


The Effect of Diet on the Cardiac Circadian Clock in Mice: A Systematic Review

 ,
,  , , and
, , and
Abstract
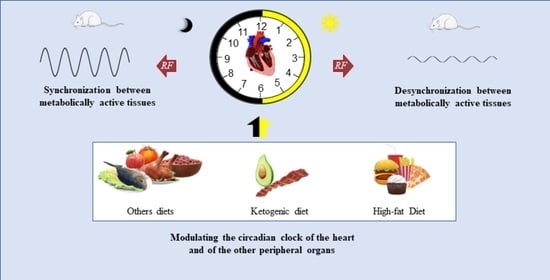
1. Introduction
2. Materials and Methods
2.1. Search Strategy and Data Extraction
- -
- For PUBMED: (“diet” OR “nutrition” OR “feeding”) AND (“circadian clock” OR “chronobiology” OR “biological clock” OR “clock proteins” OR “clock gene”) AND (“heart” OR “cardiac” OR “cardiomyocyte” OR “ventricle”) AND (“mice” OR “mouse”);
- -
- For Scopus: (“diet” OR “nutrition” OR “feeding”) AND (“circadian clock” OR “chronobiology” OR “biological clock” OR “clock proteins” OR “clock gene”) AND (“heart” OR “cardiac” OR “cardiomyocyte” OR “ventricle”) AND (“mice” OR “mouse”);
- -
- For Embase: (“diet” OR “nutrition” OR “feeding”) AND (“circadian clock” OR “chronobiology” OR “biological clock” OR “clock proteins” OR “clock gene”) AND (“heart” OR “cardiac” OR “cardiomyocyte” OR “ventricle”) AND (“mice” OR “mouse”).
2.2. Quality Assessment of Studies
3. Results
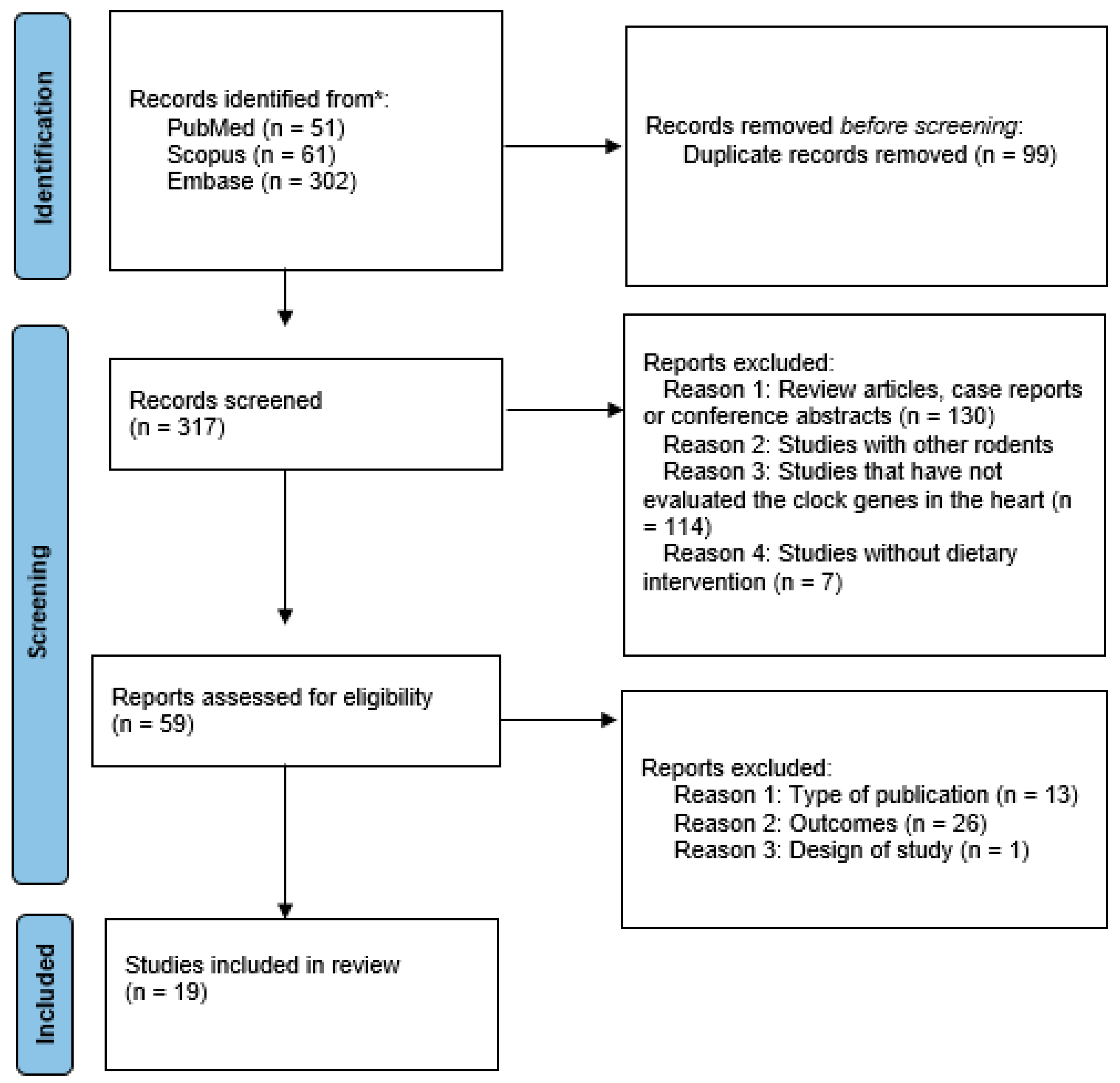
3.1. Literature Data
3.2. Quality Assessment of Studies
4. Discussion
5. Limitations and Future Research
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Panda, S.; Hogenesch, J.B.; Kay, S.A. Circadian Rhythms from Flies to Human. Nature 2002, 417, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Froy, O. Metabolism and Circadian Rhythms—Implications for Obesity. Endocr. Rev. 2010, 31, 1–24. [Google Scholar] [CrossRef]
- Espinoza Gallardo, A.C.; López-Espinoza, A.; Vázquez-Cisneros, L.C.; Zepeda-Salvador, A.P.; Santillano-Herrera, D. Light-Dark Cycle Inversion Effect on Food Intake and Body Weight in Rats. Nutr. Hosp. 2021, 38, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Stenvers, D.J.; Scheer, F.A.J.L.; Schrauwen, P.; la Fleur, S.E.; Kalsbeek, A. Circadian Clocks and Insulin Resistance. Nat. Rev. Endocrinol. 2019, 15, 75–89. [Google Scholar] [CrossRef] [PubMed]
- de Assis, L.V.M.; Oster, H. The Circadian Clock and Metabolic Homeostasis: Entangled Networks. Cell. Mol. Life Sci. 2021, 78, 4563–4587. [Google Scholar] [CrossRef] [PubMed]
- Duffield, G.E.; Best, J.D.; Meurers, B.H.; Bittner, A.; Loros, J.J.; Dunlap, J.C. Circadian Programs of Transcriptional Activation, Signaling, and Protein Turnover Revealed by Microarray Analysis of Mammalian Cells. Curr. Biol. 2002, 12, 551–557. [Google Scholar] [CrossRef]
- West, A.C.; Smith, L.; Ray, D.W.; Loudon, A.S.I.; Brown, T.M.; Bechtold, D.A. Misalignment with the External Light Environment Drives Metabolic and Cardiac Dysfunction. Nat. Commun. 2017, 8, 417. [Google Scholar] [CrossRef]
- Azzi, A.; Dallmann, R.; Casserly, A.; Rehrauer, H.; Patrignani, A.; Maier, B.; Kramer, A.; Brown, S.A. Circadian Behavior Is Light-Reprogrammed by Plastic DNA Methylation. Nat. Neurosci. 2014, 17, 377–382. [Google Scholar] [CrossRef]
- Schibler, U.; Ripperger, J.; Brown, S.A. Peripheral Circadian Oscillators in Mammals: Time and Food. J. Biol. Rhythms 2003, 18, 250–260. [Google Scholar] [CrossRef]
- Stephan, F.K. The “Other” Circadian System: Food as a Zeitgeber. J. Biol. Rhythms 2002, 17, 284–292. [Google Scholar] [CrossRef]
- Froy, O. The Relationship between Nutrition and Circadian Rhythms in Mammals. Front. Neuroendocrinol. 2007, 28, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Melkani, G.C.; Panda, S. Time-Restricted Feeding for Prevention and Treatment of Cardiometabolic Disorders. J. Physiol. 2017, 595, 3691–3700. [Google Scholar] [CrossRef] [PubMed]
- Duncan, M.J.; Smith, J.T.; Narbaiza, J.; Mueez, F.; Bustle, L.B.; Qureshi, S.; Fieseler, C.; Legan, S.J. Restricting Feeding to the Active Phase in Middle-Aged Mice Attenuates Adverse Metabolic Effects of a High-Fat Diet. Physiol. Behav. 2016, 167, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sundaram, S.; Yan, L. Time-Restricted Feeding Reduces Adiposity in Mice Fed a High-Fat Diet. Nutr. Res. 2016, 36, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.-Y.; Villegas-Montoya, C.; Boland, B.B.; Blasier, Z.; Egbejimi, O.; Gonzalez, R.; Kueht, M.; McElfresh, T.A.; Brewer, R.A.; Chandler, M.P.; et al. Influence of Dark Phase Restricted High Fat Feeding on Myocardial Adaptation in Mice. J. Mol. Cell Cardiol. 2013, 55, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]
- Hooijmans, C.R.; Rovers, M.M.; de Vries, R.B.M.; Leenaars, M.; Ritskes-Hoitinga, M.; Langendam, M.W. SYRCLE’s Risk of Bias Tool for Animal Studies. BMC Med. Res. Methodol. 2014, 14, 43. [Google Scholar] [CrossRef]
- Goh, B.C.; Wu, X.; Evans, A.E.; Johnson, M.L.; Hill, M.R.; Gimble, J.M. Food Entrainment of Circadian Gene Expression Altered in PPARα−/− Brown Fat and Heart. Biochem. Biophys. Res. Commun. 2007, 360, 828–833. [Google Scholar] [CrossRef][Green Version]
- Reilly, D.F.; Curtis, A.M.; Cheng, Y.; Westgate, E.J.; Rudic, R.D.; Paschos, G.; Morris, J.; Ouyang, M.; Thomas, S.A.; FitzGerald, G.A. Peripheral Circadian Clock Rhythmicity Is Retained in the Absence of Adrenergic Signaling. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 121–126. [Google Scholar] [CrossRef]
- Bray, M.S.; Ratcliffe, W.F.; Grenett, M.H.; Brewer, R.A.; Gamble, K.L.; Young, M.E. Quantitative Analysis of Light-Phase Restricted Feeding Reveals Metabolic Dyssynchrony in Mice. Int. J. Obes. 2013, 37, 843–852. [Google Scholar] [CrossRef]
- Mukherji, A.; Kobiita, A.; Chambon, P. Shifting the Feeding of Mice to the Rest Phase Creates Metabolic Alterations, Which, on Their Own, Shift the Peripheral Circadian Clocks by 12 Hours. Proc. Natl. Acad. Sci. USA 2015, 112, E6683–E6690. [Google Scholar] [CrossRef] [PubMed]
- Noyan, H.; El-Mounayri, O.; Isserlin, R.; Arab, S.; Momen, A.; Cheng, H.S.; Wu, J.; Afroze, T.; Li, R.-K.; Fish, J.E.; et al. Cardioprotective Signature of Short-Term Caloric Restriction. PLoS ONE 2015, 10, e0130658. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; Deng, F.; Zhou, M.; Huang, R.; Ma, X.; Tian, H.; Tan, Y.; Chen, X.; Deng, D.; Shui, G.; et al. A Multi-Tissue Multi-Omics Analysis Reveals Distinct Kineztics in Entrainment of Diurnal Transcriptomes by Inverted Feeding. iScience 2021, 24, 102335. [Google Scholar] [CrossRef]
- Hou, L.; Lu, C.; Huang, Y.; Chen, S.; Hua, L.; Qian, R. Effect of Hyperlipidemia on the Expression of Circadian Genes in Apolipoprotein E Knock-out Atherosclerotic Mice. Lipids Health Dis. 2009, 8, 60. [Google Scholar] [CrossRef] [PubMed]
- Oishi, K.; Uchida, D.; Ohkura, N.; Horie, S. PPARα Deficiency Augments a Ketogenic Diet-Induced Circadian PAI-1 Expression Possibly through PPARγ Activation in the Liver. Biochem. Biophys. Res. Commun. 2010, 401, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Oishi, K.; Miyazaki, K.; Uchida, D.; Ohkura, N.; Wakabayashi, M.; Doi, R.; Matsuda, J.; Ishida, N. PERIOD2 Is a Circadian Negative Regulator of PAI-1 Gene Expression in Mice. J. Mol. Cell. Cardiol. 2009, 46, 545–552. [Google Scholar] [CrossRef]
- Tsai, J.-Y.; Kienesberger, P.C.; Pulinilkunnil, T.; Sailors, M.H.; Durgan, D.J.; Villegas-Montoya, C.; Jahoor, A.; Gonzalez, R.; Garvey, M.E.; Boland, B.; et al. Direct Regulation of Myocardial Triglyceride Metabolism by the Cardiomyocyte Circadian Clock. J. Biol. Chem. 2010, 285, 2918–2929. [Google Scholar] [CrossRef]
- Wang, D.; Chen, S.; Liu, M.; Liu, C. Maternal Obesity Disrupts Circadian Rhythms of Clock and Metabolic Genes in the Offspring Heart and Liver. Chronobiol. Int. 2015, 32, 615–626. [Google Scholar] [CrossRef]
- Oishi, K.; Yasumoto, Y.; Higo-Yamamoto, S.; Yamamoto, S.; Ohkura, N. Feeding Cycle-Dependent Circulating Insulin Fluctuation Is Not a Dominant Zeitgeber for Mouse Peripheral Clocks except in the Liver: Differences between Endogenous and Exogenous Insulin Effects. Biochem. Biophys. Res. Commun. 2017, 483, 165–170. [Google Scholar] [CrossRef]
- Reitz, C.J.; Alibhai, F.J.; de Lima-Seolin, B.G.; Nemec-Bakk, A.; Khaper, N.; Martino, T.A. Circadian Mutant Mice with Obesity and Metabolic Syndrome Are Resilient to Cardiovascular Disease. Am. J. Physiol.-Heart Circ. Physiol. 2020, 319, H1097–H1111. [Google Scholar] [CrossRef]
- Mia, S.; Sonkar, R.; Williams, L.; Latimer, M.N.; Frayne Robillard, I.; Diwan, A.; Frank, S.J.; des Rosiers, C.; Young, M.E. Impact of Obesity on Day-night Differences in Cardiac Metabolism. FASEB J. 2021, 35, e21298. [Google Scholar] [CrossRef] [PubMed]
- Oishi, K.; Uchida, D.; Ohkura, N.; Doi, R.; Ishida, N.; Kadota, K.; Horie, S. Ketogenic Diet Disrupts the Circadian Clock and Increases Hypofibrinolytic Risk by Inducing Expression of Plasminogen Activator Inhibitor-1. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1571–1577. [Google Scholar] [CrossRef] [PubMed]
- Marques, F.Z.; Nelson, E.; Chu, P.-Y.; Horlock, D.; Fiedler, A.; Ziemann, M.; Tan, J.K.; Kuruppu, S.; Rajapakse, N.W.; El-Osta, A.; et al. High-Fiber Diet and Acetate Supplementation Change the Gut Microbiota and Prevent the Development of Hypertension and Heart Failure in Hypertensive Mice. Circulation 2017, 135, 964–977. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Hussein, A.I.; Horowitz, N.; Carroll, D.; Gower, A.C.; Demissie, S.; Gerstenfeld, L.C. Hypophosphatemia Regulates Molecular Mechanisms of Circadian Rhythm. Sci. Rep. 2018, 8, 13756. [Google Scholar] [CrossRef] [PubMed]
- Murata, K.; Mimura, A.; Suzuki, H.; Mikami, N.; Hamada, Y.; Kato, K.; Iki, N.; Ishida, M.; Daitoku, Y.; Tanimoto, Y.; et al. Efficient Induction of Proximity-Dependent Labelling by Biotin Feeding in BMAL1-BioID Knock-in Mice. J. Biochem. 2021, 170, 453–461. [Google Scholar] [CrossRef]
- Latimer, M.N.; Sonkar, R.; Mia, S.; Frayne, I.R.; Carter, K.J.; Johnson, C.A.; Rana, S.; Xie, M.; Rowe, G.C.; Wende, A.R.; et al. Branched Chain Amino Acids Selectively Promote Cardiac Growth at the End of the Awake Period. J. Mol. Cell Cardiol. 2021, 157, 31–44. [Google Scholar] [CrossRef]
- Gonzalez-Lopez, E.; Vrana, K.E. Dopamine Beta-hydroxylase and Its Genetic Variants in Human Health and Disease. J. Neurochem. 2020, 152, 157–181. [Google Scholar] [CrossRef] [PubMed]
- Durgan, D.J.; Young, M.E. The Cardiomyocyte Circadian Clock: Emerging Roles in Health and Disease. Circ. Res. 2010, 106, 647–658. [Google Scholar] [CrossRef]
- Han, X.; Turdi, S.; Hu, N.; Guo, R.; Zhang, Y.; Ren, J. Influence of Long-Term Caloric Restriction on Myocardial and Cardiomyocyte Contractile Function and Autophagy in Mice. J. Nutr. Biochem. 2012, 23, 1592–1599. [Google Scholar] [CrossRef]
- Eckle, T.; Hartmann, K.; Bonney, S.; Reithel, S.; Mittelbronn, M.; Walker, L.A.; Lowes, B.D.; Han, J.; Borchers, C.H.; Buttrick, P.M.; et al. Adora2b-Elicited Per2 Stabilization Promotes a HIF-Dependent Metabolic Switch Crucial for Myocardial Adaptation to Ischemia. Nat. Med. 2012, 18, 774–782. [Google Scholar] [CrossRef]
- Pan, X.; Jiang, X.C.; Hussain, M.M. Impaired Cholesterol Metabolism and Enhanced Atherosclerosis in Clock Mutant Mice. Circulation 2013, 128, 1758–1769. [Google Scholar] [CrossRef] [PubMed]
- Schober, A.; Blay, R.M.; Saboor Maleki, S.; Zahedi, F.; Winklmaier, A.E.; Kakar, M.Y.; Baatsch, I.M.; Zhu, M.; Geißler, C.; Fusco, A.E.; et al. MicroRNA-21 Controls Circadian Regulation of Apoptosis in Atherosclerotic Lesions. Circulation 2021, 144, 1059–1073. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Tang, Q.; Nie, J.; Zhang, C.; Zhou, X.; Yu, S.; Sun, J.; Cheng, X.; Dong, N.; Hu, Y.; et al. BMAL1-Downregulation Aggravates Porphyromonas Gingivalis -Induced Atherosclerosis by Encouraging Oxidative Stress. Circ. Res. 2020, 126, E15–E29. [Google Scholar] [CrossRef] [PubMed]
- Oishi, K.; Ohkura, N.; Kasamatsu, M.; Fukushima, N.; Shirai, H.; Matsuda, J.; Horie, S.; Ishida, N. Tissue-Specific Augmentation of Circadian PAI-1 Expression in Mice with Streptozotocin-Induced Diabetes. Thromb. Res. 2004, 114, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; ZhuGe, F.; Sun, L.; Ni, Y.; Fu, O.; Gao, G.; Chen, J.; Kato, H.; Fu, Z. Enhanced Effect of Daytime Restricted Feeding on the Circadian Rhythm of Streptozotocin-Induced Type 2 Diabetic Rats. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E1027–E1035. [Google Scholar] [CrossRef][Green Version]
- le Minh, N. Glucocorticoid Hormones Inhibit Food-Induced Phase-Shifting of Peripheral Circadian Oscillators. EMBO J. 2001, 20, 7128–7136. [Google Scholar] [CrossRef]
- Kalra, S.; Singla, R.; Rosha, R.; Dhawan, M. Ketogenic Diet: Situational Analysis of Current Nutrition Guidelines. J. Pak. Med. Assoc. 2018, 68, 1836–1839. Available online: https://www.researchgate.net/publication/329288515_Ketogenic_diet_Situational_analysis_of_current_nutrition_guidelines (accessed on 28 October 2022).
- Choi, Y.J.; Jeon, S.M.; Shin, S. Impact of a Ketogenic Diet on Metabolic Parameters in Patients with Obesity or Overweight and with or without Type 2 Diabetes: A Meta-Analysis of Randomized Controlled Trials. Nutrients 2020, 12, 2005. [Google Scholar] [CrossRef]
- Bechtold, D.A. Energy-Responsive Timekeeping. J. Genet. 2008, 87, 447–458. [Google Scholar] [CrossRef]
- Schibler, U. The 2008 Pittendrigh/Aschoff Lecture: Peripheral Phase Coordination in the Mammalian Circadian Timing System. J. Biol. Rhythms 2009, 24, 3–15. [Google Scholar] [CrossRef]
- Ba, L.; Gao, J.; Chen, Y.; Qi, H.; Dong, C.; Pan, H.; Zhang, Q.; Shi, P.; Song, C.; Guan, X.; et al. Allicin Attenuates Pathological Cardiac Hypertrophy by Inhibiting Autophagy via Activation of PI3K/Akt/MTOR and MAPK/ERK/MTOR Signaling Pathways. Phytomedicine 2019, 58, 152765. [Google Scholar] [CrossRef] [PubMed]
- Venn, B.J. Macronutrients and Human Health for the 21st Century. Nutrients 2020, 12, 2363. [Google Scholar] [CrossRef] [PubMed]
- Rautiainen, S.; Manson, J.E.; Lichtenstein, A.H.; Sesso, H.D. Dietary Supplements and Disease Prevention—A Global Overview. Nat. Rev. Endocrinol. 2016, 12, 407–420. [Google Scholar] [CrossRef]
- May, D.G.; Scott, K.L.; Campos, A.R.; Roux, K.J. Comparative Application of BioID and TurboID for Protein-Proximity Biotinylation. Cells 2020, 9, 1070. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Parameters | Inclusion Criteria | Exclusion Criteria |
|---|---|---|
| Population | Studies conducted with mice | Studies conducted with other rodents |
| Intervention | Diet, Restricted feeding | No dietary intervention |
| Comparison | Not applicable | Not applicable |
| Results | Clock genes | Other genes |
| Study Design | Case-control studies and experimental trials Published after 2007 | Literature review, cross-sectional studies, letters to the editor Published before 2007, studies not in the English language |
| Author/Year | Species/Strain/Sex | Objectives | Diets/Composition | Dietetic Intervention | Methodology |
|---|---|---|---|---|---|
| Goh et al., 2007 [18] | Male C57Bl/6N × Sv/129, C57Bl/6N × Sv/129 homozygous Pparα-null mice. | To investigate the impact of PPARα deficiency on the expression of genes encoding circadian transcription factors and PPARα responsive proteins in peripheral tissues following temporally restricted food access. | Regular chow diet (Purina 5015) | Mice were compared under ad libitum or restricted food access for the expression of circadian transcription factor-encoding mRNAs. | Gene expression by qPCR |
| Reilly et al., 2008 [19] | Males Dbh+/−, Dbh−/−, C57Bl/6J mice | To investigate the role of the SCN on the entrainment of the heart and vasculature oscillators. | Regular chow diet or RF (for 6 h). | Mice were submitted to RF during the light period (7 a.m. to 7 p.m.) or ad libitum for 5 days. On the 5th day, mice were placed in DD for 24 h and then euthanized. | Gene expression by qPCR |
| Oishi et al., 2009a [26] | Male C57Bl/6J and Per2 transgenic (overexpressing rat PER2) mice. | To investigate the function of PER2 in the circadian regulation of fibrinolysis and obesity-induced hypofibrinolysis. | Normal diet (CE-2; Clean Japan Inc., Tokyo, Japan); high-fat/high-sucrose diet (with 54.5% fat, 28.3% carbohydrates, and 17.2% protein; F2HFHSD; Oriental Yeast Co., Ltd., Tokyo, Japan) | Mice were fed with normal diet, or HFSD for 12 weeks to induce obesity and then euthanized. | Gene and protein expression by qPCR and Western blotting, respectively; measurement of blood metabolic parameters; measurement of plasma and tissue PAI-1; stimulation with 10 ng/mL TGF-β1. |
| Oishi et al., 2009b [32] | Male Jcl:ICR mice. | To investigate the effect of a ketogenic diet on fibrinolytic functions. | Normal diet (CE-2; 12.6% fat, 58.3% carbohydrate, and 29.3% protein, Clea Japan Inc); ketogenic diet (KD; 94.8% fat, 0.1% carbohydrate, and 4.8% protein, w/w; modified AIN-93G; Oriental Yeast Co. Ltd.). | Mice were fed with normal or KD under LD 12:12 for 14 days. Mice were euthanized and the tissues were collected. Mice with control or KD stayed under LD 12:12, and then they were transferred to constant DD. | Gene expression by qPCR; measurement of blood metabolic parameters and plasma PAI-1; drinking behavior analyses by Chronobiology Kits (Stanford Software Systems). |
| Hou et al., 2009 [24] | Male ApoE−/−, C57Bl/6J mice. | To investigate the effect of hyperlipidemia on the expression of circadian genes in an atherosclerotic mouse model. | Regular chow diet, high-fat diet (HFD with 0.15% cholesterol and 21% fat). | ApoE−/− mice were fed with regular diet or HFD, and C57Bl/6J mice were fed with regular diet in an LD cycle for 2 weeks. Mice were transferred to a DD cycle for 3 weeks and then euthanized. | Analysis of serum lipid, analyses of atherosclerotic plaques or foam cells by oil red O staining, gene expression by qPCR. |
| Tsai et al., 2010 [27] | Male WT, CCM mice | To investigate the role of the cardiomyocyte circadian clock in the metabolic adaptation of the heart to chronic high-fat feeding. | Standard murine diet (Teklad Lab Animal Diets, Harlan Laboratories, Indianapolis, IN), HFD (with 45% fat, Research Diets, New Brunswick, NJ), control diet (with 10% fat, Research Diets, New Brunswick, NJ) | Mice were fed with a standard diet, HFD, or control diet for 16 weeks ad libitum and then euthanized. | Echocardiography; plasma hormone, substrate and AMPK activity measurements; microarray analyses; gene and protein expression by qPCR and Western blotting, respectively; myocardial triglyceride content measurements; isolated working mouse heart perfusions; lipid extraction, fractionation, and profiling; analyses of heart sections by oil red O staining. |
| Oishi et al., 2010 [25] | Male Pparα-null, WT mice. | To investigate the role of PPARα in ketogenic diet-induced PAI-1 gene expression. | Normal diet (AIN-93M, Oriental Yeast Co., Ltd., Tokyo, Japan), ketogenic diet (KD; 94.8% fat, 0.1% carbohydrate and 4.8% protein, w/w; modified AIN-93G; Oriental Yeast Co Ltd.). | Mice were fed with normal diet supplemented with bezafibrate or a KD. The PPARγ antagonist bisphenol A diglycidyl ether was added to diets for 5 days. Mice were euthanized and the tissues were collected. | Gene expression by qPCR; measurement of plasma PAI-1. |
| Bray et al., 2013 [20] | Male C57Bl/6J mice. | To investigate the effects of RF during the light (sleep) phase on body weight gain, diurnal variation in energy balance, gene expression, and humoral factors. | Standard rodent diet (Harlan NIH-31 irradiated open formula mouse/rat diet, 4.7% fat) | Mice were fed with a standard diet under light-phase (ZT0-ZT12) or dark-phase (ZT12-ZT24) RF during 9 days and then euthanized. | Continuous 24-h monitoring of behavioral and physiological parameters; gene expression by qPCR; humoral factor measurements |
| Wang et al., 2015 [28] | Female C57Bl/6J mice, and 35 male pups postnatal day 17. | To investigate the effect of maternal obesity on the circadian clock and metabolism in the heart and liver of the offspring. | Normal diet (ND, D12450B, fat content 10%; Research Diets Inc., New Brunswick, NJ, USA) or HFD (D12492, fat content 60%) | The animals were fed normal diet or HFD for 6 weeks until mating, during gestation and lactation (until day 16 of lactation). On day 16 of lactation, both groups were fed ND. The pups were suckled until day 21 of life and then fed ND until the day of euthanasia. | Gene and protein expression by qPCR and Western blot, respectively; measurement of the body weight; serological analyses. |
| Noyan et al., 2015 [22] | Male C57Bl/6J mice. | To investigate the effect of the short-term, mild CR before induction of experimental MI to protect the heart from ischemic injury and to understand the underlying molecular pathways. | Regular chow diet or CR (30% less than the calculated mean daily AL food consumption) | Mice were fed ad libitum, stressed AL diet, or CR diet for 7 days, prior to MI via permanent coronary ligation and then euthanized. | Protein and gene expression by Western blotting; mRNA and microRNA analyses; signaling assessments in myocardial tissue; pathway enrichment analysis; infarct size (2 days post-MI); cardiac hemodynamics (before and 2 days post-MI); protein abundance of caspase. |
| Mukherji et al., 2015 [21] | Male C57Bl/6J, Pparαhep−/−, Pparαiec−/−, Bmal1hep−/−, Bmal1iec−/−, RevErbαhep−/−, RevErbαiec−/−, adrenalectomized mice | To investigate molecular mechanisms generated by shifting feeding to the rest phase and how this environmental cue alters the expression of circadian clock components, thereby leading to obesity and metabolic syndrome-like pathology. | Control diet | Mice were fed with a control diet ad libitum or RF during the 12 h light phase (ZT0-ZT12) under LD 12h:12h. | Isolation of IECs; determination of RNA transcripts; protein immunoblots; measurement of NAD+; FGF21 measurement; chip assays; plasma metabolic analysis. |
| Oishi et al., 2017 [29] | Male C57Bl/6J mice | To investigate the involvement of feeding cycle-dependent endogenous insulin rhythms in the circadian regulation of peripheral clocks, and the effect of exogenous insulin on the expression of clock genes. | High-fat/high-sucrose diet (F2HFHSD; Oriental Yeast Co., Ltd., Tokyo, Japan), normal diet (Clea Japan Inc., Tokyo, Japan). | Mice were fed HFSD for 8 h during nighttime (ZT14-22) or daytime (ZT2-10) for one week. After this period mice were euthanized and the tissues were collected. | Measurement of blood hormones; gene expression by qPCR; assay of phosphorylated AKT; wheel-running activity analyses. |
| Marques et al., 2017 [33] | Male C57Bl/6J mice | To investigate the effect of a high-fiber diet and supplementation with the short-chain fatty acid (acetate) on the gut microbiota and the prevention of cardiovascular disease. | Control diet (normal chow, 47.6%), high-fiber diet (72.7% fiber, SF11-025; Specialty Feeds, Perth, Australia), or SCFA supplementation (200 mmol/L magnesium acetate, Meck Millipore, 1.05819.1000) | Mice were fed with a high-fiber diet or acetate supplementation for 3 weeks before sham or DOCA surgery. | Morphological analyses in the heart, kidney, and lung; histological analyses in the heart and kidney; bioinformatic analyses; renal and cardiac transcriptome; analyses of the composition of the gut microbiota. |
| Noguchi et al., 2018 [34] | Male C57Bl/6J, Per2:Luciferase knock-in, A/J, C3H/HeJ mice. | To investigate whether the levels of systemic phosphate regulate the circadian cycle in peripheral tissues. | Normal diet (with 0.4% phosphate), low phosphate diet (with 0.06% phosphate; Teklad 2018; 0.65% phosphorus). | Mice were fed with normal diet or low phosphate diet. The phosphate-restricted diet group was fed two days before fracture and for 16 more days. A normal diet was reintroduced afterward. | Circadian rhythm studies were performed 10 days post-fracture; serum analyses; analyses of cartilage and bone by CECT and µCT; gene expression by qPCR; microarray analyses; analysis of periodicity of Per2 in callus organ culture. |
| Reitz et al., 2020 [30] | Male ClockΔ19/Δ19 and WT mice. | To investigate whether ClockΔ19/Δ19 mice develop or are resilient to obesity induced cardiovascular disease. | HFD (45% fat, 20% protein, 35% carbohydrate, TD.06415, Envigo Teklad Diets), normal standard chow diet (10% fat, 20% protein, 70% carbohydrates, TD.08806, Envigo Teklad Diets) | Mice were fed with HFD or normal standard chow diet for 24 weeks and then euthanized. | CLAMS for evaluation of food/calorie intake and metabolic parameters; metabolic measurements; morphometric, hemodynamics, histological analyses; echocardiography; gene and protein expression by qPCR and Western blotting, respectively; microarray and bioinformatics analyses. |
| Mia et al., 2021 [31] | Male C57Bl/6J mice | To investigate diurnal metabolic inflexibility in the heart in obesity. | Control diet (10% calories from fat, Research Diets, New Brunswick, NJ; catalog number D12450K) or HFD (45% calories from fat, Research Diets, New Brunswick, NJ; catalog number D12451) | Mice were fed with control or HFD ad libitum for 18 weeks, followed by access only during the 12 h dark phase for 2 weeks. | Behavioral and metabolic monitoring; histological analysis, transcriptome, echocardiographic image, gene and protein expression, lipidomic analyses; NEFA quantification; isolated heart perfusion. |
| Xin et al., 2021 [23] | Male and female C57Bl/6J mice | To investigate the transition kinetics during inverted feeding. | Normal chow diet (Rodent maintenance diet; Hunan SJA Laboratory Animal Co. LTD). | Female mice were fed with normal chow diet for 7 days, following DRF or NRF for 7 or 36 days. They were submitted to LL for 9 days, following DRF and LL for 9 additional days. Male mice were fed with normal chow diet for 7 days, following DRF or NRF for 7 additional days. After this period, female and male mice were euthanized. | Transcriptome and metabolomic profiling; food intake, body weight, and locomotor activity analyses; global profiling of transcripts; untargeted metabolomics; targeted lipidomics; Acyl-CoA quantification by LC/MS; gene expression by qPCR; transcriptome analyses; circadian rhythmicity analyses; phase set and cistrome enrichment analyses; heatmap of expression profile. |
| Murata et al., 2021 [35] | BMAL1-BioID, C57Bl/6J, homozygous 540 EGFP-pAflox mice | To investigate the effect of biotin diet on protein biotinylation in several tissues in the BMAL1-BioID mouse model. | Chow diet, high-biotin diet (0.5% biotin; Fujifilm Wako Pure Chemical, #021-08712). | Mice were fed with a biotin-rich diet or chow diet ad libitum for 7 days and then euthanized. | Protein expression by Western blotting; histological analyses; streptavidin blot analysis; biotin labeling assay in vitro; co-immunoprecipitation. |
| Latimer et al., 2021 [36] | Male C57Bl/6J, CBK, CON mice | To investigate the effect of time of day of dietary BCAA consumption on physiological responses (cardiac growth) and its pathological implications | Low BCAA diet (Teklad TD.150662 custom diet; with leucine, isoleucine, and valine 3-fold lower than the standard diet), high BCAA diet (Teklad TD.170323 custom diet; with leucine, isoleucine, and valine 2-fold higher than the standard diet) or a standard diet (Teklad TD.170323 custom diet). | The dietary intervention occurred acutely and chronically. Acute intervention: mice were fed with an early high BCAA diet or early low BCAA diet, and a late high BCAA diet or late low BCAA diet for 4 h. Chronic intervention: mice were fed with an early high BCAA diet, and a late high BCAA diet for 4 weeks or 6 weeks. | Transverse aortic constriction; CLAMS for evaluation of food/calorie intake, energy expenditure, and physical activity; spectrometry (plasma BCAA levels); quantitative magnetic resonance imaging (lean and fat body mass); gene and protein expression by qPCR and Western blot, respectively; histological evaluation. |
| Dietary Interventions/Diet | Author/Year | Alterations in the Circadian Clock/Gene Expression of the Heart | Alterations in the Circadian Clock/Gene Expression of the Other Tissues | Positive (+), Negative (−) or Neutral (N) Effect of the Diet/Dietary Intervention |
|---|---|---|---|---|
| Restricting feeding | Goh et al., 2007 [18] | WT mice with RF: shifted the acrophase of circadian gene expression by 5.8 ± 2.0 h and reduced the amplitude of Bmal1. Pparα-null mice with RF: shifted the acrophase of circadian gene expression 14.2 ± 2.5 h, reduced the amplitude Rev-erbα, and increased that of Bmal1. | WT and Pparα-null mice: similar expression profiles and amplitudes for Per1 and Per3 genes with mean acrophase of ZT11.4 and ZT10.6, respectively (liver, BAT, and eWAT) reflecting a 10 h time difference from Bmal1 and Npas2. | (−) |
| Restricting feeding | Reilly et al., 2008 [19] | Dbh−/− mice with RF: no changes in the expression rhythms of Per1/2, Dbp and E4bp4 genes. Expression levels of clock genes were similar to those of Dbh+/− mice. Treatment with α and β adrenergic receptor antagonists: no changes in the rhythmicity of Per1, Dbp, E4bp4, and Bmal1 genes in both genotypes. | Dbh−/− mice with RF: no changes in the expression rhythms of Per1, Per2, Dbp and E4bp4 genes (aorta and liver). Treatment with α and β adrenergic receptor antagonists: no changes in the rhythmicity of Per1, Dbp, E4bp4, and Bmal1 genes (aorta, liver, and BAT) in both genotypes. | (N) |
| High-fat, high-sucrose diet | Oishi et al., 2009a [26] | Per2/Tg mice fed ND: oscillation in Per2 mRNA expression levels in the light phase. Per2/Tg mice with HFSD: overexpression of Per2 mRNA. | Per2/Tg mice with ND or HFSD: overexpression of Per2 mRNA and attenuation of Bmal1 mRNA expression (liver and adipose tissue). | (N) |
| Ketogenic diet | Oishi et al., 2009b [32] | Jcl:ICR mice with KD: advancing acrophase expression of Per2 and Dbp genes. | Jcl:ICR mice with KD: advancing acrophase expression of Per2 and Dbp genes (liver, kidney, and adipose tissues). | (−) |
| High-fat diet | Hou et al., 2009 [24] | WT mice fed regular diet: higher Bmal1 and Per2 mRNA expression levels in CT20 and CT8, respectively. Lower levels in CT8 and CT20, respectively. ApoE−/− mice fed HFD: higher levels of Bmal1 and Per2 mRNA in CT0 and CT12, respectively. Lower levels in CT12 and CT0, respectively. No changes in Cry1 mRNA expression levels in both genotypes on regular diet. | WT mice fed regular diet: higher levels of Bmal1, Cry1, and Per2 mRNA expression in CT20, CT8, and CT12, respectively. ApoE−/− mice fed HFD: higher levels of Bmal1, Per2, and Cry1 mRNA expression in CT12. | (−) |
| High-fat diet | Tsai et al., 2010 [27] | † | † | † |
| Ketogenic diet | Oishi et al., 2010 [25] | Pparα-null and WT mice with KD: phase-advanced circadian expression for Bmal1 and low average levels of Dbp expression. | The expression of Bmal1 was phase-advanced in the liver. | (−) |
| Restricted feeding | Bray et al., 2013 [20] | WT mice with RF: altered expression of clock genes (Bmal1, Per2, Cry2, and Rev-Erbα) and clock-controlled genes (Dbp); lower and less consistent effects on the phases of clock component and output gene oscillations; average phase shifts and repression of amplitude were 3.90 ± 0.83 h and 54 ± 5%, respectively. | WT mice with RF: changes in expression of clock genes (Bmal1, Per2, Cry2, and Rev-erbα) and clock-controlled genes (Dbp); dramatic phase shifts in gene expression of clock components and output genes (liver); phase differences within the range of 6 and 11 h (mean 8.38 ± 0.84 h) (liver); smaller and less consistent effects on phase shifts of clock and output gene components (epididymal fat and gastrocnemius muscle); average phase shifts and repression of amplitude were 6.88 ± 2.06 h and 69 ± 3% (epididymal), 3.46 ± 1.41 h and 24 ± 20% (gastrocnemius muscle), respectively. | (−) |
| High-fat diet | Wang et al., 2015 [28] | P17 pups from HFD-fed dams: higher mRNA levels of Bmal1 in ZT1 with a circadian pattern and oscillates in antiphase for the Per2 gene. P35 pups from HFD-fed dams: improved phase changes while maintaining amplitude defects. | P17 pups from HFD-fed dams: Fas exhibited a rhythmic expression pattern in control animals, which peaked at ZT9, the late stage of the light phase. Pgc-1α exhibited a significantly rhythmic and lower expression (liver). P35 pups from HFD-fed dams: abolished the circadian expression rhythm of Fas and Pgc-1α (liver). | (−) |
| Restricted feeding | Noyan et al., 2015 [22] | WT mice under short-term CR: modulation in the mRNA profiles of pre-MI genes associated with the circadian clock. | WT mice under short-term CR: global change in gene expression associated with oxidative stress, immune function, apoptosis, metabolism angiogenesis, cytoskeleton and extracellular matrix. | (+) |
| Restricted feeding | Mukherji et al., 2015 [21] | WT mice with RF 8 (day 8): altered the expression of Rev-Erbα, Per1, and Per2 genes. | WT mice with RF1 (day 1): increased expression of Per1, Per2, and Rev-Erbα genes; no changes in expression of Bmal1, Cry1, and E4bp4 (liver, IECs, and pancreas). RF2 (day 2): changes in expression of clock components (Per1, Per2, Rev-Erbα, Bmal1, Cry1 e E4bp4); no recruitment of Bmal1 to the E-box region as a result of Rev-Erbα repression. After 4, 8, 15, 30, and 90 days of RF: change in expression levels of Per2, Per3, and Rev-Erbα genes (liver and intestinal epithelial cells). Glucose administration did not affect the expression pattern of Per1, Per2, and Rev-Erbα genes (liver, intestine, and pancreas). | (−) |
| High-fat, high-sucrose diet | Oishi et al., 2017 [29] | WT mice with RF: no changes in clock gene expression in both groups. Exogenous insulin administration: significantly increased the expression of the clock genes, Per1 and Per2. | WT mice with RF in NF: phase advancement of Per2 and Dbp genes (liver). Exogenous insulin administration: increased expression levels of Per1 and Per2 genes (liver, lung, WAT, and skeletal muscle). | (N) |
| High-fiber diet | Marques et al., 2017 [33] | WT mice fed high-fiber diet: upregulation of circadian rhythm in the cardiac transcriptome (q = 0.021). WT mice supplemented with acetate: upregulation of circadian rhythm in the cardiac transcriptome (q = 0.086). | WT mice fed high-fiber diet: upregulation of circadian rhythm in the renal transcriptome. WT mice supplemented with acetate: upregulation of circadian rhythm in the renal transcriptome. | (+) |
| Low phosphate diet | Noguchi et al., 2018 [34] | Mice with hypophosphatemia diet: higher expression levels of Per1, Per2, Per3, and Bmal1 genes in ZT9, ZT9, Zt16-5, and ZT21, respectively. Mice with control diet: higher expression levels of Per1, Per2, Per3, Bmal1, and Cry1 genes in ZT12, ZT15, ZT9-18, ZT24, and ZT15-24, respectively. | Mice fed hypophosphatemic diet: higher expression levels of Per2, Bmal1, and Cry genes in ZT6-12, ZT6-9, ZT18-ZT24, and ZT15-21 (callus and growth plate tissues) Mice fed control diet: higher expression levels of Per1, Per2, Per3, Bmal1, and Cry genes in ZT12, ZT15, ZT15, ZT21, and ZT15-21 (callus and growth plate tissues) | (−) |
| High-fat diet | Reitz et al., 2020 [30] | The circadian mechanism is involved in the transcriptional responses of oxidative stress and antioxidant pathways. | † | (N) |
| High-fat diet | Mia et al., 2021 [31] | WT mice with HFD: no changes in circadian clock components (Bmal1, Clock, Npas2, Nr1d1, Nr1d2, Per2, Per3, and Cry2). | † | (N) |
| Restricting feeding | Xin et al., 2021 [23] | Female mice on RF for 36 days: phase-locked to LD cycles. Female mice on LL cycle for 9 days: reversed the phase of clock genes. Cardiac transcriptome showed resistance in phase drag by reversed feeding and the fatty acid rhythm was entrained. | Female mice on RF for 36 days: phase shift of clock genes similar to animals on 7 days RF (liver and adipose tissue). Female mice on LL cycle for 9 days: increased the behavioral rhythm by 1.7 h, and did not change the phase of clock genes (liver). Induced oscillations in clock genes (adipose tissue); reversed the phase of clock genes (kidney). The liver and adipose tissue transcriptomes were entrained by the reversed feeding. The kidney transcriptome is more resistant to phase drag by reversed feeding. | (−) |
| High-biotin diet | Murata et al., 2021 [35] | BMAL1-BioID mice with biotin-rich diet: reduced endogenous BMAL1 expression. | BMAL1-BioID mice on biotin-rich diet: reduced endogenous BMAL1 expression. BMAL1-BioID mice on biotin-rich diet: reduced endogenous BMAL1 expression (liver and kidney) and showed endogenous CLOCK biotinylation in a diet-dependent manner (liver and brain). | (N) |
| Low BCAA diet | Latimer et al., 2021 [36] | CBK mice: time-of-day fluctuations in mRNA levels of Arntl, Nr1d1, and Dbp were attenuated (59%–69% reduction in amplitude). | † | (N) |
| Diet Interventions/Diet | Author/Year | Metabolic Changes | Positive (+), Negative (−), or Neutral (N) Effect of the Diet/Dietary Intervention |
|---|---|---|---|
| Restricting feeding | Goh et al., 2007 [18] | RF reduced food intake. In WT mice under conditions ad libitum, the Pparα mRNA levels showed a statistically significant circadian profile. | (+) |
| Restricting feeding | Reilly et al., 2008 [19] | † | † |
| High-fat, high-sucrose diet | Oishi et al., 2009a [26] | WT and Per2/Tg mice with HFSD: increased body weight, plasma insulin, total cholesterol, and insulin/glucose ratio. PAI-1 mRNA expression levels did not differ. Per2/Tg with HFSD or ND mice: dampening of PAI-1 expression rhythms in the heart. | (−) |
| Ketogenic diet | Oishi et al., 2009b [32] | Jcl:ICR mice with KD: reduced body weight and glucose levels, increased plasma levels of FFA and total ketone bodies, increased PAI-1 mRNA levels (heart and liver) with advanced acrophase to 4.7 h (heart), 7.9 h (kidney), and 7.8 h (epididymal fat), and led to phase advancement of the endogenous circadian clock that governs rhythmic behavior. | (-) |
| High-fat diet | Hou et al., 2009 [24] | ApoE−/− mice fed HFD: increased total and LDL-c cholesterol levels, decreased HDL-c levels, with peak concentrations in CT12, led to atherosclerotic plaque formation in the aortic root, increased diurnal expression levels of lipid metabolism-related transcription factors Pparα and Rxrα. | (−) |
| High-fat diet | Tsai et al., 2010 [27] | CCM mice fed HFD: increased caloric intake; body weight gain with increased body fat percentage; altered metabolism gene expression, cardiac triglyceride and lipid species. CCM mice fed standard diet: attenuation in gene and protein expression of myocardial enzymes related to triglyceride metabolism. HFD at the end of the active phase: increased levels of triglyceride, lipid and fatty acid metabolism genes (Dgat2, Agpat3, Hsl, S3-12, Ucp3 and Pdk4), and in the rate of substrate utilization. | (−) |
| Ketogenic diet | Oishi et al., 2010 [25] | Pparα-null mice fed KD: decreased BW, plasma FFA levels, and total ketone bodies; induced fatty liver; and increased hepatic total cholesterol levels. WT mice fed KD: increased circadian expression of PAI-1 mRNA (heart and liver). Benzafibrate: induced the expression of the Pai-1, Cy4A10, and Fgf21 genes in a PPARα-dependent manner. | (−) |
| Restricted feeding | Bray et al., 2013 [20] | WT mice on RF: changes in whole-body energy balance, higher food intake during DP, little influence on physical activity rhythms, diurnal variation in plasma glucose and triglyceride levels, body weight gain, and tissue-specific changes in metabolic genes (Accα, Glut2, Lpk, Lgs, Mcad, Dgat2, Acsl1, Atgl, Lipe, Mcp1, Glut4, Pdk4). | (−) |
| High-fat diet | Wang et al., 2015 [28] | Pregnant female mice fed HFD: hypercholesterolemia and hyperlipidemia. Pups of obese dams: expressed inflammatory cytokines (Tnfα and Il-6), showed an anti-phase pattern for Pparα and Cpt1b genes. | (−) |
| Restricted feeding | Noyan et al., 2015 [22] | WT mice under short-term CR: smaller infarct size and apoptosis, improved cardiac function at 2 d post-MI and survival and weight loss, and modulated signaling pathways associated with cardio-protection, mitochondrial function, and biogenesis. | (+) |
| Restricted feeding | Mukherji et al., 2015 [21] | WT mice with RF: increased ketone bodies, FFA, glucagon and corticosterone; decreased insulin and glucose levels stimulated the expression of Pparα, Creb, and Rev-Erbα, increased GSK3β activity. Glucose administration prevented plasma glucose reduction; increased levels of FFA, ketone bodies, corticosterone, FGF21; and inhibited transcription of Pparα and Rev-Erbα. | (−) |
| High-fat, high-sucrose diet | Oishi et al., 2017 [29] | WT mice with DRF: increased plasma insulin and GLP-1 concentration; did not affect plasma corticosterone and GIP levels. Exogenous insulin administration: induced Akt phosphorylation (heart, liver, and skeletal muscle). | (+) |
| High-fiber diet | Marques et al., 2017 [33] | WT mice with high-fiber diet: altered the composition of the gut microbiota and increased acetate levels. WT mice supplemented with acetate: altered the composition of the intestinal microbiota and increased the percentage of acetate-producing bacteria. WT mice with high-fiber diet and acetate: reduced systolic and diastolic BP levels, altered the renal transcriptome (Rasal1, Cyp414, and Cck) of genes related to renal fibrosis, fluid absorption through sodium channel regulation, anti-inflammatory action, and the cardiac transcriptome (Tcap and Timp4) of genes related to cardiac diseases, pathways acting on cell cycle, replication, translation, mRNA metabolism, respiratory electron chain, mitogen-activated protein kinase signaling, and renin-angiotensin system. | (+) |
| Low-phosphate diet | Noguchi et al., 2018 [34] | Mice with hypophosphatemic diet: increased total cartilage volume fraction, reduced TMD and BV/TV, increased osteochondromic progenitor lineage and impaired chondrocytes, reduced the size of proliferative matrix-forming cells, and affected systemic regulation of mineral metabolism. Transcriptome analyses revealed that 1.879 genes associated with diet and having a circadian pattern of regulation, those with mitochondrial function, including oxidative metabolism and canonical regulatory pathways associated with apoptotic signaling. | (−) |
| High-fat diet | Reitz et al., 2020 [30] | ClockΔ19/19 mice fed HFD or SC: hypercholesterolemia, hyperglycemia, hyperinsulinemia, increased body weight and epididymal white adipose tissue, no changes in cardiomyocyte hypertrophy, normal cardiac function, and structure. No changes in the expression of genes related to cardiac remodeling and oxidative stress (Nampt, Sirt1, Sirt2, Sirt3, Sirt4, Sirt6, Foxo1, and Foxo3). Increased concentrations of CAT and GPx proteins, and Pparγ gene transcripts. | (N) |
| High-fat diet | Mia et al., 2020 [31] | HFD increased body weight, adiposity, daily energy expenditure, reduced physical activity, and RER, increased stroke volume, left posterior ventricular wall thickness during systole, the BVW/TL ratio, cardiomyocyte area, cardiac fibrosis, and altered the temporal regulation of the cardiac transcriptome, especially of metabolism-related genes. Day–night differences affected cardiac glucose oxidation, lactate release, and the cardiac lipidome. The diurnal rhythms of lipid metabolism genes (Cd36, Mcd, Lcad, and Lipe) and plasma levels were altered. Time-of-day-restricted feeding restored body metabolic rhythms, normalized the adverse effects of cardiac remodeling (BVW/TL ratio, cardiomyocyte size, cardiac fibrosis, cardiac steatosis), and increased the day–night difference in cardiac lipid metabolism. | (−) |
| Restricting feeding | Xin et al., 2021 [23] | There was no change in body and adipose tissue weight after 1 week of RF. | (N) |
| High-biotin diet | Murata et al., 2021 [35] | A 0.5% biotin diet caused protein biotinylation in the brain, heart, testis, and liver of Bmal1-BioID mice. | (+) |
| Low BCAA diet | Latimer et al., 2021 [36] | Mice fed low BCAA diet: decreased body mass, adiposity, and modest cardiac atrophy. Consumption of dietary BCAAs at the end of the active period (dark): increased the cardiomyocyte size, promoted cardiac protein synthesis, and highly dynamic cardiac growth. | (+) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paula, A.B.R.; Resende, L.T.; Jardim, I.A.B.A.; Coelho, B.I.C.; de Coutinho Miranda, D.; Portes, A.M.O.; Teles, M.C.; Castrucci, A.M.d.L.; Isoldi, M.C. The Effect of Diet on the Cardiac Circadian Clock in Mice: A Systematic Review. Metabolites 2022, 12, 1273. https://doi.org/10.3390/metabo12121273
Paula ABR, Resende LT, Jardim IABA, Coelho BIC, de Coutinho Miranda D, Portes AMO, Teles MC, Castrucci AMdL, Isoldi MC. The Effect of Diet on the Cardiac Circadian Clock in Mice: A Systematic Review. Metabolites. 2022; 12(12):1273. https://doi.org/10.3390/metabo12121273
Chicago/Turabian StylePaula, Ana Beatriz Rezende, Letícia Teresinha Resende, Isabela Alcântara Barretto Araújo Jardim, Bianca Iara Campos Coelho, Denise de Coutinho Miranda, Alexandre Martins Oliveira Portes, Maria Cecília Teles, Ana Maria de Lauro Castrucci, and Mauro César Isoldi. 2022. "The Effect of Diet on the Cardiac Circadian Clock in Mice: A Systematic Review" Metabolites 12, no. 12: 1273. https://doi.org/10.3390/metabo12121273
APA StylePaula, A. B. R., Resende, L. T., Jardim, I. A. B. A., Coelho, B. I. C., de Coutinho Miranda, D., Portes, A. M. O., Teles, M. C., Castrucci, A. M. d. L., & Isoldi, M. C. (2022). The Effect of Diet on the Cardiac Circadian Clock in Mice: A Systematic Review. Metabolites, 12(12), 1273. https://doi.org/10.3390/metabo12121273