
Tolerant Larvae and Sensitive Juveniles: Integrating Metabolomics and Whole-Organism Responses to Define Life-Stage Specific Sensitivity to Ocean Acidification in the American Lobster

 , ,
, ,
Abstract
1. Introduction
2. Results
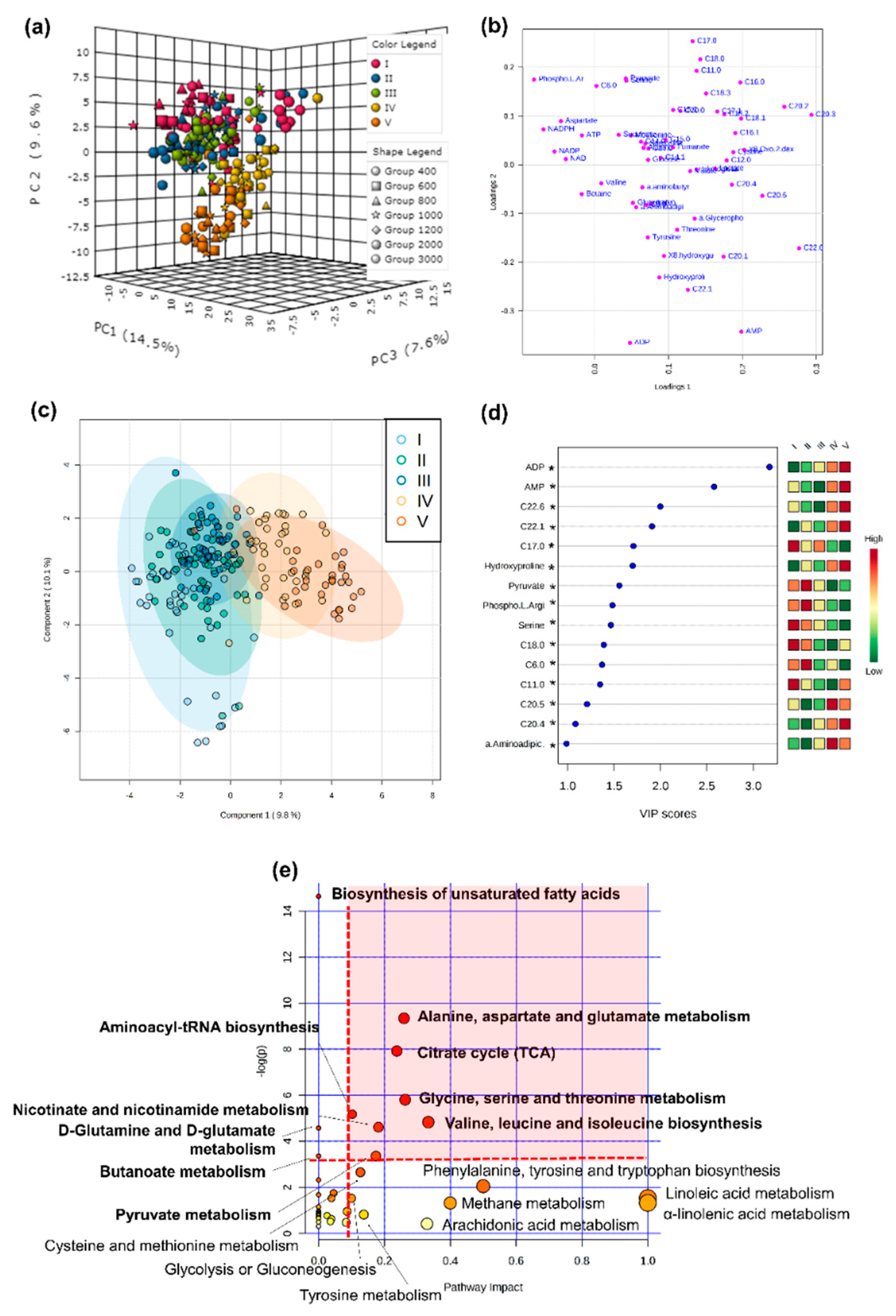
2.1. Functional Changes across Ontogeny
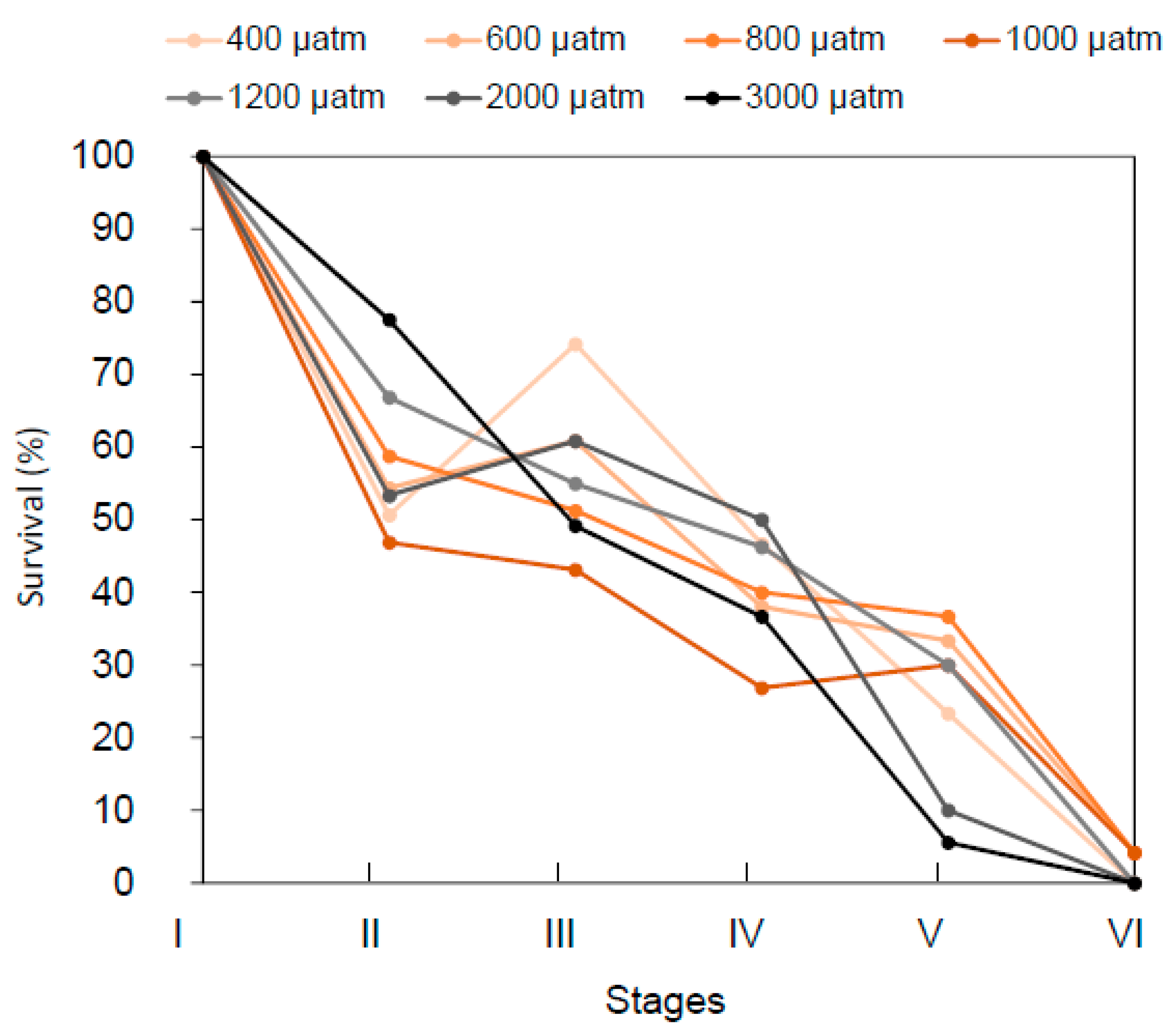
2.2. Stage-Specific pCO2 Effect on Survival Rates
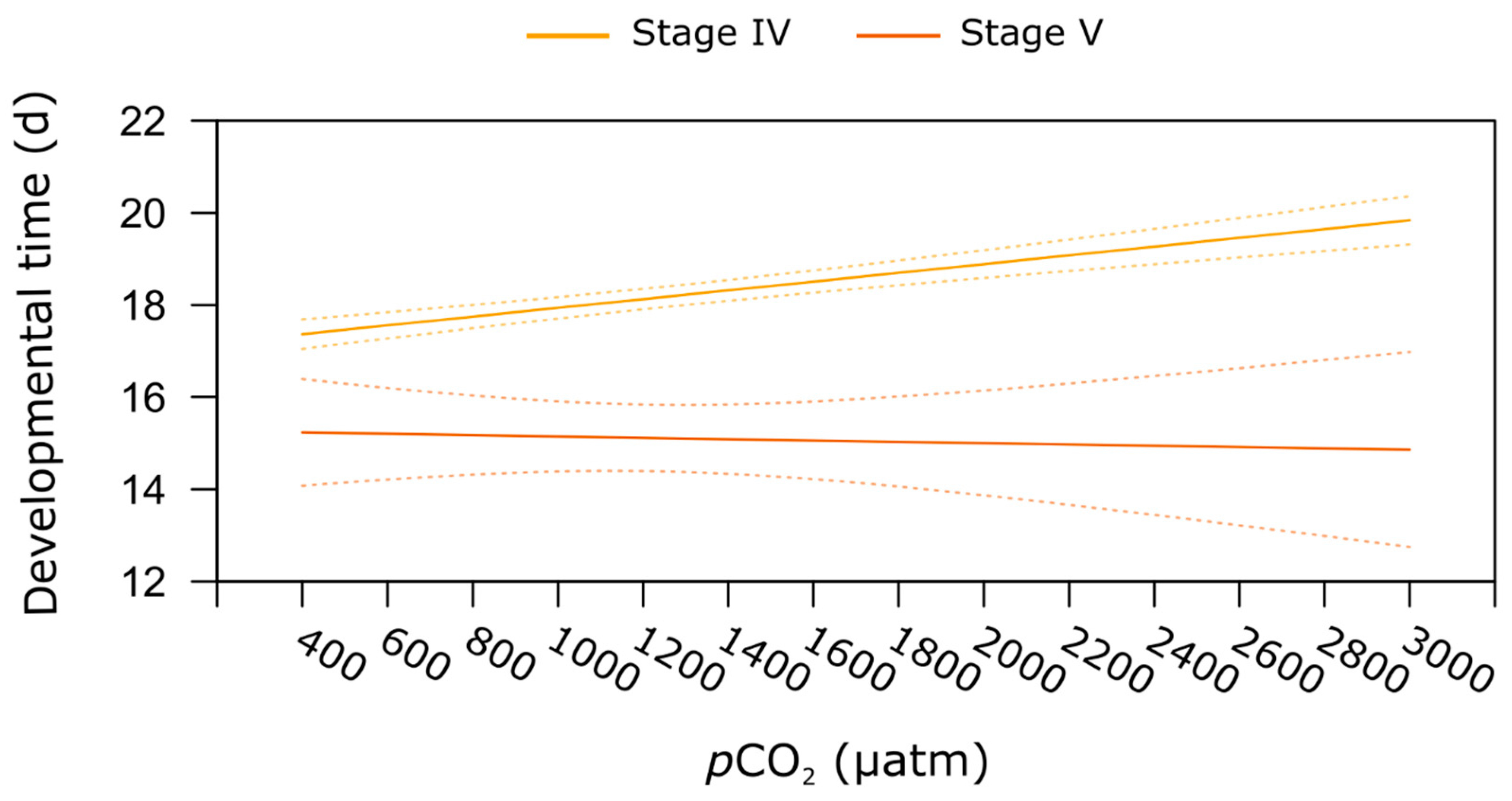
2.3. Stage-Specific pCO2 Effect on Developmental Time
2.4. Stage-Specific pCO2 Effect on Carapace Morphometrics
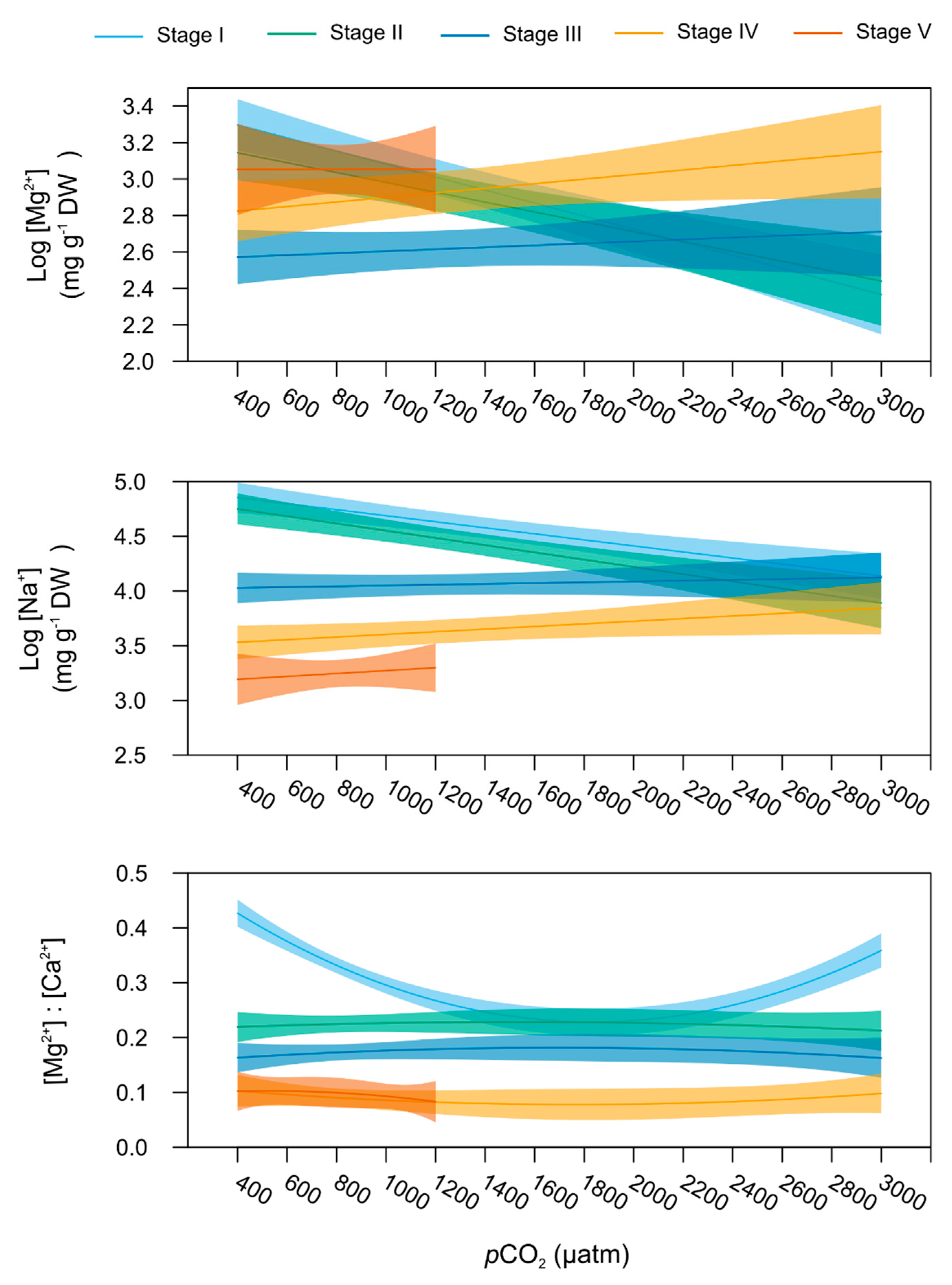
2.5. Stage-Specific pCO2 Effect on Carapace Mineralisation
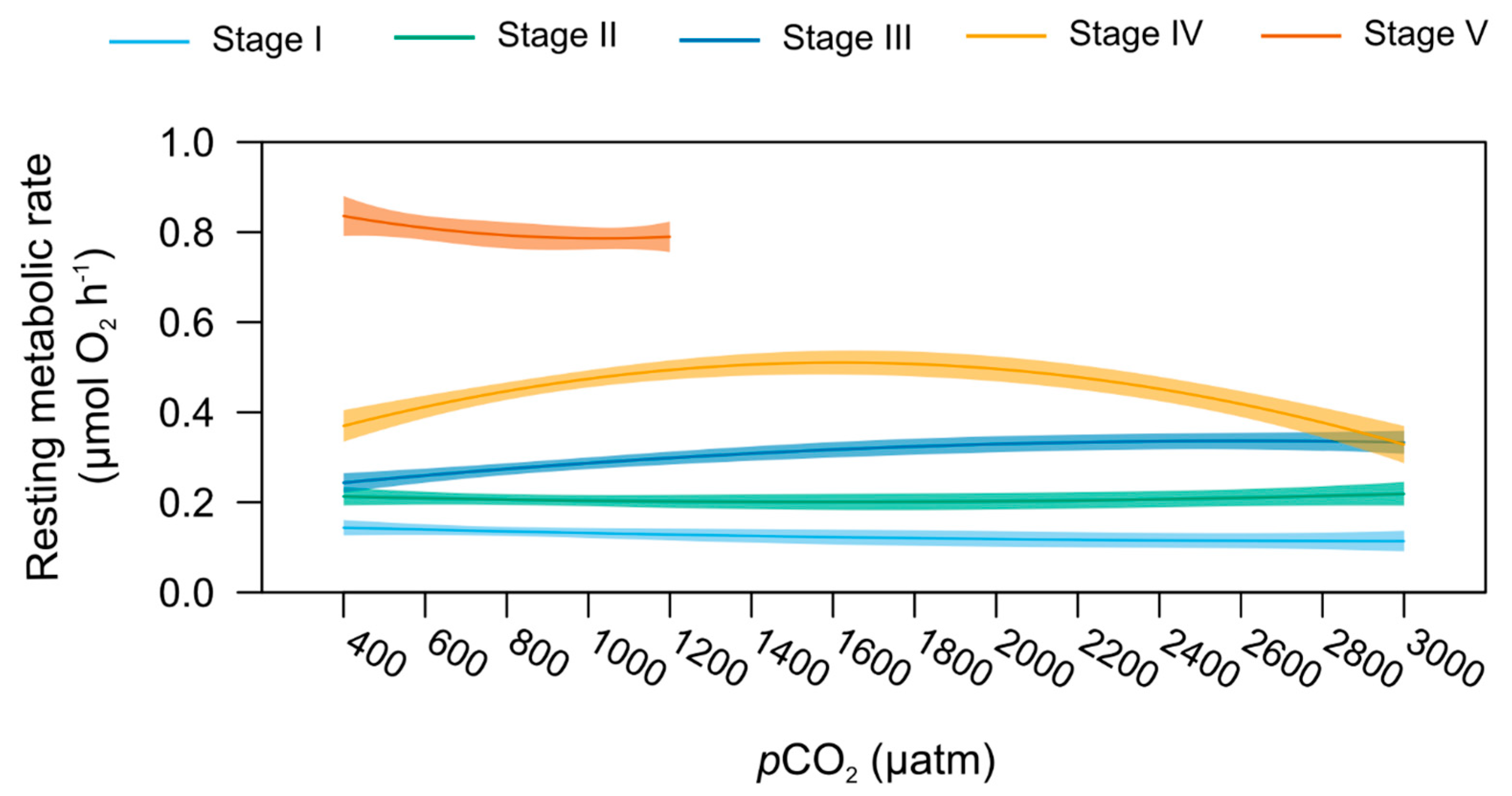
2.6. Stage-Specific pCO2 Effect on Resting Metabolic and Feeding Rates
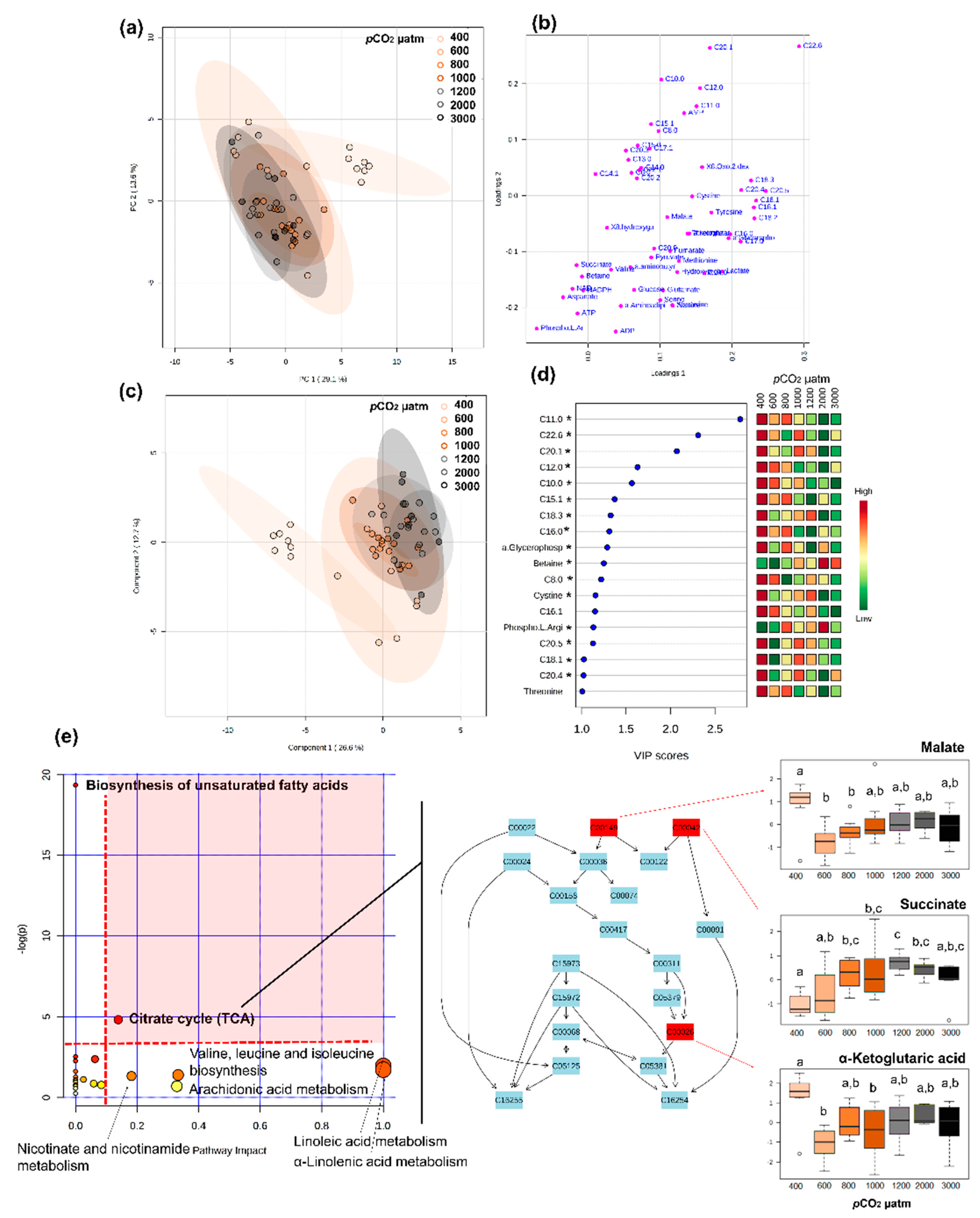
2.7. Stage-Specific pCO2 Effect on Metabolomic Fingerprints
3. Discussion
3.1. Metabolomic Reprogramming throughout Development in the American Lobster
3.2. Larval Stages I and II: The Establishment of the Internal Regulation Machinery
3.3. Stage III Larvae: The Metamorphosis as a Turning Point in the Ontogeny
3.4. Passed Metamorphosis: Post-Larval and Juvenile Stages Can Cope with Elevated pCO2 but Are More Sensitive Than Larvae
4. Materials and Methods
4.1. Specimen Collection
4.2. Experimental Design, Setup and Protocol
4.3. Seawater Parameter Monitoring
4.4. Survival
4.5. Feeding Rate (Only for Juveniles)
4.6. Resting Metabolic Rates (MO2)
4.7. Morphological Traits
4.8. Carapace Mineralisation
4.9. Statistical Analyses for Life-History and Physiological Traits
4.10. Metabolomic Fingerprints
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pechenik, J.A. On the advantages and disadvantages of larval stages in benthic marine invertebrate life cycles. Mar. Ecol. -Prog. Ser. 1999, 177, 269–297. [Google Scholar] [CrossRef]
- Marshall, D.J.; Morgan, S.G. Ecological and evolutionary consequences of linked life-history stages in the sea. Curr. Biol. 2011, 21, R718–R725. [Google Scholar] [CrossRef]
- Young, C.M. Larval ecology of marine invertebrates: A sesquicentennial history. Ophelia 1990, 32, 1–48. [Google Scholar] [CrossRef]
- Charmantier, G.; Charmantier-Daures, M.; Aiken, D.E. Metamorphosis in the Lobster Homarus (Decapoda): A Review. J. Crustacean Biol. 1991, 11, 481–495. [Google Scholar] [CrossRef]
- Pechenik, J.A. Larval experience and latent effects: Metamorphosis is not a new beginning. Integr. Comp. Biol. 2006, 46, 323–333. [Google Scholar] [CrossRef]
- Byrne, M. Impact of ocean warming and ocean acidification on marine invertebrate life history stages: Vulnerabilities and potential for persistence in a changing ocean. In Oceanography and Marine Biology: An Annual Review; Gibson, R., Atkinson, R., Gordon, J., Smith, I., Hughes, D., Eds.; Taylor & Francis: Abingdon, UK, 2011. [Google Scholar]
- Charmantier, G.; Haond, C.; Lignot, J.; Charmantier-Daures, M. Ecophysiological adaptation to salinity throughout a life cycle: A review in homarid lobsters. J. Exp. Biol. 2001, 204, 967–977. [Google Scholar] [CrossRef]
- Whiteley, N.M. Physiological and ecological responses of crustaceans to ocean acidification. Mar. Ecol. Prog. Ser. 2011, 430, 257–271. [Google Scholar] [CrossRef]
- Bindoff, N.L.; Cheung, W.W.L.; Kairo, J.G.; Aristegui, G.; Guinder, V.A.; Hallberg, R.; Hilmi, N.; Jiao, N.; Karim, M.S.; Levin, L.A.; et al. Changing Ocean, Marine Ecosystems, and Dependent Communities. In IPCC Special Report on the Ocean and Cryosphere in a Changing Climate; Pörtner, H.-O., Roberts, D.C., Masson-Delmotte, V., Zhai, P., Tignor, M., Poloczanska, E., Mintenbeck, K., Alegría, A., Nicolai, M., Okem, A., et al., Eds.; Intergovernmental Panel on Climate Change, Press CU: Cambridge, UK; New York, NY, USA, 2019; pp. 447–588. [Google Scholar]
- Burger, F.A.; John, J.G.; Frölicher, T.L. Increase in ocean acidity variability and extremes under increasing atmospheric CO2. Biogeosciences 2020, 17, 4633–4662. [Google Scholar] [CrossRef]
- Diaz, R.J.; Rosenberg, R. Spreading dead zones and consequences for marine ecosystems. Science 2008, 321, 926–929. [Google Scholar] [CrossRef] [PubMed]
- Kroeker, K.J.; Kordas, R.; Crim, R.; Hendriks, I.E.; Ramajo, L.; Singh, G.S.; Duarte, C.M.; Gattuso, J.-P. Impacts of ocean acidification on marine organisms: Quantifying sensitivities and interaction with warming. Glob. Chang. Biol. 2013, 19, 1884–1896. [Google Scholar] [CrossRef]
- Stocker, T.F.; Qin, D.; Plattner, G.-K.; Tignor, M.M.B.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M. Climate Change 2013. The Physical Science Basis. In Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Press CU: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Orr, J.C.; Fabry, V.J.; Aumont, O.; Bopp, L.; Doney, S.; Feely, R.A.; Gnanadesikan, A.; Gruber, N.; Ishida, A.; Joos, F.; et al. Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms. Nature 2005, 437, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Feely, R.A.; Doney, S.C.; Cooley, S.R. Ocean acidification: Present conditions and future changes in a high-CO2 world. Oceanography 2009, 22, 36–47. [Google Scholar] [CrossRef]
- Duarte, C.M.; Hendriks, I.E.; Moore, T.S.; Olsen, Y.S.; Steckbauer, A.; Ramajo, L.; Carstensen, J.; Trotter, J.A.; McCulloch, M. Is ocean acidification an open-ocean syndrome? Understanding anthropogenic impacts on seawater pH. Estuaries Coasts 2013, 36, 221–236. [Google Scholar] [CrossRef]
- Waldbusser, G.G.; Salisbury, J.E. Ocean acidification in the coastal zone from an organism’s perspective: Multiple system parameters, frequency domains, and habitats. Annu. Rev. Mar. Sci. 2013, 6, 221–247. [Google Scholar] [CrossRef] [PubMed]
- Spalding, C.; Finnegan, S.; Fischer, W.W. Energetic costs of calcification under ocean acidification. Glob. Biogeochem. Cycles 2017, 31, 866–877. [Google Scholar] [CrossRef]
- Melzner, F.; Mark, F.C.; Seibel, B.A.; Tomanek, L. Ocean Acidification and Coastal Marine Invertebrates: Tracking CO2 Effects from Seawater to the Cell. Annu. Rev. Mar. Science 2020, 12, 499–523. [Google Scholar] [CrossRef]
- Pandori, L.L.; Sorte, C.J. The weakest link: Sensitivity to climate extremes across life stages of marine invertebrates. Oikos 2019, 128, 621–629. [Google Scholar] [CrossRef]
- Coffey, W.D.; Nardone, J.A.; Yarram, A.; Long, W.C.; Swiney, K.M.; Foy, R.J.; Dickinson, G.H. Ocean acidification leads to altered micromechanical properties of the mineralized cuticle in juvenile red and blue king crabs. J. Exp. Mar. Biol. Ecol. 2017, 495, 1–12. [Google Scholar] [CrossRef]
- Donohue, P.; Calosi, P.; Bates, A.; Laverock, B.; Rastrick, S.P.S.; Mark, F.C.; Strobel, A.; Widdicombe, S. Physiological and behavioural impacts of exposure to elevated pCO2 on an important ecosystem engineer the burrowing shrimp Upogebia deltaura. Aquat. Biol. 2012, 15, 73–86. [Google Scholar] [CrossRef]
- Hammer, K.M.; Pedersen, S.A. Deep-water prawn Pandalus borealis displays a relatively high pH regulatory capacity in response to CO2-induced acidosis. Mar. Ecol. -Prog. Ser. 2013, 492, 139–151. [Google Scholar] [CrossRef]
- Knapp, J.L.; Bridges, C.R.; Krohn, J.; Hoffman, L.C.; Auerswald, L. The effects of hypercapnia on the West Coast rock lobster (Jasus lalandii) through acute exposure to decreased seawater pH—Physiological and biochemical responses. J. Exp. Mar. Biol. Ecol. 2016, 476, 58–64. [Google Scholar] [CrossRef]
- Lowder, K.B.; Allen, M.C.; Day, J.M.; Deheyn, D.D.; Taylor, J.R.; Woodson, H.E.B. Assessment of ocean acidification and warming on the growth, calcification, and biophotonics of a California grass shrimp. ICES J. Mar. Science 2017, 74, 1150–1158. [Google Scholar] [CrossRef]
- Miller, J.J.; Maher, M.; Bohaboy, E.; Friedman, C.S.; Mcelhany, P. Exposure to low pH reduces survival and delays development in early life stages of Dungeness crab (Cancer magister). Mar. Biol. 2016, 163, 118. [Google Scholar] [CrossRef]
- Green, B.S.; Gardner, C.; Hochmuth, J.D.; Linnane, A. Environmental effects on fished lobsters and crabs. Rev. Fish Biol. Fish. 2014, 24, 613–638. [Google Scholar] [CrossRef]
- Whiteley, N.M.; Taylor, E. Responses to environmental stresses: Oxygen, temperature and pH. Physiol. Nat. Hist. Crustacea 2015, 4, 320–358. [Google Scholar]
- Aiken, D. Proecdysis, setal development, and molt prediction in the American lobster (Homarus americanus). J. Fish. Board Can. 1973, 30, 1337–1344. [Google Scholar] [CrossRef]
- Roer, R.; Dillaman, R. The structure and calcification of the crustacean cuticle. Am. Zool. 1984, 24, 893–909. [Google Scholar] [CrossRef]
- Arnberg, M.; Calosi, P.; Spicer, J.I.; Tandberg, A.H.S.; Nilsen, M.; Westerlund, S.; Bechmann, R.K. Elevated temperature elicits greater effects than decreased pH on the development, feeding and metabolism of northern shrimp (Pandalus borealis) larvae. Mar. Biol. 2013, 160, 2037–2048. [Google Scholar] [CrossRef]
- Arnold, K.; Findlay, H.; Spicer, J.; Daniels, C.; Boothroyd, D. Effect of CO 2-related acidification on aspects of the larval development of the European lobster, Homarus gammarus (L.). Biogeosciences 2009, 6, 1747–1754. [Google Scholar] [CrossRef]
- Rato, L.D.; Novais, S.C.; Lemos, M.F.; Alves, L.M.; Leandro, S.M. Homarus gammarus (Crustacea: Decapoda) larvae under an ocean acidification scenario: Responses across different levels of biological organization. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 203, 29–38. [Google Scholar] [CrossRef]
- Small, D.P.; Calosi, P.; Boothroyd, D.; Widdicombe, S.; Spicer, J.I. Stage-Specific Changes in Physiological and Life-History Responses to Elevated Temperature and Pco2 during the Larval Development of the European LobsterHomarus gammarus (L.). Physiol. Biochem. Zool. 2015, 88, 494. [Google Scholar] [CrossRef]
- Walther, K.; Anger, K.; Portner, H.O. Effects of ocean acidification and warming on the larval development of the spider crab Hyas araneus from different latitudes (54 degrees vs. 79 degrees N). Mar. Ecol. -Prog. Ser. 2010, 417, 159–170. [Google Scholar] [CrossRef]
- Niemisto, M.; Fields, D.M.; Clark, K.F.; Waller, J.D.; Greenwood, S.J.; Wahle, R.A. American lobster postlarvae alter gene regulation in response to ocean warming and acidification. Ecol. Evol. 2021, 11, 806–819. [Google Scholar] [CrossRef]
- Caputi, N.; Lestang, S.; Frusher, S.; Wahle, R.A. The impact of climate change on exploited lobster stocks. In Lobsters: Biology, Management, Aquaculture and Fisheries, 2nd ed.; John Wiley and Sons: Hoboken, NJ, USA, 2013; pp. 84–112. [Google Scholar]
- Comeau, M.; Savoie, F. Movement of American lobster (Homarus americanus) in the southwestern Gulf of St. Lawrence. Fish. Bull. 2002, 100, 181–192. [Google Scholar]
- Keppel, E.A.; Scrosati, R.A.; Courtenay, S.C. Ocean acidification decreases growth and development in American lobster (Homarus americanus) larvae. J. Northwest Atl. Fish. Sci. 2012, 44, 61–66. [Google Scholar] [CrossRef]
- Mclean, E.L.; Katenka, N.V.; Seibel, B.A. Decreased growth and increased shell disease in early benthic phase Homarus americanus in response to elevated CO2. Mar. Ecol. -Prog. Ser. 2018, 596, 113–126. [Google Scholar] [CrossRef]
- Menu-Courey, K.; Noisette, F.; Piedalue, S.; Daoud, D.; Blair, T.; Blier, P.U.; Azetsu-Scott, K.; Calosi, P. Energy metabolism and survival of the juvenile recruits of the American lobster (Homarus americanus) exposed to a gradient of elevated seawater pCO2. Mar. Environ. Res. 2019, 143, 111–123. [Google Scholar] [CrossRef]
- Waller, J.D.; Wahle, R.A.; Mcveigh, H.; Fields, D.M. Linking rising pCO2 and temperature to the larval development and physiology of the American lobster (Homarus americanus). ICES J. Mar. Sci. 2016, 74, 1210–1219. [Google Scholar] [CrossRef]
- Bednaršek, N.; Calosi, P.; Feely, R.A.; Ambros, R.; Byrne, M.; Dupont, S.; Padilla-Gamiño, J.L.; Spicer, J.I.; Kessouri, F.; Roethler, M.; et al. Synthesis of Thresholds of Ocean Acidification Impacts on Calcifying Echinoderms. Front. Mar. Sci. 2021, 8, 261. [Google Scholar] [CrossRef]
- Bednaršek, N.; Feely, R.A.; Howes, E.L.; Hunt, B.P.V.; Kessouri, F.; León, P.; Lischka, S.; Maas, A.E.; McLaughlin, K.; Nezlin, N.P.; et al. Systematic Review and Meta-Analysis Toward Synthesis of Thresholds of Ocean Acidification Impacts on Calcifying Pteropods and Interactions with Warming. Front. Mar. Sci. 2019, 6, 6. [Google Scholar] [CrossRef]
- Christen, N.; Calosi, P.; Mcneill, C.L.; Widdicombe, S. Structural and functional vulnerability to elevated pCO2 in marine benthic communities. Mar. Biol. 2013, 160, 2113–2128. [Google Scholar] [CrossRef]
- Dupont, S.; Pörtner, H.-O. A snapshot of ocean acidification research. Mar. Biol. 2013, 160, 1765–1771. [Google Scholar] [CrossRef]
- Walther, K.; Sartoris, F.J.; Bock, C.; Portner, H.O. Impact of anthropogenic ocean acidification on thermal tolerance of the spider crab Hyas araneus. Biogeosciences 2009, 6, 2207–2215. [Google Scholar] [CrossRef]
- Spicer, J.I.; Gaston, K.J. Physiological Diversity and Its Ecological Implications; Blackwell Science: Oxford, UK, 1999; p. 241. [Google Scholar]
- Mcwilliam, P.S.; Phillips, B.F. Spiny lobster development: Mechanisms inducing metamorphosis to the puerulus: A review. Rev. Fish Biol. Fish. 2007, 17, 615–632. [Google Scholar] [CrossRef]
- Jung, H.; Lyons, R.E.; Hurwood, D.A.; Mather, P.B. Genes and growth performance in crustacean species: A review of relevant genomic studies in crustaceans and other taxa. Rev. Aquac. 2013, 5, 77–110. [Google Scholar] [CrossRef]
- Dhillon, S.S.; Torell, F.; Donten, M.; Lundstedt-Enkel, K.; Bennett, K.; Rännar, S.; Trygg, J.; Lundstedt, T. Metabolic profiling of zebrafish embryo development from blastula period to early larval stages. PLoS ONE 2019, 14, e0213661. [Google Scholar] [CrossRef] [PubMed]
- Byrne, M. Global change ecotoxicology: Identification of early life history bottlenecks in marine invertebrates, variable species responses and variable experimental approaches. Mar. Environ. Res. 2012, 76, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Byrne, M.; Przeslawski, R. Multistressor impacts of warming and acidification of the ocean on marine invertebrates’ life histories. Integr. Comp. Biol. 2013, 53, 582–596. [Google Scholar] [CrossRef] [PubMed]
- Long, C.W.; Swiney, K.M.; Foy, R.J. Effects of ocean acidification on the embryos and larvae of red king crab, Paralithodes camtschaticus. Mar. Pollut. Bull. 2013, 69, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Bechmann, R.K.; Taban, I.C.; Westerlund, S.; Godal, B.F.; Arnberg, M.; Vingen, S.; Ingvarsdottir, A.; Baussant, T. Effects of ocean acidification on early life stages of shrimp (Pandalus borealis) and mussel (Mytilus edulis). J. Toxicol. Environ. Health Part A 2011, 74, 424–438. [Google Scholar] [CrossRef]
- Long, W.C.; Swiney, K.M.; Foy, R.J. Effects of high pCO2 on Tanner crab reproduction and early life history, Part II: Carryover effects on larvae from oogenesis and embryogenesis are stronger than direct effects. ICES J. Mar. Sci. J. Cons. 2016, 73, 836–848. [Google Scholar] [CrossRef]
- Schiffer, M.; Harms, L.; Pörtner, H.O.; Lucassen, M.; Mark, F.C.; Storch, D. Tolerance of Hyas araneus zoea I larvae to elevated seawater PCO2 despite elevated metabolic costs. Mar. Biol. 2013, 160, 1943–1953. [Google Scholar] [CrossRef]
- Page, T.M.; Worthington, S.; Calosi, P.; Stillman, J.H. Effects of elevated pCO2 on crab survival and exoskeleton composition depend on shell function and species distribution: A comparative analysis of carapace and claw mineralogy across four porcelain crab species from different habitats. ICES J. Mar. Sci. J. Cons. 2016, 74, 1021–1032. [Google Scholar] [CrossRef]
- Tsang, H.H.; Welch, M.J.; Munday, P.L.; Ravasi, T.; Schunter, C. Proteomic Responses to Ocean Acidification in the Brain of Juvenile Coral Reef Fish. Front. Mar. Sci. 2020, 7, 605. [Google Scholar] [CrossRef]
- Aze, T.; Barry, J.; Bellerby, R.G.; Brander, L.; Byrne, M.; Dupont, S.; Gattuso, J.P.; Gibbs, S.; Hansson, L.; Hattam, C.; et al. An Updated Synthesis of the Impacts of Ocean Acidification on Marine Biodiversity (CBD Technical Series; 75); Secretariat of the Convention on Biological Diversity: Rio de Janeiro, Brazil, 2014. [Google Scholar]
- Zhan, Y.; Cui, D.; Xing, D.; Zhang, J.; Zhang, W.; Li, Y.; Li, C.; Chang, Y. CO2-driven ocean acidification repressed the growth of adult sea urchin Strongylocentrotus intermedius by impairing intestine function. Mar. Pollut. Bull. 2020, 153, 110944. [Google Scholar] [CrossRef] [PubMed]
- Lannig, G.; Eilers, S.; Pörtner, H.-O.; Sokolova, I.M.; Bock, C. Impact of ocean acidification on energy metabolism of oyster, Crassostrea gigas-Changes in metabolic pathways and thermal response. Mar. Drugs 2010, 8, 2318–2339. [Google Scholar] [CrossRef]
- Mills, E.; O’neill, L.A.J. Succinate: A metabolic signal in inflammation. Trends Cell Biol. 2014, 24, 313–320. [Google Scholar] [CrossRef]
- Laino, A.; Lopez-Zavala, A.A.; Garcia-Orozco, K.D.; Carrasco-Miranda, J.S.; Santana, M.; Stojanoff, V.; Sotelo-Mundo, R.R.; Garcia, C.F. Biochemical and structural characterization of a novel arginine kinase from the spider Polybetes pythagoricus. PeerJ 2017, 5, e3787. [Google Scholar] [CrossRef] [PubMed]
- Schneider, A.; Wiesner, R.J.; Grieshaber, M.K. On the role of arginine kinase in insect flight muscle. Insect Biochem. 1989, 19, 471–480. [Google Scholar] [CrossRef]
- Lin, W.; Ren, Z.; Mu, C.; Ye, Y.; Wang, C. Effects of Elevated pCO2 on the Survival and Growth of Portunus trituberculatus. Front. Physiol. 2020, 11, 750. [Google Scholar] [CrossRef] [PubMed]
- Pane, E.F.; Barry, J.P. Extracellular acid–base regulation during short-term hypercapnia is effective in a shallow-water crab, but ineffective in a deep-sea crab. Mar. Ecol. -Prog. Ser. 2007, 334, 1–9. [Google Scholar] [CrossRef]
- Zittier, Z.M.; Hirse, T.; Pörtner, H.-O. The synergistic effects of increasing temperature and CO 2 levels on activity capacity and acid–base balance in the spider crab, Hyas araneus. Mar. Biol. 2013, 160, 2049–2062. [Google Scholar] [CrossRef]
- Harrington, A.M.; Hamlin, H.J. Ocean acidification alters thermal cardiac performance, hemocyte abundance, and hemolymph chemistry in subadult American lobsters Homarus americanus H. Milne Edwards, 1837 (Decapoda: Malcostraca: Nephropidae). J. Crustacean Biol. 2019, 39, 468–476. [Google Scholar] [CrossRef]
- Charmantier, G.; Charmantier-Daures, M.; Bouaricha, N.; Thuet, P.; Trilles, J.-P.; Aiken, D. Ontogeny of osmoregulation and salinity tolerance in two decapod crustaceans: Homarus americanus and Penaeus japonicus. Biol. Bull. 1988, 175, 102–110. [Google Scholar] [CrossRef]
- Trigg, S.A.; McElhany, P.; Maher, M.; Perez, D.; Busch, D.S.; Nichols, K.M. Uncovering mechanisms of global ocean change effects on the Dungeness crab (Cancer magister) through metabolomics analysis. Sci Rep. 2019, 9, 10717. [Google Scholar] [CrossRef]
- Hammer, K.M.; Pedersen, S.A.; Størseth, T.R. Elevated seawater levels of CO2 change the metabolic fingerprint of tissues and hemolymph from the green shore crab Carcinus maenas. Comp. Biochem. Physiol. Part D Genom. Proteom. 2012, 7, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Gaytán, M.F.; Hernández-Palomares, M.L.; Soñanez-Organis, J.G.; Muhlia-Almazán, A.; Sánchez-Paz, A.; Stephens-Camacho, N.A.; Valenzuela-Soto, E.M.; Rosas-Rodríguez, J.A. Molecular characterization and organ-specific expression of the gene that encodes betaine aldehyde dehydrogenase from the white shrimp Litopenaeus vannamei in response to osmotic stress. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2015, 189, 40–46. [Google Scholar] [CrossRef]
- Stephens-Camacho, N.A.; Muhlia-Almazan, A.; Sanchez-Paz, A.; Rosas-Rodríguez, J.A. Surviving environmental stress: The role of betaine aldehyde dehydrogenase in marine crustaceans. Invertebr. Surviv. J. 2015, 12, 66–74. [Google Scholar]
- Burg, M.B.; Ferraris, J.D. Intracellular organic osmolytes: Function and regulation. J. Biol. Chem. 2008, 283, 7309–7313. [Google Scholar] [CrossRef]
- Small, D.P.; Calosi, P.; Boothroyd, D.; Widdicombe, S.; Spicer, J.I. The sensitivity of the early benthic juvenile stage of the European lobster Homarus gammarus (L.) to elevated pCO 2 and temperature. Mar. Biol. 2016, 163, 53. [Google Scholar] [CrossRef]
- Incze, L.S.; Wolff, N.; Wahle, R.A. Can scientific observations of early life stages be scaled up to the level of a fished population? A case study using Homarus americanus. Fish. Res. 2003, 65, 33–46. [Google Scholar] [CrossRef]
- Cobb, J.S.; Wahle, R.A. Early Life History and Recruitment Processes of Clawed Lobsters. Crustaceana 1994, 67, 1–25. [Google Scholar] [CrossRef]
- Wahle, R.A.; Steneck, R.S. Habitat restrictions in early benthic life: Experiments on habitat selection and in situ predation with the American lobster. J. Exp. Mar. Biol. Ecol. 1992, 157, 91–114. [Google Scholar] [CrossRef]
- Carter, H.A.; Ceballos-Osuna, L.; Miller, N.A.; Stillman, J.H. Impact of ocean acidification on metabolism and energetics during early life stages of the intertidal porcelain crab Petrolisthes cinctipes. J. Exp. Biol. 2013, 216, 1412–1422. [Google Scholar] [CrossRef] [PubMed]
- Small, D.; Calosi, P.; White, D.; Spicer, J.I.; Widdicombe, S. Impact of medium-term exposure to CO2 enriched seawater on the physiological functions of the velvet swimming crab Necora puber. Aquat. Biol. 2010, 10, 11–21. [Google Scholar] [CrossRef]
- Wood, H.L.; Eriksson, S.P.; Nordborg, M.; Styf, H.K. The effect of environmental stressors on the early development of the Norway lobster Nephrops norvegicus (L.). J. Exp. Mar. Biol. Ecol. 2015, 473, 35–42. [Google Scholar] [CrossRef]
- Spicer, J.I.; Raffo, A.; Widdicombe, S. Influence of CO2-related seawater acidification on extracellular acid–base balance in the velvet swimming crab Necora puber. Mar. Biol. 2007, 151, 1117–1125. [Google Scholar] [CrossRef]
- Whiteley, N.M.; Egginton, S.; Taylor, E.W.; Raven, J. Acid-base regulation in crustaceans: The role of bicarbonate ions. In Seminar Series-Society for Experimental Biology; Cambridge University Press: Cambridge, UK, 1999; Volume 68, pp. 233–256. [Google Scholar]
- Calosi, P.; Melatunan, S.; Turner, L.M.; Artioli, Y.; Davidson, R.; Byrne, J.J.; Viant, M.R.; Widdicombe, S.; Rundle, S.D. Regional adaptation defines sensitivity to future ocean acidification. Nat. Commun. 2017, 8, 13994. [Google Scholar] [CrossRef]
- Blackford, J.C.; Jones, N.; Proctor, R.; Holt, J.; Widdicombe, S.; Lowe, D.J.; Rees, A.P. An initial assessment of the potential environmental impact of CO2 escape from marine carbon capture and storage systems. Proc. Inst. Mech. Eng. Part A J. Power Energy 2009, 223, 269–280. [Google Scholar] [CrossRef]
- Hauri, C.; Gruber, N.; Vogt, M.; Doney, S.C.; Feely, R.A.; Lachkar, Z.; Leinweber, A.; McDonnell, A.M.P.; Munnich, M.; Plattner, G.-K. Spatiotemporal variability and long-term trends of ocean acidification in the California Current System. Biogeosciences 2013, 10, 193–216. [Google Scholar] [CrossRef]
- Romano, N.; Zeng, C. Cannibalism of Decapod Crustaceans and Implications for Their Aquaculture: A Review of its Prevalence, Influencing Factors, and Mitigating Methods. Rev. Fish. Sci. Aquac. 2017, 25, 42–69. [Google Scholar] [CrossRef]
- Dickson, A.G.; Sabine, C.L.; Christian, J.R. (Eds.) Guide to Best Practices for Ocean CO2 Measurements; North Pacific Marine Science Organization: Sidney, Australia, 2007. [Google Scholar]
- Lewis, E.; Wallace, D.W.R. Program Developed for CO2 System Calculations; Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory, U.S. Department of Energy: Oak Ridge, TN, USA, 1998. [Google Scholar]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Speakman, J.R.; Mcqueenie, J. Limits to Sustained Metabolic Rate: The Link between Food Intake, Basal Metabolic Rate, and Morphology in Reproducing Mice, Mus musculus. Physiol. Zool. 1996, 69, 746–769. [Google Scholar] [CrossRef]
- Weiss, H.M.; Lozano-Álvarez, E.; Briones-Fourzán, P.; Negrete-Soto, F. Using red light with fixed-site video cameras to study the behavior of the spiny lobster, Panulirus argus, and associated animals at night and inside their shelters. Mar. Technol. Soc. J. 2006, 40, 86–95. [Google Scholar] [CrossRef]
- Pinheiro, J.C.; Bates, D.M.; Debroy, S.; Sarkar, D.; Team, R.C. Linear and Nonlinear Mixed Effects Models-Package “nlme”; Version 3.1-104; CRAN: Vienna, Austria, 2012. [Google Scholar]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Lu, W.; Kimball, E.; Rabinowitz, J.D. A high-performance liquid chromatography-tandem mass spectrometry method for quantitation of nitrogen-containing intracellular metabolites. J. Am. Soc. Mass Spectrom. 2016, 17, 37–50. [Google Scholar] [CrossRef][Green Version]
- Thibault, C.; Massamba-N’siala, G.; Noisette, F.; Vermandele, F.; Babin, M.; Calosi, P. Within- and trans-generational responses to combined global changes are highly divergent in two congeneric species of marine annelids. Mar. Biol. 2020, 167, 41. [Google Scholar] [CrossRef]
- Wei, R.; Wang, J.; Su, M.; Jia, E.; Chen, S.; Chen, T.; Ni, Y. Missing value imputation approach for mass spectrometry-based metabolomics data. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Worley, B.; Powers, R. Multivariate analysis in metabolomics. Curr. Metab. 2013, 1, 92–107. [Google Scholar]
- Worley, B.; Powers, R. PCA as a practical indicator of OPLS-DA model reliability. Curr. Metab. 2016, 4, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Wishart, D.S. MetPA: A web-based metabolomics tool for pathway analysis and visualization. Bioinformatics 2010, 26, 2342–2344. [Google Scholar] [CrossRef]
- Pörtner, H.-O.; Farrell, A.P. Physiology and climate change. Science 2008, 322, 690–692. [Google Scholar] [CrossRef]
- Giménez, K.A.; Torres, G.L. Linking life history traits in successive phases of a complex life cycle: Effects of larval biomass on early juvenile development in an estuarine crab, Chasmagnathus granulata. Oikos 2004, 104, 570–580. [Google Scholar] [CrossRef]
- Jarrett, J.N. Seasonal variation in larval condition and postsettlement performance of the barnacle Semibalanus balanoides. Ecology 2003, 84, 384–390. [Google Scholar] [CrossRef]
- Huey, R.B.; Bennett, A.F. Bart’s familiar quotations: The enduring biological wisdom of George A. Bartholomew. Physiol. Biochem. Zool. 2008, 81, 519–525. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Best Model Fitted | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| pCO2 | Stage | pCO2: Stage | Covariate | ||||||||||
| df | F | p | df | F | p | df | F | p | df | df | F | p | |
| Survival cumulated | 1 | 31.2 | <0.001 | no | |||||||||
| Survival between stages | 1 | 29.87 | <0.001 | 4 | 2129.87 | <0.001 | 4 | 666.51 | <0.001 | no | |||
| Developmental time | 1 | 11.572 | <0.001 | 1 | 15.884 | <0.001 | 1 | 0.908 | 0.342 | no | |||
| Morphometrics | |||||||||||||
| Total length (Lt) | 1 | 13.981 | <0.001 | 4 | 582.854 | <0.001 | 4 | 0.327 | 0.860 | no | |||
| ceph/Lt | 1 | 0.317 | 0.574 | 4 | 94.258 | <0.001 | 4 | 1.947 | 0.102 | no | |||
| abd/Lt | 1 | 2.291 | 0.131 | 4 | 23.342 | <0.001 | 4 | 1.673 | 0.155 | no | |||
| ceph/abd | 1 | 1.436 | 0.232 | 4 | 65.443 | <0.001 | 4 | 1.434 | 0.222 | no | |||
| Rostrum | 1 | 3.733 | 0.054 | 4 | 14.608 | <0.001 | 4 | 1.467 | 0.211 | Lt | 1 | 356.727 | <0.001 |
| Cephalothorax | 1 | 0.330 | 0.568 | 4 | 40.443 | <0.001 | 4 | 2.837 | 0.024 | Lt | 1 | 5075.599 | <0.001 |
| Abdomen | 1 | 0.847 | 0.358 | 4 | 21.266 | <0.001 | 4 | 3.538 | 0.007 | Lt | 1 | 2885.833 | <0.001 |
| Tail | 1 | 0.007 | 0.932 | 4 | 24.358 | <0.001 | 4 | 4.526 | 0.001 | Lt | 1 | 1095.440 | <0.001 |
| Claw | 1 | 1.166 | 0.281 | 4 | 120.891 | <0.001 | 4 | 0.793 | 0.530 | Lt | 1 | 4108.430 | <0.001 |
| Mineralisation | |||||||||||||
| log(Ca2+) | 1 | 0.766 | 0.384 | 4 | 5.955 | <0.001 | 4 | 1.478 | 0.215 | wbm | 1 | 55.695 | <0.001 |
| log(Mg2+) | 1 | 4.357 | 0.040 | 4 | 2.962 | 0.024 | 4 | 2.748 | 0.033 | wbm | 1 | 0.334 | 0.565 |
| log(Na+) | 1 | 5.137 | 0.026 | 4 | 2.111 | 0.086 | 4 | 2.720 | 0.034 | wbm | 1 | 102.544 | <0.001 |
| log(K+) | 1 | 4.665 | 0.033 | 4 | 3.970 | 0.005 | 4 | 1.753 | 0.145 | wbm | 1 | 95.232 | <0.001 |
| log(Sr+) | 1 | 0.541 | 0.464 | 4 | 5.849 | <0.001 | 4 | 1.257 | 0.293 | wbm | 1 | 49.116 | <0.001 |
| [Mg2+]:[Ca2+] | 1 | 5.049 | 0.008 | 4 | 19.040 | <0.001 | 4 | 2.316 | 0.026 | wbm | 1 | 125.105 | <0.001 |
| Feeding rate | 1 | 0.169 | 0.683 | 4 | 1.620 | 0.209 | 4 | 1.143 | 0.290 | wbm | 1 | 5.377 | 0.024 |
| Resting metabolic rate (MO2) | 1 | 1.608 | 0.202 | 4 | 4.440 | 0.002 | 4 | 3.210 | 0.001 | wbm | 1 | 1196.088 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noisette, F.; Calosi, P.; Madeira, D.; Chemel, M.; Menu-Courey, K.; Piedalue, S.; Gurney-Smith, H.; Daoud, D.; Azetsu-Scott, K. Tolerant Larvae and Sensitive Juveniles: Integrating Metabolomics and Whole-Organism Responses to Define Life-Stage Specific Sensitivity to Ocean Acidification in the American Lobster. Metabolites 2021, 11, 584. https://doi.org/10.3390/metabo11090584
Noisette F, Calosi P, Madeira D, Chemel M, Menu-Courey K, Piedalue S, Gurney-Smith H, Daoud D, Azetsu-Scott K. Tolerant Larvae and Sensitive Juveniles: Integrating Metabolomics and Whole-Organism Responses to Define Life-Stage Specific Sensitivity to Ocean Acidification in the American Lobster. Metabolites. 2021; 11(9):584. https://doi.org/10.3390/metabo11090584
Chicago/Turabian StyleNoisette, Fanny, Piero Calosi, Diana Madeira, Mathilde Chemel, Kayla Menu-Courey, Sarah Piedalue, Helen Gurney-Smith, Dounia Daoud, and Kumiko Azetsu-Scott. 2021. "Tolerant Larvae and Sensitive Juveniles: Integrating Metabolomics and Whole-Organism Responses to Define Life-Stage Specific Sensitivity to Ocean Acidification in the American Lobster" Metabolites 11, no. 9: 584. https://doi.org/10.3390/metabo11090584
APA StyleNoisette, F., Calosi, P., Madeira, D., Chemel, M., Menu-Courey, K., Piedalue, S., Gurney-Smith, H., Daoud, D., & Azetsu-Scott, K. (2021). Tolerant Larvae and Sensitive Juveniles: Integrating Metabolomics and Whole-Organism Responses to Define Life-Stage Specific Sensitivity to Ocean Acidification in the American Lobster. Metabolites, 11(9), 584. https://doi.org/10.3390/metabo11090584