
Metabolomic Profile in Venous Thromboembolism (VTE)

 ,
, 
Abstract
1. Introduction
2. Metabolomics and Related Challenges
Challenges
3. Deep Venous Thrombosis and Pulmonary Embolism
3.1. Deep Venous Thrombosis in Animal Studies
3.1.1. Metabolome Profiling of Venous Thrombus and Vein Walls
3.1.2. Metabolome Profiling of Whole Blood, Serum
3.1.3. Metabolome Profiling of Urine
3.2. Pulmonary Embolism in Animal Studies
3.3. Effects of Anti-Thrombotic Drug Compounds on the Metabolome Profile
3.4. Deep Venous Thrombosis in Human Studies
3.5. Pulmonary Embolism in Human Studies
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Jiang, X.; Zeleznik, O.A.; Lindström, S.; Lasky-Su, J.; Hagan, K.; Clish, C.B.; Eliassen, A.H.; Kraft, P.; Kabrhel, C. Metabolites Associated with the Risk of Incident Venous Thromboembolism: A Metabolomic Analysis. J. Am. Heart Assoc. 2018, 7, e010317. [Google Scholar] [CrossRef]
- Beckman, M.G.; Hooper, W.C.; Critchley, S.E.; Ortel, T.L. Venous thromboembolism: A public health concern. Am. J. Prev. Med. 2010, 38, S495–S501. [Google Scholar] [CrossRef]
- Kirlikaya, B.; Langridge, B.; Davies, A.H.; Onida, S. Metabolomics as a tool to improve decision making for the vascular surgeon—Wishful thinking or a dream come true? Vasc. Pharmacol. 2019, 116, 1–3. [Google Scholar] [CrossRef]
- Heit, J.A.; Armasu, S.M.; Asmann, Y.W.; Cunningham, J.M.; Matsumoto, M.E.; Petterson, T.M.; De Andrade, M. A genome-wide association study of venous thromboembolism identifies risk variants in chromosomes 1q24. 2 and 9q. J. Thromb. Haemost. 2012, 10, 1521–1531. [Google Scholar] [CrossRef] [PubMed]
- Sung, Y.; Spagou, K.; Kafeza, M.; Kyriakides, M.; Dharmarajah, B.; Shalhoub, J.; Diaz, J.A.; Wakefield, T.W.; Holmes, E.; Davies, A.H. Deep Vein Thrombosis Exhibits Characteristic Serum and Vein Wall Metabolic Phenotypes in the Inferior Vena Cava Ligation Mouse Model. Eur. J. Vasc. Endovasc. Surg. 2018, 55, 703–713. [Google Scholar] [CrossRef]
- Baldwin, M.J.; Moore, H.M.; Rudarakanchana, N.; Gohel, M.; Davies, A.H. Post-thrombotic syndrome: A clinical review. J. Thromb. Haemost. 2013, 11, 795–805. [Google Scholar] [CrossRef]
- Ma, N.; Yang, Y.; Liu, X.; Li, S.; Qin, Z.; Li, J. Plasma metabonomics and proteomics studies on the anti-thrombosis mechanism of aspirin eugenol ester in rat tail thrombosis model. J. Proteom. 2020, 215, 103631. [Google Scholar] [CrossRef] [PubMed]
- von Brühl, M.L.; Stark, K.; Steinhart, A.; Chandraratne, S.; Konrad, I.; Lorenz, M.; Khandoga, A.; Tirniceriu, A.; Coletti, R.; Köllnberger, M.; et al. Monocytes, neutrophils, and platelets cooperate to initiate and propagate venous thrombosis in mice in vivo. J. Exp. Med. 2012, 209, 819–835. [Google Scholar] [CrossRef] [PubMed]
- Kyrle, P.A.; Eichinger, S. Deep vein thrombosis. Lancet 2005, 365, 1163–1174. [Google Scholar] [CrossRef]
- Linkins, L.A.; Takach Lapner, S. Review of D-dimer testing: Good, Bad, and Ugly. Int. J. Lab. Hematol. 2017, 39, 98–103. [Google Scholar] [CrossRef]
- Bujak, R.; García-Álvarez, A.; Rupérez, F.J.; Nuño-Ayala, M.; García, A.; Ruiz-Cabello, J.; Fuster, V.; Ibáñez, B.; Barbas, C. Metabolomics Reveals Metabolite Changes in Acute Pulmonary Embolism. J. Proteome Res. 2014, 13, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Deguchi, H.; Banerjee, Y.; Trauger, S.; Siuzdak, G.; Kalisiak, E.; Fernández, J.A.; Hoang, L.; Tran, M.; Yegneswaran, S.; Elias, D.J.; et al. Acylcarnitines are anticoagulants that inhibit factor Xa and are reduced in venous thrombosis, based on metabolomics data. Blood 2015, 126, 1595–1600. [Google Scholar] [CrossRef]
- Germain, M.; Chasman, D.I.; De Haan, H.; Tang, W.; Lindström, S.; Weng, L.-C.; De Andrade, M.; De Visser, M.C.; Wiggins, K.L.; Suchon, P. Meta-analysis of 65,734 individuals identifies TSPAN15 and SLC44A2 as two susceptibility loci for venous thromboembolism. Am. J. Hum. Genet. 2015, 96, 532–542. [Google Scholar] [CrossRef]
- Scheres, L.J.J.; Lijfering, W.M.; Cannegieter, S.C. Current and future burden of venous thrombosis: Not simply predictable. Res. Pract. Thromb. Haemost. 2018, 2, 199–208. [Google Scholar] [CrossRef]
- National Clinical Guideline Centre (UK). Venous Thromboembolic Diseases: The Management of Venous Thromboembolic Diseases and the Role of Thrombophilia Testing; Royal College of Physicians: London, UK, 2012. [Google Scholar]
- Previtali, E.; Bucciarelli, P.; Passamonti, S.M.; Martinelli, I. Risk factors for venous and arterial thrombosis. Blood Transfus. 2011, 9, 120–138. [Google Scholar] [CrossRef] [PubMed]
- Lewis Gregory, D.; Asnani, A.; Gerszten Robert, E. Application of Metabolomics to Cardiovascular Biomarker and Pathway Discovery. J. Am. Coll. Cardiol. 2008, 52, 117–123. [Google Scholar] [CrossRef]
- Quintero, M.; Tasic, L.; Annichino-Bizzacchi, L.M. Thrombosis: Current knowledge based on metabolomics by nuclear magnetic resonance (NMR) spectroscopy and mass spectrometry (MS). Thromb. Update 2020, 1, 100011. [Google Scholar] [CrossRef]
- Wendelboe, A.M.; Raskob, G.E. Global Burden of Thrombosis. Circ. Res. 2016, 118, 1340–1347. [Google Scholar] [CrossRef]
- Martinez-Pinna, R.; Barbas, C.; Blanco-Colio, L.M.; Tunon, J.; Ramos-Mozo, P.; Lopez, J.A.; Meilhac, O.; Michel, J.B.; Egido, J.; Martin-Ventura, J.L. Proteomic and metabolomic profiles in atherothrombotic vascular disease. Curr. Atheroscler. Rep. 2010, 12, 202–208. [Google Scholar] [CrossRef]
- Leo, G.C.; Darrow, A.L. NMR-based metabolomics of urine for the atherosclerotic mouse model using apolipoprotein-E deficient mice. Magn. Reson. Chem. 2009, 47, S20–S25. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.W.; Wang, Z.; Levison, B.S.; Koeth, R.A.; Britt, E.B.; Fu, X.; Wu, Y.; Hazen, S.L. Intestinal microbial metabolism of phosphatidylcholine and cardiovascular risk. N. Engl. J. Med. 2013, 368, 1575–1584. [Google Scholar] [CrossRef]
- Obi, A.T.; Stringer, K.A.; Diaz, J.A.; Finkel, M.A.; Farris, D.M.; Yeomans, L.; Wakefield, T.; Myers, D.D., Jr. 1D-¹H-nuclear magnetic resonance metabolomics reveals age-related changes in metabolites associated with experimental venous thrombosis. J. Vasc. Surg. Venous Lymphat. Disord. 2016, 4, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Würtz, P.; Havulinna, A.S.; Soininen, P.; Tynkkynen, T.; Prieto-Merino, D.; Tillin, T.; Ghorbani, A.; Artati, A.; Wang, Q.; Tiainen, M. Metabolite profiling and cardiovascular event risk: A prospective study of 3 population-based cohorts. Circulation 2015, 131, 774–785. [Google Scholar] [CrossRef] [PubMed]
- Fraser, K.; Roy, N.C.; Goumidi, L.; Verdu, A.; Suchon, P.; Leal-Valentim, F.; Trégouët, D.A.; Morange, P.E.; Martin, J.C. Plasma Biomarkers and Identification of Resilient Metabolic Disruptions in Patients with Venous Thromboembolism Using a Metabolic Systems Approach. Arter. Thromb. Vasc. Biol. 2020, 40, 2527–2538. [Google Scholar] [CrossRef]
- Clish, C.B. Metabolomics: An emerging but powerful tool for precision medicine. Cold Spring Harb. Mol. Case Stud. 2015, 1, a000588. [Google Scholar] [CrossRef]
- Bahado-Singh, R.O.; Graham, S.F.; Han, B.; Turkoglu, O.; Ziadeh, J.; Mandal, R.; Er, A.; Wishart, D.S.; Stahel, P.L. Serum metabolomic markers for traumatic brain injury: A mouse model. Metabolomics 2016, 12, 100. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Holmes, E.; Kinross, J.M.; Darzi, A.W.; Takats, Z.; Lindon, J.C. Metabolic phenotyping in clinical and surgical environments. Nature 2012, 491, 384–392. [Google Scholar] [CrossRef]
- Posma, J.M.; Garcia-Perez, I.; Ebbels, T.M.; Lindon, J.C.; Stamler, J.; Elliott, P.; Holmes, E.; Nicholson, J.K. Optimized phenotypic biomarker discovery and confounder elimination via covariate-adjusted projection to latent structures from metabolic spectroscopy data. J. Proteome Res. 2018, 17, 1586–1595. [Google Scholar] [CrossRef]
- Dallmann, R.; Viola, A.U.; Tarokh, L.; Cajochen, C.; Brown, S.A. The human circadian metabolome. Proc. Natl. Acad. Sci. USA 2012, 109, 2625–2629. [Google Scholar] [CrossRef] [PubMed]
- Brunner, M.P.; Shah, S.H.; Craig, D.M.; Stevens, R.D.; Muehlbauer, M.J.; Bain, J.R.; Newgard, C.B.; Kraus, W.E.; Granger, C.B.; Sketch, M.H., Jr.; et al. Effect of heparin administration on metabolomic profiles in samples obtained during cardiac catheterization. Circ. Cardiovasc. Genet. 2011, 4, 695–700. [Google Scholar] [CrossRef]
- Grossman, M.I.; Moeller, H.C.; Palm, L. Effect of lipemia and heparin on free fatty acid concentration of serum in humans. Proc. Soc. Exp. Biol. Med. 1955, 90, 106–109. [Google Scholar] [CrossRef]
- Chekmeneva, E.; dos Santos Correia, G.A.; Chan, Q.; Wijeyesekera, A.; Tin, A.; Young, J.H.; Elliott, P.; Nicholson, J.K.; Holmes, E. Optimization and application of direct infusion nanoelectrospray HRMS method for large-scale urinary metabolic phenotyping in molecular epidemiology. J. Proteome Res. 2017, 16, 1646–1658. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, K.; Sugita, C.; Yamashita, A.; Moriguchi-Goto, S.; Furukoji, E.; Sakae, T.; Gi, T.; Hirai, T.; Asada, Y. Higher lactate and purine metabolite levels in erythrocyte-rich fresh venous thrombus: Potential markers for early deep vein thrombosis. Thromb. Res. 2019, 177, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, Y.; Xie, Z.; Ming, X.; Li, Z.; Kong, Y. A tryptophan derivative TD-26 attenuates thrombus formation by inhibiting both PI3K/Akt signaling and binding of fibrinogen to integrin αIIbβ3. Biochem. Biophys. Res. Commun. 2015, 465, 516–522. [Google Scholar] [CrossRef]
- Low, P.S. Structure and function of the cytoplasmic domain of band 3: Center of erythrocyte membrane—Peripheral protein interactions. Biochim. Biophys. Acta Rev. Biomembr. 1986, 864, 145–167. [Google Scholar] [CrossRef]
- Messana, I.; Orlando, M.; Cassiano, L.; Pennacchietti, L.; Zuppi, C.; Castagnola, M.; Giardina, B. Human erythrocyte metabolism is modulated by the O2-linked transition of hemoglobin. FEBS Lett. 1996, 390, 25–28. [Google Scholar] [CrossRef]
- Fukami, M.H.; Holmsen, H.; Salganicoff, L. Adenine nucleotide metabolism of blood platelets IX. Time course of secretion and changes in energy metabolism in thrombin-treated platelets. Biochim. Biophys. Acta Gen. Subj. 1976, 444, 633–643. [Google Scholar] [CrossRef]
- Crowell, J.W.; Houston, B. Effect of acidity on blood coagulation. Am. J. Physiol. Leg. Content 1961, 201, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Engström, M.; Schött, U.; Romner, B.; Reinstrup, P. Acidosis Impairs the Coagulation: A Thromboelastographic Study. J. Trauma 2006, 61, 624–628. [Google Scholar] [CrossRef] [PubMed]
- Fredholm, B.B. Adenosine, an endogenous distress signal, modulates tissue damage and repair. Cell Death Differ. 2007, 14, 1315–1323. [Google Scholar] [CrossRef]
- Cramer, T.; Yamanishi, Y.; Clausen, B.E.; Förster, I.; Pawlinski, R.; Mackman, N.; Haase, V.H.; Jaenisch, R.; Corr, M.; Nizet, V.; et al. HIF-1α Is Essential for Myeloid Cell-Mediated Inflammation. Cell 2003, 112, 645–657. [Google Scholar] [CrossRef]
- Fuentes, E.; Pereira, J.; Mezzano, D.; Alarcón, M.; Caballero, J.; Palomo, I. Inhibition of platelet activation and thrombus formation by adenosine and inosine: Studies on their relative contribution and molecular modeling. PLoS ONE 2014, 9, e112741. [Google Scholar] [CrossRef]
- Johnston-Cox, H.A.; Ravid, K. Adenosine and blood platelets. Purinergic Signal. 2011, 7, 357–365. [Google Scholar] [CrossRef]
- Gabazza, E.C.; Hayashi, T.; Ido, M.; Adachi, Y.; Suzuki, K. Adenosine inhibits thrombin-induced expression of tissue factor on endothelial cells by a nitric oxide-mediated mechanism. Clin. Sci. 2002, 102, 167–175. [Google Scholar] [CrossRef]
- Fuentes, E.; Badimon, L.; Caballero, J.; Padró, T.; Vilahur, G.; Alarcón, M.; Pérez, P.; Palomo, I. Protective mechanisms of adenosine 5′-monophosphate in platelet activation and thrombus formation. Thromb. Haemost. 2014, 112, 491–507. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, D.; Song, P.; Zou, M.H. Tryptophan-kynurenine pathway is dysregulated in inflammation, and immune activation. Front. Biosci. Landmark 2015, 20, 1116–1143. [Google Scholar] [CrossRef]
- Watanabe, Y.; Koyama, S.; Yamashita, A.; Matsuura, Y.; Nishihira, K.; Kitamura, K.; Asada, Y. Indoleamine 2,3-dioxygenase 1 in coronary atherosclerotic plaque enhances tissue factor expression in activated macrophages. Res. Pract. Thromb. Haemost. 2018, 2, 726–735. [Google Scholar] [CrossRef] [PubMed]
- Ray, P.D.; Huang, B.-W.; Tsuji, Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell. Signal. 2012, 24, 981–990. [Google Scholar] [CrossRef]
- Raha, S.; Robinson, B.H. Mitochondria, oxygen free radicals, disease and ageing. Trends Biochem. Sci. 2000, 25, 502–508. [Google Scholar] [CrossRef]
- Neurauter, G.; Grahmann, A.V.; Klieber, M.; Zeimet, A.; Ledochowski, M.; Sperner-Unterweger, B.; Fuchs, D. Serum phenylalanine concentrations in patients with ovarian carcinoma correlate with concentrations of immune activation markers and of isoprostane-8. Cancer Lett. 2008, 272, 141–147. [Google Scholar] [CrossRef]
- Liu, W.; Le, A.; Hancock, C.; Lane, A.N.; Dang, C.V.; Fan, T.W.-M.; Phang, J.M. Reprogramming of proline and glutamine metabolism contributes to the proliferative and metabolic responses regulated by oncogenic transcription factor c-MYC. Proc. Natl. Acad. Sci. USA 2012, 109, 8983–8988. [Google Scholar] [CrossRef]
- Paneni, F.; Costantino, S.; Cosentino, F. Molecular pathways of arterial aging. Clin. Sci. 2015, 128, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Romano, K.A.; Vivas, E.I.; Amador-Noguez, D.; Rey, F.E. Intestinal microbiota composition modulates choline bioavailability from diet and accumulation of the proatherogenic metabolite trimethylamine-N-oxide. MBio 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Romano, K.A.; Martinez-del Campo, A.; Kasahara, K.; Chittim, C.L.; Vivas, E.I.; Amador-Noguez, D.; Balskus, E.P.; Rey, F.E. Metabolic, Epigenetic, and Transgenerational Effects of Gut Bacterial Choline Consumption. Cell Host Microbe 2017, 22, 279.e277–290.e277. [Google Scholar] [CrossRef] [PubMed]
- Skye, S.M.; Zhu, W.; Romano, K.A.; Guo, C.-J.; Wang, Z.; Jia, X.; Kirsop, J.; Haag, B.; Lang, J.M.; DiDonato, J.A.; et al. Microbial Transplantation with Human Gut Commensals Containing CutC Is Sufficient to Transmit Enhanced Platelet Reactivity and Thrombosis Potential. Circ. Res. 2018, 123, 1164–1176. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Gregory, J.C.; Org, E.; Buffa, J.A.; Gupta, N.; Wang, Z.; Li, L.; Fu, X.; Wu, Y.; Mehrabian, M.; et al. Gut Microbial Metabolite TMAO Enhances Platelet Hyperreactivity and Thrombosis Risk. Cell 2016, 165, 111–124. [Google Scholar] [CrossRef]
- Kasahara, K.; Rey, F.E. The emerging role of gut microbial metabolism on cardiovascular disease. Curr. Opin. Microbiol. 2019, 50, 64–70. [Google Scholar] [CrossRef]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; DuGar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.-M. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef]
- Hoppel, C. The role of carnitine in normal and altered fatty acid metabolism. Am. J. Kidney Dis. 2003, 41, S4–S12. [Google Scholar] [CrossRef]
- El-Hattab, A.W.; Scaglia, F. Disorders of carnitine biosynthesis and transport. Mol. Genet. Metab. 2015, 116, 107–112. [Google Scholar] [CrossRef]
- Rutkowsky, J.M.; Knotts, T.A.; Ono-Moore, K.D.; McCoin, C.S.; Huang, S.; Schneider, D.; Singh, S.; Adams, S.H.; Hwang, D.H. Acylcarnitines activate proinflammatory signaling pathways. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E1378–E1387. [Google Scholar] [CrossRef] [PubMed]
- Aguer, C.; McCoin, C.S.; Knotts, T.A.; Thrush, A.B.; Ono-Moore, K.; McPherson, R.; Dent, R.; Hwang, D.H.; Adams, S.H.; Harper, M.-E. Acylcarnitines: Potential implications for skeletal muscle insulin resistance. FASEB J. 2015, 29, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Hannun, Y.A.; Obeid, L.M. Principles of bioactive lipid signalling: Lessons from sphingolipids. Nat. Rev. Mol. Cell Biol. 2008, 9, 139–150. [Google Scholar] [CrossRef]
- Augé, N.; Nègre-Salvayre, A.; Salvayre, R.; Levade, T. Sphingomyelin metabolites in vascular cell signaling and atherogenesis. Prog. Lipid Res. 2000, 39, 207–229. [Google Scholar] [CrossRef]
- Egom, E.E.A.; Chacko, S.; Stringer, S.E.; El-Omar, M.; Chirico, D.; Clarke, B.; Neyses, L.; Cruickshank, K.; Lei, M.; Fath-Ordoubadi, F. Serum sphingolipids level as a novel potential marker for early detection of human myocardial ischaemic injury. Front. Physiol. 2013, 4, 130. [Google Scholar] [CrossRef] [PubMed]
- Lowenstein, C.J.; Morrell, C.N.; Yamakuchi, M. Regulation of Weibel–Palade Body Exocytosis. Trends Cardiovasc. Med. 2005, 15, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Nakane, M.; Kubota, M.; Nakagomi, T.; Tamura, A.; Hisaki, H.; Shimasaki, H.; Ueta, N. Lethal forebrain ischemia stimulates sphingomyelin hydrolysis and ceramide generation in the gerbil hippocampus. Neurosci. Lett. 2000, 296, 89–92. [Google Scholar] [CrossRef]
- Wakefield, T.W.; Myers, D.D.; Henke, P.K. Role of selectins and fibrinolysis in VTE. Thromb. Res. 2009, 123, S35–S40. [Google Scholar] [CrossRef]
- Myers, D.D.; Culmer, D.; Diaz, J.; Hawley, A.; Jackson, T.; Shuster, K.; Sigler, R.; Wakefield, T. Vein Wall and Circulating P-selectin Promote Venous Thrombogenesis during Aging in a Rodent Model. J. Vasc. Surg. Venous Lymphat. Disord. 2013, 1, 114. [Google Scholar] [CrossRef]
- Cao, J.; Lü, X.G.; Li, Y.; Jin, Q.Q.; Chu, X.Y.; Wang, Y.Y.; Sun, J.H. Study on Urinary Metabolic Profile in Rats with Deep Venous Thrombosis Based on Pattern Recognition. Fa Yi Xue Za Zhi 2018, 34, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Tuder, R.M.; Davis, L.A.; Graham, B.B. Targeting energetic metabolism: A new frontier in the pathogenesis and treatment of pulmonary hypertension. Am. J. Respir. Crit. Care Med. 2012, 185, 260–266. [Google Scholar] [CrossRef]
- Yuan, J.X.-J.; Ward, J.P. Membrane Receptors, Channels and Transporters in Pulmonary Circulation; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2010; Volume 661. [Google Scholar]
- Agrawal, A.; Guttapalli, A.; Narayan, S.; Albert, T.J.; Shapiro, I.M.; Risbud, M.V. Normoxic stabilization of HIF-1α drives glycolytic metabolism and regulates aggrecan gene expression in nucleus pulposus cells of the rat intervertebral disk. Am. J. Physiol. Cell Physiol. 2007, 293, C621–C631. [Google Scholar] [CrossRef]
- Marsboom, G.; Wietholt, C.; Haney, C.R.; Toth, P.T.; Ryan, J.J.; Morrow, E.; Thenappan, T.; Bache-Wiig, P.; Piao, L.; Paul, J. Lung 18F-fluorodeoxyglucose positron emission tomography for diagnosis and monitoring of pulmonary arterial hypertension. Am. J. Respir. Crit. Care Med. 2012, 185, 670–679. [Google Scholar] [CrossRef]
- Funk, C.D. Prostaglandins and leukotrienes: Advances in eicosanoid biology. Science 2001, 294, 1871–1875. [Google Scholar] [CrossRef]
- Lambert, I.; Pedersen, S.; Poulsen, K. Activation of PLA2 isoforms by cell swelling and ischaemia/hypoxia. Acta Physiol. 2006, 187, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Bartke, N.; Hannun, Y.A. Bioactive sphingolipids: Metabolism and function. J. Lipid Res. 2009, 50, S91–S96. [Google Scholar] [CrossRef] [PubMed]
- Geoffroy, K.; Wiernsperger, N.; Lagarde, M.; El Bawab, S. Bimodal effect of advanced glycation end products on mesangial cell proliferation is mediated by neutral ceramidase regulation and endogenous sphingolipids. J. Biol. Chem. 2004, 279, 34343–34352. [Google Scholar] [CrossRef] [PubMed]
- Martinov, M.V.; Vitvitsky, V.M.; Banerjee, R.; Ataullakhanov, F.I. The logic of the hepatic methionine metabolic cycle. Biochim. Et Biophys. Acta Proteins Proteom. 2010, 1804, 89–96. [Google Scholar] [CrossRef]
- Gu, S.X.; Stevens, J.W.; Lentz, S.R. Regulation of thrombosis and vascular function by protein methionine oxidation. Blood 2015, 125, 3851–3859. [Google Scholar] [CrossRef]
- Keijzer, M.B.A.; den Heijer, M.; Borm, G.F.; Blom, H.J.; Vollset, S.E.; Hermus, A.R.M.M.; Ueland, P.M. Low fasting methionine concentration as a novel risk factor for recurrent venous thrombosis. Thromb. Haemost. 2006, 96, 492–497. [Google Scholar] [CrossRef][Green Version]
- Ozkan, Y.; Yardim-Akaydin, S.; Imren, E.; Torun, M.; Simşek, B. Increased plasma homocysteine and allantoin levels in coronary artery disease: Possible link between homocysteine and uric acid oxidation. Acta Cardiol. 2006, 61, 432–439. [Google Scholar] [CrossRef]
- Reuter, S.E.; Evans, A.M. Long-chain acylcarnitine deficiency in patients with chronic fatigue syndrome. Potential involvement of altered carnitine palmitoyltransferase-I activity. J. Intern. Med. 2011, 270, 76–84. [Google Scholar] [CrossRef]
- Minkler, P.E.; Kerner, J.; North, K.N.; Hoppel, C.L. Quantitation of long-chain acylcarnitines by HPLC/fluorescence detection: Application to plasma and tissue specimens from patients with carnitine palmitoyltransferase-II deficiency. Clin. Chim. Acta 2005, 352, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Chace, D.H.; DiPerna, J.C.; Mitchell, B.L.; Sgroi, B.; Hofman, L.F.; Naylor, E.W. Electrospray tandem mass spectrometry for analysis of acylcarnitines in dried postmortem blood specimens collected at autopsy from infants with unexplained cause of death. Clin. Chem. 2001, 47, 1166–1182. [Google Scholar] [CrossRef] [PubMed]
- Ijiri, Y.; Naemura, A.; Yamashita, T.; Meguro, S.; Watanabe, H.; Tokimitsu, I.; Yamamoto, J. Dietary diacylglycerol extenuates arterial thrombosis in apoE and LDLR deficient mice. Thromb. Res. 2006, 117, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Ijiri, Y.; Naemura, A.; Yamashita, T.; Ikarugi, H.; Meguro, S.; Tokimitsu, I.; Yamamoto, J. Mechanism of the antithrombotic effect of dietary diacylglycerol in atherogenic mice. Pathophysiol. Haemost. Thromb. 2006, 35, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Doggen, C.J.; Smith, N.L.; Lemaitre, R.N.; Heckbert, S.R.; Rosendaal, F.R.; Psaty, B.M. Serum lipid levels and the risk of venous thrombosis. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 1970–1975. [Google Scholar] [CrossRef] [PubMed]
- McColl, M.; Sattar, N.; Ellison, J.; Tait, R.; Walker, I.; Packard, C.; Greer, I. Lipoprotein (a), cholesterol and triglycerides in women with venous thromboembolism. Blood Coagul. Fibrinolysis Int. J. Haemost. Thromb. 2000, 11, 225–229. [Google Scholar]
- de Visser, M.C.; Rosendaal, F.R.; Bertina, R.M. A reduced sensitivity for activated protein C in the absence of factor V Leiden increases the risk of venous thrombosis. Blood J. Am. Soc. Hematol. 1999, 93, 1271–1276. [Google Scholar]
- Sanmarco, M.; Alessi, M.-C.; Harle, J.R.; Sapin, C.; Aillaud, M.-F.; Gentile, S.; Juhan-Vague, I.; Weiller, P.-J. Antibodies to phosphatidylethanolamine as the only antiphospholipid antibodies found in patients with unexplained thromboses. Thromb. Haemost. 2001, 85, 800–805. [Google Scholar]
- Sanmarco, M.; Gayet, S.; Alessi, M.C.; Audrain, M.; de Maistre, E.; Gris, J.C.; de Groot, P.G.; Hachulla, E.; Harlé, J.R.; Sié, P.; et al. Antiphosphatidylethanolamine antibodies are associated with an increased odds ratio for thrombosis. A multicenter study with the participation of the European Forum on antiphospholipid antibodies. Thromb. Haemost. 2007, 97, 949–954. [Google Scholar]
- Hansen-Krone, I.J.; Enga, K.F.; Südduth-Klinger, J.M.; Mathiesen, E.B.; Njølstad, I.; Wilsgaard, T.; Watkins, S.; Brækkan, S.K.; Hansen, J.-B. High fish plus fish oil intake is associated with slightly reduced risk of venous thromboembolism: The Tromsø Study. J. Nutr. 2014, 144, 861–867. [Google Scholar] [CrossRef]
- Andriamampandry, M.D.; Leray, C.; Freund, M.; Cazenave, J.-P.; Gachet, C. Antithrombotic effects of (n-3) polyunsaturated fatty acids in rat models of arterial and venous thrombosis. Thromb. Res. 1999, 93, 9–16. [Google Scholar] [CrossRef]
- Sanchez, C.; Poggi, M.; Morange, P.-E.; Defoort, C.; Martin, J.-C.; Tanguy, S.; Dutour, A.; Grino, M.; Alessi, M.-C. Diet modulates endogenous thrombin generation, a biological estimate of thrombosis risk, independently of the metabolic status. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2394–2404. [Google Scholar] [CrossRef] [PubMed]
- Kolachalama, V.B.; Shashar, M.; Alousi, F.; Shivanna, S.; Rijal, K.; Belghasem, M.E.; Walker, J.; Matsuura, S.; Chang, G.H.; Gibson, C.M. Uremic solute-aryl hydrocarbon receptor-tissue factor axis associates with thrombosis after vascular injury in humans. J. Am. Soc. Nephrol. 2018, 29, 1063–1072. [Google Scholar] [CrossRef] [PubMed]
- Voils, S.A.; Shahin, M.H.; Garrett, T.J.; Frye, R.F. Metabolomic association between venous thromboembolism in critically ill trauma patients and kynurenine pathway of tryptophan metabolism. Thromb. Res. 2018, 165, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Addi, T.; Dou, L.; Burtey, S. Tryptophan-derived uremic toxins and thrombosis in chronic kidney disease. Toxins 2018, 10, 412. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Wang, Z.; Tang, W.W.; Hazen, S.L. Gut microbe-generated trimethylamine N-oxide from dietary choline is prothrombotic in subjects. Circulation 2017, 135, 1671–1673. [Google Scholar] [CrossRef]
- Zeleznik, O.A.; Poole, E.M.; Lindstrom, S.; Kraft, P.; Van Hylckama Vlieg, A.; Lasky-Su, J.A.; Harrington, L.B.; Hagan, K.; Kim, J.; Parry, B.A.; et al. Metabolomic analysis of 92 pulmonary embolism patients from a nested case–control study identifies metabolites associated with adverse clinical outcomes. J. Thromb. Haemost. 2018, 16, 500–507. [Google Scholar] [CrossRef]
- Vanni, S.; Viviani, G.; Baioni, M.; Pepe, G.; Nazerian, P.; Socci, F.; Bartolucci, M.; Bartolini, M.; Grifoni, S. Prognostic Value of Plasma Lactate Levels Among Patients with Acute Pulmonary Embolism: The Thrombo-Embolism Lactate Outcome Study. Ann. Emerg. Med. 2013, 61, 330–338. [Google Scholar] [CrossRef]


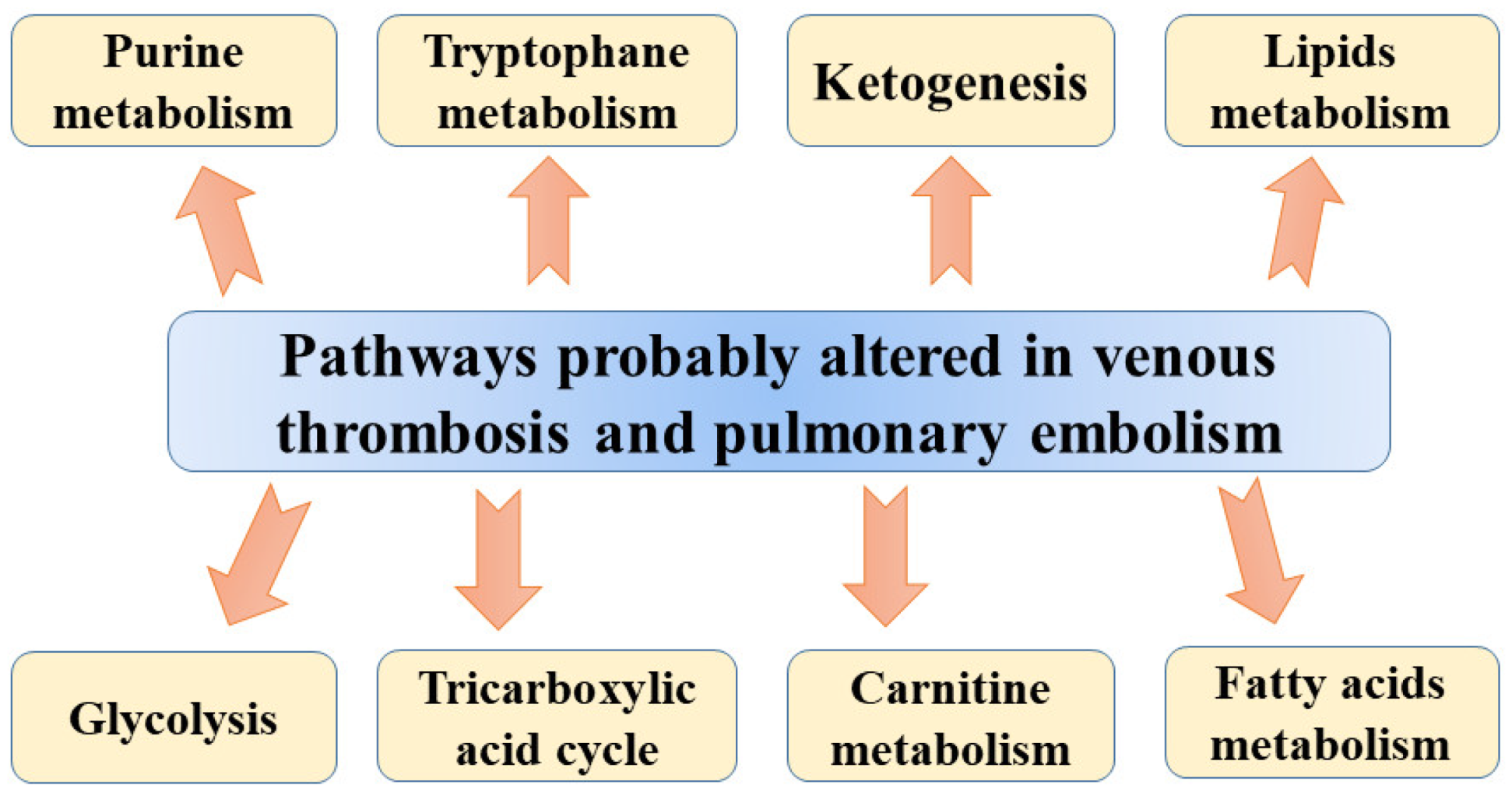{kind=link}
| Differences | Venous Thrombosis | Ref | Arterial Thrombosis | Ref |
|---|---|---|---|---|
| Causes | The occlusion of blood flow of the lower limbs | [15] | Arterial occlusion, typically triggered by erosion/rupture of an atherosclerotic plaque, or as a result of the embolization of a thrombus in the heart or other arteries. | [16,17,18] |
| Most common consequences | Acute symptoms: swelling and pain. The blood clot can disrupt and migrate causing PE. | [15] | Local tissue ischemia, and ischemic heart and ischemic stroke. | [18,19] |
| Examples of alterations in metabolomic serum profile | Altered levels of triacylglycerols, phosphatidylethanolamines, and amino acids (tryptophan) | [1] | Altered levels of xanthine and ascorbate (possible markers of atherosclerotic plaque formation). Altered levels of HDL, choline, taurine, glycine and glucose (specific biomarkers of the initial state of the disease). | [20] |
| Reduced levels of adenosine, adenine, and intermediate molecules of the tricarboxylic acid (citrate, succinate and fumarate). In creased levels of L-carnitine, sphingomyelins, phosphatidylcholines and triglycerides. | [5] | Methionine, alanine and valine (disease progression). | [21] | |
| Altered levels of acylcarnitines (palmitoleoyl carnitine, and decanoyl carnitine) | [15] | Altered levels of citric acid, 4-hydroxyproline, aspartic acid, and fructose, L-alanine, L-arabitol, scyllo-inositol, 2-hydroxyphenilacetic acid, 3-hydroxybutiryc acid and N-acetylneuraminic acid, short-chain dicarboxylacylcarnitine and TMAO. | [22] | |
| Increased levels of glutamine, phenylalanine, and proline | [23] | Altered levels of phenylalanine and monounsaturated fatty acids. | [24] | |
| Altered levels of betaine and/or trimethylamine N-oxide (TMAO). Altered levels of n-3 long-chain fatty acid lipids. | [25] |
| Venous Thrombosis | ||||
|---|---|---|---|---|
| Metabolite Type | Human Studies | Ref | Animal Studies | Ref |
| Central carbon metabolites (lactic acid, citric acid, glucose 6-phosphate) | No data found | - | Sample: Rabbit venous blood and jugular venous thrombus Results: ↑ lactic acid and ↓ glycolytic metabolite levels (citrate acid, glucose 6-phosphate) in VT vs. VB (p < 0.05)– possibly related to enhanced glycolysis in intrathrombus erythrocytes | [34] |
| Sample: Serum from DVT mice Results: ↓ tricyclic acid cycle (TCA) intermediates: citrate (1.5 times), succinate (2.3 times), and fumarate (2.8 times) | [5] | |||
| Purine nucleotides and their metabolites (hypoxanthine, guanine, AMP, guanine monophosphate (GMP) | No data found | - | Sample: Rabbit venous blood and jugular venous thrombus Results: ↑ levels of AMP, GMP, hypoxanthine, and guanine levels in VT vs. VB (p < 0.05). | [34] |
| - | Sample: Serum from DVT mice Results: ↓ adenosine (9.6 fold in DVT), adenine (10.6 fold) Sample: Vein wall from DVT mice Results: ↑ adenosine in vein wall (2.2 fold) | [5] | ||
| Tryptophan and its metabolites (serotonin, 3-hydroxykynurenine, tryptophan) | No data found | - | Sample: Rabbit venous blood and jugular venous thrombus Results: ↑ levels of tryptophan metabolites (serotonin and kynurenine pathway metabolites) in VT vs. VB (p < 0.05). ↓ tryptophan level in VT (p < 0.05). | [34] |
| Other amino acids | No data found | - | Sample: Mice model of VT-whole blood Results: ↑ glutamine, phenylalanine, and proline in old mice vs. young mice | [23] |
| Lipid metabolism, lipids, fatty acids (choline, ethanolamine phosphate, L-carnitine, sphingomyelins) | Sample: Idiopathic adult VTE cases (plasma) Results: ↓ long-chain ACs (10:1, 12:0, 12:2, 18:1, and 18:2) (p = 0.01, 0.004, 0.001, 0.04, and 0.02, respectively) | [12] | Sample: Rabbit venous blood and jugular venous thrombus Results: ↑ choline levels in VT vs. VB (p < 0.05). | [34] |
| Sample: Adults with incident VTE cases (blood) Results: altered levels of 12 diacylglycerols, 42 triacylglycerols, 23 phosphatidylethanolamines, 36 phosphatidylcholines, 12 cholesterol esters, 7 sphingomyelins, 9 lysophosphatidylcholines, 6 lysophosphatidylethanolamines, and 22 carnitines | [1] | Sample: Serum from DVT mice Results: ↑ levels of L-carnitine (67.0 fold change), sphingomyelins (1.5 fold), phosphatidylcholines, and triglycerides (1.8 fold) Sample: vein wall of DVT mice Results: ↑ levels of ceramide and sphingomyelins | [5] | |
| Sample: Fasting plasma samples from patients with VTE Results: ↑ levels of phosphatidylcholines and triglycerides containing fatty acyl moieties composed of long-chain polyunsaturated fatty acids of both the n-6 and n-3 series | [25] | |||
| Other | Sample: Fasting plasma samples from patients with VTE Results: ↑ TMAO levels (2 fold) | [25] | Sample: Rabbit venous blood and jugular venous thrombus Results: ↑ choline levels in VT vs. VB (p < 0.05). | [34] |
| Pulmonary Embolism | ||||
| Central carbon metabolites (lactic acid, citric acid, glucose 6-phosphate) | Sample: Patients with low-risk PE vs. intermediate/high-risk PE Results: ↓alpha-ketoglutarate, malate, isocitrate, fumarate and cis-aconitate in high-risk patients | [101] | Sample: Pig model of pulmonary embolism (serum) Results: ↑ citrate, malate, fumarate, and α-ketoglutarate-associated with a reduced TCA cycle turnover ↑ pyruvate and lactate-shift in glucose metabolism, beneficial for the generation of ATP and the prevention of mitochondrial ROS production | [11] |
| Purine nucleotides and their metabolites (hypoxanthine, guanine, AMP, guanine monophosphate (GMP) | Sample: Patients with low-risk PE vs. intermediate/high-risk PE Results: ↑ xanthosine and hypoxanthine in intermediate/high-risk PE | [101] | No data found | |
| Lipid metabolism, lipids, fatty acids (choline, ethanolamine phosphate, L-carnitine, sphingomyelins) | Sample: Adults with incident VTE cases (blood) Results: 12 diacylglycerols, 42 triacylglycerols, 23 phosphatidylethanolamines, 36 phosphatidylcholines, 12 cholesterol esters, 7 sphingomyelins, 9 lysophosphatidylcholines, 6 lysophosphatidylethanolamines, and 22 carnitines | [1] | Sample: Pig model of pulmonary embolism (serum) Results: ↑ free fatty acids (FFA), such as docosatetraenoic and docosapentaenoic acids, as well as palmitic and oleic acids-increased lipolysis ↑ leukotriene C4 and ↓ in dihydroxyoctadecadienoic acid, hydroxyoctadecenoic acid, and oxoheptadecatrienoic acid ↓ sphingomyelin and ceramide-1-phosphate and ↑ sphingosine | [11] |
| Sample: Patients with low-risk PE vs. intermediate/high-risk PE Results: Enriched fatty acid metabolism (Acyl Carnitine) pathway ↓ arachidonoylcholine, oleoylcholine, palmitoylcholine, docosahexaenoylcholine and dihomo-linolenoyl-choline in the intermediate/high-risk group | [101] | No data found | ||
| Other | No data found | Sample: Pig model of pulmonary embolism (serum) Results: ↑ β-hydroxybutyrate (β-HB) and acetoacetate (ketone bodies) -probably due to hypoxia-mediated alterations | [11] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franczyk, B.; Gluba-Brzózka, A.; Ławiński, J.; Rysz-Górzyńska, M.; Rysz, J. Metabolomic Profile in Venous Thromboembolism (VTE). Metabolites 2021, 11, 495. https://doi.org/10.3390/metabo11080495
Franczyk B, Gluba-Brzózka A, Ławiński J, Rysz-Górzyńska M, Rysz J. Metabolomic Profile in Venous Thromboembolism (VTE). Metabolites. 2021; 11(8):495. https://doi.org/10.3390/metabo11080495
Chicago/Turabian StyleFranczyk, Beata, Anna Gluba-Brzózka, Janusz Ławiński, Magdalena Rysz-Górzyńska, and Jacek Rysz. 2021. "Metabolomic Profile in Venous Thromboembolism (VTE)" Metabolites 11, no. 8: 495. https://doi.org/10.3390/metabo11080495
APA StyleFranczyk, B., Gluba-Brzózka, A., Ławiński, J., Rysz-Górzyńska, M., & Rysz, J. (2021). Metabolomic Profile in Venous Thromboembolism (VTE). Metabolites, 11(8), 495. https://doi.org/10.3390/metabo11080495