, M.W. Regulation of NMDA receptors by tyrosine kinases and phosphatases. Nature 1994, 369, 233–235. [Google Scholar] [CrossRef]
- Cataldi, M.; Taglialatela, M.; Guerriero, S.; Amoroso, S.; Lombardi, G.; di Renzo, G.; Annunziato, L. Protein-tyrosine Kinases Activate while Protein-tyrosine Phosphatases Inhibit L-type Calcium Channel Activity in Pituitary GH3 Cells. J. Biol. Chem. 1996, 271, 9441–9446. [Google Scholar] [CrossRef]
- Moss, S.J.; Gorrie, G.H.; Amato, A.; Smart, T.G. Modulation of GABAA receptors by tyrosine phosphorylation. Nature 1995, 377, 344–348. [Google Scholar] [CrossRef] [PubMed]
- Wan, Q.; Man, H.Y.; Braunton, J.; Wang, W.; Salter, M.W.; Becker, L.; Wang, Y.T. Modulation of GABAAReceptor Function by Tyrosine Phosphorylation of β Subunits. J. Neurosci. 1997, 17, 5062–5069. [Google Scholar] [CrossRef]
- Li, Y.-J.; Yang, K.; Long, X.-M.; Xiao, G.; Huang, S.-J.; Zeng, Z.-Y.; Liu, Z.-Y.; Sun, Z.-L. Toxicity assessment of gelsenicine and the search for effective antidotes. Hum. Exp. Toxicol. 2022, 41. [Google Scholar] [CrossRef]
- Hashimoto, R.; Fujimaki, K.; Jeong, M.R.; Christ, L.; Chuang, D.-M. Lithium-induced inhibition of Src tyrosine kinase in rat cerebral cortical neurons: A role in neuroprotection against N -methyl-D -aspartate receptor-mediated excitotoxicity. FEBS Lett. 2003, 538, 145–148. [Google Scholar] [CrossRef]
- Connolly, N.M.C.; Prehn, J.H.M. The metabolic response to excitotoxicity—Lessons from single-cell imaging. J. Bioenerg. Biomembr. 2015, 47, 75–88. [Google Scholar] [CrossRef]
- Armada-Moreira, A.; Gomes, J.I.; Pina, C.C.; Savchak, O.K.; Gonçalves-Ribeiro, J.; Rei, N.; Pinto, S.; Morais, T.P.; Martins, R.S.; Ribeiro, F.F.; et al. Going the Extra (Synaptic) Mile: Excitotoxicity as the Road Toward Neurodegenerative Diseases. Front. Cell. Neurosci. 2020, 14, 90. [Google Scholar] [CrossRef] [PubMed]
- McBain, C.J.; Mayer, M.L. N-methyl-D-aspartic acid receptor structure and function. Physiol. Rev. 1994, 74, 723–760. [Google Scholar] [CrossRef] [PubMed]
- Berliocchi, L.; Bano, D.; Nicotera, P. Ca2+ signals and death programmes in neurons. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 2255–2258. [Google Scholar] [CrossRef]
- Choi, D.; Koh, J.; Peters, S. Pharmacology of glutamate neurotoxicity in cortical cell culture: Attenuation by NMDA antagonists. J. Neurosci. 1988, 8, 185–196. [Google Scholar] [CrossRef]
- Bunda, A.; LaCarubba, B.; Bertolino, M.; Akiki, M.; Bath, K.; Lopez-Soto, J.; Lipscombe, D.; Andrade, A. Cacna1b alternative splicing impacts excitatory neurotransmission and is linked to behavioral responses to aversive stimuli. Mol. Brain 2019, 12, 81. [Google Scholar] [CrossRef]
- Zuo, M.-T.; Wu, Y.; Wang, Z.-Y.; Wang, N.; Huang, S.-J.; Yu, H.; Zhao, X.-J.; Huang, C.-Y.; Liu, Z.-Y. A comprehensive toxicity evaluation in rats after long-term oral Gelsemium elegans exposure. Biomed. Pharmacother. 2021, 137, 111284. [Google Scholar] [CrossRef]
- Fan, J.; Gladding, C.M.; Wang, L.; Zhang, L.Y.; Kaufman, A.M.; Milnerwood, A.J.; Raymond, L.A. P38 MAPK is involved in enhanced NMDA receptor-dependent excitotoxicity in YAC transgenic mouse model of Huntington disease. Neurobiol. Dis. 2012, 45, 999–1009. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Yang, K.; Cao, J.-J.; Wang, Z.-Y.; Wu, Y.; Sun, Z.-L.; Liu, Z.-Y. Integration of Metabolomics and Transcriptomicsto Comprehensively Evaluate the Metabolic Effects of Gelsemium elegans on Pigs. Animals 2021, 11, 1192. [Google Scholar] [CrossRef]
- Ren, L.; Li, G.P.; Chen, S.N.; Xu, W.; Wang, H.S.; Huang, M.X.; Liao, H.J.; Wu, S.S. Material Basis Analysis of Toxicity Attenuation and Efficacy Reservation of Processing of Gelsemium elegans Rhizomes. Chin. J. Exp. Tradit. Med. Form. 2020, 26, 117–124. [Google Scholar]
- Zhou, X.; Ding, Q.; Chen, Z.; Yun, H.; Wang, H. Involvement of the GluN2A and GluN2B subunits in synaptic and extrasynaptic N-methyl-d-aspartate receptor function and neuronal excitotoxicity. J. Biol. Chem. 2013, 288, 24151–24159. [Google Scholar] [CrossRef]
- Vandresen-Filho, S.; Severino, P.C.; Constantino, L.C.; Martins, W.C.; Molz, S.; Dal-Cim, T.; Bertoldo, D.B.; Silva, F.R.M.B.; Tasca, C.I. N-Methyl-d-aspartate preconditioning prevents quinolinic acid-induced deregulation of glutamate and calcium homeostasis in mice hippocampus. Neurotox. Res. 2014, 27, 118–128. [Google Scholar] [CrossRef]
- Boeck, C.; Kroth, E.H.; Bronzatto, M.J.; Vendite, D. Adenosine receptors co-operate with NMDA preconditioning to protect cerebellar granule cells against glutamate neurotoxicity. Neuropharmacology 2005, 49, 17–24. [Google Scholar] [CrossRef]
- Kushnareva, Y.E.; Wiley, S.; Ward, M.; Andreyev, A.Y.; Murphy, A.N. Excitotoxic injury to mitochondria isolated from cultured neurons. J. Biol. Chem. 2005, 280, 28894–28902. [Google Scholar] [CrossRef]
- Gao, L.; Liu, F.; Hou, P.P.; Manaenko, A.; Xiao, Z.P.; Wang, F.; Xu, T.L.; Hu, Q. Neurons Release Injured Mitochondria as “Help-Me” Signaling After Ischemic Stroke. Front. Aging Neurosci. 2022, 14. [Google Scholar] [CrossRef]
- Yang, L.-K.; Lu, L.; Yue, J.; Wang, X.-S.; Qi, J.-Y.; Yang, F.; Liu, S.-B. Activation of microglial G-protein-coupled receptor 30 protects neurons against excitotoxicity through NF-κB/MAPK pathways. Brain Res. Bull. 2021, 172, 22–30. [Google Scholar] [CrossRef]
- Satoh, T.; Nakatsuka, D.; Watanabe, Y.; Nagata, I.; Kikuchi, H.; Namura, S. Neuroprotection by MAPK/ERK kinase inhibition with U0126 against oxidative stress in a mouse neuronal cell line and rat primary cultured cortical neurons. Neurosci. Lett. 2000, 288, 163–166. [Google Scholar] [CrossRef]
- Stanciu, M.; Wang, Y.; Kentor, R.; Burke, N.; Watkins, S.; Kress, G.; Reynolds, I.; Klann, E.; Angiolieri, M.R.; Johnson, J.W.; et al. Persistent activation of erk contributes to glutamate-induced oxidative toxicity in a neuronal cell line and primary cortical neuron cultures. J. Biol. Chem. 2000, 275, 12200–12206. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO. | Compound | CID | MW | MF |
|---|---|---|---|---|
| 1 | 11-Hydroxyhumantenine | 5318224 | 370.4 | C21H26N2O4 |
| 2 | 11-Hydroxyrankinidine | 5318332 | 356.4 | C20H24N2O4 |
| 3 | 11-Methoxy-19-(R)-Hydroxygelselegine | 5319453 | 404.5 | C21H28N2O6 |
| 4 | 11-Methoxygelsemamide | 5319437 | 355.4 | C21H25NO4 |
| 5 | 11-Methoxyhumantenine | 44583832 | 384.5 | C22H28N2O4 |
| 6 | 14β-Hydroxygelsedine | 126023 | 344.4 | C19H24N2O4 |
| 7 | 15-Hydroxyhumantenine | 101606434 | 370.4 | C21H26N2O4 |
| 8 | 16-Epi-Voacarpine | 5317127 | 368.4 | C21H24N2O4 |
| 9 | 19-(R)-Hydroxydihydrogelsemine | 5318191 | 340.4 | C20H24N2O3 |
| 10 | 19-(R)-Hydroxydihydrogelsevirine | 5318192 | 370.4 | C21H26N2O4 |
| 11 | 19-(R)-Hydroxydihydrokoumine | 5318193 | 324.4 | C20H24N2O2 |
| 12 | 19-(S)-Hydroxydihydrogelsevirine | 5318192 | 370.4 | C21H26N2O4 |
| 13 | 19-(S)-Hydroxydihydrokoumine | 5318193 | 324.4 | C20H24N2O2 |
| 14 | 19-(Z)-Akuammidine | 44583830 | 352.4 | C21H24N2O3 |
| 15 | 19-(Z)-Taberpsychine | 5321582 | 310.4 | C20H26N2O |
| 16 | 19-Oxo-Gelsenicine | 5320330 | 340.4 | C19H20N2O4 |
| 17 | 20-Hydroxydihydrorankinidine | 101606432 | 358.4 | C20H26N2O4 |
| 18 | Akuammidine N-Oxide | 11268654 | 368.4 | C21H24N2O4 |
| 19 | Akuammidinen-Oxide | 102423744 | 368.4 | C21H24N2O4 |
| 20 | Dihydrokoumine | 5316727 | 308.4 | C20H24N2O |
| 21 | Elegansamine | 5317023 | 508.6 | C29H36N2O6 |
| 22 | Epiwilsonine | 5315317 | 343.4 | C20H25NO4 |
| 23 | Gelsamydine | 5317540 | 508.6 | C29H36N2O6 |
| 24 | Gelsedine | 21589070 | 328.4 | C19H24N2O3 |
| 25 | Gelsemamide | 5317542 | 340.4 | C20H24N2O3 |
| 26 | Gelsemicine | 5462428 | 358.4 | C20H26N2O4 |
| 27 | Gelsemine | 5390854 | 322.4 | C20H22N2O2 |
| 28 | 4-(S)-Gelsemine N-Oxide | 5317545 | 338.4 | C20H22N2O3 |
| 29 | 4-(R)-Gelsemine N-Oxide | 5317545 | 338.4 | C20H22N2O3 |
| 30 | Gelsemoxonine | 44583831 | 358.4 | C19H22N2O5 |
| 31 | Gelsevirine | 14217344 | 352.4 | C21H24N2O3 |
| 32 | Humantenidine | 44584549 | 342.4 | C19H22N2O4 |
| 33 | Humantenine | 44593672 | 354.4 | C21H26N2O3 |
| 34 | Humantenirine | 11132403 | 370.4 | C21H26N2O4 |
| 35 | Humantenmine | 158212 | 326.4 | C19H22N2O3 |
| 36 | Koumidine | 44584550 | 294.4 | C19H22N2O |
| 37 | Koumine N-Oxide | 5318847 | 322.4 | C20H22N2O2 |
| 38 | N-Desmethoxyhumantenine | 5316593 | 324.4 | C20H24N2O2 |
| 39 | N-Desmethoxyrankinidine | 5316594 | 310.4 | C19H22N2O2 |
| 40 | Oxoglaucine | 97662 | 351.4 | C20H17NO5 |
| 41 | Rankinidine | 6439112 | 340.4 | C20H24N2O3 |
| 42 | Sempervirine(ii) | 168919 | 272.3 | C19H16N2 |
| 43 | Tabersonine | 25201472 | 337.4 | C21H25N2O2+ |
| 44 | 14-Hydroxygelsemicine | 597741 | 374.4 | C20H26N2O5 |
| 45 | Gelsenicine | 21123652 | 326.4 | C19H22N2O3 |
| 46 | Gelegamine D | 101467880 | 356.4 | C20H24N2O4 |
| 47 | Gelegamine E | 101467881 | 370.4 | C20H22N2O5 |
| 48 | GS-1 | 12070887 | 386.4 | C20H22N2O6 |
| 49 | GS-2 | 12070888 | 372.4 | C20H24N2O5 |
| 50 | 11-Hydroxygelsenicine | 102004554 | 342.4 | C19H22N2O4 |
| 51 | 11,14-Dihydroxygelsenicine | 101727430 | 358.4 | C19H22N2O5 |
| 52 | 14-Hydroxygelsenicine | 14217347 | 342.4 | C19H22N2O4 |
| 53 | 14-Acetoxygelsenicine | 11962104 | 384.4 | C21H24N2O5 |
| 54 | 14,15-Dihydroxygelsenicine | 44583829 | 358.4 | C19H22N2O5 |
| 55 | Gelsedilam | 102254466 | 314.34 | C17H18N2O4 |
| 56 | Gelsecrotonidine | 101449927 | 396.4 | C22H24N2O5 |
| 57 | 14-Hydroxygelsecrotonidine | 101449929 | 412.4 | C22H24N2O6 |
| 58 | 11-Methoxygelsecrotonidine | 101449930 | 426.5 | C23H26N2O6 |
| 59 | 14α-Hydroxygelsamydine | 44559138 | 524.6 | C29H36N2O7 |
| 60 | 19α-Hydroxygelsamydine | 102003053 | 524.6 | C29H36N2O7 |
| 61 | Gelegamine C | 101467879 | 514.4 | C21H27IN2O5 |
| 62 | 14-Acetoxygelselegine | 101727431 | 430.5 | C23H30N2O6 |
| 63 | 14α-Hydroxyelegansamine | 44559137 | 524.6 | C29H36N2O7 |
| 64 | Gelseoxazolidinine | 102297300 | 428.5 | C23H28N2O6 |
| 65 | Gelseziridine | 101951238 | 342.4 | C19H22N2O4 |
| 66 | GS-3 | 101751032 | 388.4 | C20H24N2O6 |
| 67 | Gelselenidine | 101951237 | 368.4 | C21H24N2O4 |
| 68 | Gelsesyringalidine | 136704418 | 490.5 | C28H30N2O6 |
| 69 | Gelsevanillidine | 136811988 | 460.5 | C27H28N2O5 |
| 70 | Gelsefuranidine | 102254468 | 420.5 | C24H24N2O5 |
| 71 | 14-Dehydroxygelsefuranidine | 102417029 | 404.5 | C24H24N2O4 |
| 72 | Gelsemolenine A | 101951239 | 384.4 | C21H24N2O5 |
| 73 | Gelsemolenine B | 101951240 | 370.4 | C20H22N2O5 |
| 74 | Gelseiridone | 101397829 | 538.6 | C29H34N2O8 |
| 75 | 21-Oxogelsemine | 11078214 | 336.4 | C20H20N2O3 |
| 76 | 21-Oxogelsevirine | 184299 | 366.4 | C21H22N2O4 |
| 77 | Gelsebanine | 16086585 | 504.6 | C30H36N2O5 |
| 78 | 6-Hydroxyhumantenine | 101855842 | 370.4 | C21H26N2O5 |
| 79 | 19(E)-Humantenine | 101520842 | 354.4 | C21H26N2O3 |
| 80 | Gelegamine A | 101467877 | 384.4 | C21H24N2O5 |
| 81 | Gelegamine B | 101467878 | 384.4 | C21H24N2O6 |
| 82 | Kounaminal | 102260292 | 363.5 | C22H25N3O2 |
| 83 | Gelsempervine A | 131636659 | 382.5 | C22H26N2O4 |
| 84 | Gelsempervine B | 101727385 | 424.5 | C24H28N2O5 |
| 85 | Gelsempervine C | 12444814 | 382.5 | C22H26N2O4 |
| 86 | Gelsempervine D | 101744809 | 424.5 | C24H28N2O5 |
| 87 | N-Methoxyanhydrovobasinediol | 102004539 | 338.4 | C21H26N2O2 |
| 88 | Dehydrokoumidine | 119077162 | 292.4 | C19H20N2O |
| 89 | Sempervilam | 11483103 | 288.3 | C19H16N2O |
| 90 | Ourouparine | 71436261 | 329.4 | C21H17N2O2+ |
| 91 | Gelsebamine | 16086588 | 255.35 | C14H25NO3 |
| 92 | Koumine | 91895267 | 306.4 | C20H22N2O |
| 93 | Humantendine | 5490912 | 342.4 | C19H22N2O4 |
| 94 | Gelsevirine N-Oxide | 101951241 | 368.4 | C21H24N2O4 |
| NO. | Name | BetweennessCentrality | ClosenessCentrality | Degree |
|---|---|---|---|---|
| 1 | MAPK3 | 0.0634 | 0.5272 | 60 |
| 2 | SRC | 0.0695 | 0.5105 | 56 |
| 3 | MAPK1 | 0.0381 | 0.5013 | 52 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, X.-J.; Huang, C.-Y.; Zuo, M.-T.; Gong, M.-D.; Huang, S.-J.; Tang, M.-H.; Liu, Z.-Y. Network Pharmacology and Experimental Verification to Unveil the Mechanism of N-Methyl-D-Aspartic Acid Rescue Humantenirine-Induced Excitotoxicity. Metabolites 2023, 13, 195. https://doi.org/10.3390/metabo13020195
Qi X-J, Huang C-Y, Zuo M-T, Gong M-D, Huang S-J, Tang M-H, Liu Z-Y. Network Pharmacology and Experimental Verification to Unveil the Mechanism of N-Methyl-D-Aspartic Acid Rescue Humantenirine-Induced Excitotoxicity. Metabolites. 2023; 13(2):195. https://doi.org/10.3390/metabo13020195
Chicago/Turabian StyleQi, Xue-Jia, Chong-Yin Huang, Meng-Ting Zuo, Meng-Die Gong, Si-Juan Huang, Mo-Huan Tang, and Zhao-Ying Liu. 2023. "Network Pharmacology and Experimental Verification to Unveil the Mechanism of N-Methyl-D-Aspartic Acid Rescue Humantenirine-Induced Excitotoxicity" Metabolites 13, no. 2: 195. https://doi.org/10.3390/metabo13020195
APA StyleQi, X.-J., Huang, C.-Y., Zuo, M.-T., Gong, M.-D., Huang, S.-J., Tang, M.-H., & Liu, Z.-Y. (2023). Network Pharmacology and Experimental Verification to Unveil the Mechanism of N-Methyl-D-Aspartic Acid Rescue Humantenirine-Induced Excitotoxicity. Metabolites, 13(2), 195. https://doi.org/10.3390/metabo13020195