
Nuclear Receptor Pathways Mediating the Development of Boar Taint


Abstract
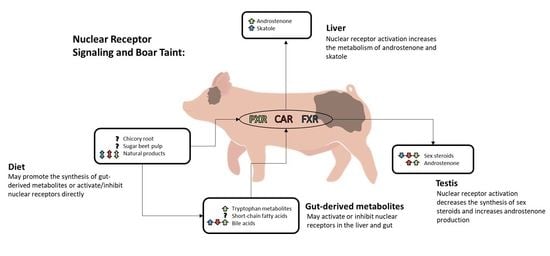
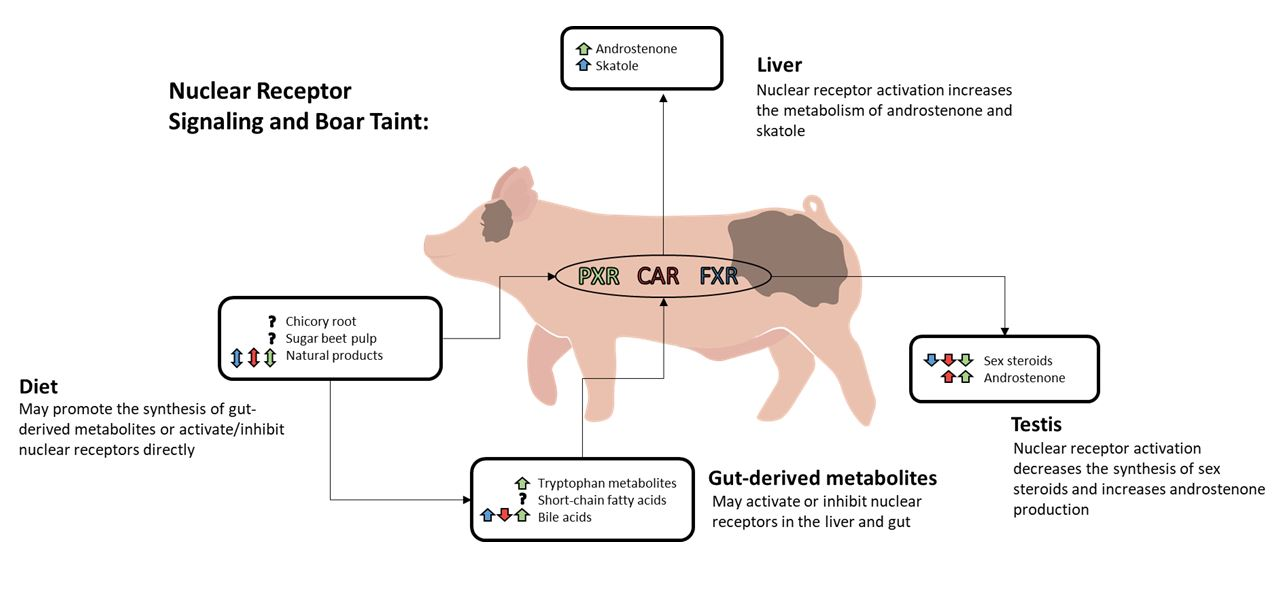
:
{kind=link}
{kind=link}
1. Introduction
2. Nuclear Receptor Signaling and Boar Taint
2.1. Nuclear Receptor-Mediated Regulation of Testicular Steroid Synthesis
2.2. Nuclear Receptor-Mediated Regulation of Boar Taint Metabolism
2.2.1. Nuclear Receptor-Mediated Effects on Skatole Metabolism
2.2.2. Nuclear Receptor-Mediated Effects on Androstenone Metabolism
2.3. Coregulatory Proteins and Nuclear Receptor Crosstalk
3. The Gut–Liver Axis
3.1. Gut-Derived Tryptophan Metabolites
3.2. Short-Chain Fatty Acids
3.3. Bile Acids
3.4. Diet
4. Recommendations for Future Research
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mangelsdorf, D.J.; Thummel, C.; Beato, M.; Herrlich, P.; Schutz, G.; Umesono, K.; Blumberg, B.; Kastner, P.; Mark, M.; Chambon, P.; et al. The Nuclear Receptor Superfamily: The Second Decade. Cell 1995, 83, 835–839. [Google Scholar] [CrossRef]
- Sever, R.; Glass, C.K. Signaling by Nuclear Receptors. Cold Spring Harb. Perspect. Biol. 2013, 5, a016709. [Google Scholar] [CrossRef] [PubMed]
- Bain, D.L.; Heneghan, A.F.; Connaghan-Jones, K.D.; Miura, M.T. Nuclear Receptor Structure: Implications for Function. Annu. Rev. Physiol. 2007, 69, 201–220. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, Y. Natural and Artificial Zinc Finger Proteins. RIKEN Rev. 2001, 35, 102–104. [Google Scholar]
- Xu, J.; Li, Q. Review of the In Vivo Functions of the P160 Steroid Receptor Coactivator Family. Mol. Endocrinol. 2003, 17, 1681–1692. [Google Scholar] [CrossRef] [PubMed]
- Kraus, W.L.; McInerney, E.M.; Katzenellenbogen, B.S. Ligand-Dependent, Transcriptionally Productive Association of the Amino-and Carboxy-Terminal Regions of a Steroid Hormone Nuclear Receptor. Proc. Natl. Acad. Sci. USA. 1995, 92, 12314–12318. [Google Scholar] [CrossRef] [PubMed]
- Tasset, D.; Tora, L.; Fromental, C.; Scheer, E.; Chambon, P. Distinct Classes of Transcriptional Activating Domains Function by Different Mechanisms. Cell 1990, 62, 1177–1187. [Google Scholar] [CrossRef]
- Benecke, A.; Chambon, P.; Gronemeyer, H. Synergy Between Estrogen Receptor an Activation Functions AF1 and AF2 Mediated by Transcription Intermediary Factor TIF2. EMBO Rep. 2000, 1, 151–157. [Google Scholar] [CrossRef]
- Frigo, D.E.; Bondesson, M.; Williams, C. Nuclear Receptors: From Molecular Mechanisms to Therapeutics. Essays Biochem. 2021, 65, 847–856. [Google Scholar] [CrossRef]
- Wang, Y.D.; Chen, W.D.; Moore, D.D.; Wendong, H. FXR: A Metabolic Regulator and Cell Protector. Cell Res. 2008, 18, 1087–1095. [Google Scholar] [CrossRef]
- Kliewer, S.A.; Moore, J.T.; Wade, L.; Staudinger, J.; Watson, M.A.; Jones, S.A.; McKee, D.D.; Oliver, B.B.; Wilson, T.M.; Zetterström, R.H.; et al. An Orphan Nuclear Receptor Activated by Pregnanes Defines a Novel Steroid Signaling Pathway. Cell 1998, 9, 73–82. [Google Scholar] [CrossRef] [Green Version]
- di Masi, A.; De Marnis, E.; Ascenzi, P.; Marino, M. Nuclear Receptors CAR and PXR: Molecular, Functional and Biomedical Aspects. Mol. Asp. Med. 2009, 30, 297–343. [Google Scholar] [CrossRef] [PubMed]
- Smutny, T.; Mani, S.; Pavek, P. Post-Translational and Post-Transcriptional Modifications of Pregnane X Receptor (PXR) in Regulation of the Cytochrome P450 Superfamily. Curr. Drug Metab. 2013, 14, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, M.; Mukherjee, S.; Wang, H.; Li, H.; Sun, K.; Benechet, A.P.; Qiu, Z.; Maher, L.; Redinbo, M.R.; Phillips, R.S.; et al. Symbiotic Bacterial Metabolites Regulate Gastrointestinal Barrier Function Via the Xenobiotic Sensor PXR and Toll-Like Receptor 4. Immunity 2014, 41, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Motta, S.; Callea, L.; Tagliabue, S.G.; Bonati, L. Exploring the PXR Ligand Binding Mechanism with Advanced Molecular Dynamics Methods. Sci. Rep. 2018, 8, 16207. [Google Scholar] [CrossRef]
- Forman, B.M.; Tzameli, I.; Choi, H.; Chen, J.; Simha, D.; Seol, W.; Evans, R.M.; Moore, D.D. Androstane Metabolites Bind to And Deactivate the Nuclear Receptor CAR-β. Nature 1998, 395, 612–615. [Google Scholar] [CrossRef]
- Sueyoshi, T.; Kawamoto, T.; Zelko, I.; Honkakoski, P.; Negishi, M. The Repressed Nuclear Receptor CAR Responds to Phenobarbital in Activating the Human CYP2B6 gene. J. Biol. Chem. 1999, 274, 6043–6046. [Google Scholar] [CrossRef]
- Moore, L.B.; Parks, D.J.; Jones, S.A.; Bledsoe, R.K.; Consler, T.G.; Stimmel, J.B.; Goodwin, B.; Liddle, C.; Blanchard, S.G.; Wilson, T.M.; et al. Orphan Nuclear Receptors Constitutive Androstane Receptor and Pregnane X Receptor Share Xenobiotic and Steroid Ligands. J. Biol. Chem. 2000, 275, 15122–15127. [Google Scholar] [CrossRef]
- Tzamelia, I.; Pissios, P.; Schuetz, E.G.; Moore, D.D. The Xenobiotic Compound 1,4-Bis[2-(3,5-Dichloropyridyloxy)]Benzene Is an Agonist Ligand for the Nuclear Receptor CAR. Mol. Cell. Biol. 2000, 20, 2951–2958. [Google Scholar] [CrossRef]
- Fisher, C.D.; Augustine, L.M.; Maher, J.M.; Nelson, D.M.; Slitt, A.L.; Klaassen, C.D.; Lehman-Mckeeman, L.D.; Cherrington, N.J. Induction of Drug-Metabolizing Enzymes by Garlic and Allyl Sulfide Compounds Via Activation of Constitutive Androstane Receptor and Nuclear Factor E2-Related Factor 2. Drug Metab. Dispos. 2007, 35, 995–1000. [Google Scholar] [CrossRef]
- Shizu, R.; Min, J.; Sobhany, M.; Pedersen, L.C.; Mutoh, S.; Negishi, M. Interaction of the Phosphorylated DNA-Binding Domain in Nuclear Receptor CAR With Its Ligand-Binding Domain Regulates CAR Activation. J. Biol. Chem. 2018, 293, 333–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, B.; Arons, A.B.; Vincent, J.G.; Fernandez, E.J.; Shen, T. Regulatory Mechanics of Constitutive Androstane Receptors: Basal and Ligand-Directed Actions. J. Chem. Inf. Model. 2019, 59, 5174–5182. [Google Scholar] [CrossRef] [PubMed]
- Wei, P.; Zhang, J.; Egan-Hafley, M.; Liand, S.; Moore, D. The Nuclear Receptor CAR Mediates Specific Xenobiotic Induction of Drug Metabolism. Nature 2000, 407, 920–923. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.K.H.; Waxman, D.J. Synthetic Drugs and Natural Products as Modulators of Constitutive Androstane Receptor (Car) And Pregnane X Receptor (PXR). Drug Metab. Rev. 2006, 38, 51–73. [Google Scholar] [CrossRef]
- Forman, B.M.; Goode, E.; Chen, J.; Oro, A.E.; Bradley, D.J.; Perlman, T.; Noonan, D.J.; Burka, L.T.; McMoris, T.; Lamph, W.W.; et al. Identification of a Nuclear Receptor That Is Activated by Farnesol Metabolites. Cell 1995, 81, 687–693. [Google Scholar] [CrossRef]
- Wang, H.; Chen, J.; Hollister, K.; Sowers, L.C.; Forman, B.M. Endogenous Bile Acids Are Ligands for the Nuclear Receptor FXR/BAR. Mol. Cell 1999, 3, 543–553. [Google Scholar] [CrossRef]
- Makishima, M.; Okamoto, A.Y.; Repa, J.J.; Tu, H.; Learned, R.M.; Luk, A.; Hull, M.V.; Lustig, K.D.; Mangelsdorf, D.J.; Shan, B. Identification of a Nuclear Receptor for Bile Acids. Science 1999, 21, 1362–1365. [Google Scholar] [CrossRef]
- Parks, D.J.; Blanchard, S.G.; Bledsoe, R.K.; Chandra, G.; Consler, T.G.; Kliewer, S.A.; Stimmel, J.B.; Willson, T.M.; Zavacki, A.M.; Moore, D.D.; et al. Bile acids: Natural Ligands for an Orphan Nuclear Receptor. Science 1999, 284, 1365–1368. [Google Scholar] [CrossRef]
- Stofan, M.; Guo, G.L. Bile Acids and FXR: Novel Targets for Liver Diseases. Front. Med. 2020, 7, 544. [Google Scholar] [CrossRef]
- Eloranta, J.J.; Kullak-Ublick, G.A. The Role of FXR In Disorders of Bile Acid Homeostasis. Physiology 2008, 23, 286–295. [Google Scholar] [CrossRef]
- Martinot, E.; Sédes, L.; Baptissart, M.; Holota, H.; Rouaisnel, B.; Damon-Soubeyrand, C.; De Haze, A.; Saru, J.; Thibault-Carpentier, C.; Keime, C.; et al. The Bile Acid Nuclear Receptor Fxrα Is a Critical Regulator of Mouse Germ Cell Fate. Stem Cell Rep. 2017, 9, 315–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, D.; Mangelsdorf, D.J.; Meyer, U.A. Pregnane X Receptor Is a Target of the Farnesoid XD Receptor. J. Biol. Chem. 2006, 281, 19081–19091. [Google Scholar] [CrossRef] [PubMed]
- Modica, S.; Bellafante, E.; Moschetta, A. Master Regulation of Bile Acid and Xenobiotic Metabolism Via the FXR, PXR and CAR Trio. Front. Biosci. 2009, 14, 4719–4745. [Google Scholar] [CrossRef]
- Gray, M.A.; Pollock, C.B.; Shook, L.B.; Squires, E.J. Characterization of Porcine Pregnane X Receptor, Farnesoid X Receptor and Their Splice Variants. Exp. Biol. Med. 2010, 235, 718–736. [Google Scholar] [CrossRef]
- Pollock, C.B.; Rogatcheva, M.B.; Schook, L.B. Comparative Genomics of Xenobiotic Metabolism: A Porcine-Human PXR Gene Comparison. Mamm. Genome 2007, 18, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.A.; Peacock, J.N.; Squires, E.J. Characterization of the Porcine Constitutive Androstane Receptor (CAR) and Its Splice Variants. Xenobiotica 2009, 39, 915–930. [Google Scholar] [CrossRef]
- Gray, M.A.; Squires, E.J. Effects of Nuclear Receptor Transactivation on Steroid Hormone Synthesis and Gene Expression in Porcine Leydig Cells. J. Steroid Biochem. Mol. Biol. 2013, 133, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.A.; Squires, E.J. Effects of Nuclear Receptor Transactivation on Boar Taint Metabolism and Gene Expression in Porcine Hepatocytes. J. Steroid Biochem. Mol. Biol. 2013, 133, 110–119. [Google Scholar] [CrossRef]
- Gunn, M.G.; Allen, P.; Bonneau, M.; Byrne, D.V.; Cinotti, S.; Fredriksen, B.; Hansen, L.L.; Karlsson, A.H.; Linder, M.G.; Lundström, K.; et al. Welfare Aspects of the Castration of Piglets. Scientific Report on the Scientific Panel for Animal Health and Welfare on a Request from the Commission Related to Welfare Aspects of the Castration of Piglets. EFSA J. 2004, 91, 1–18. [Google Scholar]
- Gower, D.B. 16-Unsaturated C19 Steroids A Review of Their Chemistry, Biochemistry and Possible Physiological Role. J. Steroid Biochem. 1972, 3, 45–103. [Google Scholar] [CrossRef]
- Meadus, W.J.; Mason, J.I.; Squires, E.J. Cytochrome P450c17 from the Porcine and Bovine Adrenal Catalyses the Formation of 5,16-Androstadien-3β-Ol from Pregnenolone in the Presence of Cytochrome b5. J. Steroid. Biochem. Molec. Biol. 1993, 46, 565–572. [Google Scholar] [CrossRef]
- Nakajin, S.; Takahashi, M.; Shinoda, M.; Hall, P.F. Cytochrome B5 Promotes the Synthesis 16-C19 Steroids by Homogenous Cytochrome P-450 C21 Side-Chain Cleavage from Pig Testis. Biochem. Biophys. Res. Commun. 1985, 132, 708–713. [Google Scholar] [CrossRef]
- Billen, M.J.; Squires, E.J. The Role of Porcine Cytochrome B5a And Cytochrome B5b In the Regulation of Cytochrome P45017A1 activities. J. Steroid. Biochem. Molec. Biol. 2009, 113, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.F. 57 Varieties: The Human Cytochromes P450. Pharmacogenomics 2004, 5, 305–318. [Google Scholar] [CrossRef] [PubMed]
- Wiercinska, P.; Lou, Y.; Squires, E.J. The Roles of Different Porcine Cytochrome P450 Enzymes and Cytochrome B5a In Skatole Metabolism. Animal 2012, 6, 834–845. [Google Scholar] [CrossRef]
- Schelstraete, W.; De Clerck, L.; Govaert, L.; Millercam, J.; Devreese, M.; Deforce, D.; Van Bocxlaer, J.; Croubels, S. Characterization of Porcine Hepatic and Intestinal Drug Metabolizing CYP450: Comparison with Human Orthologues from A Quantitative, Activity and Selectivity Perspective. Nature 2019, 9, 9233. [Google Scholar] [CrossRef] [PubMed]
- Babol, J.; Squires, E.J.; Lundström, K. Relationship Between Oxidation and Conjugation Metabolism of Skatole in Pig Liver and Concentrations of Skatole in Fat. J. Anim. Sci. 1998, 76, 829–838. [Google Scholar] [CrossRef]
- Diaz, G.J.; Squires, E.J. Metabolism of 3-Methylindole by Porcine Microsomes: Responsible Cytochrome P450 Enzymes. Toxicol. Sci. 2000, 55, 284–292. [Google Scholar] [CrossRef]
- Terner, M.A.; Gilmore, W.J.; Lou, Y.; Squires, E.J. The Role of CYP2A And CYP2E1 in the Metabolism of 3-Methylindole in Primary Cultured Porcine Hepatocytes. Drug Metab. Dispos. 2006, 34, 848–854. [Google Scholar] [CrossRef]
- Diaz, G.J.; Squires, E.J. Phase II In Vitro Metabolism of 3-Methylindole Metabolites in Porcine Liver. Xenobiotica 2003, 33, 485–498. [Google Scholar] [CrossRef]
- Brunius, C.; Vidanarachchi, J.K.; Tomankova, J.; Lundström, K.; Andersson, K.; Zamaratskaia, G. Skatole Metabolites in Urine as a Biological Marker of Pigs with Enhanced Hepatic Metabolism. Animal 2016, 10, 1734–1740. [Google Scholar] [CrossRef] [PubMed]
- Diaz, J.G.; Skordos, K.W.; Yost, G.S.; Squires, E.J. Identification of Phase I metabolites of 3-methylindole produced by pig liver microsomes. Drug Metab. Dispos. 1999, 27, 1150–1156. [Google Scholar] [CrossRef] [PubMed]
- Bæk, C.; Hansen-Møller, J.; Friis, C.; Cornett, C.; Hansen, S.H. Identification of Selected Metabolites of Skatole in Plasma and Urine from Pigs. J. Agric. Food Chem. 1997, 45, 2332–2340. [Google Scholar] [CrossRef]
- Achour, B.; Barber, J.; Rostami-Hodjegan, A. Cytochrome P450 Pig Liver Pie: Determination of Individual Cytochrome P450 Isoform Contents in Microsomes from Two Pig Livers Using Liquid Chromatography in Conjunction with Mass Spectrometry. Drug Metab. Dispos. 2011, 39, 2130–2134. [Google Scholar] [CrossRef]
- Moe, M.; Lien, S.; Bendixen, C.; Hedegaard, J.; Hornshøj, H.; Berget, I.; Meuwissen, T.H.E.; Grindflek, E. Gene Expression Profiles in Liver of Pigs with Extreme High and Low Levels of Androstenone. BMC Vet. Res. 2008, 4, 29. [Google Scholar] [CrossRef]
- Doran, E.; Whittington, F.W.; Wood, J.D.; McGivan, J.D. Cytochrome P450IIE1 (CYP2E1) is Induced by Skatole, and This Induction is bBocked by Androstenone in Isolated Pig Hepatocytes. Chem. Biol. Interact. 2002, 140, 81–92. [Google Scholar] [CrossRef]
- Zamaratskaia, G.; Gilmore, W.J.; Lundström, K.; Squires, E.J. Effect of Testicular Steroids on Catalytic Activities of Cytochrome P450 Enzymes in Porcine Liver Microsomes. Food Chem. Toxicol. 2007, 45, 676–681. [Google Scholar] [CrossRef]
- Puccinelli, E.; Gervasi, P.G.; La Marca, M.; Beffy, P.; Longo, V. Expression and Inducibility by Phenobarbital of CYP2C33, CYP2C42, CYP2C49, CYP2B22, And CYP3As in Porcine Liver, Kidney, Small Intestine, and Nasal Tissues. Xenobiotica 2010, 40, 525–535. [Google Scholar] [CrossRef]
- Laderoute, H.; Bone, C.; Squires, E.J. The Sulfoconjugation of Androstenone and Dehydroepiandrosterone by Human and Porcine Sulfotransferase Enzymes. Steroids 2018, 136, 8–16. [Google Scholar] [CrossRef]
- Sinclair, P.A.; Gilmore, W.J.; Lin, Z.; Lou, Y.; Squires, E.J. Molecular Cloning and Regulation of Porcine SULT2A1: Relationship Between SULT2A1 Expression and Sulfoconjugation of Androstenone. J. Mol. Endocrinol. 2006, 36, 301–311. [Google Scholar] [CrossRef]
- Bone, C.; Squires, E.J. The Uptake and Deconjugation of Androstenone Sulfate in the Adipose Tissue of the Boar. Animals 2021, 11, 3158. [Google Scholar] [CrossRef] [PubMed]
- Schuler, G.; Dezhkam, Y.; Tenbusch, L.; Klymiuk, M.C.; Zimmer, B.; Hoffmann, B. Formation and Hydrolysis of Sulfonated Estrogens in the Porcine Testis and Epididymis. J. Mol. Endocrinol. 2018, 61, M13–M25. [Google Scholar] [CrossRef] [PubMed]
- Lee, F.Y.; de Aguiar Vallim, T.Q.; Chong, H.K.; Zhang, Y.; Liu, Y.; Jones, S.A.; Osbourne, T.F.; Edwards, P.A. Activation of the Farnesoid X Receptor Provides Protection Against Acetaminophen-Induced Hepatic Toxicity. Mol. Endocrinol. 2010, 24, 1626–1636. [Google Scholar] [CrossRef]
- Dasgupta, S.; Lonard, D.M.; O’Malley, B.W. Nuclear Receptor Coactivators: Master Regulators of Human Health and Disease. Annu. Rev. Med. 2014, 65, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Xu, Y. Nuclear Receptor Coactivators (NCOAs) and Corepressors (NCORs) in the Brain. Endocrinology 2020, 161, 1–12. [Google Scholar] [CrossRef]
- Heery, D.M.; Kalkhoven, E.; Hoare, S.; Parker, M.G. A Signature Motif in Transcriptional Co-Activators Mediates Binding to Nuclear Receptors. Nature 1997, 387, 733–736. [Google Scholar] [CrossRef]
- Le Douarin, B.; Nielsen, L.A.; Garnier, J.; Ichinose, H.; Jeanmougin, F.; Losson, R.; Chambon, P.A. Possible Involvement of TIF1α and TIF1β In the Epigenetic Control of Transcription by Nuclear Receptors. EMBO J. 1996, 15, 6701–6715. [Google Scholar] [CrossRef]
- Dasgupta, S.; O’Malley, B.W. Transcriptional Coregulators: Emerging Roles of SRC Family of Coactivators in Disease Pathology. J. Mol. Endocrinol. 2014, 53, R47–R59. [Google Scholar] [CrossRef]
- Perissi, V.; Rosenfeld, M.G. Controlling Nuclear Receptors: The Circular Logic of Cofactor Cycles. Nat. Rev. Mol. Cell Biol. 2005, 6, 542–554. [Google Scholar] [CrossRef]
- Rosenfeld, M.G.; Lunyak, V.V.; Glass, C.K. Sensors and Signals: A Coactivator/Corepressor/Epigenetic Code for Integrating Signal-Dependent Programs of Transcriptional Response. Genes Dev. 2006, 20, 1405–1428. [Google Scholar] [CrossRef]
- Pavek, P. Pregnane X receptor (PXR)–Mediated Gene Repression and Cross-Talk of PXR with Other Nuclear Receptors Via Coactivator Interactions. Front. Pharmacol. 2016, 7, 456. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Lazar, M.A. The Cornr Motif Controls the Recruitment of Corepressors by Nuclear Hormone Receptors. Nature 1999, 402, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Nagy, L.; Kao, H.; Love, J.D.; Li, C.; Banayo, E.; Cooch, J.T.; Krishna, V.; Chatterjee, K.; Evans, R.M.; Schwabe, W.R. Mechanism of Corepressor Binding and Release from Nuclear Hormone Receptors. Genes Dev. 1999, 13, 3209–3216. [Google Scholar] [CrossRef] [PubMed]
- Goodson, M.; Jonas, B.A.; Privalsky, M.A. Corepressors: Custom Tailoring and Alterations While You Wait. Nucl. Recept. Signal. 2005, 3, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Baptissart, M.; Martinot, E.; Vega, A.; Sédes, L.; Rouaisnel, B.; de Haze, A.; Baron, S.; Schoonjas, K.; Caira, F.; Volle, D.H. Bile Acid-Fxrα Pathways Regulate Male Sexual Maturation in Mice. Oncotarget 2016, 7, 19468–19482. [Google Scholar] [CrossRef] [PubMed]
- Ourlin, J.C.; Lasserre, F.; Pineau, T.; Fabre, J.M.; Sa-Cunha, A.; Maurel, P.; Vilarem, M.J.; Pascussi, J.M. The Small Heterodimer Partner Interacts with the Pregnane X Receptor and Represses Its Transcriptional Activity. Mol. Endocrinol. 2003, 17, 1693–1703. [Google Scholar] [CrossRef]
- Li, T.; Chiang, J.Y.L. Rifampicin Induction of CYP3A4 Requires Pregnane X Receptor Cross Talk with Hepatocyte Nuclear Factor 4α and Coactivators, and Suppression of Small Heterodimer Partner Gene Expression. Drug Metab. Dispos. 2006, 34, 756–764. [Google Scholar] [CrossRef]
- Saini, S.P.S.; Mu, Y.; Gong, H.; Toma, D.; Uppal, H.; Ren, S.; Li, S.; Poloyac, S.M.; Xie, W. Dual Role of Orphan Nuclear Receptor Pregnane X Receptor in Bilirubin Detoxification in Mice. Hepatology 2005, 41, 497–505. [Google Scholar] [CrossRef]
- Pascussi, J.M.; Gerbal-Chaloin, S.; Duret, C.; Daujat-Chavanieu, M.; Vilarem, M.J.; Maurel, P. The Tangle of Nuclear Receptors That Controls Xenobiotic Metabolism and Transport: Crosstalk and Consequences. Annu. Rev. Pharmacol. Toxicol. 2008, 48, 1–32. [Google Scholar] [CrossRef]
- Masuyama, H.; Hiramatsu, Y.; Kunitomi, M.; Kudo, T.; MacDonald, P.N. Endocrine Disrupting Chemicals, Phthalic Acid and Nonylphenol, Activate Pregnane X Receptor-Mediated Transcription. Mol. Endocrinol. 2000, 14, 421–428. [Google Scholar] [CrossRef]
- Quigley, E.M.M. Gut Bacteria in Health and Disease. Gastroenterol. Hepatol. 2013, 9, 560–569. [Google Scholar]
- Dempsey, J.L.; Cui, J.Y. Microbiome is a Functional Modifier of P450 Drug Metabolism. Curr. Pharmacol. Rep. 2019, 5, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, A.G.; Sperandio, V. Quorum Sensing and the Gut Microbiome. In Quorum Sensing, 1st ed.; Tommonaro, G., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 151–169. [Google Scholar] [CrossRef]
- Li, X.; Zhang, B.; Hu, Y.; Zhao, Y. New Insights into Gut-Bacteria-Derived Indole and Its Derivatives in Intestinal and Liver Diseases. Front. Pharmacol. 2021, 12, 769501. [Google Scholar] [CrossRef] [PubMed]
- Albillos, A.; de Gottardi, A.; Rescigno, M. The Gut-Liver Axis in Liver Disease: Pathophysiological Basis for Therapy. J. Hepatol. 2020, 72, 558–577. [Google Scholar] [CrossRef]
- Nielsen, S.D.; Bauhaus, Y.; Zamaratskaia, G.; Junqueira, M.A.; Blaabjerg, K.; Petrat-Melin, B.; Young, J.F.; Rasmussen, M.K. Constitutive Expression and Activity of Cytochrome P450 In Conventional Pigs. Res. Vet. Sci. 2017, 111, 75–80. [Google Scholar] [CrossRef]
- Dodd, D.; Spitzer, M.H.; Van Treuren, W.; Merrill, B.D.; Hryckowian, A.J.; Higginbottom, S.K.; Le, A.; Cowan, T.M.; Nolan, G.P.; Fischbach, M.A.; et al. A Gut Bacterial Pathway Metabolizes Aromatic Amino Acids into Nine Circulating Metabolites. Nature 2017, 551, 648–652. [Google Scholar] [CrossRef]
- Elsden, S.R.; Hilton, M.G.; Waller, J.M. The End Products of the Metabolism of Aromatic Amino Acids by Clostridia. Arch. Microbiol. 1976, 107, 283–288. [Google Scholar] [CrossRef]
- Wlodarska, M.; Luo, C.; Kolde, R.; d’Hennezel, E.; Annand, J.W.; Heim, C.E.; Krastel, P.; Schmitt, E.K.; Omar, A.S.; Creasey, E.; et al. Indoleacrylic Acid Produced by Commensal Peptostreptococcus Species Suppresses Inflammation. Cell Host Microbe 2017, 22, 25–37. [Google Scholar] [CrossRef]
- Rasmussen, M.K.; Daujat-Chavanieu, M.; Gerbal-Chaloin, S. Activation of the Aryl Hydrocarbon Receptor Decreases-Rifampicin-Induced CYP3A4 Expression in Primary Human Hepatocytes and Heparg. Toxicol. Lett. 2017, 227, 1–8. [Google Scholar] [CrossRef]
- Sári, Z.; Mikó, E.; Kovács, T.; Jankó, L.; Csonka, T.; Lente, G.; Sebő, É.; Tóth, J.; Tóth, D.; Árkosy, P.; et al. Indolepropionic Acid, A Metabolite of the Microbiome, Has Cytostatic Properties in Breast Cancer by Activating AHR and PXR Receptors and Inducing Oxidative Stress. Cancers 2020, 12, 2411. [Google Scholar] [CrossRef]
- Morgan, E.T.; Dempsey, J.L.; Mimche, S.M.; Lamb, T.J.; Kulkarni, S.; Cui, J.Y.; Jeong, H.; Slitt, A.L. Physiological Regulation of Drug Metabolism and Transport: Pregnancy, Microbiome, Inflammation, Infection, and Fasting. Drug Metab. Depos. 2018, 46, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Illés, P.; Krasulová, K.; Vyhlídalová, B.; Poulíková, K.; Marcalíková, A.; Pečinková, P.; Sirotová, N.; Vrzal, R.; Mani, S.; Dvořák, Z. Indole Microbial Intestinal Metabolites Expand the Repertoire of Ligands and Agonists of The Human Pregnane X Receptor. Toxicol. Lett. 2020, 334, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Lee, J. Indole as an Intercellular Signal in Microbial Communities. FEMS Microbiol. Rev. 2010, 34, 426–444. [Google Scholar] [CrossRef]
- Tsavkelova, E.; Oeser, B.; Oren-Young, L.; Israeli, M.; Sasson, Y.; Tudzynski, B.; Sharon, A. Identification and Functional Characterization of Indole-3-Acetamide-Mediated IAA Biosynthesis in Plant-Associated Fusarium species. Fungal Genet. Biol. 2012, 49, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Jensen, B.B. Prevention of Boar Taint in Pig Production. Factors Affecting the Level of Skatole. Acta Vet. Scand. 2006, 48, S6. [Google Scholar] [CrossRef]
- Zelante, T.; Iannitti, R.G.; Cunha, C.; De Luca, A.; Giovannini, G.; Pieraccini, G.; Zecchi, R.; D’Angelo, C.; Massi-Benedetti, C.; Fallarino, F.; et al. Tryptophan Catabolites from Microbiota Engage Aryl Hydrocarbon Receptor and Balance Mucosal Reactivity Via Interlukin-22. Immunity 2013, 39, 372–385. [Google Scholar] [CrossRef]
- Whitehead, T.R.; Price, N.P.; Drake, H.L.; Cotta, M.A. Catabolic Pathway for the Production of Skatole and Indoleacetic Acid by the Acetogen Clostridium drakei, Clostridium scatologenes, and Swine Manure. Appl. Environ. Microbiol. 2008, 74, 1950–1953. [Google Scholar] [CrossRef]
- Li, X.; Jensen, R.L.; Højberg, O.; Canibe, N.; Jensen, B.B. Olsenella scatoligenes sp. Nov., a 3-methylindole- (skatole) and 4-methylphenol-(p-cresol) Producing Bacterium Isolated from Pig Faeces. Int. J. Syst. Evol. Microbiol. 2015, 65, 1227–1233. [Google Scholar] [CrossRef]
- Vyhlídalová, B.; Krasulová, K.; Pečinková, P.; Marcalíková, A.; Vrzal, R.; Zemánková, L.; Vančo, J.; Trávníček, Z.; Vondráček, J.; Karasová, M.; et al. Gut Microbial Catabolites of Tryptophan Are Ligands and Agonists of the Aryl Hydrocarbon Receptor: A Detailed Characterization. Int. J. Mol. Sci. 2020, 21, 2614. [Google Scholar] [CrossRef]
- Rasmussen, M.K.; Klausen, C.L.; Ekstrand, B. Regulation of Cytochrome P450 Mrna Expression in Primary Porcine Hepatocytes by Selected Secondary Plant Metabolites from Chicory (Cichorium intybus L.). Food Chem. 2014, 146, 255–263. [Google Scholar] [CrossRef]
- Monshouwer, M.; van’t Klooster, G.A.E.; Nijmeijer, S.M.; Witkamp, R.F.; van Miert, A.S.J.P.A.M. Characterization of Cytochrome P450 Isoenzymes in Primary Cultures of Pig Hepatocytes. Toxicol. Vitr. 1998, 12, 715–723. [Google Scholar] [CrossRef]
- Richards, L.B.; Li, M.; van Esch, B.C.A.M.; Garssen, J.; Folkerts, G. The Effects of Short-Chain Fatty Acids on the Cardiovascular System. PharmaNutrition 2016, 4, 68–111. [Google Scholar] [CrossRef]
- Venegas, D.P.; De la Fuente, M.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (Scfas)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed]
- den Besten, G.; Bleeker, A.; Gerding, A.; van Eunen, K.; Havinga, R.; van Dijk, T.H.; Oosterveer, M.H.; Jonker, J.W.; Groen, A.K.; Reijngoud, D.J.; et al. Short-Chain Fatty Acids Protect Against High-Fat Diet-Induced Obesity Via Pparγ-Dependent Switch from Lipogenesis to Fat Oxidation. Diabetes 2015, 64, 2398–2406. [Google Scholar] [CrossRef]
- Jung, T.H.; Park, J.H.; Jeon, W.M.; Han, K.S. Butyrate Modulates Bacterial Adherence on LS174T Human Colorectal Cells by Stimulating Mucin Secretion and MAPK Signaling Pathway. Nutr. Res. Pract. 2015, 9, 343–349. [Google Scholar] [CrossRef]
- Tan, J.; Mckenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The Role of Short-Chain Fatty Acids in Health and Disease. In Advances in Immunology, 1st ed.; Alt, F., Murphy, K., Eds.; Academic Press: Cambridge, MA, USA, 2014; Volume 121, pp. 91–119. [Google Scholar] [CrossRef]
- Jansen, M.S.; Nagel, S.C.; Miranda, P.J.; McDonnell, D.P. Short-Chain Fatty Acids Enhance Nuclear Receptor Activity Through Mitogen-Activated Protein Kinase Activation and Histone Deacetylase Inhibition. Proc. Natl. Acad. Sci. USA. 2004, 101, 7199–7204. [Google Scholar] [CrossRef] [PubMed]
- Jin, U.H.; Park, H.; Davidson, L.A.; Callaway, E.S.; Chapkin, R.S.; Jayaraman, A.; Asante, A.; Allred, C.; Weaver, E.A.; Safe, S. Short Chain Fatty Acids Enhance Aryl Hydrocarbon (Ah) Responsiveness in Mouse Colonocytes and Caco-2 Human Colon Cancer Cells. Nature 2017, 7, 10163. [Google Scholar] [CrossRef] [PubMed]
- Marinelli, L.; Martin-Gallausiaux, C.; Bourhis, J.M.; Béguet-Crespel, F.; Blottière, H.M.; Lapaque, N. Identification of the Novel Role of Butyrate as Ahr Ligand in Human Intestinal Epithelial Cells. Nature 2019, 9, 643. [Google Scholar] [CrossRef]
- Ranhotra, H.S.; Flannigan, K.L.; Brave, M.; Mukherjee, S.; Lukin, D.J.; Hirota, S.A.; Mani, S. Xenobiotic Receptor-Mediated Regulation of Intestinal Barrier Function and Innate Immunity. Nucl. Receptor Res. 2016, 3, 101199. [Google Scholar] [CrossRef]
- Ren, J.; Sun, K.; Wu, Z.; Yao, J.; Guo, B. All 4 Bile Salt Hydrolase Proteins Are Responsible for the Hydrolysis Activity in Lactobacillus plantarum ST-III. J. Food Sci. 2011, 76, M622–M628. [Google Scholar] [CrossRef]
- Elkins, C.A.; Moser, S.A.; Savage, D.C. Genes Encoding Bile Salt Hydrolases and Conjugated Bile Salt Transporters in Lactobacillus johnsonii 100-100 and other Lactobacillus species. Microbiology 2001, 147, 3403–3412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayashree, S.; Pooja, S.; Pushpanathan, M.; Rajendhran, J.; Gunasekaran, P. Identification and Characterization of Bile Salt Hydrolase Genes from the Genome of Lactobacillus fermentum MTCC 8711. Appl. Biochem. Biotechnol. 2014, 174, 855–866. [Google Scholar] [CrossRef] [PubMed]
- Franz, C.M.A.P.; Specht, I.; Haberer, P.; Holzapfel, W.H. Bile Salt Hydrolase Activity of Enterococci Isolated from Food: Screening and Quantitative Determination. J. Food Prot. 2001, 64, 725–729. [Google Scholar] [CrossRef]
- Tanaka, H.; Hashiba, H.; Kok, J.; Mierau, I. Bile Salt Hydrolase of Bifidobacterium Longum–Biochemical and Genetic Characterization. Appl. Environ. Microbiol. 2000, 66, 2502–2512. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.B.; Yi, S.H.; Lee, B.H. Purification and Characterization of Three Different Types of Bile Salt Hydrolases from Bifidobacterium Strains. J. Dairy Sci. 2004, 87, 258–266. [Google Scholar] [CrossRef]
- Rossocha, M.; Schultz-Heienbrok, R.; von Moeller, H.; Coleman, J.P.; Saenger, W. Conjugated Bile Acid Hydrolase Is a Tetrameric N-Terminal Thiol Hydrolase with Specific Recognition of Its Cholyl but Not of Its Tauryl Product. Biochemistry 2005, 44, 5739–5748. [Google Scholar] [CrossRef]
- Jones, B.V.; Begley, M.; Hill, C.; Gahan, C.G.M.; Marchesi, J.R. Functional and Comparative Metagenomics Analysis of Bile Salt Hydrolase Activity in the Human Gut Microbiome. Proc. Natl. Acad. Sci. USA 2008, 105, 13580–13585. [Google Scholar] [CrossRef]
- Reichen, J.; Paumgartner, G. Uptake of Bile Acids by Perfused Rat Liver. Am. J. Physiol. 1976, 231, 734–742. [Google Scholar] [CrossRef]
- Salvioli, G.; Lugli, R.; Pradelli, J.M.; Gigliotti, G. Bile Acid Binding in Plasma: The Importance of Lipoproteins. FEBS Lett. 1985, 187, 272–276. [Google Scholar] [CrossRef]
- Grüner, N.; Mattner, J. Bile Acids and Microbiota: Multifaceted and Versatile Regulators of the Liver-Gut Axis. Int. J. Mol. Sci. 2021, 22, 1397. [Google Scholar] [CrossRef]
- Ajouz, H.; Mukherji, D.; Shamseddine, A. Secondary Bile Acids: An Underrecognized Cause of Colon Cancer. J. Surg. Oncol. 2014, 12, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urdaneta, V.; Cassadesús, J. Interactions Between Bacteria and Bile Salts in the Gastrointestinal and Hepatobiliary Tracts. Front. Med. 2017, 4, 163. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Zhang, H.; Xiao, D.; Wei, H.; Chen, Y. Farnesoid X Receptor (FXR): Structures and Ligands. Comput. Struct. Biotechnol. J. 2021, 19, 2148–2159. [Google Scholar] [CrossRef] [PubMed]
- Staudinger, J.L.; Goodwin, B.; Jones, S.A.; Hawkins-Brown, D.; MacKenzie, K.I.; LaTour, A.; Liu, Y.; Klaassen, C.D.; Brown, K.K.; Reinhard, J.; et al. The Nuclear Receptor PXR Is a Lithocholic Acid Sensor That Protects Against Liver Toxicity. Proc. Natl. Acad. Sci. USA 2001, 98, 3369–3374. [Google Scholar] [CrossRef]
- Barnes, S.; Buchina, E.S.; King, R.J.; McBurnett, T.; Taylor, K.B. Bile Acid Sulfotransferase I From Rat Liver Sulfates Bile Acids And 3-Hydroxy Steroids: Purification, N-Terminal Amino Acid Sequence, and Kinetic Properties. J. Lipid Res. 1989, 30, 529–540. [Google Scholar] [CrossRef]
- Pillot, T.; Ouzzine, M.; Fournel-Gigleux, S.; Lafaurie, C.; Radominska, A.; Burchell, B.; Siest, G.; Magdalou, J. Glucuronidation of Hyodeoxycholic Acid in Human Liver. J. Biol. Chem. 1993, 268, 25636–25642. [Google Scholar] [CrossRef]
- Claudel, T.; Staels, B.; Kuipers, F. The farnesoid X Receptor a Molecular Link Between Bile Acid and Lipid and Glucose Metabolism. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 2020–2031. [Google Scholar] [CrossRef]
- Moore, L.B.; Maglich, J.M.; McKee, D.D.; Wisely, B.; Wilson, T.M.; Kliewer, S.A.; Lambert, M.H.; Moore, J.T. Pregnane X Receptor (PXR), Constitutive Androstane Receptor (CAR), And Benzoate X Receptor (BXR) Define Three Pharmacologically Distinct Classes of Nuclear Receptors. Mol. Endocrinol. 2002, 16, 977–986. [Google Scholar] [CrossRef]
- Jen, K.; Squires, E.J. Efficacy of Non-Nutritive Sorbent Materials as Intestinal-Binding Agents for the Control of Boar Taint. Animal 2011, 5, 1814–1820. [Google Scholar] [CrossRef]
- Claus, R.; Lösel, D.; Lacorn, M.; Mentshel, J.; Schenkel, H. Effects of Butyrate on Apoptosis in the Pig Colon and Its Consequences for Skatole Formation and Tissue Accumulation. J. Anim. Sci. 2003, 81, 239–248. [Google Scholar] [CrossRef]
- Li, X.; Jensen, B.B.; Canibe, N. The Mode of Action of Chicory Roots on Skatole Production in Entire Male Pigs Is Neither Via Reducing the Population of Skatole-Producing Bacteria nor Via Increased Butyrate Production in the Hindgut. Appl. Environ. Microbiol. 2019, 85, e02327-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, B.; Halestrap, A.P.; Paraskeva, C. Butyrate Can Act as a Stimulator of Growth or Inducer of Apoptosis in Human Colonic Epithelial Cell Lines Depending on the Presence of Alternative Energy Sources. Carcinogenesis 1997, 18, 1265–1270. [Google Scholar] [CrossRef] [PubMed]
- Knarreborg, A.; Beck, J.; Jensen, M.T.; Laue, A.; Agergaard, N.; Jensen, B.B. Effect of Non-Starch Polysaccharides on Production and Absorption of Indolic Compounds in Entire Male Pigs. Anim. Sci. 2002, 74, 445–453. [Google Scholar] [CrossRef]
- Whittington, F.M.; Nute, G.R.; Hughes, S.I.; McGivan, J.D.; Lean, I.J.; Wood, J.D.; Doran, E. Relationship Between Skatole and Androstenone Accumulation, And Cytochrome P4502E1 Expression in Meishan X Large White Pigs. Meat Sci. 2004, 67, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, M.K.; Zamaratskaia, G.; Ekstrand, B. In Vivo Effect of Dried Chicory Root (Cichorium Intybus L.) on Xenobiotica Metabolizing Cytochrome P450 Enzymes in Porcine Liver. Toxicol. Lett. 2011, 200, 88–91. [Google Scholar] [CrossRef]
- Simonsson, U.S.H.; Lindell, M.; Raffalli-Mathieu, F.; Lannerbro, A.; Honkakoski, P.; Lang, M.A. In Vivo and Mechanistic Evidence of Nuclear Receptor CAR Induction by Artemisinin. Eur. J. Clin. Investig. 2006, 36, 647–653. [Google Scholar] [CrossRef]
- Sachar, M.; Ma, X. Nuclear Receptors in Herb-Drug Interactions. Drug Metab. Rev. 2013, 45, 73–78. [Google Scholar] [CrossRef]
- Hernandez, J.P.; Mota, L.C.; Baldwin, W.S. Activation of CAR and PXR by dietary, Environmental and Occupational Chemicals Alters Drug Metabolism, Intermediary Metabolism and Cell Proliferation. Curr. Pharmacogenomics Person. Med. 2009, 7, 81–105. [Google Scholar] [CrossRef]
- Liu, W.; Wong, C. Oleanolic Acid Is a Selective Farnesoid X Receptor Modulator. Phytother. Res. 2010, 24, 369–373. [Google Scholar] [CrossRef]
- Le Bon, A.M.; Vernevaut, M.F.; Guenot, L.; Kahane, R.; Auger, J.; Arnault, I.; Haffner, T.; Siess, M.H. Effects of Garlic Powders with Varying Alliin Contents on Hepatic Drug Metabolizing Enzymes in Rats. J. Agric. Food Chem. 2003, 51, 7617–7623. [Google Scholar] [CrossRef]
- Cherrington, N.J.; Slitt, A.L.; Maher, J.M.; Zhang, X.X.; Zhang, J.; Huang, W.; Wan, Y.J.Y.; Moore, D.D.; Klaassen, C.D. Induction of Multidrug Resistance Protein 3 (MRP3) In Vivo Is Independent of Constitutive Androstane Receptor. Drug Metab. Dispos. 2003, 31, 1315–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, L.B.; Goodwin, B.; Jones, S.A.; Wisely, G.B.; Serabjit-Singh, C.J.; Willson, T.M.; Collins, J.L.; Kliewer, S.A. St. John’s Wort Induces Hepatic Drug Metabolism Through Activation of the Pregnane X Receptor. Proc. Natl. Acad. Sci. USA 2000, 97, 7500–7502. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, H.; Moore, L.B.; Johnson, M.D.L.; Maglich, J.M.; Goodwin, B.; Ittoop, O.R.R.; Wisely, B.; Creech, K.; Parks, D.J.; et al. The Phytoestrogen Coumestrol Is a Naturally Occurring Antagonist of the Human Pregnane X Receptor. Mol. Endocrinol. 2008, 22, 838–857. [Google Scholar] [CrossRef] [PubMed]
- Rajaraman, G.; Chen, J.; Chang, T.K.H. Ginkgolide A Contributes to the Potentiation of Acetaminophen Toxicity by Ginkgo Biloba Extract in Primary Cultures of Rat Hepatocytes. Toxicol. Appl. Pharmacol. 2006, 217, 225–233. [Google Scholar] [CrossRef]
- Chang, T.K.H.; Chen, J.; Teng, X.W. Distinct Role of Bilobalide and Ginkgolide in the Modulation of Rat CYP2B1 and CYP3A23 Gene Expression by Ginkgo biloba Extract in Cultured Hepatocytes. Drug Metab. Dispos. 2006, 34, 234–242. [Google Scholar] [CrossRef]
- Li, L.; Stanton, J.D.; Tolson, A.H.; Luo, Y.; Wang, H. Bioactive Terpenoids and Flavonoids from Ginkgo Biloba Extract Induce the Expression of Hepatic Drug-Metabolizing Enzymes Through Pregnane X Receptor, Constitutive Androstane Receptor, and Aryl Hydrocarbon Receptor-Mediated Pathways. Pharm. Res. 2009, 26, 872–882. [Google Scholar] [CrossRef]
- Brobst, D.E.; Ding, X.; Creech, K.L.; Goodwin, B.; Kelley, B.; Staudinger, J.L. Guggulsterone Activates Multiple Nuclear Receptors and Induces CYP3A Gene Expression Through the Pregnane X Receptor. J. Pharmacol. Exp. Ther. 2004, 310, 528–535. [Google Scholar] [CrossRef]
- Ding, X.; Staudinger, J.L. The Ratio of Constitutive Androstane Receptor to Pregnane X Receptor Determines the Activity of Guggulsterone Against the Cyp2b10 Promoter. J. Pharmacol. Exp. Ther. 2005, 314, 120–127. [Google Scholar] [CrossRef]
- Feng, W.; Ao, H.; Peng, C. Gut Microbiota, Short-Chain Fatty Acids, And Herbal Medicines. Front. Pharmacol. 2018, 9, 1354. [Google Scholar] [CrossRef]
- Bachs, L.; Parés, A.; Elena, M.; Piera, C.; Rodsés, J. Effects of Long-Term Rifampicin Administration in Primary Biliary Cirrhosis. Gastroenterology 1992, 102, 2077–2080. [Google Scholar] [CrossRef]
- Kurian, R.; Hedrich, W.; Mackowiak, B.; Li, L.; Wang, H. CITCO as an Adjuvant Facilitates CHOP-Based Lymphoma Treatment in Hcar-Transgenic Mice. Cells 2020, 9, 2520. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Zhang, F.; Chen, L.; Yang, Q.; Su, H.; Yang, X.; He, H.; Ling, M.; Zheng, J.; Duan, C.; et al. Dietary Chenodeoxycholic Acid Improves Growth Performance and Intestinal Health by Altering Serum Metabolic Profiles and Gut Bacteria in Weaned Piglets. Anim. Nutr. 2021, 7, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Burkina, V.; Rasmussen, M.K.; Oliinychenko, Y.; Zamaratskaia, G. Porcine Cytochrome 2A19 and 2E1. Basic Clin. Pharmacol. Toxicol. 2019, 124, 32–39. [Google Scholar] [CrossRef]
- Squires, E.J.; Bone, C.; Cameron, J. Pork Production with Entire Males: Directions for Control of Boar Taint. Animals 2020, 10, 1665. [Google Scholar] [CrossRef] [PubMed]
- Drag, M.H.; Kogelman, L.J.A.; Maribo, H.; Meinert, L.; Thomsen, P.D.; Kadarmideen, H.N. Characterization of Eqttls Associated with Androstenone by RNA Sequencing in Porcine Testis. Physiol. Genom. 2019, 51, 488–499. [Google Scholar] [CrossRef]
- Drag, M.; Hansen, M.B.; Kadarmideen, H.N. Systems Genomics Study Reveals Expression Quantitative Trait Loci, Regulator Genes and Pathways Associated with Boar Taint in Pigs. PLoS ONE 2018, 13, e0192673. [Google Scholar] [CrossRef]
- Drag, M.; Skinkyté-Juskiené, R.; Do, D.N.; Kogelman, L.J.A.; Kadarmideen, H.N. Differential Expression and Co-Expression Gene Networks Reveal Candidate Biomarkers of Boar Taint in Non-Castrated Pigs. Sci. Rep. 2017, 7, 12205. [Google Scholar] [CrossRef]
- Duarte, D.A.; Schroyen, M.; Mota, R.R.; Vanderick, S.; Gengler, N. Recent Advances on Boar Taint Reduction as an Alternative to Castration: A Review. J. Appl. Genet. 2021, 62, 137–150. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bone, C.; Squires, E.J. Nuclear Receptor Pathways Mediating the Development of Boar Taint. Metabolites 2022, 12, 785. https://doi.org/10.3390/metabo12090785
Bone C, Squires EJ. Nuclear Receptor Pathways Mediating the Development of Boar Taint. Metabolites. 2022; 12(9):785. https://doi.org/10.3390/metabo12090785
Chicago/Turabian StyleBone, Christine, and E. James Squires. 2022. "Nuclear Receptor Pathways Mediating the Development of Boar Taint" Metabolites 12, no. 9: 785. https://doi.org/10.3390/metabo12090785
APA StyleBone, C., & Squires, E. J. (2022). Nuclear Receptor Pathways Mediating the Development of Boar Taint. Metabolites, 12(9), 785. https://doi.org/10.3390/metabo12090785