
A Review of Oxylipins in Alzheimer’s Disease and Related Dementias (ADRD): Potential Therapeutic Targets for the Modulation of Vascular Tone and Inflammation



Abstract
1. Introduction
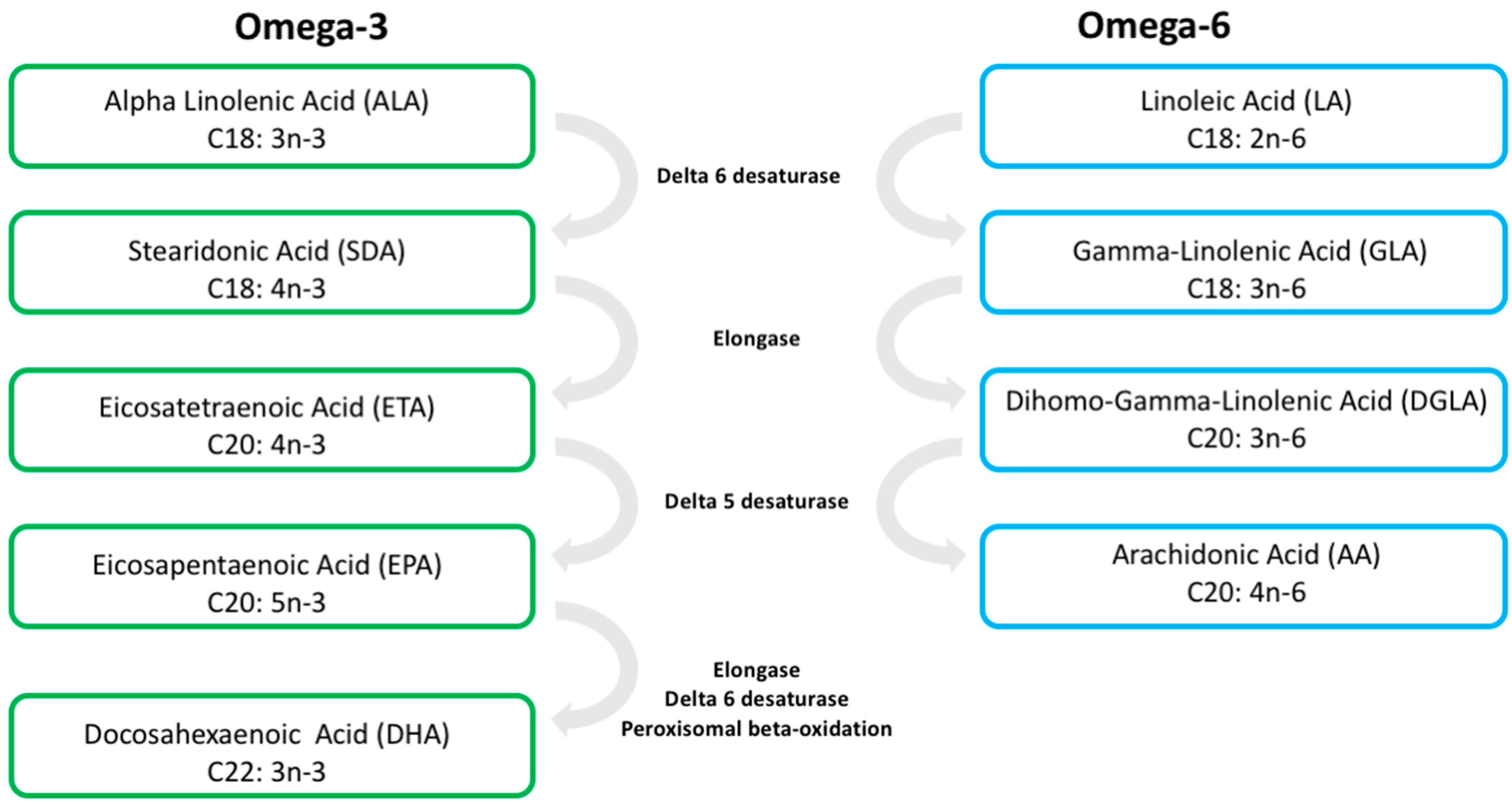
2. Long-Chain PUFAs
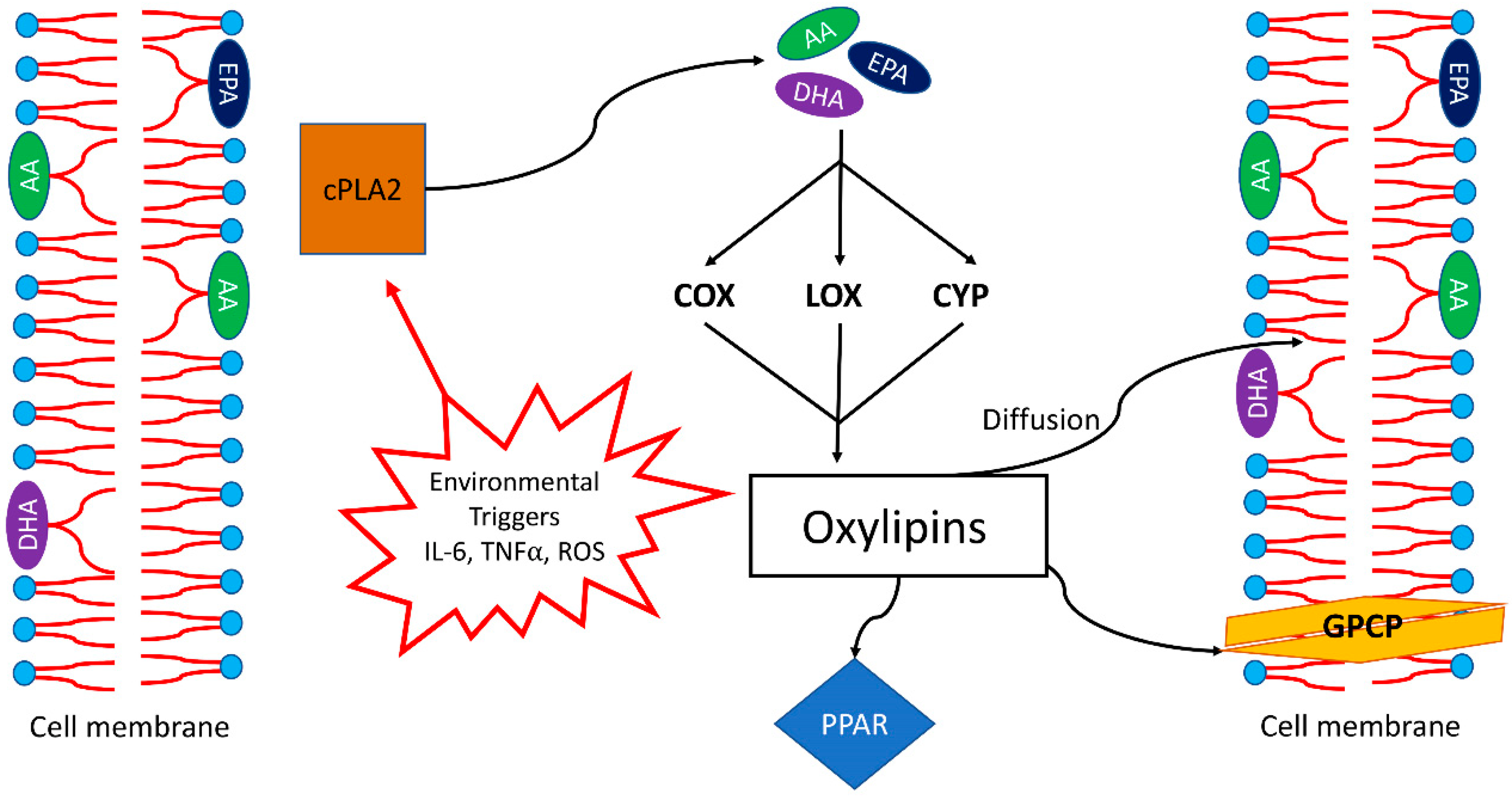
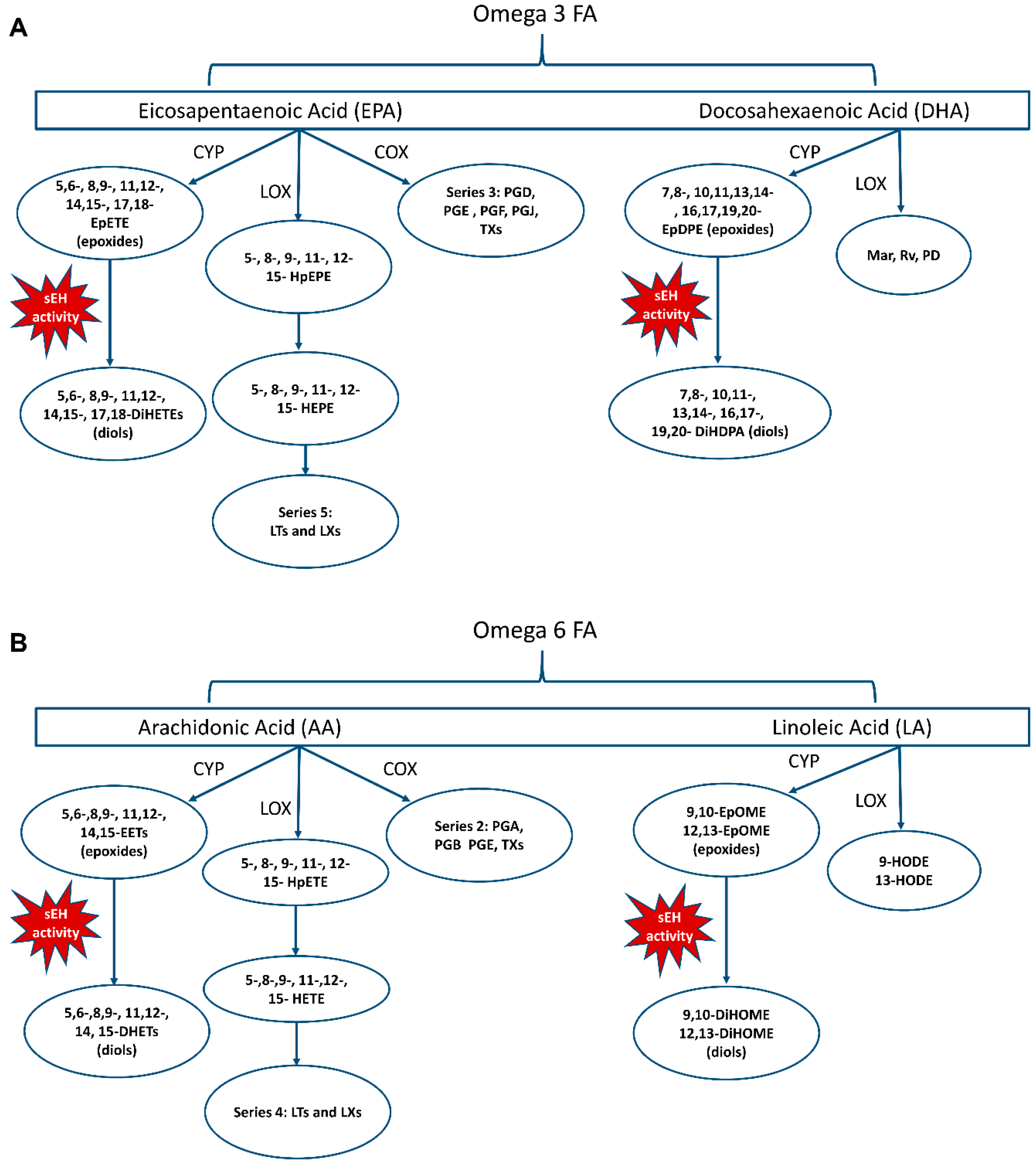
3. Oxylipins
4. n-3 PUFAs and Oxylipins: Animal Studies
5. n-3 PUFAs and Brain Fatty Acid Composition: Animal Studies
6. Oxylipins and the Gut Microbiome: Animal Studies
7. n-PUFAs in ADRD: Human Studies
8. Specific Oxylipins Associated with Vascular Risk Factors
8.1. Linoleic Acid—Omega-6-Derived Oxylipins: HODEs, EpOMES, and DiHOMES
8.2. HODES
8.3. HEPEs
8.4. CYP and sEH
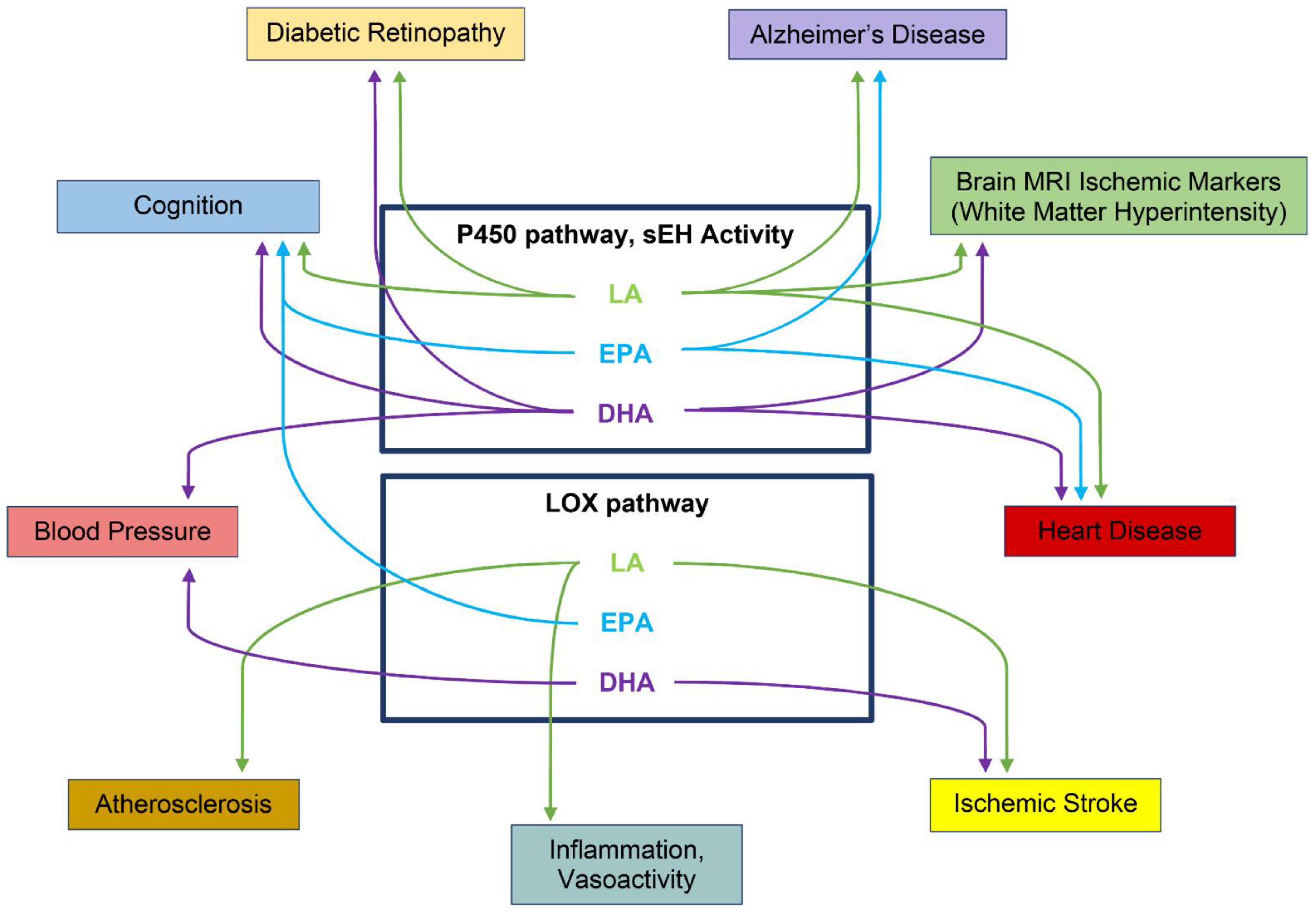
9. Oxylipins Altered in ADRD Risk and ADRD Conditions
9.1. Cardiovascular
9.2. Diabetic Retinopathy (DR)
9.3. Hypertension
9.4. Vascular Cognitive Impairment and Dementia (VCID)
9.5. Mild Cognitive Impairment (MCI) and Alzheimer’s Disease (AD)
10. Summary and Future Directions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- WHO. Dementia Facts 2021. Available online: https://www.who.int/en/news-room/fact-sheets/detail/dementia (accessed on 28 August 2022).
- Bowler, J.V. Vascular cognitive impairment. J. Neurol. Neurosurg. Psychiatry 2005, 76 (Suppl. 5), v35–v44. [Google Scholar] [CrossRef] [PubMed]
- Azarpazhooh, M.R.; Avan, A.; Cipriano, L.E.; Munoz, D.G.; Sposato, L.A.; Hachinski, V. Concomitant vascular and neurodegenerative pathologies double the risk of dementia. Alzheimer’s Dement. 2018, 14, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Gelber, R.P.; Launer, L.J.; White, L.R. The Honolulu-Asia Aging Study: Epidemiologic and neuropathologic research on cognitive impairment. Curr. Alzheimer Res. 2012, 9, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.A.; Arvanitakis, Z.; Bang, W.; Bennett, D.A. Mixed brain pathologies account for most dementia cases in community-dwelling older persons. Neurology 2007, 69, 2197–2204. [Google Scholar] [CrossRef]
- Gottesman, R.F.; Albert, M.S.; Alonso, A.; Coker, L.H.; Coresh, J.; Davis, S.M.; Deal, J.A.; McKhann, G.M.; Mosley, T.H.; Sharrett, A.R.; et al. Associations between Midlife Vascular Risk Factors and 25-Year Incident Dementia in the Atherosclerosis Risk in Communities (ARIC) Cohort. JAMA Neurol. 2017, 74, 1246–1254. [Google Scholar] [CrossRef]
- Knopman, D.S.; Roberts, R. Vascular risk factors: Imaging and neuropathologic correlates. J. Alzheimer’s Dis. 2010, 20, 699–709. [Google Scholar] [CrossRef]
- Petrovitch, H.; White, L.R.; Izmirilian, G.; Ross, G.W.; Havlik, R.J.; Markesbery, W.; Nelson, J.; Davis, D.; Hardman, J.; Foley, D.; et al. Midlife blood pressure and neuritic plaques, neurofibrillary tangles, and brain weight at death: The HAAS. Honolulu-Asia aging Study. Neurobiol. Aging 2000, 21, 57–62. [Google Scholar] [CrossRef]
- Qiu, C.; Winblad, B.; Marengoni, A.; Klarin, I.; Fastbom, J.; Fratiglioni, L. Heart failure and risk of dementia and Alzheimer disease: A population-based cohort study. Arch. Intern. Med. 2006, 166, 1003–1008. [Google Scholar] [CrossRef]
- Erten-Lyons, D.; Woltjer, R.; Kaye, J.; Mattek, N.; Dodge, H.H.; Green, S.; Tran, H.; Howieson, D.B.; Wild, K.; Silbert, L.C. Neuropathologic basis of white matter hyperintensity accumulation with advanced age. Neurology 2013, 81, 977–983. [Google Scholar] [CrossRef]
- Honig, L.S.; Kukull, W.; Mayeux, R. Atherosclerosis and AD: Analysis of data from the US National Alzheimer’s Coordinating Center. Neurology 2005, 64, 494–500. [Google Scholar] [CrossRef]
- Yarchoan, M.; Xie, S.X.; Kling, M.A.; Toledo, J.B.; Wolk, D.A.; Lee, E.B.; Van Deerlin, V.; Lee, V.M.-Y.; Trojanowski, J.Q.; Arnold, S.E. Cerebrovascular atherosclerosis correlates with Alzheimer pathology in neurodegenerative dementias. Brain 2012, 135, 3749–3756. [Google Scholar] [CrossRef]
- Wolters, F.J.; Ikram, M.A. Epidemiology of Vascular Dementia. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1542–1549. [Google Scholar] [CrossRef]
- Arnett, D.K.; Blumenthal, R.S.; Albert, M.A.; Buroker, A.B.; Goldberger, Z.D.; Hahn, E.J.; Himmelfarb, C.D.; Khera, A.; Lloyd-Jones, D.; McEvoy, J.W.; et al. 2019 ACC/AHA Guideline on the Primary Prevention of Cardiovascular Disease: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. J. Am. Coll. Cardiol. 2019, 74, e177–e232. [Google Scholar] [CrossRef]
- Kleindorfer, D.O.; Towfighi, A.; Chaturvedi, S.; Cockroft, K.M.; Gutierrez, J.; Lombardi-Hill, D.; Kamel, H.; Kernan, W.N.; Kittner, S.J.; Leira, E.C.; et al. 2021 Guideline for the Prevention of Stroke in Patients with Stroke and Transient Ischemic Attack: A Guideline from the American Heart Association/American Stroke Association. Stroke 2021, 52, e364–e467. [Google Scholar] [CrossRef] [PubMed]
- Pischon, T.; Hankinson, S.E.; Hotamisligil, G.S.; Rifai, N.; Willett, W.C.; Rimm, E.B. Habitual dietary intake of n-3 and n-6 fatty acids in relation to inflammatory markers among US men and women. Circulation 2003, 108, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P.; DiNicolantonio, J.J. The importance of a balanced omega-6 to omega-3 ratio in the prevention and management of obesity. Open Heart 2016, 3, e000385. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Exp. Biol. Med. 2008, 233, 674–688. [Google Scholar] [CrossRef] [PubMed]
- Mariamenatu, A.H.; Abdu, E.M. Overconsumption of Omega-6 Polyunsaturated Fatty Acids (PUFAs) versus Deficiency of Omega-3 PUFAs in Modern-Day Diets: The Disturbing Factor for Their “Balanced Antagonistic Metabolic Functions” in the Human Body. J. Lipids 2021, 2021, 8848161. [Google Scholar] [CrossRef]
- Simopoulos, A.P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef]
- de Lorgeril, M.; Renaud, S.; Mamelle, N.; Salen, P.; Monjaud, I.; Mamelle, N.; Martin, J.; Guidollet, J.; Touboul, P.; Delaye, J. Mediterranean alpha-linolenic acid-rich diet in secondary prevention of coronary heart disease. Lancet 1994, 343, 1454–1459. [Google Scholar] [CrossRef]
- Mozaffarian, D.; Lemaitre, R.N.; King, I.B.; Song, X.; Huang, H.; Sacks, F.M. Plasma phospholipid long-chain omega-3 fatty acids and total and cause-specific mortality in older adults: A cohort study. Ann. Intern. Med. 2013, 158, 515–525. [Google Scholar] [CrossRef]
- Santos, H.O.; Price, J.C.; Bueno, A.A. Beyond Fish Oil Supplementation: The Effects of Alternative Plant Sources of Omega-3 Polyunsaturated Fatty Acids upon Lipid Indexes and Cardiometabolic Biomarkers-An Overview. Nutrients 2020, 12, 3519. [Google Scholar] [CrossRef]
- Taha, A.Y. Linoleic acid-good or bad for the brain? NPJ Sci. Food 2020, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Gabbs, M.; Leng, S.; Devassy, J.G.; Monirujjaman, M.; Aukema, H.M. Advances in Our Understanding of Oxylipins Derived from Dietary PUFAs. Adv. Nutr. 2015, 6, 513–540. [Google Scholar] [CrossRef] [PubMed]
- Tourdot, B.E.; Ahmed, I.; Holinstat, M. The emerging role of oxylipins in thrombosis and diabetes. Front. Pharmacol. 2014, 4, 176. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.Y.; Geng, X.; Teng, T.; Yang, B.; Appenteng, M.K.; Greenlief, C.M.; Lee, J.C. Dynamic Role of Phospholipases A2 in Health and Diseases in the Central Nervous System. Cells 2021, 10, 2963. [Google Scholar] [CrossRef]
- Ostermann, A.I.; Waindok, P.; Schmidt, M.J.; Chiu, C.Y.; Smyl, C.; Rohwer, N.; Weylandt, K.-H.; Schebb, N.H. Modulation of the endogenous omega-3 fatty acid and oxylipin profile in vivo-A comparison of the fat-1 transgenic mouse with C57BL/6 wildtype mice on an omega-3 fatty acid enriched diet. PLoS ONE 2017, 12, e0184470. [Google Scholar] [CrossRef]
- Ramsden, C.E.; Hennebelle, M.; Schuster, S.; Keyes, G.S.; Johnson, C.D.; Kirpich, I.A.; Dahlen, J.E.; Horowitz, M.S.; Zamora, D.; Feldstein, A.E.; et al. Effects of diets enriched in linoleic acid and its peroxidation products on brain fatty acids, oxylipins, and aldehydes in mice. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 1206–1213. [Google Scholar] [CrossRef]
- Cudaback, E.; Jorstad, N.L.; Yang, Y.; Montine, T.J.; Keene, C.D. Therapeutic implications of the prostaglandin pathway in Alzheimer’s disease. Biochem. Pharmacol. 2014, 88, 565–572. [Google Scholar] [CrossRef]
- Caligiuri, S.P.B.; Parikh, M.; Stamenkovic, A.; Pierce, G.N.; Aukema, H.M. Dietary modulation of oxylipins in cardiovascular disease and aging. Am. J. Physiol. Heart Circ. Physiol. 2017, 313, H903–H918. [Google Scholar] [CrossRef]
- Harris, T.R.; Hammock, B.D. Soluble epoxide hydrolase: Gene structure, expression and deletion. Gene 2013, 526, 61–74. [Google Scholar] [CrossRef] [PubMed]
- McReynolds, C.; Morisseau, C.; Wagner, K.; Hammock, B. Epoxy Fatty Acids Are Promising Targets for Treatment of Pain, Cardiovascular Disease and Other Indications Characterized by Mitochondrial Dysfunction, Endoplasmic Stress and Inflammation. Adv. Exp. Med. Biol. 2020, 1274, 71–99. [Google Scholar] [CrossRef] [PubMed]
- Enayetallah, A.E.; French, R.A.; Thibodeau, M.S.; Grant, D.F. Distribution of soluble epoxide hydrolase and of cytochrome P450 2C8, 2C9, and 2J2 in human tissues. J. Histochem. Cytochem. 2004, 52, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Sura, P.; Sura, R.; Enayetallah, A.E.; Grant, D.F. Distribution and expression of soluble epoxide hydrolase in human brain. J. Histochem. Cytochem. 2008, 56, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Imig, J.D.; Zhao, X.; Capdevila, J.H.; Morisseau, C.; Hammock, B.D. Soluble epoxide hydrolase inhibition lowers arterial blood pressure in angiotensin II hypertension. Hypertension 2002, 39, 690–694. [Google Scholar] [CrossRef]
- Imig, J.D.; Zhao, X.; Zaharis, C.Z.; Olearczyk, J.J.; Pollock, D.M.; Newman, J.; Kim, I.-H.; Watanabe, T.; Hammock, B.D. An orally active epoxide hydrolase inhibitor lowers blood pressure and provides renal protection in salt-sensitive hypertension. Hypertension 2005, 46, 975–981. [Google Scholar] [CrossRef]
- Martini, R.P.; Ward, J.; Siler, D.A.; Eastman, J.M.; Nelson, J.W.; Borkar, R.N.; Alkayed, N.J.; Dogan, A.; Cetas, J.S. Genetic variation in soluble epoxide hydrolase: Association with outcome after aneurysmal subarachnoid hemorrhage. J. Neurosurg. 2014, 121, 1359–1366. [Google Scholar] [CrossRef]
- Saleh, R.N.M.; West, A.L.; Ostermann, A.I.; Schebb, N.H.; Calder, P.C.; Minihane, A.M. APOE Genotype Modifies the Plasma Oxylipin Response to Omega-3 Polyunsaturated Fatty Acid Supplementation in Healthy Individuals. Front. Nutr. 2021, 8, 723813. [Google Scholar] [CrossRef]
- Quinn, J.F.; Raman, R.; Thomas, R.G.; Yurko-Mauro, K.; Nelson, E.B.; Van Dyck, C.; Galvin, J.E.; Emond, J.; Jack, C.R.; Weiner, M.; et al. Docosahexaenoic acid supplementation and cognitive decline in Alzheimer disease: A randomized trial. JAMA 2010, 304, 1903–1911. [Google Scholar] [CrossRef] [PubMed]
- Bazan, N.G. The docosanoid neuroprotectin D1 induces homeostatic regulation of neuroinflammation and cell survival. Prostaglandins Leukot. Essent. Fat. Acids 2013, 88, 127–129. [Google Scholar] [CrossRef][Green Version]
- Bazan, N.G. Docosanoids and elovanoids from omega-3 fatty acids are pro-homeostatic modulators of inflammatory responses, cell damage and neuroprotection. Mol. Asp. Med. 2018, 64, 18–33. [Google Scholar] [CrossRef]
- Noverr, M.C.; Erb-Downward, J.R.; Huffnagle, G.B. Production of eicosanoids and other oxylipins by pathogenic eukaryotic microbes. Clin. Microbiol. Rev. 2003, 16, 517–533. [Google Scholar] [CrossRef]
- Bellien, J.; Joannides, R. Epoxyeicosatrienoic acid pathway in human health and diseases. J. Cardiovasc. Pharmacol. 2013, 61, 188–196. [Google Scholar] [CrossRef]
- Gonzalez-Fernandez, E.; Liu, Y.; Auchus, A.P.; Fan, F.; Roman, R.J. Vascular contributions to cognitive impairment and dementia: The emerging role of 20-HETE. Clin. Sci. 2021, 135, 1929–1944. [Google Scholar] [CrossRef] [PubMed]
- Imig, J.D. Epoxyeicosanoids in hypertension. Physiol. Res. 2019, 68, 695–704. [Google Scholar] [CrossRef]
- MacVicar, B.A.; Newman, E.A. Astrocyte regulation of blood flow in the brain. Cold Spring Harb. Perspect. Biol. 2015, 7, a020388. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.M.; Liu, X.; Alkayed, N.J. Cytochrome P450 eicosanoids in cerebrovascular function and disease. Pharmacol. Ther. 2017, 179, 31–46. [Google Scholar] [CrossRef]
- Biringer, R.G. The Role of Eicosanoids in Alzheimer’s Disease. Int. J. Environ. Res. Public Health 2019, 16, 2560. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Ghosh, A.; Lin, J.; Zhang, C.; Pan, Y.; Thakur, A.; Singh, K.; Hong, H.; Tang, S. 5-lipoxygenase pathway and its downstream cysteinyl leukotrienes as potential therapeutic targets for Alzheimer’s disease. Brain Behav. Immun. 2020, 88, 844–855. [Google Scholar] [CrossRef] [PubMed]
- Michael, J.; Marschallinger, J.; Aigner, L. The leukotriene signaling pathway: A druggable target in Alzheimer’s disease. Drug Discov. Today 2019, 24, 505–516. [Google Scholar] [CrossRef]
- Tarantini, S.; Tran, C.H.T.; Gordon, G.R.; Ungvari, Z.; Csiszar, A. Impaired neurovascular coupling in aging and Alzheimer’s disease: Contribution of astrocyte dysfunction and endothelial impairment to cognitive decline. Exp. Gerontol. 2017, 94, 52–58. [Google Scholar] [CrossRef]
- Chamani, S.; Bianconi, V.; Tasbandi, A.; Pirro, M.; Barreto, G.E.; Jamialahmadi, T.; Sahebkar, A. Resolution of Inflammation in Neurodegenerative Diseases: The Role of Resolvins. Mediat. Inflamm. 2020, 2020, 3267172. [Google Scholar] [CrossRef]
- Simonetto, M.; Infante, M.; Sacco, R.L.; Rundek, T.; Della-Morte, D. A Novel Anti-Inflammatory Role of Omega-3 PUFAs in Prevention and Treatment of Atherosclerosis and Vascular Cognitive Impairment and Dementia. Nutrients 2019, 11, 2279. [Google Scholar] [CrossRef] [PubMed]
- Nguemeni, C.; Delplanque, B.; Rovere, C.; Simon-Rousseau, N.; Gandin, C.; Agnani, G.; Nahon, J.L.; Heurteaux, C.; Blondeau, N. Dietary supplementation of alpha-linolenic acid in an enriched rapeseed oil diet protects from stroke. Pharmacol. Res. 2010, 61, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Ren, H.; Yao, X.; Shi, Z.; Liang, F.; Kang, J.X.; Wan, J.-B.; Pei, Z.; Su, K.-P.; Su, H. Enriched Brain Omega-3 Polyunsaturated Fatty Acids Confer Neuroprotection against Microinfarction. EBioMedicine 2018, 32, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Saito, H.; Minami, M.; Togashi, H.; Nakamura, N.; Ueno, K.; Shimamura, K.; Nemoto, M.; Parvez, H. Docosahexaenoic acid attenuated hypertension and vascular dementia in stroke-prone spontaneously hypertensive rats. Neurotoxicol. Teratol. 2002, 24, 683–693. [Google Scholar] [CrossRef]
- de Wilde, M.C.; Farkas, E.; Gerrits, M.; Kiliaan, A.J.; Luiten, P.G. The effect of n-3 polyunsaturated fatty acid-rich diets on cognitive and cerebrovascular parameters in chronic cerebral hypoperfusion. Brain Res. 2002, 947, 166–173. [Google Scholar] [CrossRef]
- Hu, J.; Dziumbla, S.; Lin, J.; Bibli, S.I.; Zukunft, S.; de Mos, J.; Awwad, K.; Frömel, T.; Jungmann, A.; Devraj, K.; et al. Inhibition of soluble epoxide hydrolase prevents diabetic retinopathy. Nature 2017, 552, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Zuloaga, K.L.; Zhang, W.; Roese, N.E.; Alkayed, N.J. Soluble epoxide hydrolase gene deletion improves blood flow and reduces infarct size after cerebral ischemia in reproductively senescent female mice. Front. Pharmacol. 2014, 5, 290. [Google Scholar] [CrossRef]
- Iliff, J.J.; Alkayed, N.J. Soluble Epoxide Hydrolase Inhibition: Targeting Multiple Mechanisms of Ischemic Brain Injury with a Single Agent. Future Neurol. 2009, 4, 179–199. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.W.; Zhang, W.; Alkayed, N.J.; Koerner, I.P. Peroxisomal translocation of soluble epoxide hydrolase protects against ischemic stroke injury. J. Cereb. Blood Flow Metab. 2015, 35, 1416–1420. [Google Scholar] [CrossRef]
- Takeyama, E.; Islam, A.; Watanabe, N.; Tsubaki, H.; Fukushima, M.; Mamun, M.A.; Sato, S.; Eto, F.; Yao, I.; Ito, T.K.; et al. Dietary Intake of Green Nut Oil or DHA Ameliorates DHA Distribution in the Brain of a Mouse Model of Dementia Accompanied by Memory Recovery. Nutrients 2019, 11, 2371. [Google Scholar] [CrossRef]
- Lee, A.Y.; Choi, J.M.; Lee, J.; Lee, M.H.; Lee, S.; Cho, E.J. Effects of Vegetable Oils with Different Fatty Acid Compositions on Cognition and Memory Ability in Abeta25-35-Induced Alzheimer’s Disease Mouse Model. J. Med. Food 2016, 19, 912–921. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.Y.; Lee, M.H.; Lee, S.; Cho, E.J. Alpha-Linolenic Acid from Perilla frutescens var. japonica Oil Protects Abeta-Induced Cognitive Impairment through Regulation of APP Processing and Abeta Degradation. J. Agric. Food Chem. 2017, 65, 10719–10729. [Google Scholar] [CrossRef] [PubMed]
- Qu, M.H.; Yang, X.; Wang, Y.; Tang, Q.; Han, H.; Wang, J.; Du Wang, G.; Xue, C.; Gao, Z. Docosahexaenoic Acid-Phosphatidylcholine Improves Cognitive Deficits in an Abeta23-35-Induced Alzheimer’s Disease Rat Model. Curr. Top. Med. Chem. 2016, 16, 558–564. [Google Scholar] [CrossRef]
- Walczewska, A.; Stepien, T.; Bewicz-Binkowska, D.; Zgorzynska, E. The role of docosahexaenoic acid in neuronal function. Postepy Hig. I Med. Dosw. 2011, 65, 314–327. [Google Scholar] [CrossRef] [PubMed]
- Domenichiello, A.F.; Chen, C.T.; Trepanier, M.O.; Stavro, P.M.; Bazinet, R.P. Whole body synthesis rates of DHA from alpha-linolenic acid are greater than brain DHA accretion and uptake rates in adult rats. J. Lipid Res. 2014, 55, 62–74. [Google Scholar] [CrossRef] [PubMed]
- Williard, D.E.; Harmon, S.D.; Kaduce, T.L.; Preuss, M.; Moore, S.A.; Robbins, M.E.; Spector, A.A. Docosahexaenoic acid synthesis from n-3 polyunsaturated fatty acids in differentiated rat brain astrocytes. J. Lipid Res. 2001, 42, 1368–1376. [Google Scholar] [CrossRef]
- Moore, S.A.; Yoder, E.; Murphy, S.; Dutton, G.R.; Spector, A.A. Astrocytes, not neurons, produce docosahexaenoic acid (22:6 omega-3) and arachidonic acid (20:4 omega-6). J. Neurochem. 1991, 56, 518–524. [Google Scholar] [CrossRef]
- Moore, S.A.; Yoder, E.; Spector, A.A. Role of the blood-brain barrier in the formation of long-chain omega-3 and omega-6 fatty acids from essential fatty acid precursors. J. Neurochem. 1990, 55, 391–402. [Google Scholar] [CrossRef]
- Sihag, J.; Di Marzo, V. (Wh)olistic (E)ndocannabinoidome-Microbiome-Axis Modulation through (N)utrition (WHEN) to Curb Obesity and Related Disorders. Lipids Health Dis. 2022, 21, 9. [Google Scholar] [CrossRef] [PubMed]
- Wilmanski, T.; Diener, C.; Rappaport, N.; Patwardhan, S.; Wiedrick, J.; Lapidus, J.; Earls, J.C.; Zimmer, A.; Glusman, G.; Robinson, M.; et al. Gut microbiome pattern reflects healthy ageing and predicts survival in humans. Nat. Metab. 2021, 3, 274–286. [Google Scholar] [CrossRef] [PubMed]
- Sampson, T.; Debelius, J.; Thron, T.; Janssen, S.; Shastri, G.; Ilhan, Z.; Challis, C.; Schretter, C.; Rocha, S.; Gradinaru, V.; et al. Gut microbiota regulate motor deficits and neurinflammation in a model of Parkinson’s disease. Cell 2016, 167, 1469–1480. [Google Scholar] [CrossRef] [PubMed]
- Torres, E.; Akinyeke, T.; Stagaman, K.; Duvoisin, R.; Meshul, C.K.; Sharpton, T.J.; Raber, J. Effects of sub-chronic MPTP exposure on behavioral and cognitive performance and the microbiome of wild-type and mGlu8 knockout female and male mice. Front. Behav. Neurosci. 2018, 12, 140. [Google Scholar] [CrossRef]
- Kundu, P.; Torres, E.; Stagaman, K.; Kasschau, K.; Okhovat, M.; Holden, S.; Ward, S.; Nevonen, K.; Davis, B.; Saito, T.; et al. Integrated analysis of behavioral, epigenetic, and gut microbiome analyses in AppNL-G-F, AppNL-F, and wild type mice. Sci. Rep. 2021, 11, 4678. [Google Scholar] [CrossRef]
- Kundu, P.; Stagaman, K.; Kasschau, K.; Holden, S.; Shulzhenko, N.; Sharpton, T.; Raber, J. Fecal implants from AppNL-G-F and AppNL-G-F/E4 donor mice sufficient to induce behavioral phenotypes in germ-free mice. Front. Behav. Neurosci. 2022, 16, 791128. [Google Scholar] [CrossRef] [PubMed]
- Muscogiuri, G.; Cantone, E.; Cassarano, S.; Tuccinardi, D.; Barrea, L.; Savastano, S.; Colao, A.; on behalf of the OPERA group. Gut microbiota: A new path to treat obesity. Int. J. Obes. Suppl. 2019, 9, 10–19. [Google Scholar] [CrossRef]
- Torres-Fuentes, C.; Schellekens, H.; Dinan, T.; Cryan, J.F. The microbiota–gut–brain axis in obesity. Lancet Gastroenterol. Hepatol. 2017, 2, 747–756. [Google Scholar] [CrossRef]
- Kamada, N.; Seo, S.; Chen, G.; Nunez, G. Role of the gut microbiota in immunity and inflammatory disease. Nat. Rev. Immunol. 2013, 13, 321–335. [Google Scholar] [CrossRef]
- Clemente, J.; Manasson, J.; Scher, J. The role of the gut microbiome in systemic inflammatory disease. BMJ 2018, 360, j5145. [Google Scholar] [CrossRef]
- Avila-Roman, J.; Arreaza-Gil, V.; Cortes-Espinar, A.; Soliz-Rueda, J.; Mulero, M.; Muguerza, B.; Arola-Arnal, A.; Arola, L.; Torres-Fuentes, C. Impact of gut microbiota on plasma oxylipins profile under healthy and obesogenic conditions. Clin. Nutr. 2021, 40, 1475–1486. [Google Scholar] [CrossRef]
- Deol, P.; Fahrmann, J.; Yang, J.; Evans, J.; Rizo, A.; Grapov, D.; Salemi, M.; Wanichthanarak, K.; Fiehn, O.; Phinney, B.; et al. Omega-6 and omega-3 oxylipins are implicated in soybean oil-induced obesity in mice. Sci. Rep. 2017, 7, 12488. [Google Scholar] [CrossRef]
- He, R.; Chen, Y.; Cai, Q. The role of the LTB4-BLT1 axis in health and disease. Pharmacol. Res. 2020, 158, 104857. [Google Scholar] [CrossRef]
- Marrone, C.; Coccurello, R. Dietary Fatty Acids and Microbiota-Brain Communication in Neuropsychiatric Diseases. Biomolecules 2020, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Tariska, P.; Urbanics, K.; Knolmayer, J.; Meszaros, A. Clinical symptoms of Alzheimer disease. Orv. Hetil. 1995, 136, 867–873. [Google Scholar]
- Syeda, T.; Sanchez-Tapia, M.; Orta, I.; Granados-Portillo, O.; Perez-Jimenez, L.; Rodriguez-Callejas, J.D.; Toribio, S.; Silva-Lucero, M.-D.; Rivera, A.-L.; Tovar, A.R.; et al. Bioactive Foods Decrease Liver and Brain Alterations Induced by a High-Fat-Sucrose Diet through Restoration of Gut Microbiota and Antioxidant Enzymes. Nutrients 2022, 14, 22. [Google Scholar] [CrossRef] [PubMed]
- Babcock, T.; Helton, W.S.; Espat, N.J. Eicosapentaenoic acid (EPA): An antiinflammatory omega-3 fat with potential clinical applications. Nutrition 2000, 16, 1116–1118. [Google Scholar] [CrossRef]
- Abdelhamid, A.S.; Brown, T.J.; Brainard, J.S.; Biswas, P.; Thorpe, G.C.; Moore, H.J.; Deane, K.H.; Summerbell, C.D.; Worthington, H.V.; Song, F.; et al. Omega-3 fatty acids for the primary and secondary prevention of cardiovascular disease. Cochrane Database Syst. Rev. 2020, 3, CD003177. [Google Scholar] [CrossRef]
- Shen, S.; Gong, C.; Jin, K.; Zhou, L.; Xiao, Y.; Ma, L. Omega-3 Fatty Acid Supplementation and Coronary Heart Disease Risks: A Meta-Analysis of Randomized Controlled Clinical Trials. Front. Nutr. 2022, 9, 809311. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.E.; Van Elswyk, M.; Alexander, D.D. Long-chain omega-3 fatty acids eicosapentaenoic acid and docosahexaenoic acid and blood pressure: A meta-analysis of randomized controlled trials. Am. J. Hypertens. 2014, 27, 885–896. [Google Scholar] [CrossRef]
- Hadley, K.B.; Ryan, A.S.; Forsyth, S.; Gautier, S.; Salem, N., Jr. The Essentiality of Arachidonic Acid in Infant Development. Nutrients 2016, 8, 216. [Google Scholar] [CrossRef] [PubMed]
- Sambra, V.; Echeverria, F.; Valenzuela, A.; Chouinard-Watkins, R.; Valenzuela, R. Docosahexaenoic and Arachidonic Acids as Neuroprotective Nutrients throughout the Life Cycle. Nutrients 2021, 13, 986. [Google Scholar] [CrossRef] [PubMed]
- Attwell, D.; Buchan, A.M.; Charpak, S.; Lauritzen, M.; Macvicar, B.A.; Newman, E.A. Glial and neuronal control of brain blood flow. Nature 2010, 468, 232–243. [Google Scholar] [CrossRef]
- Stackhouse, T.L.; Mishra, A. Neurovascular Coupling in Development and Disease: Focus on Astrocytes. Front. Cell Dev. Biol. 2021, 9, 702832. [Google Scholar] [CrossRef] [PubMed]
- Barberger-Gateau, P.; Raffaitin, C.; Letenneur, L.; Berr, C.; Tzourio, C.; Dartigues, J.F.; Alpérovitch, A. Dietary patterns and risk of dementia: The Three-City cohort study. Neurology 2007, 69, 1921–1930. [Google Scholar] [CrossRef]
- Devore, E.E.; Grodstein, F.; van Rooij, F.J.; Hofman, A.; Rosner, B.; Stampfer, M.J.; Witteman, J.C.; Breteler, M.M. Dietary intake of fish and omega-3 fatty acids in relation to long-term dementia risk. Am. J. Clin. Nutr. 2009, 90, 170–176. [Google Scholar] [CrossRef]
- Morris, M.C.; Evans, D.A.; Bienias, J.L.; Tangney, C.C.; Bennett, D.A.; Wilson, R.S.; Aggarwal, N.; Schneider, J. Consumption of fish and n-3 fatty acids and risk of incident Alzheimer disease. Arch. Neurol. 2003, 60, 940–946. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, J.; Qiu, J.; Li, Y.; Wang, J.; Jiao, J. Intakes of fish and polyunsaturated fatty acids and mild-to-severe cognitive impairment risks: A dose-response meta-analysis of 21 cohort studies. Am. J. Clin. Nutr. 2016, 103, 330–340. [Google Scholar] [CrossRef]
- Freund-Levi, Y.; Eriksdotter-Jonhagen, M.; Cederholm, T.; Basun, H.; Faxen-Irving, G.; Garlind, A.; Vedin, I.; Vessby, B.; Wahlund, L.-O.; Palmblad, J. Omega-3 fatty acid treatment in 174 patients with mild to moderate Alzheimer disease: OmegAD study: A randomized double-blind trial. Arch. Neurol. 2006, 63, 1402–1408. [Google Scholar] [CrossRef] [PubMed]
- Andrieu, S.; Guyonnet, S.; Coley, N.; Cantet, C.; Bonnefoy, M.; Bordes, S.; Bories, L.; Cufi, M.-N.; Dantoine, T.; Dartigues, J.-F.; et al. Effect of long-term omega 3 polyunsaturated fatty acid supplementation with or without multidomain intervention on cognitive function in elderly adults with memory complaints (MAPT): A randomised, placebo-controlled trial. Lancet Neurol. 2017, 16, 377–389. [Google Scholar] [CrossRef]
- Hennebelle, M.; Zhang, Z.; Metherel, A.H.; Kitson, A.P.; Otoki, Y.; Richardson, C.E.; Yang, J.; Lee, K.S.S.; Hammock, B.D.; Zhang, L.; et al. Linoleic acid participates in the response to ischemic brain injury through oxidized metabolites that regulate neurotransmission. Sci. Rep. 2017, 7, 4342. [Google Scholar] [CrossRef]
- Bojic, L.A.; McLaren, D.G.; Harms, A.C.; Hankemeier, T.; Dane, A.; Wang, S.P.; Rosa, R.; Previs, S.F.; Johns, U.G.; Castro-Perez, J.M. Quantitative profiling of oxylipins in plasma and atherosclerotic plaques of hypercholesterolemic rabbits. Anal. Bioanal. Chem. 2016, 408, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Rolin, J.; Maghazachi, A.A. Implications of chemokines, chemokine receptors, and inflammatory lipids in atherosclerosis. J. Leukoc. Biol. 2014, 95, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Shibata, N.; Toi, S.; Shibata, T.; Uchida, K.; Itabe, H.; Sawada, T.; Kawamata, T.; Okada, Y.; Uchiyama, S.; Kobayashi, M. Immunohistochemical detection of 13(R)-hydroxyoctadecadienoic acid in atherosclerotic plaques of human carotid arteries using a novel specific antibody. Acta Histochem. Cytochem. 2009, 42, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Vangaveti, V.N.; Shashidhar, V.M.; Rush, C.; Malabu, U.H.; Rasalam, R.R.; Collier, F.; Baune, B.T.; Kennedy, R.L. Hydroxyoctadecadienoic acids regulate apoptosis in human THP-1 cells in a PPARgamma-dependent manner. Lipids 2014, 49, 1181–1192. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, H.; Chan, L. The role of 15-lipoxygenase in atherogenesis: Pro- and antiatherogenic actions. Curr. Opin. Lipidol. 1997, 8, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, H.; Heydeck, D.; Hugou, I.; Gniwotta, C. In vivo action of 15-lipoxygenase in early stages of human atherogenesis. J. Clin. Investig. 1997, 99, 888–893. [Google Scholar] [CrossRef]
- Waddington, E.I.; Croft, K.D.; Sienuarine, K.; Latham, B.; Puddey, I.B. Fatty acid oxidation products in human atherosclerotic plaque: An analysis of clinical and histopathological correlates. Atherosclerosis 2003, 167, 111–120. [Google Scholar] [CrossRef]
- Szczuko, M.; Kotlega, D.; Palma, J.; Zembron-Lacny, A.; Tylutka, A.; Golab-Janowska, M.; Drozd, A. Lipoxins, RevD1 and 9, 13 HODE as the most important derivatives after an early incident of ischemic stroke. Sci. Rep. 2020, 10, 12849. [Google Scholar] [CrossRef]
- Kim-Campbell, N.; Gretchen, C.; Ritov, V.B.; Kochanek, P.M.; Balasubramani, G.K.; Kenny, E.; Sharma, M.; Viegas, M.; Callaway, C.; Kagan, V.E.; et al. Bioactive Oxylipins in Infants and Children with Congenital Heart Disease Undergoing Pediatric Cardiopulmonary Bypass. Pediatr. Crit. Care Med. 2020, 21, 33–41. [Google Scholar] [CrossRef]
- Shinto, L.; Lahna, D.; Murchison, C.F.; Dodge, H.; Hagen, K.; David, J.; Kaye, J.; Quinn, J.F.; Wall, R.; Silbert, L.C. Oxidized Products of Omega-6 and Omega-3 Long Chain Fatty Acids Are Associated with Increased White Matter Hyperintensity and Poorer Executive Function Performance in a Cohort of Cognitively Normal Hypertensive Older Adults. J. Alzheimer’s Dis. 2020, 74, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Borsini, A.; Nicolaou, A.; Camacho-Munoz, D.; Kendall, A.C.; Di Benedetto, M.G.; Giacobbe, J.; Su, K.-P.; Pariante, C.M. Omega-3 polyunsaturated fatty acids protect against inflammation through production of LOX and CYP450 lipid mediators: Relevance for major depression and for human hippocampal neurogenesis. Mol. Psychiatry 2021, 26, 6773–6788. [Google Scholar] [CrossRef] [PubMed]
- Endo, J.; Sano, M.; Isobe, Y.; Fukuda, K.; Kang, J.X.; Arai, H.; Arita, M. 18-HEPE, an n-3 fatty acid metabolite released by macrophages, prevents pressure overload-induced maladaptive cardiac remodeling. J. Exp. Med. 2014, 211, 1673–1687. [Google Scholar] [CrossRef]
- Welty, F.K.; Schulte, F.; Alfaddagh, A.; Elajami, T.K.; Bistrian, B.R.; Hardt, M. Regression of human coronary artery plaque is associated with a high ratio of (18-hydroxy-eicosapentaenoic acid + resolvin E1) to leukotriene B4. Fed. Am. Soc. Exp. Biol. 2021, 35, e21448. [Google Scholar] [CrossRef] [PubMed]
- Takenaga, M.; Hirai, A.; Terano, T.; Tamura, Y.; Kitagawa, H.; Yoshida, S. Comparison of the in vitro effect of eicosapentaenoic acid (EPA)-derived lipoxygenase metabolites on human platelet function with those of arachidonic acid. Thromb. Res. 1986, 41, 373–384. [Google Scholar] [CrossRef]
- Atone, J.; Wagner, K.; Hashimoto, K.; Hammock, B.D. Cytochrome P450 derived epoxidized fatty acids as a therapeutic tool against neuroinflammatory diseases. Prostaglandins Other Lipid Mediat. 2020, 147, 106385. [Google Scholar] [CrossRef]
- Sun, C.P.; Zhang, X.Y.; Morisseau, C.; Hwang, S.H.; Zhang, Z.J.; Hammock, B.D.; Ma, X.-C. Discovery of Soluble Epoxide Hydrolase Inhibitors from Chemical Synthesis and Natural Products. J. Med. Chem. 2021, 64, 184–215. [Google Scholar] [CrossRef]
- Caligiuri, S.P.; Aukema, H.M.; Ravandi, A.; Guzman, R.; Dibrov, E.; Pierce, G.N. Flaxseed consumption reduces blood pressure in patients with hypertension by altering circulating oxylipins via an alpha-linolenic acid-induced inhibition of soluble epoxide hydrolase. Hypertension 2014, 64, 53–59. [Google Scholar] [CrossRef]
- Caligiuri, S.P.; Rodriguez-Leyva, D.; Aukema, H.M.; Ravandi, A.; Weighell, W.; Guzman, R.; Pierce, G.N. Dietary Flaxseed Reduces Central Aortic Blood Pressure without Cardiac Involvement but Through Changes in Plasma Oxylipins. Hypertension 2016, 68, 1031–1038. [Google Scholar] [CrossRef]
- Zhang, Y.; Guallar, E.; Blasco-Colmenares, E.; Harms, A.C.; Vreeken, R.J.; Hankemeier, T.; Tomaselli, G.F.; Cheng, A. Serum-Based Oxylipins Are Associated with Outcomes in Primary Prevention Implantable Cardioverter Defibrillator Patients. PLoS ONE 2016, 11, e0157035. [Google Scholar] [CrossRef]
- Yu, D.; Hennebelle, M.; Sahlas, D.J.; Ramirez, J.; Gao, F.; Masellis, M.; Cogo-Moreira, H.; Swartz, R.H.; Herrmann, N.; Chan, P.C.; et al. Soluble Epoxide Hydrolase-Derived Linoleic Acid Oxylipins in Serum Are Associated with Periventricular White Matter Hyperintensities and Vascular Cognitive Impairment. Transl. Stroke Res. 2019, 10, 522–533. [Google Scholar] [CrossRef] [PubMed]
- Silbert, L.C.; Wall, R.; Lahna, D.; Dodge, H.; Hagen, K.; David, J.; Kaye, J.; Shinto, L. Docosahexaenoic acid oxylipins are associated with white matter microstructural integrity in a cohort of cognitively normal, hypertensive, young-old. Alzheimer’s Dement. 2020, 16 (Suppl. 4), e044965. [Google Scholar] [CrossRef]
- Zhao, T.; Wang, Y.; Guo, X.; Li, H.; Jiang, W.; Xiao, Y.; Deng, B.; Sun, Y. Altered oxylipin levels in human vitreous indicate imbalance in pro-/anti-inflammatory homeostasis in proliferative diabetic retinopathy. Exp. Eye Res. 2022, 214, 108799. [Google Scholar] [CrossRef] [PubMed]
- Borkowski, K.; Taha, A.Y.; Pedersen, T.L.; De Jager, P.L.; Bennett, D.A.; Arnold, M.; Kaddurah-Daouk, R.; Newman, J.W. Serum metabolomic biomarkers of perceptual speed in cognitively normal and mildly impaired subjects with fasting state stratification. Sci. Rep. 2021, 11, 18964. [Google Scholar] [CrossRef] [PubMed]
- Borkowski, K.; Pedersen, T.L.; Seyfried, N.T.; Lah, J.J.; Levey, A.I.; Hales, C.M.; Dammer, E.B.; Blach, C.; Louie, G.; Kaddurah-Daouk, R.; et al. Association of plasma and CSF cytochrome P450, soluble epoxide hydrolase, and ethanolamide metabolism with Alzheimer’s disease. Alzheimer’s Res. Ther. 2021, 13, 149. [Google Scholar] [CrossRef]
- DeCarli, C.; Murphy, D.G.; Tranh, M.; Grady, C.L.; Haxby, J.V.; Gillette, J.A.; Salerno, J.A.; Gonzales-Aviles, A.; Honvitz, B.; Rapoport, S.I.; et al. The effect of white matter hyperintensity volume on brain structure, cognitive performance, and cerebral metabolism of glucose in 51 healthy adults. Neurology 1995, 45, 2077–2084. [Google Scholar] [CrossRef]
- Silbert, L.C.; Dodge, H.H.; Perkins, L.G.; Sherbakov, L.; Lahna, D.; Erten-Lyons, D.; Woltjer, R.; Shinto, L.; Kaye, J.A. Trajectory of white matter hyperintensity burden preceding mild cognitive impairment. Neurology 2012, 79, 741–747. [Google Scholar] [CrossRef]
- Silbert, L.C.; Nelson, C.; Howieson, D.B.; Moore, M.M.; Kaye, J.A. Impact of white matter hyperintensity volume progression on rate of cognitive and motor decline. Neurology 2008, 71, 108–113. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Population | Methods | Findings | Conclusions |
|---|---|---|---|---|
| Kuhn et al. (1997) [108] | Advanced atheroma from patients undergoing carotid endarterectomy, Berlin samples (N = 17), age 50–70 years Young human lesions obtained from PDAY study samples (N = 19) (Wissler 1994), age 15–34 years | Advanced atheroma samples, carotid artery Post-mortem atherosclerotic samples |
| LOX activity is enzymatically active in young human lesions; may be import for early atherogenesis |
| Waddington et al. (2003) [109] | Patients undergoing carotid endarterectomy, age 70 years, Royal Perth Hospital, Australia Recent TIA or stroke (N = 29) Asymptomatic stenosis (N = 17) | Atherosclerotic plaques, carotid artery Plaque severity, pathological classification |
| Did not identify FA oxidation products associated with plaque instability and symptomatic CVD |
| Shibata et al. (2009) [105] | Patients undergoing carotid endarterectomy (N = 6) | Atherosclerotic plaques, carotid artery |
| 13(R)-HODE, non-enzymatically formed in the presence of oxidative stress, is a major component of atherosclerotic lesions |
| Caligiuri et al. (2014) [119] | FlaxPAD participants (Leyva 2011) Original cohort (N = 110), age 66.4 years N = 98 at baseline with oxylipin data N = 76 at 6 months with oxylipin data Flaxseed-enriched vs. control diet | Plasma Systolic BP Diastolic BP |
| Flaxseed LA may inhibit sEH activity, resulting in altered oxylipin concentrations that contribute to blood pressure reduction in patients with PAD |
| Caligiuri et al. (2016) [120] | FlaxPAD participants (Leyva 2011) Original cohort (N = 110), age 66.4 years N = 62, baseline and 6 months N = 41/62 with HTN diagnosis | Plasma Radial tonometry (cBP) |
| Study provides support for oxylipins as therapeutic targets in HTN |
| Zhang et al. (2016) [121] | PROSE-ICD study (N = 479), age 60, patients with heart failure and implanted defibrillator | serum |
| Novel oxylipin markers identified associated with ventricular fibrillation and all-cause mortality in patients with heart failure |
| Hu et al. (2017) [59] | Wilmer Eye Institute, Johns Hopkins N = 6, non-DR, age 75.7 ± 3.1 years N = 7, mild DR, age 57.2 ± 13.0 years N = 6, severe DR, age 62.2 ± 11.8 years | Post-mortem retinal samples |
| DR severity associated with increased sEH expression |
| Hu et al. 2017 [59] | Henan Eye Institute, Henan, China N = 17 DR, age 57.0 ± 9.9 years N = 14 non-DM macular disease, age 58.9 ± 8.0 years | Vitreous humor samples from patients undergoing vitrectomy |
| DR associated with increased DHA sEH-derived oxylipin |
| Yu et al. (2019) [122] | Patients with recent TIA and high WMH (N = 29), age 71.8 years Healthy elderly controls with low WMH (N = 25), age 71.7 years MMSE > 19, no cortical stroke or Alzheimer’s disease diagnosis | Serum 3T MRI Psychometric testing |
| Oxylipin changes associated with higher sEH activity may be use as markers for age-related VCI |
| Szczuko et al. (2020) [110] | Patient with ischemic stroke (N = 75), District Hospital, Poland Control Group (N = 35), University of the Third Age, Poland | Fasting plasma |
| Study confirms the involvement of FFA-derivative mediators of inflammation associated with ischemic stroke |
| Shinto et al. (2020) [112] | Community-based sample, Portland, OR N = 37, controlled HTN, age 65.6 ± 7.1 years, no dementia | Fasting plasma 3T MRI |
| Specific oxylipin products, and not parent compounds LA or DHA, are associated with MRI and cognitive markers of dementia risk, including that from VCI, in young–old individuals |
| Silbert et al. (2020) [123] | Community-based sample, Portland, OR N = 37, controlled HTN, age 65.6 ± 7.1 years, no dementia (N = 36 with DTI) | Fasting plasma 3T MRI TBSS DTI |
| DHA-derived oxylipins, not DHA are associated with WM integrity in hypertensive young-old individuals at risk for VCI |
| Kim-Campbell et al. (2020) [111] | Children undergoing CPB surgery at the Children’s Hospital of Pittsburg N = 34, age 2.5 years (0.6–12.0) | Blood collected start and end of CPB PHb, plasma oxylipins, VIS |
| 9:13-HODE ratio is a relevant biomarker of vascular tension and inflammation in a pediatric population undergoing CPB surgery |
| Welty et al. (2021) [115] | Subset of participants in HEARTS trial; EPA + DHA treatment in patients with CAD (Alfaddagh 2017), BIDMC, Boston, MA 31 subjects with the highest (N = 16) and lowest plasma omega-3 fatty acid index EPA + DHA/total fatty acids (N = 15) at 30-month follow-up. mean age in both groups, around 63 years | Plasma omega-3 fatty acid index (EPA + DHA/total fatty acid level), and oxylipins Coronary plaque volume measured using CCTA |
| A low ratio of specialized pro-resolving mediators to proinflammatory mediator, specifically (18-HEPE + RvE1)/LTB4, is a potential novel risk factor associated with coronary plaque progression in patients with CAD |
| Zhao et al. (2022) [124] | Department of Ophthalmology, Second Xiangya Hospital of Central South University Vitreous humor samples from patients with PDR (N = 41) and non-diabetic controls (N = 22) | Vitreous humor samples obtained during vitrectomy |
| LOX- and CYP-products were the most affected oxylipins, altered balance between inflammatory and anti-inflammatory oxylipins in PDR |
| Borkowski et al. (2021) [125] | Participants from the RUSH ROS and MAP (N = 198). Age 78.2 (7.2), 22% male, 95% white, NCI and MCI. 212 serum samples: fasted (N = 59), non-fasted (N = 80), unknown if fasted (N = 73); | Fasted and non-fasted oxylipins, endocannabinoids, bile acids and steroids Cognitive battery |
| Omega-3 sEH (diols) are predictive of poorer perceptual speed performance in samples obtained in fasting state; developed tool to estimate is sample is fasted, if unknown |
| Borkowski et al. (2021) [126] | Emory Healthy Brain Study (Emory ADRC) and Emory Cognitive Neurology Clinic CSF: AD, N = 150, age 68.2 (1.3), 47% male, 70% ApoE4, control, N = 139, age 65 (1.4), 28% male, 25% ApoE4 Plasma: AD, N = 148, age 68 (1.4), 48% male, 70% ApoE4, control, N = 133 age 66 (1.5), 27% male, 24% ApoE4 | Oxylipins, endocannabinoids, bile acids, steroids. Estimated fasted samples [125] MoCA |
| AD is associated with elevations in fasting plasma and CSF CYP sEH activity, supporting vascular dysfunction as a factor in AD |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shinto, L.H.; Raber, J.; Mishra, A.; Roese, N.; Silbert, L.C. A Review of Oxylipins in Alzheimer’s Disease and Related Dementias (ADRD): Potential Therapeutic Targets for the Modulation of Vascular Tone and Inflammation. Metabolites 2022, 12, 826. https://doi.org/10.3390/metabo12090826
Shinto LH, Raber J, Mishra A, Roese N, Silbert LC. A Review of Oxylipins in Alzheimer’s Disease and Related Dementias (ADRD): Potential Therapeutic Targets for the Modulation of Vascular Tone and Inflammation. Metabolites. 2022; 12(9):826. https://doi.org/10.3390/metabo12090826
Chicago/Turabian StyleShinto, Lynne H., Jacob Raber, Anusha Mishra, Natalie Roese, and Lisa C. Silbert. 2022. "A Review of Oxylipins in Alzheimer’s Disease and Related Dementias (ADRD): Potential Therapeutic Targets for the Modulation of Vascular Tone and Inflammation" Metabolites 12, no. 9: 826. https://doi.org/10.3390/metabo12090826
APA StyleShinto, L. H., Raber, J., Mishra, A., Roese, N., & Silbert, L. C. (2022). A Review of Oxylipins in Alzheimer’s Disease and Related Dementias (ADRD): Potential Therapeutic Targets for the Modulation of Vascular Tone and Inflammation. Metabolites, 12(9), 826. https://doi.org/10.3390/metabo12090826