Abstract
Myopathies have risen strongly in recent years, likely linked to selection for appetite. For white striping (WS), causes have been identified; but for wooden breast (WB), the cause remains speculative. We used metabolomics to study the breast muscle of 51 birds that were scored for both at 35 days of age to better understand potential causes. A partial least square discriminant analysis revealed that WS and WB had distinct metabolic profiles, implying different etiologies. Arginine and proline metabolism were affected in both, although differently: WB increased arginine in breast muscle implying that the birds did not use this pathway to increase tissue blood flow. Antioxidant defenses were impeded as shown by low anserine and beta-alanine. In contrast, GSH and selenium concentrations were increased. Serine, linked to anti-inflammatory properties, was increased. Taurine, which can stabilize the cell’s sarcolemma as well as modulate potassium channels and cellular calcium homeostasis, was also increased. Mineral data and depressed phosphatidylethanolamine, cAMP, and creatine-phosphate suggested compromised energy metabolism. WB also had drastically lower diet-derived lipids, suggesting compromised lipid digestion. In conclusion, WB may be caused by impaired lipid digestion triggered by a very high appetite: the ensuing deficiencies may well impair blood flow into muscle resulting in irreparable damage.
1. Introduction
Poultry meat is highly regarded as one of the most nutritious protein sources. The demand for poultry meat has risen in recent decades due to the low cost, good nutritional profile (high-protein content coupled with a balanced n-6 to n-3 polyunsaturated fatty acids ratio, low fat, low cholesterol, and presence of some functional components), suitability for further processing and no cultural or religious restrictions [1,2,3]. Additionally, together with milk, poultry products are recognized as one of the most environmentally efficient livestock products with regard to carbon footprint and resource depletion [4]. As for any product, the environmental impact is strongly influenced by the total amount and quality of the finished food product [5]. Thus, the growing demand for chicken meat and the push for lower production costs has led to the genetic selection of broilers for quantitative traits. It is worth noting that the selection procedures in favor of fast-growing broiler chicken strains have led to an extraordinary improvement in animal performance [6]. This significant increase in growth rate has been achieved by inducing muscle hypertrophy; and, unintentionally, genetic selection might have also induced meat quality issues such as white striping (WS) and wooden breast (WB) [7,8,9]. The alterations in the chemical composition and technological properties of the meat from birds affected by these myopathies may prevent the final product from reaching the consumer, and, therefore, increase wastage, thereby compromising sustainability [10,11,12]. The incidence of these myopathies has been estimated to lead to an economic loss of more than USD 200 million/year in the United States [13]. White striping is recognized by white striations that run parallel to the muscle fiber [14]. The underlying etiology of WS has been previously described by our group, and the proposed mechanism has been echoed by others [9,15,16].
Unfortunately, information around the underlying etiology of WB is less clear in the literature. WB is an abnormality in which the Pectoralis major muscle in broilers presents a pale and hardened consistency in cranial and/or caudal areas often accompanied by petechial hemorrhages and exudate [17,18]. Affected muscle have moderate to severe multifocal regenerative myodegeneration and necrosis with a variable amount of interstitial connective tissue accumulation or fibrosis. WB incidence has increased over the last years and, unfortunately, fillets with this abnormality are rejected from human consumption completely. The lack of automated systems at the slaughterhouse to detect WB complicates the quantification of this condition. However, a recent study using near infrared spectroscopy reported a prevalence of 9% among 10,483 fillets from high-yielding strains targeted to reach body weights of 2.72–4.53 kg [19]. WB fillets also show pathological changes including a reduction in protein and ash as well as an increase in fat and collagen [20]. It is often assumed that a link exists between the etiology of WS and WB, as both are frequently present together [13,21]. It is generally accepted that the prevalence of WB correlates positively with feed intakes and thus growth rates at a given age [22]. Studies have seen incidences in WB at 9 weeks of age of 85%, with more than 42% ranking severe or very severe; but already at 3 weeks of age, alterations in lipid-related genes are observed [23]. WB has been linked to rapid muscle growth inducing oxidative stress or hypoxia due to insufficient vascularization [3]. Since there are still many uncertainties about the underlying etiology, metabolomics was used to analyze both unaffected as well as affected breast muscle tissue. The overall objective of this study was to use non-targeted metabolomics to identify biological pathways that are involved in the underlying etiology of WB in broiler chickens.
2. Materials and Methods
2.1. Animals and Sample Collection
Sample collection and analysis has already been described earlier [15]. In brief, broilers used in this study were all ROSS 308 males, raised on a research farm (Poultry Research Centre; Casarrubios del Monte, Toledo, Spain) to 35 days. Males were chosen as they typically have a faster growth rate and higher incidence of myopathies. Diets fed were in line with local commercial practices (Table 1).

Table 1.
Ingredient and nutrient composition of the experimental diets as fed to the birds sampled for metabolomics. Data are in % except where noted.
On day 35, birds were euthanized in compliance with welfare rules. Time between euthanization and sample collection was as short as possible to ensure proper sample quality (on average 2–2.5 min). Muscle samples of the pectoralis major muscle were collected for 51 birds, which were randomly selected (two birds per pen). These muscle samples were scored for WS and WB. For WB, filets that were flexible throughout were scored as “normal” while filets in which hardness was present were scored as “moderate/severe” based on criteria described in the literature [24]; moderate/severe will be referred to as severe below. The samples were frozen using liquid nitrogen and stored at −80 °C until further processing.
An additional 47 birds were collected from a trial using diets with a similar nutrient composition. Those birds (mixed sex, Ross 308, 42 days of age) were scored for WS and WB as outlined above, and muscle samples were analyzed for vitamin K2 (MK-7) using LC-MS. Samples were grinded and 20 mL of acetonitrile was added. This was mixed for 60 min and then analyzed.
2.2. Sample Analysis
Metabolomic analysis of the 51 samples was performed by Metabolomic Discoveries (Potsdam-Golm, Germany), which is described in more detail by Boerboom (Boerboom et al., 2018). Muscle samples were also analyzed for phosphorus (P), sodium (Na), potassium (K), calcium (Ca), magnesium (Mg), copper (Cu), iron (Fe), zinc (Zn), cobalt (Co), nickel (Ni), and selenium (Se). Samples were weighed, destructed using 16 N HNO3, and then analyzed by inductively coupled plasma mass spectrometry using a NexION 350 D (Perkin-Elmer, Waltham, MA, USA) according to method NEN-EN 15510 [25].
2.3. Data Analysis
Metabolomics data were analyzed using MetaboAnalyst (Xia Lab, McGill, Ste. Anne de Bellevue, Quebec, Canada) [26,27,28,29,30]. Data were normalized on internal standards, log transformed and auto-scaled to remove heteroscedasticity and reduce bias. The p-value was adjusted using the false discovery rate (FDR) correction and was deemed significant if p < 0.05 [31]. Pathway analysis was done using MetaboAnalyst. The significance was calculated using pathway enrichment analysis which groups functionally related metabolites and tests whether they are significantly enriched. The impact was calculated by taking the pathway structure into account, thereby assigning more impact to changes in key positions in a pathway. As a measure for this, betweenness centrality was chosen, which takes into consideration the global network structure and not only the immediate neighbor of the metabolite like degree centrality does [30,32]. Additionally, a Partial Least Squares—Discriminant Analysis (PLS-DA) was performed using JMP 16.2 Pro (SAS Inst., Cary, NC, USA) using both WB and WS as discrete response parameters. Full cross validation and two rounds of very important variable selection (threshold > 1) were used. Data were also analyzed by multi-factor ANOVA using WS, WB, and their interaction WS × WB. Vitamin K2 data were analyzed using ANOVA including WB, WS, gender, and their 2-way interactions; this model was simplified prior to final analysis to only WB, as other factors were not significant (p > 0.2).
3. Results
Birds with severe WB averaged 2500 ± 62 g at sampling (mean ± SEM; day 35), birds scored as normal averaged 2380 ± 28 g (p = 0.08). The animals not selected were weighed on a pen basis and averaged 2289 ± 20 g; daily gain averaged 64.5 ± 0.58 g/d, feed intake averaged 94.5 ± 0.76 g/d, and FCR averaged 1.466 ± 0.005.
Out of a total of 51 samples (Table 2), 14 samples (27%) received a WB score of severe. Moderate WS was observed in 24 samples (47%), and severe WS was observed in eight birds (16%). Ten birds exhibited both WB and WS (of which one severe WS). The PLS-DA analysis (Figure 1) indicated that WB was placed effectively orthogonal to WS.
Table 2.
Bird count for the various myopathy scores.
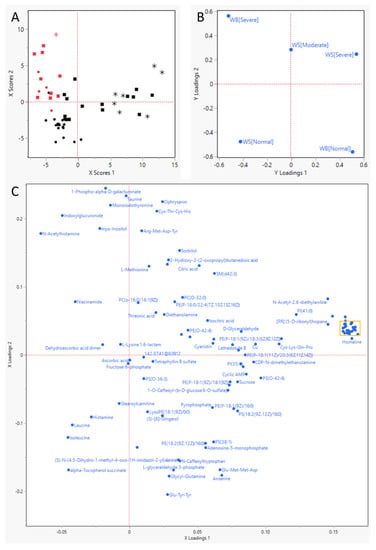

Figure 1.
Partial-Least Squares—Discriminant Analysis results. (A) score plot for the individual birds, represented by red (severe WB) or black (no WB) symbols; circles (• or •) indicate that they had no WS, squares (▪ or ▪) moderate WS, and stars (* or *) severe WS. (B) Y Loadings plot for the response parameters, severity of Wooden Breast (WB) and White Striping (WS). This plot uses the same layout as plots (A,C) and shows how the response variables (WB and WS) associate with the individual animals (plot (A)) and the various metabolites that survived the very important parameter selection (plot (C)). (C) Loadings plot for the metabolites remaining after two rounds of very important parameter selection). (D) close-up of the yellow rectangle in (C).
In the metabolomics data, bias was removed by the pre-processing steps described before [15], limiting the difference in absolute magnitude and difference in variance, thereby improving the quality of the dataset. The total number of annotated metabolites used in the analysis was 497. An ANOVA was used to determine the significant differences between the metabolites in normal birds and WB-affected birds. Afterwards, relative fold change was taken into account, and a volcano plot was created. The cut-off was set at an FDR-corrected p-value of 0.05 and a fold change threshold of 2. This yielded 65 metabolites (Supplementary Table S1). A positive fold change indicates that the severe WB group had increased concentrations of metabolites compared to the normal group. Several metabolites quantified were plant metabolites with poorly defined metabolic function. These will not be discussed in detail. It is worth noting that there is a high number of small peptides and of fat-soluble components that appear to be affected by the occurrence of WB.
Pathway analysis (Table 3) showed that histidine metabolism was most significantly affected. The highest impacted pathway was the taurine and hypo-taurine metabolism, which is somewhat confounded by the method applied, as these pathways are small and as such will more easily rank high. Individual pathway visualization was performed for the impacted pathways to ensure a more precise interpretation of the data. This highlighted the metabolism of arginine and proline, taurine, β-alanine, and glutathione. The pathways with a low impact were removed due to their lower biological effect.
Table 3.
Pathway analysis results using MetaboAnalyst. The p-value was calculated using pathway enrichment analysis and the impact was calculated using pathway topological analysis using betweenness centrality and using Gallus Gallus as a pathway library.
The results of the mineral analysis indicated significant changes occurring between normal and WB-affected breast tissues (Figure 2). The levels of sodium and selenium were elevated in WB-affected tissues (p < 0.05), while the levels of phosphorus, potassium, magnesium, copper, and nickel were lowered in WB-affected tissues (p < 0.05).

Figure 2.
Effect of WB on tissue mineral level. Ratios are in relation to the normal WB group (* p < 0.05).
The ANOVA analysis indicated that approximately 20 metabolites were decreased by over 95%, while 18 increased by more than 100% (Figure 3), most of them significantly so (such drastic downshifts were not seen in WS). In Figure 3, it is apparent that the compounds decreased are mainly lipophilic compounds with a uniquely dietary and/or microbial origin; several of these lipophilic compounds clustered strongly in Figure 1D. The most strongly increased metabolite was guanosine triphosphate adenosine and also taurine and the lysine catabolite 2-amino adipic acid stand out.

Figure 3.
Metabolites most strongly decreased (<5% vs. normal) and most strongly increased (>100% vs. normal) as a result of WB (* p < 0.05).
An in-depth look at the two metabolites with the strongest drop, vitamin K1 2.3-epoxide, and sorbitan palmitate (Figure 4), shows that both metabolites were effectively 0 in all birds with WB. In the normal birds, a wide range of concentrations was found.

Figure 4.
Two of the metabolites most severely depressed in birds with Wooden Breast (WB). Affected birds registered 0 for both of those metabolites, while normal birds displayed a wide spread in concentration. Circles (•) indicate that birds had no White Striping (WS), squares (▪) moderate WS, and stars (*) severe WS.
To further investigate a vitamin K2 deficiency as a possible factor in WB, we measured vit. K2 in birds collected in a separate trial (47 birds, of which 15 had WB; Figure 5). Neither sex nor WS affected vit. K2 levels. Birds without visible WB again showed a wide spread in vit. K2 levels, while in the affected birds the majority did not have detectable vit. K2 (Control: 10.6 µg/kg, WB: 0.8 µg/kg, p = 0.0003).

Figure 5.
Vitamin K2 (as MK-7) measurement in a separate group of birds scored for Wooden Breast (WB).
4. Discussion
Meat myopathies such as WS and WB are of huge concern to the broiler industry. Their occurrence is linked to the high appetite and consequently fast growth of the current genetic strains of birds used, and the resulting compromised blood supply and hypoxia are commonly brought forward as a starting point for their development [9,33]. The results of the PLS-DA analysis in the current study imply that WB and WS do not share a similar metabolic profile, but rather, express different changes in metabolite profiles, even if pathophysiological similarities might occur between the two diseases. Our results also imply that the metabolomics profile can be used to differentiate the two groups, in line with observations in the literature [34].
Arginine and proline metabolism were shown to be affected by WB occurrence, apparently like previous findings for WS [15]. However, the metabolites that changed within these pathways for WS were not the same metabolites as the ones changed in WB. Arginine to citrulline conversion was hypothesized to be increased in WS due to the observation of lower arginine levels and higher citrulline levels in WS cases [15]. In contrast, arginine levels showed an increase in WB-affected tissues (p < 0.05). This appears to indicate that there was still arginine available for conversion to citrulline for production of nitric oxide, required for enhanced blood flow [35,36]. Arginine in animals is used in two direct metabolic pathways: 1. Arginine is decomposed into ornithine and urea by arginase, and 2. Arginine and molecular oxygen generate citrulline and nitric oxide (NO) by nitric oxide synthase (NOS) [37]. However, no change in ornithine or citrulline was observed, indicating that none of the previously cited metabolic pathways of arginine utilization were being used.
Anserine (beta-alanyl-methyl-L-histidine) was reduced in WB-affected tissues (p < 0.05). Anserine is important for antioxidant functions, pH buffering and anti-glycation agents [38,39]. Anserine levels tend to be lower when birds get older resulting in reduced antioxidant capacity [40]. This reduction is in accordance with literature on WB, where a reduction in anserine has been observed more frequently [9,41]. A potential contributor to the lower levels of anserine that were observed in this study could be the lower levels of beta-alanine, which was also observed (p < 0.05). The rate of anserine synthesis in skeletal muscle is dependent on the amount of circulating beta-alanine. In line with these findings, a recently published article on metabolomics in WB-affected birds indicated that 3-methyl-histidine was the top metabolite being affected in plasma, with eight more metabolites involved in histidine metabolism being affected, further indicating the effect WB occurrence has on histidine metabolism [34]. Surprisingly, though, a study feeding high levels of histidine indicated no effects on meat quality, even though a higher level of carnosine was observed [42].
Phosphatidylethanolamine (PE) is the second most abundant glycerophospholipid in eukaryotic cells [43]. It has diverse cellular functions that include serving as a precursor for the formation of phosphatidylcholine, influencing membrane topology and promoting cell and organelle membrane fusion, oxidative phosphorylation, mitochondrial biogenesis, and autophagy. It is also an important precursor for other lipids [43,44,45]. The results of the metabolomics indicate that the levels of PE in WB-affected tissues are lowered. In case of limited availability, mitochondrial activity is negatively affected given the importance of PE in mitochondrial stability and activity [43]. This could have compromised cellular energy metabolism in WB-affected tissues, in line with lowered levels of cAMP and phospho-creatine. Furthermore, it has been observed that the level of PE is important for membrane stability, and as such, the lower levels of PE observed could have led to membrane destabilization, leading to WB [46]. This is further substantiated by the mineral results, which show a reduction in phosphorus, magnesium, and copper along with an increase in sodium. These changes fit with an impairment in metabolic activity of the cell as phosphate and magnesium are key in energy metabolism while copper is an important cofactor for various oxidases. Perturbed energy metabolism can subsequently lead to an efflux of potassium and an influx of calcium (ns) and sodium into the cells [47]. In order to compensate for this, the cells might have produced high amounts of glutathione (GSH) as this is a master regulator of metabolism within the cell [48]. Aside from its function as a regulator of metabolism, it is also a cysteine-derived antioxidant and important for detoxification, protection from oxidative stress and maintenance of redox balance. In line with this antioxidant effect, a significant increase in selenium was observed in WB birds. Synthesis of GSH is driven by the activity of the pentose phosphate pathway [49], which has been shown to be influenced by WB in previously published literature [34,41]. They indicated that WB affected purine metabolism, which was a result of the increased activity of the pentose phosphate pathway or a decreased nucleotide utilization. In this dataset, serine levels tended to be higher in WB-affected tissues (p = 0.05) and they play an important role in the metabolism of purines and pyrimidines, further substantiating the effect WB has on purine metabolism [50]. Numerous studies have indicated that glycine can exert anti-inflammatory and antioxidant effects, protecting against hypoxia induced injuries, albeit in renal tissue [51]. It could also be that serine expression is increased in WB-affected tissue to protect against the hypoxic state that is created by damaged vascular support system [17]. The conversion of serine into glycine leads to an increase in Ca influx, something that is also observed in the WB-affected tissues in this study (Figure 2).
As described for WS, taurine is often released at higher levels to protect against damage of hypoxic nature, and in addition, it plays a pivotal role in Ca homeostasis [2,15]. Levels of taurine, a metabolite of methionine, increased similarly for both WB and WS (p < 0.05). The key effect of taurine is stabilization of the sarcolemma. Besides this, taurine also modulates the activity of various types of K channels and, in particular, those able to couple metabolic state of striated fibers to electrical activity. Lastly, it controls intracellular calcium homeostasis by modulating calcium handling mechanisms and consequently excitation-contraction coupling [52]. Taurine uptake into cells also has an osmotic function, and high levels may reflect swollen cells, which is a common observation in WB-affected tissues [53]. Another affected metabolite in the cysteine and methionine metabolism is 3-methylthiopropionic acid, which is almost absent in the WB-affected tissue (p < 0.01). 3-methylthiopropionic acid is an intermediate in the transamination pathway of methionine metabolism. As methionine was not affected, a possible explanation for low levels of 3-methylthiopropionic acid is that the transamination pathway for methionine catabolism was downregulated and methionine instead was converted to taurine, in line with increased taurine levels [54].
The drastic reduction in various diet-derived lipids as observed in the ANOVA analysis implies that lipid digestion was impaired, which may well be linked to the very high feed intake of birds affected by WB. Indeed, Nir et al. (1973) and Krogdahl (1985) showed that digestion of fat is typically first limiting when intestinal passage rate is increased [55,56]. This observation also aligns with changes in lipid-related genes [23,57]. Liu et al. also showed strong changes in lipids, but their analysis focused on lipids that are not per se obtained from the diet [58]. Several of the affected lipids are linked to specific metabolic functions, like a vitamin, in either the bird or in the feedstuffs the birds consumed. These lipids may well be harder to digest than those used as energy sources, aggravated by very high feed intake. For example, for vitamin E a bioavailability of only 5% was obtained [59]. In line with this, Thompson (1989) highlighted numerous case reports describing patients developing deficiencies of vitamins D, E, and K as a consequence of lipid malabsorption [60]. As shown in Figure 4, birds with WB had effectively tissue levels of zero for the most severely affected lipids. In the ‘normal’ birds, interestingly, also several birds registered at 0, but also a substantial number of birds had higher levels. A possible explanation, in line with field observations, is that deficiencies of these diet-derived lipids and thus problems with WB start earlier in some birds than others; those that appeared normal at slaughter may well have developed WB if they had been slaughtered a week later due to aggravating deficiencies of these lipids. A vitamin K2 analysis of 47 samples of breast muscle tissue obtained from an independent flock of birds confirmed that birds with severe WB had practically zero vit. K2, while normal birds again had a wide spread in vit. K2 levels (Figure 5). Vit. K2 is involved in calcium metabolism; when animals are deficient, it leads to calcium deposits in blood vessels, in line with what Sihvo and Mutryn showed, and in line with data from Abasht et al. (2021) showing vascular endothelial dysfunction [17,57,61,62]. This may well impede blood flow into breast muscle tissue, resulting in atrophy, in line with the observations that creatine kinase (a marker for muscle injury) increased with WB severity [63]. Several of the observations in the metabolic pathways fit with this observation as does the appearance of petechia and hemorrhages.
Another interesting lead is the enormous increase in guanosine triphosphate adenosine (Figure 3; also seen in WS). This class of molecules is used to cap mRNA protecting it from degradation [64]. In addition, they are strong agonists of the purinergic system involved in vasoregulation [65]. High levels may well mean that the animal was trying to counter hypoxia by vasodilation and vascular regeneration, but unsuccessfully did so as blood vessels were damaged by calcium deposits linked to a vitamin K2 deficiency.
5. Conclusions
Taken together, the results of our analyses seem to indicate that birds suffering from WB suffer from a problem with the digestion of lipids such as vitamin K2. Although not evaluated in this experiment, these digestive problems could potentially be the result of feed intake that exceeds the digestive capacity of the birds. The resulting lipid deficiencies may well impair vascular health and antioxidant defenses, resulting in tissue hypoxia and irreparable damage. Tissue hypoxia is something that WS and WB have in common, but the magnitude/extend of the hypoxic state as well as the expression do differ. In the case of WS, the hypoxia appears more manageable, allowing the tissue to retain some functionality and growth, albeit with an altered nutrient profile. In the case of WB, the hypoxia appears more severe resulting in tissue that has severely compromised functionality, in line with other observations [66]. Problems with fat digestion as a result of excessive feed intake deserve further attention and, hence, should be investigated further.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/metabo13010020/s1, Table S1: Significantly affected metabolites based on volcano plot between the groups (FDR corrected p < 0.05, Log2FC threshold is 1).
Author Contributions
Conceptualization, G.M.B., A.N.-V. and T.A.T.G.v.K.; methodology, G.M.B., A.N.-V. and T.A.T.G.v.K.; data formal analysis, G.M.B. and T.A.T.G.v.K.; writing—original draft preparation, G.M.B. and T.A.T.G.v.K.; writing—review and editing, A.N.-V.; visualization, G.M.B. and T.A.T.G.v.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
All animal care procedures were carried out as described in the protocol approved by Centro de Estudios Biosanitarios (Madrid, Spain) by the External Ethical Committee, and approved by the Junta de Castilla-La Mancha Animal Welfare department as compliable with the Real Decreto RD 53/2013, Animal Care and Use Committee (98R064-0).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data are not publicly available due to privacy.
Acknowledgments
The Poultry Research Centre (Casarrubios del Monte, Toledo, Spain) staff for animal care and sampling and the staff of the research laboratory Nutreco, Boxmeer, for the mineral analysis.
Conflicts of Interest
All authors were employed by Nutreco at the time the research was carried out, the company that funded and hosted the experiment.
References
- Petracci, M.; Mudalal, S.; Bonfiglio, A.; Cavani, C. Occurrence of white striping under commercial conditions and its impact on breast meat quality in broiler chickens. Poult. Sci. 2013, 92, 1670–1675. [Google Scholar] [CrossRef] [PubMed]
- Petracci, M.; Mudalal, S.; Soglia, F.; Cavani, C. Meat quality in fast-growing broiler chickens. World’s Poult. Sci. J. 2015, 71, 363–374. [Google Scholar] [CrossRef]
- Petracci, M.; Soglia, F.; Madruga, M.; Carvalho, L.; Ida, E.; Estévez, M. Wooden-breast, white striping, and spaghetti meat: Causes, consequences and consumer perception of emerging broiler meat abnormalities. Compr. Rev. Food Sci. Food Saf. 2019, 18, 565–583. [Google Scholar] [CrossRef] [PubMed]
- Roma, R.; Corrado, S.; De Boni, A.; Forleo, M.B.; Fantin, V.; Moretti, M.; Palmieri, N.; Vitali, A.; Camillo, D.C. Life cycle assessment in the livestock and derived edible products sector. In Life Cycle Assessment in the Agri-Food Sector; Springer: Berlin/Heidelberg, Germany, 2015; pp. 251–332. [Google Scholar]
- Costantini, M.; Ferrante, V.; Guarino, M.; Bacenetti, J. Environmental sustainability assessment of poultry productions through life cycle approaches: A critical review. Trends Food Sci. Technol. 2021, 110, 201–212. [Google Scholar] [CrossRef]
- Tixier-Boichard, M. From the jungle fowl to highly performing chickens: Are we reaching limits? World’s Poult. Sci. J. 2020, 76, 2–17. [Google Scholar] [CrossRef]
- Alnahhas, N.; Berri, C.; Chabault, M.; Chartrin, P.; Boulay, M.; Bourin, M.C.; Bihan-Duval, L. Genetic parameters of white striping in relation to body weight, carcass composition, and meat quality traits in two broiler lines divergently selected for the ultimate pH of the pectoralis major muscle. BMC Genet. 2016, 17, 61. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.R.; Suyemoto, M.M.; Sarsour, A.H.; Cordova, H.A.; Oviedo-Rondón, E.O.; Wineland, M.; Barnes, H.J.; Borst, L.B. Temporal characterization of wooden breast myopathy (“woody breast”) severity and correlation with growth rate and lymphocytic phlebitis in three commercial broiler strains and a random-bred broiler strain. Avian Pathol. 2019, 48, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Soglia, F.; Petracci, M.; Davoli, R.; Zappaterra, M. A critical review of the mechanisms involved in the occurrence of growth-related abnormalities affecting broiler chicken breast muscles. Poult. Sci. 2021, 100, 101180. [Google Scholar] [CrossRef] [PubMed]
- Geronimo, B.C.; Prudencio, S.H.; Soares, A.L. Biochemical and technological characteristics of wooden breast chicken fillets and their consumer acceptance. J. Food Sci. Technol. 2022, 59, 1185–1192. [Google Scholar] [CrossRef] [PubMed]
- Maharjan, P.; Beitia, A.; Weil, J.; Suesuttajit, N.; Hilton, K.; Caldas, J.; Umberson, C.; Martinez, D.; Kong, B.; Owens, C.M. Woody breast myopathy broiler show age-dependent adaptive differential gene expression in Pectoralis major and altered in-vivo triglyceride kinetics in adipogenic tissues. Poult. Sci. 2021, 100, 101092. [Google Scholar] [CrossRef]
- Petracci, M.; Mudalal, S.; Babini, E.; Cavani, C. Effect of white striping on chemical composition and nutritional value of chicken breast meat. Ital. J. Anim. Sci. 2014, 13, 3138. [Google Scholar] [CrossRef]
- Kuttappan, V.; Hargis, B.; Owens, C. White striping and woody breast myopathies in the modern poultry industry: A review. Poult. Sci. 2016, 95, 2724–2733. [Google Scholar] [CrossRef] [PubMed]
- Kuttappan, V.A.; Lee, Y.S.; Erf, G.F.; Meullenet, J.F.; McKee, S.R.; Owens, C.M. Consumer acceptance of visual appearance of broiler breast meat with varying degrees of white striping. Poult. Sci. 2012, 91, 1240–1247. [Google Scholar] [CrossRef] [PubMed]
- Boerboom, G.; van Kempen, T.; Navarro-Villa, A.; Pérez-Bonilla, A. Unraveling the cause of white striping in broilers using metabolomics. Poult. Sci. 2018, 97, 3977–3986. [Google Scholar] [CrossRef]
- Muroya, S.; Ueda, S.; Komatsu, T.; Miyakawa, T.; Ertbjerg, P. MEATabolomics: Muscle and meat metabolomics in domestic animals. Metabolites 2020, 10, 188. [Google Scholar] [CrossRef]
- Sihvo, H.-K.; Lindén, J.; Airas, N.; Immonen, K.; Valaja, J.; Puolanne, E. Wooden breast myodegeneration of pectoralis major muscle over the growth period in broilers. Veter Pathol. 2017, 54, 119–128. [Google Scholar] [CrossRef]
- Sihvo, H.K.; Immonen, K.; Puolanne, E. Myodegeneration with fibrosis and regeneration in the pectoralis major muscle of broilers. Veter Pathol. 2013, 51, 619–623. [Google Scholar] [CrossRef]
- Wold, J.P.; Veiseth-Kent, E.; Høst, V.; Løvland, A. Rapid on-line detection and grading of wooden breast myopathy in chicken fillets by near-infrared spectroscopy. PLoS ONE 2017, 12, e0173384. [Google Scholar] [CrossRef]
- Soglia, F.; Laghi, L.; Canonico, L.; Cavani, C.; Petracci, M. Functional property issues in broiler breast meat related to emerging muscle abnormalities. Food Res. Int. 2016, 89, 1071–1076. [Google Scholar] [CrossRef]
- Livingston, M.L.; Landon, C.; Barnes, H.; Brake, J. White striping and wooden breast myopathies of broiler breast muscle is affected by time-limited feeding, genetic background, and egg storage. Poult. Sci. 2019, 98, 217–226. [Google Scholar] [CrossRef]
- Brothers, B.; Zhuo, Z.; Papah, M.B.; Abasht, B. RNA-seq analysis reveals spatial and sex differences in pectoralis major muscle of broiler chickens contributing to difference in susceptibility to wooden breast disease. Front. Physiol. 2019, 10, 764. [Google Scholar] [CrossRef] [PubMed]
- Papah, M.B.; Abasht, B. Dysregulation of lipid metabolism and appearance of slow myofiber-specific isoforms accompany the development of Wooden Breast myopathy in modern broiler chickens. Sci. Rep. 2019, 9, 17170. [Google Scholar] [CrossRef] [PubMed]
- Tijare, V.V.; Yang, F.L.; Kuttappan, V.A.; Alvarado, C.Z.; Coon, C.N.; Owens, C.M. Meat quality of broiler breast fillets with white striping and woody breast muscle myopathies. Poult. Sci. 2016, 95, 2167–2173. [Google Scholar] [CrossRef] [PubMed]
- Bikker, P.; ten Tije, N.; Tijkorte, A. Fosforbenutting Bij Biologisch Gehouden Vleesvarkens; Wageningen Livestock Research: Wageningen, The Netherlands, 2017. [Google Scholar]
- van den Berg, R.A.; Hoefsloot, H.C.; Westerhuis, J.A.; Smilde, A.K.; van der Werf, M.J. Centering, scaling, and transformations: Improving the biological information content of metabolomics data. BMC Genomic. 2006, 7, 142. [Google Scholar] [CrossRef]
- Xia, J.; Mandal, R.; Sinelnikov, I.V.; Broadhurst, D.; Wishart, D.S. MetaboAnalyst 2.0—A comprehensive server for metabolomic data analysis. Nucleic Acids Res. 2012, 40, W127–W133. [Google Scholar] [CrossRef]
- Xia, J.; Psychogios, N.; Young, N.; Wishart, D.S. MetaboAnalyst: A web server for metabolomic data analysis and interpretation. Nucleic Acids Res. 2009, 37, W652–W660. [Google Scholar] [CrossRef]
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0—Making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef]
- Xia, J.; Wishart, D.S. Metabolomic data processing, analysis, and interpretation using MetaboAnalyst. Curr. Protoc. Bioinform. 2011, 34, 14.10.1–14.10.48. [Google Scholar] [CrossRef]
- Vinaixa, M.; Samino, S.; Saez, I.; Duran, J.; Guinovart, J.J.; Yanes, O. A Guideline to Univariate Statistical Analysis for LC/MS-Based Untargeted Metabolomics-Derived Data. Metabolites 2012, 2, 775. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef]
- Zampiga, M.; Soglia, F.; Petracci, M.; Meluzzi, A.; Sirri, F. Effect of different arginine-to-lysine ratios in broiler chicken diets on the occurrence of breast myopathies and meat quality attributes. Poult. Sci. 2019, 98, 2691–2697. [Google Scholar] [CrossRef] [PubMed]
- Lake, J.A.; Yan, Y.; Dekkers, J.C.; Qiu, J.; Brannick, E.M.; Abasht, B. Identification of circulating metabolites associated with wooden breast and white striping. PLoS ONE 2022, 17, e0274208. [Google Scholar] [CrossRef] [PubMed]
- Bloomer, R.J.; Farney, T.M.; Trepanowski, J.F.; McCarthy, C.G.; Canale, R.E.; Schilling, B.K. Comparison of pre-workout nitric oxide stimulating dietary supplements on skeletal muscle oxygen saturation, blood nitrate/nitrite, lipid peroxidation, and upper body exercise performance in resistance trained men. J. Int. Soc. Sports Nutr. 2010, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Sun, W.-D.; Li, J.-C.; Pan, J.-Q.; Liu, Y.-J.; Wang, J.-Y.; Wang, X.-L. L-arginine prevents reduced expression of endothelial nitric oxide synthase (NOS) in pulmonary arterioles of broilers exposed to cool temperatures. Veter J. 2007, 173, 151–157. [Google Scholar] [CrossRef]
- Guo, Y.; Shi, B.; Yan, S.; Xu, Y.; Li, J.; Li, T. Effects of arginine on cytokines and nitric oxide synthesis in broilers. J. Anim. Plant Sci. 2015, 25, 366–371. [Google Scholar]
- Haug, A.; R⊘dbotten, R.; Mydland, L.T.; Christophersen, O.A. Increased broiler muscle carnosine and anserine following histidine supplementation of commercial broiler feed concentrate. Acta Agric. 2008, 58, 71–77. [Google Scholar] [CrossRef]
- Kai, S.; Watanabe, G.; Kubota, M.; Kadowaki, M.; Fujimura, S. Effect of dietary histidine on contents of carnosine and anserine in muscles of broilers. Anim. Sci. J. 2015, 86, 541–546. [Google Scholar] [CrossRef]
- Kim, S.-K.; Kim, Y.-M.; Baek, I.-K.; Auh, J.-H. Carnosine and anserine in chicken: Distribution, age-dependency and their anti-glycation activity. J. Food Sci. Anim. Resour. 2012, 32, 45–48. [Google Scholar] [CrossRef]
- Abasht, B.; Mutryn, M.F.; Michalek, R.D.; Lee, W.R. Oxidative stress and metabolic perturbations in wooden breast disorder in chickens. PLoS ONE 2016, 11, e0153750. [Google Scholar] [CrossRef]
- Qi, Y.; Zhang, K.; Tian, G.; Bai, S.; Ding, X.; Wang, J.; Peng, H.; Lv, L.; Xuan, Y.; Zeng, Q. Effects of dietary corn germ meal levels on growth performance, serum biochemical parameters, meat quality, and standardized ileal digestibility of amino acids in Pekin ducks. Poult. Sci. 2022, 101, 101779. [Google Scholar] [CrossRef]
- Calzada, E.; Onguka, O.; Claypool, S.M. Phosphatidylethanolamine metabolism in health and disease. Int. Rev. Cell Mol. Biol. 2016, 321, 29–88. [Google Scholar] [CrossRef] [PubMed]
- Böttinger, L.; Horvath, S.E.; Kleinschroth, T.; Hunte, C.; Daum, G.; Pfanner, N.; Becker, T. Phosphatidylethanolamine and cardiolipin differentially affect the stability of mitochondrial respiratory chain supercomplexes. J. Mol. Biol. 2012, 423, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Imhof, I.; Canivenc-Gansel, E.; Meyer, U.; Conzelmann, A. Phosphatidylethanolamine is the donor of the phosphorylethanolamine linked to the α1, 4-linked mannose of yeast GPI structures. Glycobiology 2000, 10, 1271–1275. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Agellon, L.B.; Allen, T.M.; Umeda, M.; Jewell, L.; Mason, A.; Vance, D.E. The ratio of phosphatidylcholine to phosphatidylethanolamine influences membrane integrity and steatohepatitis. Cell Metab. 2006, 3, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Chavez, L.O.; Leon, M.; Einav, S.; Varon, J. Beyond muscle destruction: A systematic review of rhabdomyolysis for clinical practice. Crit. Care 2016, 20, 135. [Google Scholar] [CrossRef]
- Romero-Aristizabal, C.; Marks, D.S.; Fontana, W.; Apfeld, J. Regulated spatial organization and sensitivity of cytosolic protein oxidation in Caenorhabditis elegans. Nat. Commun. 2014, 5, 5020. [Google Scholar] [CrossRef]
- Rohatgi, N.; Nielsen, T.K.; Bjørn, S.P.; Axelsson, I.; Paglia, G.; Voldborg, B.G.; Palsson, B.O.; Rolfsson, Ó. Biochemical characterization of human gluconokinase and the proposed metabolic impact of gluconic acid as determined by constraint based metabolic network analysis. PLoS ONE 2014, 9, e98760. [Google Scholar] [CrossRef]
- De Koning, T.J.; Snell, K.; Duran, M.; Berger, R.; Poll-The, B.-T.; Surtees, R. L-serine in disease and development. Biochem. J. 2003, 371, 653–661. [Google Scholar] [CrossRef]
- Mishra, R.C.; Tripathy, S.; Quest, D.; Desai, K.M.; Akhtar, J.; Dattani, I.D.; Gopalakrishnan, V. L-Serine lowers while glycine increases blood pressure in chronic L-NAME-treated and spontaneously hypertensive rats. Am. J. Hypertens. 2008, 26, 2339–2348. [Google Scholar] [CrossRef]
- Conte Camerino, D.; Tricarico, D.; Pierno, S.; Desaphy, J.F.; Liantonio, A.; Pusch, M.; Burdi, R.; Camerino, C.; Fraysse, B.; De Luca, A. Taurine and skeletal muscle disorders. Neurochem. Res. 2004, 29, 135–142. [Google Scholar] [CrossRef]
- Lambert, I.H. Regulation of the cellular content of the organic osmolyte taurine in mammalian cells. Neurochem. Res. 2004, 29, 27–63. [Google Scholar] [CrossRef] [PubMed]
- Toue, S.; Kodama, R.; Amao, M.; Kawamata, Y.; Kimura, T.; Sakai, R. Screening of toxicity biomarkers for methionine excess in rats. J. Nutr. 2006, 136, 1716S–1721S. [Google Scholar] [CrossRef] [PubMed]
- Krogdahl, Å. Digestion and absorption of lipids in poultry. J. Nutr. 1985, 115, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Nir, I.; Nitsan, Z.; Vax, A. The influence of force feeding and of protein supplementation to the diet on the metabolisable energy of diets, digestibility of nutrients, nitrogen retention and digestive enzymes output in geese. Ann. Biol. Anim. Biochim. Biophys. 1973, 13, 465–479. [Google Scholar] [CrossRef]
- Abasht, B.; Papah, M.B.; Qiu, J. Evidence of vascular endothelial dysfunction in Wooden Breast disorder in chickens: Insights through gene expression analysis, ultra-structural evaluation and supervised machine learning methods. PLoS ONE 2021, 16, e0243983. [Google Scholar] [CrossRef]
- Liu, R.; Kong, F.; Xing, S.; He, Z.; Bai, L.; Sun, J.; Tan, X.; Zhao, D.; Zhao, G.; Wen, J. Dominant changes in the breast muscle lipid profiles of broiler chickens with wooden breast syndrome revealed by lipidomics analyses. J. Anim. Sci. Biotechnol. 2022, 13, 93. [Google Scholar] [CrossRef]
- van Kempen, T.A.; Reijersen, M.; de Bruijn, C.; De Smet, S.; Michiels, J.; Traber, M.G.; Lauridsen, C. Vitamin E plasma kinetics in swine show low bioavailability and short half-life of all-rac-α-tocopheryl acetate. J. Anim. Sci. 2016, 94, 4188–4195. [Google Scholar] [CrossRef]
- Thompson, G.R. Lipid related consequences of intestinal malabsorption. Gut 1989, 30, 29–34. [Google Scholar] [CrossRef]
- Mutryn, M.F.; Brannick, E.M.; Fu, W.; Lee, W.R.; Abasht, B. Characterization of a novel chicken muscle disorder through differential gene expression and pathway analysis using RNA-sequencing. BMC Genom. 2015, 16, 399. [Google Scholar] [CrossRef]
- Mandatori, D.; Pelusi, L.; Schiavone, V.; Pipino, C.; Di Pietro, N.; Pandolfi, A. The dual role of vitamin K2 in “bone-vascular crosstalk”: Opposite effects on bone loss and vascular calcification. Nutrients 2021, 13, 1222. [Google Scholar] [CrossRef]
- Kong, F.; Zhao, G.; He, Z.; Sun, J.; Wang, X.; Liu, D.; Zhu, D.; Liu, R.; Wen, J. Serum Creatine Kinase as a Biomarker to Predict Wooden Breast in vivo for Chicken Breeding. Front. Physiol. 2021, 12, 711711. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, S.; Torres, A.G.; Ribas de Pouplana, L. Inosine in biology and disease. Genes 2021, 12, 600. [Google Scholar] [CrossRef] [PubMed]
- Jankowski, V.; Van Der Giet, M.; Mischak, H.; Morgan, M.; Zidek, W.; Jankowski, J. Dinucleoside polyphosphates: Strong endogenous agonists of the purinergic system. Br. J. Pharmacol. 2009, 157, 1142–1153. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dong, X.; Puolanne, E.; Ertbjerg, P. Effect of wooden breast degree on lipid and protein oxidation and citrate synthase activity of chicken pectoralis major muscle. LWT 2022, 154, 112884. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).