


Chronometabolism: The Timing of the Consumption of Meals Has a Greater Influence Than Glycemic Index (GI) on the Postprandial Metabolome


 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Blood Sample Collection
2.3. Sample Measurement
2.4. Data Generation and Quality Control
2.5. Statistical Analysis
3. Results
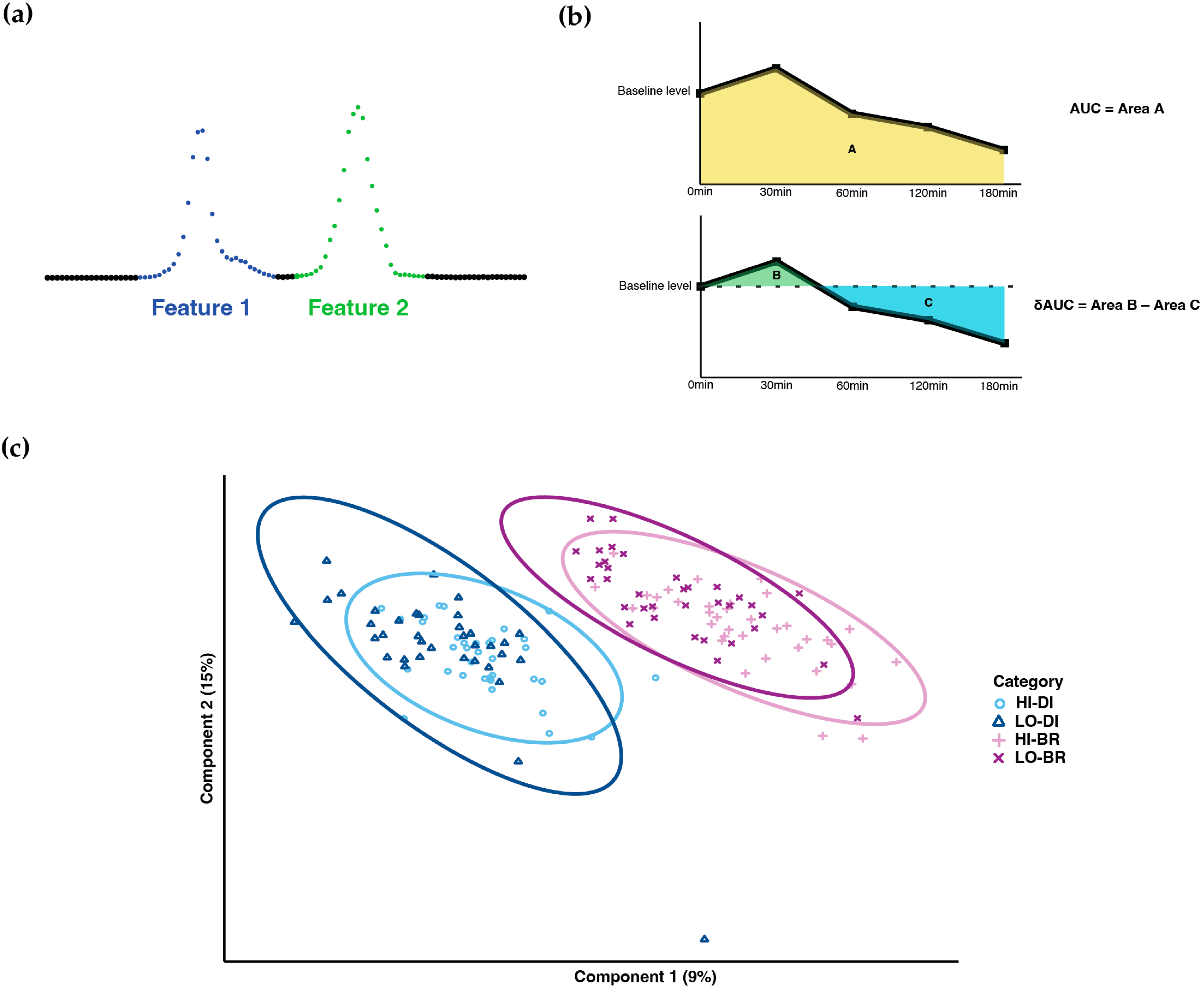
3.1. Metabolome Profiling
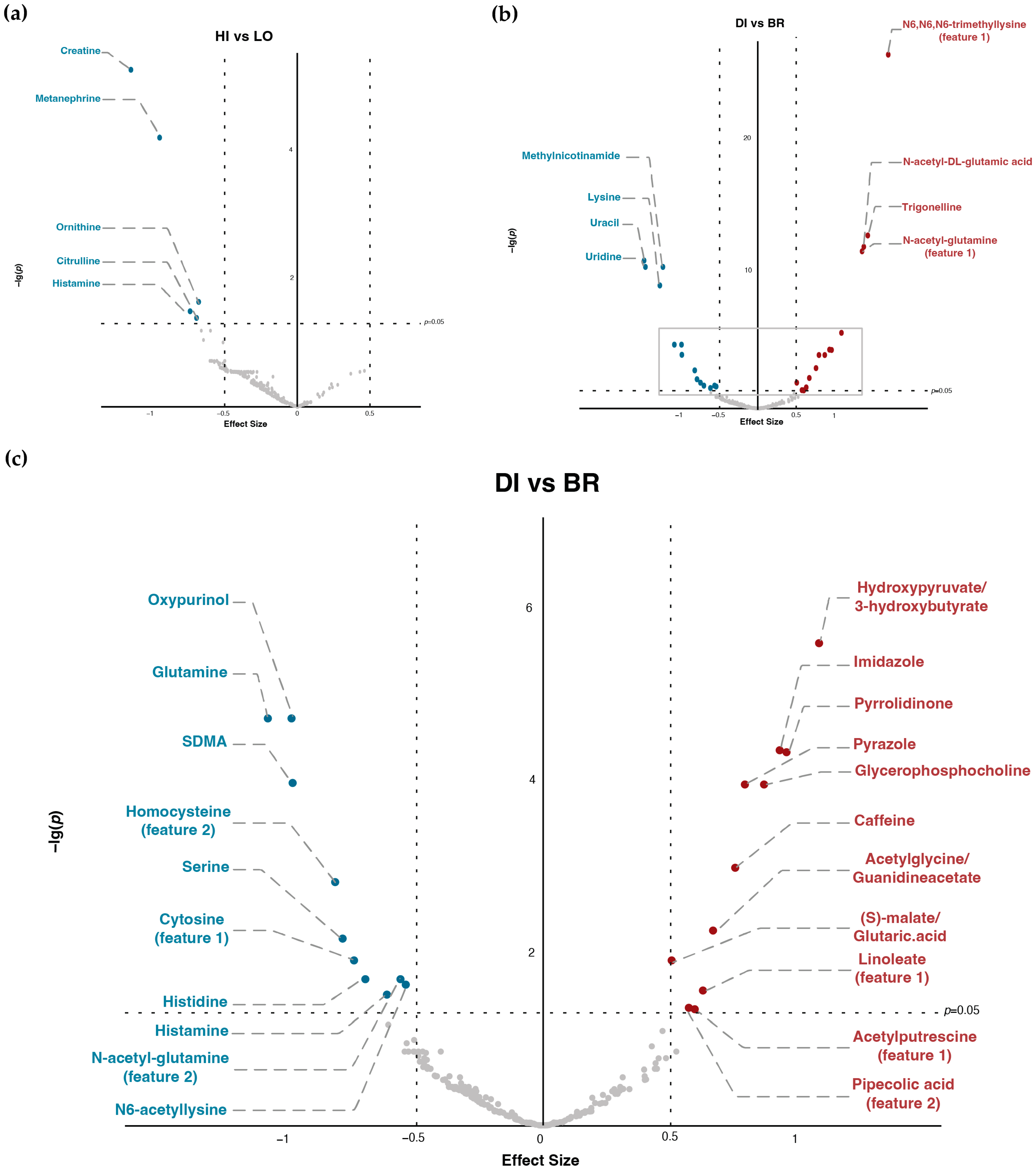
3.2. Differences in Metabolites and Related Pathway under Different GI Content and TM Timing
3.3. Top Metabolites and Pathways Identified by (δ)AUC Difference of Overall Comparison
4. Discussion
4.1. Novelty
4.2. Future Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blaak, E.E.; Antoine, J.-M.; Benton, D.; Björck, I.; Bozzetto, L.; Brouns, F.; Diamant, M.; Dye, L.; Hulshof, T.; Holst, J.J.; et al. Impact of postprandial glycaemia on health and prevention of disease. Obes. Rev. 2012, 13, 923–984. [Google Scholar] [CrossRef] [PubMed]
- Barclay, A.W.; Augustin, L.S.A.; Brighenti, F.; Delport, E.; Henry, C.J.; Sievenpiper, J.L.; Usic, K.; Yuexin, Y.; Zurbau, A.; Wolever, T.M.S.; et al. Dietary Glycaemic Index Labelling: A Global Perspective. Nutrients 2021, 13, 3244. [Google Scholar] [CrossRef] [PubMed]
- Ighodaro, O.M. Molecular pathways associated with oxidative stress in diabetes mellitus. Biomed. Pharmacother. 2018, 108, 656–662. [Google Scholar] [CrossRef]
- Yoon, S.; Eom, G.H.; Kang, G. Nitrosative Stress and Human Disease: Therapeutic Potential of Denitrosylation. Int. J. Mol. Sci. 2021, 22, 9794. [Google Scholar] [CrossRef] [PubMed]
- Hurrle, S.; Hsu, W.H. The etiology of oxidative stress in insulin resistance. Biomed. J. 2017, 40, 257–262. [Google Scholar] [CrossRef]
- Yong, Y.N.; Henry, C.J.; Haldar, S. Is There a Utility of Chrono-Specific Diets in Improving Cardiometabolic Health? Mol. Nutr. Food Res. 2022, 66, 2200043. [Google Scholar] [CrossRef] [PubMed]
- Skene, D.J.; Skornyakov, E.; Chowdhury, N.R.; Gajula, R.P.; Middleton, B.; Satterfield, B.C.; Porter, K.I.; Van Dongen, H.P.A.; Gaddameedhi, S. Separation of circadian- and behavior-driven metabolite rhythms in humans provides a window on peripheral oscillators and metabolism. Proc. Natl. Acad. Sci. USA 2018, 115, 7825–7830. [Google Scholar] [CrossRef]
- Davies, S.K.; Ang, J.E.; Revell, V.L.; Holmes, B.; Mann, A.; Robertson, F.P.; Cui, N.; Middleton, B.; Ackermann, K.; Kayser, M.; et al. Effect of sleep deprivation on the human metabolome. Proc. Natl. Acad. Sci. USA 2014, 111, 10761–10766. [Google Scholar] [CrossRef]
- Sato, T.; Greco, C.M. Expanding the link between circadian rhythms and redox metabolism of epigenetic control. Free. Radic. Biol. Med. 2021, 170, 50–58. [Google Scholar] [CrossRef]
- Sutton, E.F.; Beyl, R.; Early, K.S.; Cefalu, W.T.; Ravussin, E.; Peterson, C.M. Early Time-Restricted Feeding Improves Insulin Sensitivity, Blood Pressure, and Oxidative Stress Even without Weight Loss in Men with Prediabetes. Cell Metab. 2018, 27, 1212–1221.e1213. [Google Scholar] [CrossRef]
- Dyar, K.A.; Eckel-Mahan, K.L. Circadian Metabolomics in Time and Space. Front. Neurosci. 2017, 11, 369. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Parr, E.B.; Devlin, B.L.; Hawley, J.A.; Sassone-Corsi, P. Human metabolomics reveal daily variations under nutritional challenges specific to serum and skeletal muscle. Mol. Metab. 2018, 16, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kinouchi, K.; Mikami, Y.; Kanai, T.; Itoh, H. Circadian rhythms in the tissue-specificity from metabolism to immunity: Insights from omics studies. Mol. Asp. Med. 2021, 80, 100984. [Google Scholar] [CrossRef] [PubMed]
- Haldar, S.; Egli, L.; De Castro, C.A.; Tay, S.L.; Koh, M.X.N.; Darimont, C.; Mace, K.; Henry, C.J. High or low glycemic index (GI) meals at dinner results in greater postprandial glycemia compared with breakfast: A randomized controlled trial. BMJ Open Diabetes Res. Care 2020, 8, e001099. [Google Scholar] [CrossRef] [PubMed]
- Teo, G.; Chew, W.S.; Burla, B.J.; Herr, D.; Tai, E.S.; Wenk, M.R.; Torta, F.; Choi, H. MRMkit: Automated Data Processing for Large-Scale Targeted Metabolomics Analysis. Anal. Chem. 2020, 92, 13677–13682. [Google Scholar] [CrossRef] [PubMed]
- Broadhurst, D.; Goodacre, R.; Reinke, S.N.; Kuligowski, J.; Wilson, I.D.; Lewis, M.R.; Dunn, W.B. Guidelines and considerations for the use of system suitability and quality control samples in mass spectrometry assays applied in untargeted clinical metabolomic studies. Metabolomics 2018, 14, 72. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Wunderlich, C.; Schober, K.; Lange, S.A.; Drab, M.; Braun-Dullaeus, R.C.; Kasper, M.; Schwencke, C.; Schmeisser, A.; Strasser, R.H. Disruption of caveolin-1 leads to enhanced nitrosative stress and severe systolic and diastolic heart failure. Biochem. Biophys. Res. Commun. 2006, 340, 702–708. [Google Scholar] [CrossRef]
- Wang, F.; Yuan, Q.; Chen, F.; Pang, J.; Pan, C.; Xu, F.; Chen, Y. Fundamental Mechanisms of the Cell Death Caused by Nitrosative Stress. Front. Cell Dev. Biol. 2021, 9, 742483. [Google Scholar] [CrossRef]
- Manders, R.J.; Little, J.P.; Forbes, S.C.; Candow, D.G. Insulinotropic and Muscle Protein Synthetic Effects of Branched-Chain Amino Acids: Potential Therapy for Type 2 Diabetes and Sarcopenia. Nutrients 2012, 4, 1664–1678. [Google Scholar] [CrossRef]
- Takahashi, M.; Ozaki, M.; Kang, M.-I.; Sasaki, H.; Fukazawa, M.; Iwakami, T.; Lim, P.J.; Kim, H.-K.; Aoyama, S.; Shibata, S. Effects of Meal Timing on Postprandial Glucose Metabolism and Blood Metabolites in Healthy Adults. Nutrients 2018, 10, 1763. [Google Scholar] [CrossRef] [PubMed]
- Isherwood, C.M.; Van der Veen, D.R.; Johnston, J.D.; Skene, D.J. Twenty-four-hour rhythmicity of circulating metabolites: Effect of body mass and type 2 diabetes. FASEB J. 2017, 31, 5557–5567. [Google Scholar] [CrossRef] [PubMed]
- Fleszar, M.G.; Wiśniewski, J.; Zboch, M.; Diakowska, D.; Gamian, A.; Krzystek-Korpacka, M. Targeted metabolomic analysis of nitric oxide/L-arginine pathway metabolites in dementia: Association with pathology, severity, and structural brain changes. Sci. Rep. 2019, 9, 13764. [Google Scholar] [CrossRef]
- Ardigo, D.; Stüehlinger, M.; Franzini, L.; Valtueña, S.; Piatti, P.M.; Pachinger, O.; Reaven, G.M.; Zavaroni, I. ADMA is independently related to flow-mediated vasodilation in subjects at low cardiovascular risk. Eur. J. Clin. Investig. 2007, 37, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Millar-Craig, M.; Bishop, C.; Raftery, E.B. Circadian Variation of Blood-Pressure. Lancet 1978, 311, 795–797. [Google Scholar] [CrossRef]
- Davis, R.; Murgia, C.; Dordevic, A.L.; Bonham, M.P.; Huggins, C.E. Diurnal variation in gene expression of human peripheral blood mononuclear cells after eating a standard meal compared with a high protein meal: A cross-over study. Clin. Nutr. 2021, 40, 4349–4359. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Torres, I.; Manzano-Pech, L.; Rubio-Ruíz, M.E.; Soto, M.E.; Guarner-Lans, V. Nitrosative Stress and Its Association with Cardiometabolic Disorders. Molecules 2020, 25, 2555. [Google Scholar] [CrossRef]
- Wilking, M.; Ndiaye, M.; Mukhtar, H.; Ahmad, N. Circadian rhythm connections to oxidative stress: Implications for human health. Antioxid. Redox. Signal 2013, 19, 192–208. [Google Scholar] [CrossRef]
- Mariotti, F. Animal and Plant Protein Sources and Cardiometabolic Health. Adv. Nutr. 2019, 10, S351–S366. [Google Scholar] [CrossRef]
- Tricò, D.; Frascerra, S.; Baldi, S.; Mengozzi, A.; Nesti, L.; Mari, A.; Natali, A. The insulinotropic effect of a high-protein nutrient preload is mediated by the increase of plasma amino acids in type 2 diabetes. Eur. J. Nutr. 2019, 58, 2253–2261. [Google Scholar] [CrossRef]
- Matés, J.M.; Pérez-Gómez, C.; de Castro, I.N.; Asenjo, M.; Márquez, J. Glutamine and its relationship with intracellular redox status, oxidative stress and cell proliferation/death. Int. J. Biochem. Cell. Biol. 2002, 34, 439–458. [Google Scholar] [CrossRef] [PubMed]
- Ch, R.; Chevallier, O.; Elliott, C.T. Metabolomics reveal circadian control of cellular metabolism. Trends Analyt. Chem. 2020, 130, 115986. [Google Scholar] [CrossRef]
- Badoud, F.; Lam, K.P.; DiBattista, A.; Perreault, M.; Zulyniak, M.A.; Cattrysse, B.; Stephenson, S.; Britz-McKibbin, P.; Mutch, D.M. Serum and Adipose Tissue Amino Acid Homeostasis in the Metabolically Healthy Obese. J. Proteome Res. 2014, 13, 3455–3466. [Google Scholar] [CrossRef] [PubMed]
- Gar, C.; Rottenkolber, M.; Prehn, C.; Adamski, J.; Seissler, J.; Lechner, A. Serum and plasma amino acids as markers of prediabetes, insulin resistance, and incident diabetes. Crit. Rev. Clin. Lab. Sci. 2018, 55, 21–32. [Google Scholar] [CrossRef]
- Clarke, H.; Hickner, R.C.; Ormsbee, M.J. The Potential Role of Creatine in Vascular Health. Nutrients 2021, 13, 857. [Google Scholar] [CrossRef]
- Alqudah, A.; Wedyan, M.; Qnais, E.; Jawarneh, H.; McClements, L. Plasma Amino Acids Metabolomics’ Important in Glucose Management in Type 2 Diabetes. Front. Pharmacol. 2021, 12, 695418. [Google Scholar] [CrossRef]
- Cao, Y.-F.; Li, J.; Zhang, Z.; Liu, J.; Sun, X.-Y.; Feng, X.-F.; Luo, H.-H.; Yang, W.; Li, S.-N.; Yang, X.; et al. Plasma Levels of Amino Acids Related to Urea Cycle and Risk of Type 2 Diabetes Mellitus in Chinese Adults. Front. Endocrinol. 2019, 10, 50. [Google Scholar] [CrossRef]
- Fiehn, O.; Garvey, W.T.; Newman, J.W.; Lok, K.H.; Hoppel, C.L.; Adams, S.H. Plasma Metabolomic Profiles Reflective of Glucose Homeostasis in Non-Diabetic and Type 2 Diabetic Obese African-American Women. PLoS ONE 2010, 5, e15234. [Google Scholar] [CrossRef]
- Bjørnestad, E.Ø.; Dhar, I.; Svingen, G.F.T.; Pedersen, E.R.; Svenningsson, M.M.; Tell, G.S.; Ueland, P.M.; Ørn, S.; Sulo, G.; Laaksonen, R.; et al. Trimethyllysine predicts all-cause and cardiovascular mortality in community-dwelling adults and patients with coronary heart disease. Eur. Heart J. 2021, 1, oeab007. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, M.; Wang, A.; Xue, J.; Cheng, S.; Cheng, A.; Gao, J.; Zhang, Q.; Zhan, R.; Meng, X.; et al. Association between Plasma Trimethyllysine and Prognosis of Patients with Ischemic Stroke. J. Am. Heart Assoc. 2021, 10, e020979. [Google Scholar] [CrossRef]
- Li, X.S.; Wang, Z.; Cajka, T.; Buffa, J.A.; Nemet, I.; Hurd, A.G.; Gu, X.; Skye, S.M.; Roberts, A.B.; Wu, Y.; et al. Untargeted metabolomics identifies trimethyllysine, a TMAO-producing nutrient precursor, as a predictor of incident cardiovascular disease risk. JCI Insight 2018, 3, e99096. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.; Leong, J.; Jialal, I. Amino acid levels in nascent metabolic syndrome: A contributor to the pro-inflammatory burden. J. Diabetes Complicat. 2018, 32, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, Y.; Hao, F.; Zhou, X.; Han, X.; Tang, H.; Ji, L. Human Serum Metabonomic Analysis Reveals Progression Axes for Glucose Intolerance and Insulin Resistance Statuses. J. Proteome Res. 2009, 8, 5188–5195. [Google Scholar] [CrossRef] [PubMed]
- Bulló, M.; Papandreou, C.; Ruiz-Canela, M.; Guasch-Ferré, M.; Li, J.; Hernández-Alonso, P.; Toledo, E.; Liang, L.; Razquin, C.; Corella, D.; et al. Plasma Metabolomic Profiles of Glycemic Index, Glycemic Load, and Carbohydrate Quality Index in the PREDIMED Study. J. Nutr. 2020, 151, 50–58. [Google Scholar] [CrossRef]
- Lépine, G.; Tremblay-Franco, M.; Bouder, S.; Dimina, L.; Fouillet, H.; Mariotti, F.; Polakof, S. Investigating the Postprandial Metabolome after Challenge Tests to Assess Metabolic Flexibility and Dysregulations Associated with Cardiometabolic Diseases. Nutrients 2022, 14, 472. [Google Scholar] [CrossRef]
- Diederichs, T.; Herder, C.; Roßbach, S.; Roden, M.; Wudy, S.A.; Nöthlings, U.; Alexy, U.; Buyken, A.E. Carbohydrates from Sources with a Higher Glycemic Index during Adolescence: Is Evening Rather than Morning Intake Relevant for Risk Markers of Type 2 Diabetes in Young Adulthood? Nutrients 2017, 9, 591. [Google Scholar] [CrossRef]
- Escobar-Martínez, I.; Arreaza-Gil, V.; Muguerza, B.; Arola-Arnal, A.; Bravo, F.I.; Torres-Fuentes, C.; Suárez, M. Administration Time Significantly Affects Plasma Bioavailability of Grape Seed Proanthocyanidins Extract in Healthy and Obese Fischer 344 Rats. Mol. Nutr. Food Res. 2022, 66, 2100552. [Google Scholar] [CrossRef]
- Hironao, K.-Y.; Ashida, H.; Yamashita, Y. The cacao procyanidin extract-caused anti-hyperglycemic effect was changed by the administration timings. J. Clin. Biochem. Nutr. 2020, 67, 61–66. [Google Scholar] [CrossRef]
- Takahashi, M.; Ozaki, M.; Miyashita, M.; Fukazawa, M.; Nakaoka, T.; Wakisaka, T.; Matsui, Y.; Hibi, M.; Osaki, N.; Shibata, S. Effects of timing of acute catechin-rich green tea ingestion on postprandial glucose metabolism in healthy men. J. Nutr. Biochem. 2019, 73, 108221. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Metabolite | Effect Size 1 | p 2 | Relevant Metabolic Pathway |
|---|---|---|---|
| AUC | |||
| N6,N6,N6-trimethyllysine (feature 1) | 1.70 | 7.08 × 10−27 *** | Lysine metabolism, carnitine synthesis |
| Trigonelline | 1.44 | 1.58 × 10−13 *** | Tryptophan-nicotinamide metabolism |
| N-acetyl-DL-glutamic acid | 1.38 | 1.09 × 10−12 *** | Arginine-NO pathway |
| N-acetyl-glutamine (feature 1) | 1.36 | 2.34 × 10−12 *** | Arginine-NO pathway |
| Hydroxypyruvate/3-hydroxybutyrate | 1.09 | 2.43 × 10−6 *** | Serine metabolism/fatty acid biosynthesis, ketone metabolism |
| Pyrrolidinone | 0.96 | 4.48 × 10−5 *** | - |
| Imidazole | 0.94 | 4.25 × 10−5 *** | Histidine catabolism |
| Glycerophosphocholine | 0.87 | 1.06 × 10−4 *** | Retinol and choline metabolism |
| Pyrazole | 0.80 | 1.06 × 10−4 *** | - |
| Caffeine | 0.76 | 9.82 × 10−4 *** | Tryptophan-nicotinamide metabolism |
| Acetylglycine/Guanidineacetate | 0.67 | 5.25 × 10−3 ** | Guanidineacetate: arginine, proline, glycine, threonine and serine metabolism |
| Linoleate (feature 1) | 0.63 | 0.03 * | - |
| Acetylputrescine (feature 1) | 0.60 | 0.04 * | Arginine/proline metabolism |
| Pipecolic acid (feature 2) | 0.58 | 0.04 * | Lysine metabolism (by intestinal microflora) |
| (S)-malate/Glutaric acid | 0.51 | 0.01 * | Glucose metabolism/Glutamate, tryptophan and lysine metabolism |
| Ornithine | 0.47 | 0.08 | Arginine-NO pathway, urea cycle |
| 1-Methyladenosine | −0.51 | 0.10 | - |
| N6-acetyllysine | −0.54 | 0.02 * | Lysine metabolism |
| N-acetyl-glutamine (feature 2) | −0.56 | 0.02 * | Arginine-NO pathway |
| ADMA | −0.61 | 0.06 | Arginine-NO pathway |
| Histamine | −0.62 | 0.03 * | Histidine catabolism |
| Histidine | −0.70 | 0.02 * | Histidine metabolism |
| Cytosine (feature 1) | −0.75 | 0.01 * | Pyrimidine synthesis/salvage pathway |
| Serine | −0.79 | 6.51 × 10−3 ** | Purine and pyrimidine synthesis pathway |
| Homocysteine (feature 2) | −0.82 | 1.44 × 10−3 ** | Methionine catabolism |
| SDMA | −0.99 | 1.02 × 10−4 *** | Arginine-NO pathway |
| Oxypurinol | −1.00 | 1.82 × 10−5 *** | Purine synthesis/salvage pathway |
| Glutamine | −1.09 | 1.82 × 10−5 *** | Arginine-NO pathway |
| Methylnicotinamide | −1.24 | 3.32 × 10−11 *** | Tryptophan-nicotinamide metabolism |
| Lysine | −1.28 | 7.73 × 10−10 *** | Lysine metabolism, carnitine synthesis |
| Uracil | −1.47 | 3.32 × 10−11 *** | Pyrimidine synthesis/salvage pathway |
| Uridine | −1.48 | 1.12 × 10−11 *** | Pyrimidine synthesis/salvage pathway |
| δAUC 3 | |||
| Uridine | 0.73 | 0.08 | Pyrimidine synthesis/salvage pathway |
| Methylnicotinamide | −0.71 | 0.08 | Tryptophan-nicotinamide metabolism |
| Lysine | −0.72 | 0.08 | Lysine metabolism, carnitine synthesis |
| Creatine | −0.93 | 1.72 × 10−3 ** | Arginine-NO pathway |
| Ornithine | −1.12 | 1.52 × 10−5 *** | Arginine-NO pathway, urea cycle |
| Metabolite | Effect Size 1 | p 2 | Relevant Metabolic Pathway |
|---|---|---|---|
| AUC | |||
| Lysine | −0.51 | 0.10 | Lysine metabolism |
| Imidazole | −0.60 | 0.06 | Histidine catabolism |
| Trans-4-hydroxyproline (feature 1) | −0.65 | 0.09 | Collagen synthesis/degradation, cell signalling |
| Arginine | −0.66 | 0.06 | Arginine-NO pathway |
| Ornithine | −0.68 | 0.02 * | Arginine-NO pathway, urea cycle |
| Citrulline | −0.69 | 0.04 * | Arginine-NO pathway, urea cycle |
| Histamine | −0.74 | 0.03 * | Histidine catabolism |
| Metanephrine | −0.95 | 5.81 × 10−5 *** | Tyrosine metabolism |
| Creatine | −1.15 | 5.05 × 10−6 *** | Creatine metabolism |
| δAUC 3 | |||
| Creatine | −0.93 | 3.78 × 10−3 ** | Arginine-NO pathway |
| Ornithine | −0.77 | 0.02 * | Arginine-NO pathway, urea cycle |
| Metabolite Ratio 1 | Effect Size 2 | p 3 |
|---|---|---|
| Arginine:SDMA | 1.23 | 1.91 × 10−10 *** |
| Arginine:ADMA | 0.88 | 2.19 × 10−4 *** |
| Arginine:NG,NG-Dimethylarginine | 0.66 | 0.01 * |
| Arginine:Dimethylarginine | 0.58 | 0.04 * |
| Homoarginine:Lysine | 0.50 | 4.70 × 10−2 * |
| Glutamine:Glutamate | −0.93 | 7.61 × 10−5 *** |
| Glutamate:Acetylglutamine | −1.37 | 8.35 × 10−13 *** |
| Lysine:Arginine | −1.42 | 5.45 × 10−19 *** |
| Glutamate:Acetylglutamate | −1.47 | 2.37 × 10−14 *** |
| Glutamine:Acetylglutamate | −1.76 | 6.88 × 10−26 *** |
| Glutamine:Acetylglutamine | −1.77 | 3.11 × 10−27 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yong, Y.N.; Dong, J.; Pakkiri, L.S.; Henry, C.J.; Haldar, S.; Drum, C.L. Chronometabolism: The Timing of the Consumption of Meals Has a Greater Influence Than Glycemic Index (GI) on the Postprandial Metabolome. Metabolites 2023, 13, 490. https://doi.org/10.3390/metabo13040490
Yong YN, Dong J, Pakkiri LS, Henry CJ, Haldar S, Drum CL. Chronometabolism: The Timing of the Consumption of Meals Has a Greater Influence Than Glycemic Index (GI) on the Postprandial Metabolome. Metabolites. 2023; 13(4):490. https://doi.org/10.3390/metabo13040490
Chicago/Turabian StyleYong, Yi Ning, Jiangwen Dong, Leroy Sivappiragasam Pakkiri, Christiani Jeyakumar Henry, Sumanto Haldar, and Chester Lee Drum. 2023. "Chronometabolism: The Timing of the Consumption of Meals Has a Greater Influence Than Glycemic Index (GI) on the Postprandial Metabolome" Metabolites 13, no. 4: 490. https://doi.org/10.3390/metabo13040490
APA StyleYong, Y. N., Dong, J., Pakkiri, L. S., Henry, C. J., Haldar, S., & Drum, C. L. (2023). Chronometabolism: The Timing of the Consumption of Meals Has a Greater Influence Than Glycemic Index (GI) on the Postprandial Metabolome. Metabolites, 13(4), 490. https://doi.org/10.3390/metabo13040490