
Hypoxia Performance Curve: Assess a Whole-Organism Metabolic Shift from a Maximum Aerobic Capacity towards a Glycolytic Capacity in Fish

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
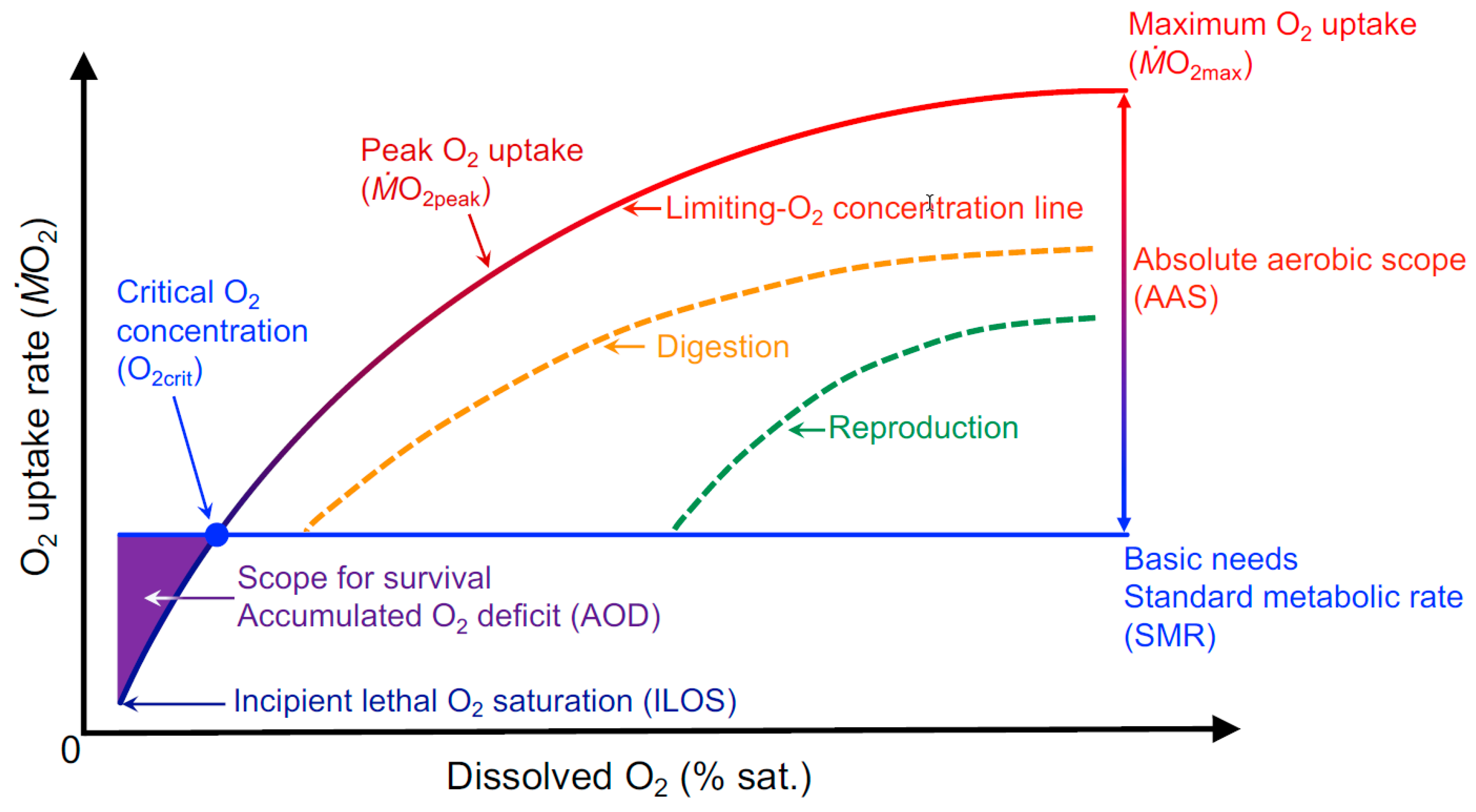
2. Current Hypoxia Tolerance Assessments Explore Only a Small Portion of Hypoxic Performance Curve

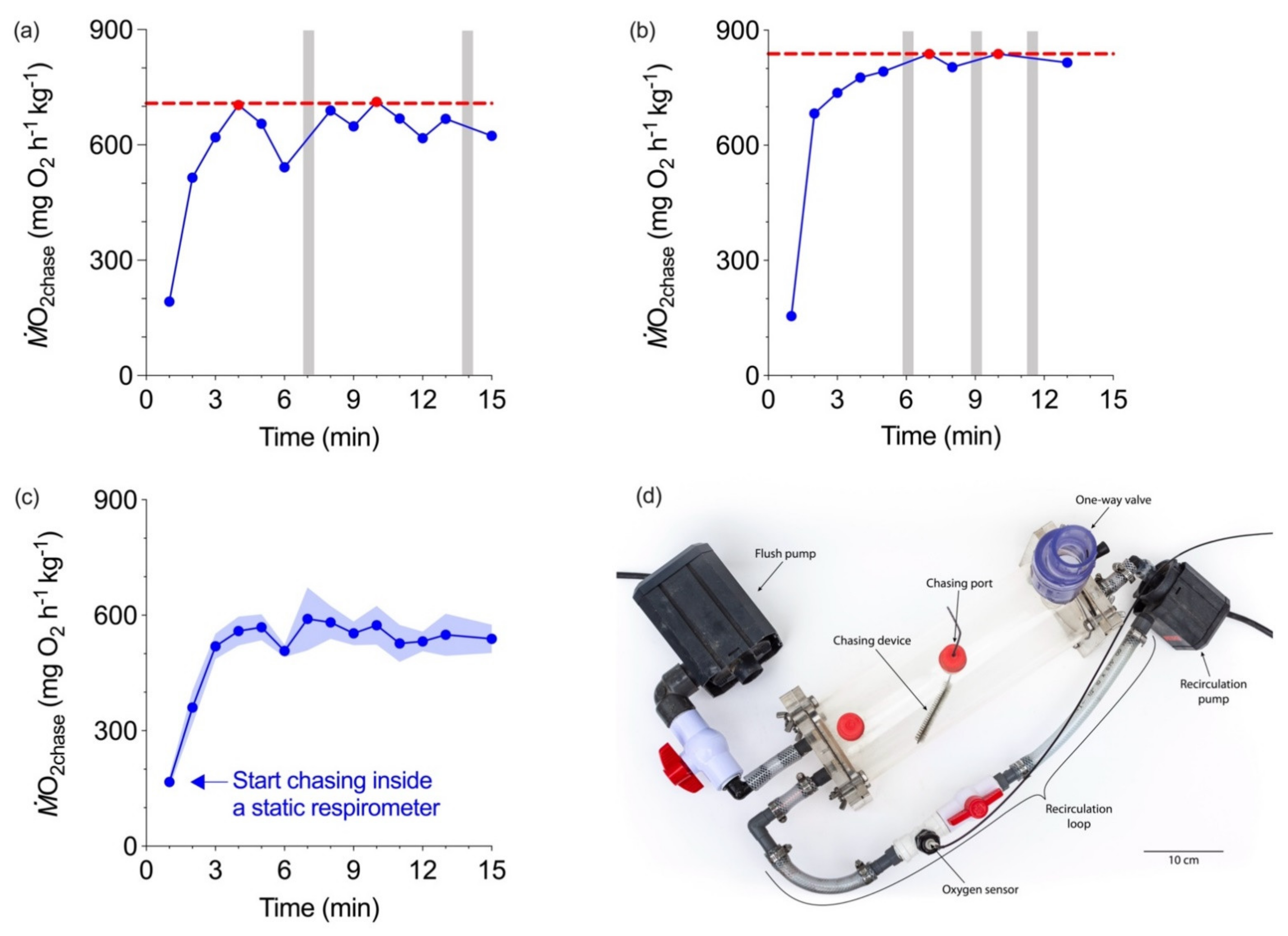
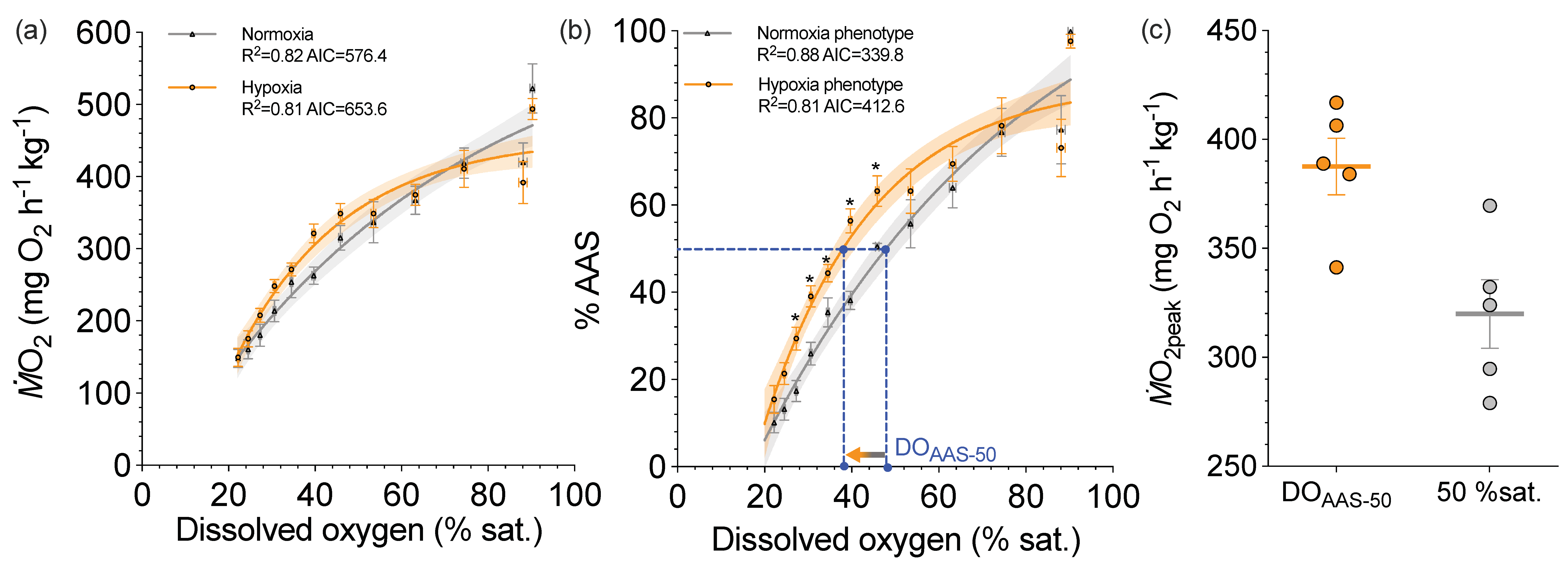
3. Individualizing the Limiting-Oxygen Concentration (LOC) Performance Curve
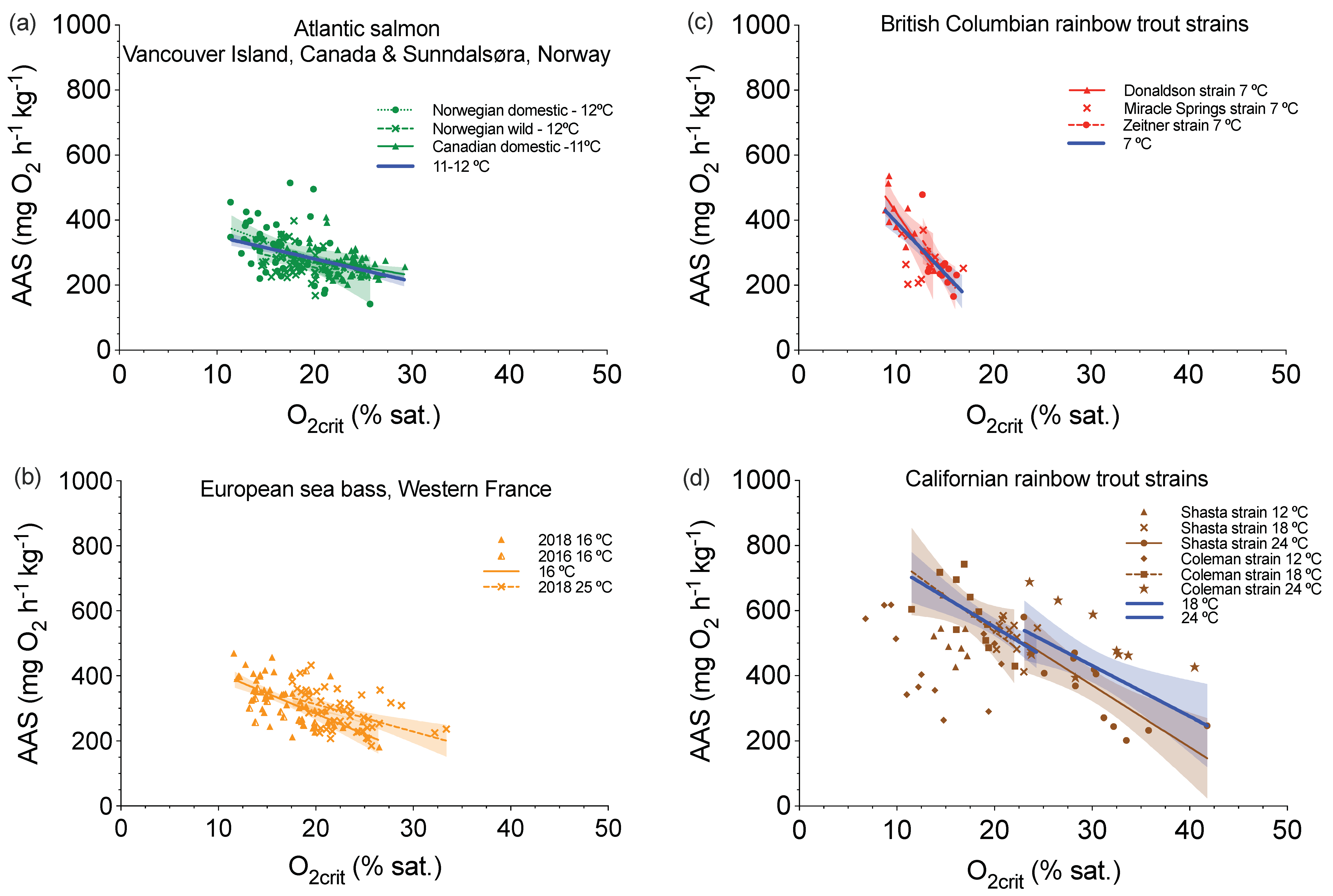
4. A Benefit of Characterizing Aerobic and Glycolytic Capacities on the Same Individuals
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AAS | absolute aerobic scope |
| AOD | accumulated O2 deficit |
| CaO2 | arterial blood O2 content |
| Ccrit | critical O2 concentration |
| CvO2 | venous O2 content |
| DO | dissolved O2 level |
| DOAAS-50 | the water-dissolved O2 that supports 50% of the individual’s AAS |
| ILOS | incipient lethal O2 saturation |
| LOC | limiting-O2 concentration |
| MMR | maximum metabolic rate |
| ṀO2max | maximum O2 uptake |
| ṀO2 | O2 uptake |
| ṀO2peak | peak O2 uptake |
| O2 | oxygen |
| O2crit | critical dissolved O2 |
| P50 | a partial O2 pressure where hemoglobin is 50% saturated with O2 |
| Pcrit | critical O2 tension |
| PO2 | partial pressure of O2, or O2 tension |
| RMR | routine metabolic rate |
| SMR | standard metabolic rate |
| Ucrit | critical swimming speed |
References
- Eliason, E.J.; Clark, T.; Hague, M.J.; Hanson, L.M.; Gallagher, Z.S.; Jeffries, K.M.; Gale, M.K.; Patterson, D.A.; Hinch, S.G.; Farrell, A.P. Differences in Thermal Tolerance Among Sockeye Salmon Populations. Science 2011, 332, 109–112. [Google Scholar] [CrossRef]
- Crossin, G.T.; Hinch, S.G.; Cooke, S.J.; Welch, D.W.; Patterson, D.A.; Jones, S.; Lotto, A.G.; Leggatt, R.A.; Mathes, M.T.; Shrimpton, J.M.; et al. Exposure to high temperature influences the behaviour, physiology, and survival of sockeye salmon during spawning migration. Can. J. Zool. 2008, 86, 127–140. [Google Scholar] [CrossRef]
- Eliason, E.J.; Farrell, A.P. Oxygen uptake in Pacific salmon Oncorhynchus spp.: When ecology and physiology meet. J. Fish Biol. 2016, 88, 359–388. [Google Scholar] [CrossRef] [PubMed]
- Schmidtko, S.; Stramma, L.; Visbeck, M. Decline in global oceanic oxygen content during the past five decades. Nat. Cell Biol. 2017, 542, 335–339. [Google Scholar] [CrossRef]
- Bond, N.A.; Cronin, M.F.; Freeland, H.; Mantua, N. Causes and impacts of the 2014 warm anomaly in the NE Pacific. Geophys. Res. Lett. 2015, 42, 3414–3420. [Google Scholar] [CrossRef]
- Clack, J.A. Devonian climate change, breathing, and the origin of the tetrapod stem group. Integr. Comp. Biol. 2007, 47, 510–523. [Google Scholar] [CrossRef]
- Randall, D.J.; Rummer, J.L.; Wilson, J.; Wang, S.; Brauner, C.J. A unique mode of tissue oxygenation and the adaptive radiation of teleost fishes. J. Exp. Biol. 2014, 217, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Stecyk, J.A.W. Cardiovascular responses to limiting oxygen levels. In Fish Physiology; Gamperl, A.K., Gillis, T.E., Farrell, A.P., Brauner, C.J., Eds.; Academic Press: Cambridge, MA, USA, 2017; Chapter 5; Volume 36, pp. 299–371. [Google Scholar] [CrossRef]
- Chabot, D.; Claireaux, G. Ecophysiology. In Atlantic Cod; John Wiley & Sons: Hoboken, NJ, USA, 2018; pp. 27–86. [Google Scholar] [CrossRef]
- Fry, F.E.J.; Hart, J.S. The Relation of Temperature to Oxygen Consumption in the Goldfish. Biol. Bull. 1948, 94, 66–77. [Google Scholar] [CrossRef]
- Fry, F. The Effect of Environmental Factors on the Physiology of Fish. Fish Physiol. 1971, 6, 1–98. [Google Scholar] [CrossRef]
- Gamperl, A.K.; Driedzic, W. Cardiovascular Function and Cardiac Metabolism. In Fish Physiology; Richards, J.G., Farrell, A.P., Brauner, C.J., Eds.; Academic Press: Cambridge, MA, USA, 2009; Chapter 7; Volume 27, pp. 301–360. [Google Scholar] [CrossRef]
- Holeton, G.F.; Randall, D.J. The Effect of Hypoxia Upon the Partial Pressure of Gases in the Blood and Water Afferent and Efferent to the Gills of Rainbow Trout. J. Exp. Biol. 1967, 46, 317–327. [Google Scholar] [CrossRef]
- Hughes, G.M.; Sunders, R.L. Responses of the Respiratory Pumps to Hypoxia in the Rainbow Trout (Salmo Gairdneri). J. Exp. Biol. 1970, 53, 529–545. [Google Scholar] [CrossRef]
- Sundin, L.I. Responses of the branchial circulation to hypoxia in the Atlantic cod, Gadus morhua. Am. J. Physiol. Integr. Comp. Physiol. 1995, 268, R771–R778. [Google Scholar] [CrossRef] [PubMed]
- Randall, D.; Shelton, G. The effects of changes in environmental gas concentrations on the breathing and heart rate of a teleost fish. Comp. Biochem. Physiol. 1963, 9, 229–239. [Google Scholar] [CrossRef]
- Butler, P.J.; Taylor, E.W. The effect of progressive hypoxia on respiration in the dogfish (scyliorhinus canicula) at different seasonal temperatures. J. Exp. Biol. 1975, 63, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Randall, D. The Control of Respiration and Circulation in Fish During Exercise and Hypoxia. J. Exp. Biol. 1982, 100, 275–288. [Google Scholar] [CrossRef]
- Farrell, A.P. Tribute to P. L. Lutz: A message from the heart—Why hypoxic bradycardia in fishes? J. Exp. Biol. 2007, 210, 1715–1725. [Google Scholar] [CrossRef]
- Sandblom, E.; Axelsson, M. Effects of hypoxia on the venous circulation in rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2005, 140, 233–239. [Google Scholar] [CrossRef]
- Sandblom, E.; Axelsson, M. Adrenergic control of venous capacitance during moderate hypoxia in the rainbow trout (Oncorhynchus mykiss): Role of neural and circulating catecholamines. Am. J. Physiol. Integr. Comp. Physiol. 2006, 291, R711–R718. [Google Scholar] [CrossRef][Green Version]
- Scott, A.L.; Rogers, W.A. Haematological effects of prolonged sublethal hypoxia on channel catfish* Ictalurus punctatus (Rafinesque). J. Fish Biol. 1981, 18, 591–601. [Google Scholar] [CrossRef]
- Soldatov, A. The effect of hypoxia on red blood cells of flounder: A morphologic and autoradiographic study. J. Fish Biol. 1996, 48, 321–328. [Google Scholar] [CrossRef]
- Farrell, A.P.; Richards, J.G. Defining hypoxia: An integrative synthesis of the responses of fish to hypoxia. In Fish Physiology; Richards, J.G., Farrell, A.P., Brauner, C.J., Eds.; Academic Press: Cambridge, MA, USA, 2009; Chapter 11; Volume 27, pp. 487–503. [Google Scholar] [CrossRef]
- Schwerte, T.; Überbacher, D.; Pelster, B. Non-invasive imaging of blood cell concentration and blood distribution in zebrafish Danio rerio incubated in hypoxic conditions in vivo. J. Exp. Biol. 2003, 206, 1299–1307. [Google Scholar] [CrossRef] [PubMed]
- Hochachka, P.W. Defense strategies against hypoxia and hypothermia. Science 1986, 231, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Sollid, J.; De Angelis, P.; Gundersen, K.; Nilsson, G.E. Hypoxia induces adaptive and reversible gross morphological changes in crucian carp gills. J. Exp. Biol. 2003, 206, 3667–3673. [Google Scholar] [CrossRef] [PubMed]
- Brauner, C.J.; Rombough, P.J. Ontogeny and paleophysiology of the gill: New insights from larval and air-breathing fish. Respir. Physiol. Neurobiol. 2012, 184, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Anttila, K.; Lewis, M.; Prokkola, J.; Kanerva, M.; Seppänen, E.; Kolari, I.; Nikinmaa, M. Warm acclimation and oxygen depletion induce species-specific responses in salmonids. J. Exp. Biol. 2015, 218, 1471–1477. [Google Scholar] [CrossRef]
- Gamperl, A.K.; Todgham, A.E.; Parkhouse, W.S.; Dill, R.; Farrell, A.P. Recovery of trout myocardial function following anoxia: Preconditioning in a non-mammalian model. Am. J. Physiol. Integr. Comp. Physiol. 2001, 281, R1755–R1763. [Google Scholar] [CrossRef]
- Gillis, T.E.; Johnston, E.F. Cardiac Preconditioning, Remodeling and Regeneration. In Fish Physiology; Gamperl, A.K., Gillis, T.E., Farrell, A.P., Brauner, C.J., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 36, pp. 185–233. [Google Scholar] [CrossRef]
- McKenzie, D.J.; Wong, S.; Randall, D.J.; Egginton, S.; Taylor, E.W.; Farrell, A.P. The effects of sustained exercise and hypoxia upon oxygen tensions in the red muscle of rainbow trout. J. Exp. Biol. 2004, 207, 3629–3637. [Google Scholar] [CrossRef]
- McKenzie, D.J.; Zhang, Y.; Eliason, E.J.; Schulte, P.M.; Claireaux, G.; Blasco, F.R.; Nati, J.J.; Farrell, A.P. Intraspecific variation in tolerance of warming in fishes. J. Fish Biol. 2020. [Google Scholar] [CrossRef]
- Lefevre, S.; Wang, T.; McKenzie, D.J. The role of mechanistic physiology in investigating impacts of global warming on fishes. J. Exp. Biol. 2021, 224. [Google Scholar] [CrossRef] [PubMed]
- Seibel, B.A.; Deutsch, C. Oxygen supply capacity in animals evolves to meet maximum demand at the current oxygen partial pressure regardless of size or temperature. J. Exp. Biol. 2020, 223. [Google Scholar] [CrossRef] [PubMed]
- Fry, F. Effects of the environment on animal activity. Publ. Ont. Fish. Res. Lab. 1947, 55, 1–62. [Google Scholar]
- Ultsch, G.R.; Boschung, H.; Ross, M.J.; Ross, H.B.J. Metabolism, Critical Oxygen Tension, and Habitat Selection in Darters (Etheostoma). Ecology 1978, 59, 99–107. [Google Scholar] [CrossRef]
- Ultsch, G.R.; Regan, M.D. The utility and determination of Pcrit in fishes. J. Exp. Biol. 2019, 222. [Google Scholar] [CrossRef] [PubMed]
- Wood, C.M. The fallacy of the Pcrit—Are there more useful alternatives? J. Exp. Biol. 2018, 221. [Google Scholar] [CrossRef] [PubMed]
- Mueller, C.A.; Seymour, R.S. The Regulation Index: A New Method for Assessing the Relationship between Oxygen Con-sumption and Environmental Oxygen. Physiol. Biochem. Zool. 2011, 84, 522–532. [Google Scholar] [CrossRef]
- Chabot, D.; Steffensen, J.F.; Farrell, A.P. The determination of standard metabolic rate in fishes. J. Fish Biol. 2016, 88, 81–121. [Google Scholar] [CrossRef] [PubMed]
- Steffensen, J.F. Some errors in respirometry of aquatic breathers: How to avoid and correct for them. Fish Physiol. Biochem. 1989, 6, 49–59. [Google Scholar] [CrossRef]
- Steffensen, J.F.; Bushnell, P.G.; Schurmann, H. Oxygen consumption in four species of teleosts from Greenland: No evidence of metabolic cold adaptation. Polar Biol. 1994, 14, 49–54. [Google Scholar] [CrossRef]
- Reemeyer, J.E.; Rees, B.B. Standardizing the determination and interpretation of Pcrit in fishes. J. Exp. Biol. 2019, 222. [Google Scholar] [CrossRef]
- Negrete, B.; Esbaugh, A.J. A methodological evaluation of the determination of critical oxygen threshold in an estuarine teleost. Biol. Open 2019, 8. [Google Scholar] [CrossRef]
- Claireaux, G.; Chabot, D. Responses by fishes to environmental hypoxia: Integration through Fry’s concept of aerobic metabolic scope. J. Fish Biol. 2016, 88, 232–251. [Google Scholar] [CrossRef]
- Chabot, D.; Zhang, Y.; Farrell, A.P. Valid oxygen uptake measurements: Using high r 2 values with good intentions can bias upward the determination of standard metabolic rate. J. Fish Biol. 2021, 98, 1206–1216. [Google Scholar] [CrossRef]
- Réale, D.; Garant, D.; Humphries, M.M.; Bergeron, P.; Careau, V.; Montiglio, P.-O. Personality and the emergence of the pace-of-life syndrome concept at the population level. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 4051–4063. [Google Scholar] [CrossRef] [PubMed]
- Polverino, G.; Santostefano, F.; Díaz-Gil, C.; Mehner, T. Ecological conditions drive pace-of-life syndromes by shaping rela-tionships between life history, physiology and behaviour in two populations of Eastern mosquitofish. Sci. Rep. 2018, 8, 14673. [Google Scholar] [CrossRef]
- Seibel, B.A.; Andres, A.; Birk, M.A.; Burns, A.L.; Shaw, C.T.; Timpe, A.W.; Welsh, C.J. Oxygen supply capacity breathes new life into critical oxygen partial pressure (Pcrit). J. Exp. Biol. 2021, 224. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y. Interpreting Species, Intraspecific and Intra-Individual Variability by Comprehensively Characterizing a Fish’s Respiratory Phenotype with Valid Measures of Oxygen Uptake; University of British Columbia: Vancouver, BC, Canada, 2021. [Google Scholar] [CrossRef]
- Neill, W.H.; Miller, J.M.; Van Der Veer, H.W.; Winemiller, K.O. Ecophysiology of marine fish recruitment: A conceptual framework for understanding interannual variability. Neth. J. Sea Res. 1994, 32, 135–152. [Google Scholar] [CrossRef]
- Claireaux, G.; Lagardère, J.-P. Influence of temperature, oxygen and salinity on the metabolism of the European sea bass. J. Sea Res. 1999, 42, 157–168. [Google Scholar] [CrossRef]
- Pörtner, H.O.; Farrell, A.P. Physiology and Climate Change. Science 2008, 322, 690–692. [Google Scholar] [CrossRef] [PubMed]
- Hochachka, P.W. Scope for survival: A conceptual “Mirror” to Fry’s scope for activity. Trans. Am. Fish. Soc. 1990, 119, 622–628. [Google Scholar] [CrossRef]
- Richards, J.G.; Sardella, B.A.; Schulte, P.M. Regulation of pyruvate dehydrogenase in the common killifish, Fundulus hetero-clitus, during hypoxia exposure. Am. J. Physiol. Integr. Comp. Physiol. 2008, 295, R979–R990. [Google Scholar] [CrossRef]
- McBryan, T.L.; Healy, T.M.; Haakons, K.L.; Schulte, P.M. Warm acclimation improves hypoxia tolerance in Fundulus hetero-clitus. J. Exp. Biol. 2016, 219, 474–484. [Google Scholar] [CrossRef]
- Regan, M.D.; Richards, J.G. Rates of hypoxia induction alter mechanisms of O2 uptake and the critical O2 tension of goldfish. J. Exp. Biol. 2017, 220, 2536–2544. [Google Scholar] [CrossRef]
- Milinkovitch, T.; Marras, S.; Antognarelli, F.; Lefrançois, C.; Le Floch, S.; Domenici, P. The effects of hypoxia on aerobic metabolism in oil-contaminated sea bass (Dicentrarchus labrax). Chemosphere 2020, 253, 126678. [Google Scholar] [CrossRef]
- Hochachka, P.W.; Buck, L.T.; Doll, C.J.; Land, S.C. Unifying theory of hypoxia tolerance: Molecular/metabolic defense and rescue mechanisms for surviving oxygen lack. Proc. Natl. Acad. Sci. USA 1996, 93, 9493–9498. [Google Scholar] [CrossRef]
- Stecyk, J.A.; Galli, G.L.; Shiels, H.A.; Farrell, A.P. Cardiac survival in anoxia-tolerant vertebrates: An electrophysiological perspective. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol. 2008, 148, 339–354. [Google Scholar] [CrossRef] [PubMed]
- Speers-Roesch, B.; Sandblom, E.; Lau, G.Y.; Farrell, A.P.; Richards, J.G. Effects of environmental hypoxia on cardiac energy metabolism and performance in tilapia. Am. J. Physiol. Integr. Comp. Physiol. 2010, 298, R104–R119. [Google Scholar] [CrossRef] [PubMed]
- Lefevre, S.; Stecyk, J.; Torp, M.-K.; Løvold, L.Y.; Sørensen, C.; Johansen, I.B.; Stensløkken, K.-O.; Couturier, C.S.; Sloman, K.A.; Nilsson, G.E. Re-oxygenation after anoxia induces brain cell death and memory loss in the anoxia-tolerant crucian carp. J. Exp. Biol. 2017, 220, 3883–3895. [Google Scholar] [CrossRef]
- Speers-Roesch, B.; Norin, T.; Driedzic, W.R. The benefit of being still: Energy savings during winter dormancy in fish come from inactivity and the cold, not from metabolic rate depression. Proc. R. Soc. B Boil. Sci. 2018, 285, 20181593. [Google Scholar] [CrossRef] [PubMed]
- Claireaux, G.; Théron, M.; Prineau, M.; Dussauze, M.; Merlin, F.-X.; Le Floch, S. Effects of oil exposure and dispersant use upon environmental adaptation performance and fitness in the European sea bass, Dicentrarchus labrax. Aquat. Toxicol. 2013, 130–131, 160–170. [Google Scholar] [CrossRef]
- Nonnotte, G.; Maxime, V.; Truchot, J.; Williot, P.; Peyraud, C. Respiratory responses to progressive ambient hypoxia in the sturgeon, Acipenser baeri. Respir. Physiol. 1993, 91, 71–82. [Google Scholar] [CrossRef]
- Maxime, V.; Pichavant, K.; Boeuf, G.; Nonnotte, G. Effects of hypoxia on respiratory physiology of turbot, Scophthalmus maximus. Fish Physiol. Biochem. 2000, 22, 51–59. [Google Scholar] [CrossRef]
- Routley, M.H.; Nilsson, G.E.; Renshaw, G.M. Exposure to hypoxia primes the respiratory and metabolic responses of the epaulette shark to progressive hypoxia. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2002, 131, 313–321. [Google Scholar] [CrossRef]
- Milligan, C. Metabolic recovery from exhaustive exercise in rainbow trout. Comp. Biochem. Physiol. Part A Physiol. 1996, 113, 51–60. [Google Scholar] [CrossRef]
- Jain, K.E.; Farrell, A.P. Influence of seasonal temperature on the repeat swimming performance of rainbow trout Oncorhynchus mykiss. J. Exp. Biol. 2003, 206, 3569–3579. [Google Scholar] [CrossRef]
- Black, E.C. Hyperactivity as a Lethal Factor in Fish. J. Fish. Res. Board Can. 1958, 15, 573–586. [Google Scholar] [CrossRef]
- Wood, C.; Turner, J.D.; Graham, M.S. Why do fish die after severe exercise? J. Fish Biol. 1983, 22, 189–201. [Google Scholar] [CrossRef]
- Shoubridge, E.A.; Hochachka, P.W. Ethanol: Novel end product of vertebrate anaerobic metabolism. Science 1980, 209, 308–309. [Google Scholar] [CrossRef] [PubMed]
- Stecyk, J.A.W.; Larsen, B.C.; Nilsson, G.E. Intrinsic contractile properties of the crucian carp (Carassius carassius) heart during anoxic and acidotic stress. Am. J. Physiol. Integr. Comp. Physiol. 2011, 301, R1132–R1142. [Google Scholar] [CrossRef]
- Vornanen, M.; Stecyk, J.; Nilsson, G.E. The Anoxia-Tolerant Crucian Carp (Carassius carassius L.). In Fish Physiology; Richards, J.G., Farrell, A.P., Brauner, C.J., Eds.; Academic Press: Cambridge, MA, USA, 2009; Chapter 9; Volume 27, pp. 397–441. [Google Scholar] [CrossRef]
- Jackson, D.C. How a Turtle’s Shell Helps It Survive Prolonged Anoxic Acidosis. Physiology 2000, 15, 181–185. [Google Scholar] [CrossRef]
- Cox, G.K.; Sandblom, E.; Richards, J.G.; Farrell, A.P. Anoxic survival of the Pacific hagfish (Eptatretus stoutii). J. Comp. Physiol. B 2010, 181, 361–371. [Google Scholar] [CrossRef]
- Zhang, Y.; Mauduit, F.; Farrell, A.P.; Chabot, D.; Ollivier, H.; Rio-Cabello, A.; Le Floch, S.; Claireaux, G. Exposure of European sea bass (Dicentrarchus labrax) to chemically dispersed oil has a chronic residual effect on hypoxia tolerance but not aerobic scope. Aquat. Toxicol. 2017, 191, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Ultsch, G.R.; Jackson, D.C. Long-term submergence at 3 degrees C of the turtle Chrysemys picta bellii in normoxic and severely hypoxic water. III. Effects of changes in ambient PO2 and subsequent air breathing. J. Exp. Biol. 1982, 97, 87–99. [Google Scholar] [CrossRef]
- Bushnell, P.G.; Steffensen, J.F.; Johansen, K. Oxygen Consumption and Swimming Performance in Hypoxia-Acclimated Rainbow Trout Salmo Gairdneri. J. Exp. Biol. 1984, 113, 225–235. [Google Scholar] [CrossRef]
- Motyka, R.; Norin, T.; Petersen, L.H.; Huggett, D.B.; Gamperl, A.K. Long-term hypoxia exposure alters the cardiorespiratory physiology of steelhead trout (Oncorhynchus mykiss), but does not affect their upper thermal tolerance. J. Therm. Biol. 2017, 68, 149–161. [Google Scholar] [CrossRef]
- Regan, M.D.; Gill, I.; Richards, J.G. Calorespirometry reveals that goldfish prioritize aerobic metabolism over metabolic rate depression in all but near-anoxic environments. J. Exp. Biol. 2017, 220, 564–572. [Google Scholar] [CrossRef]
- Borowiec, B.G.; McClelland, G.B.; Rees, B.B.; Scott, G.R. Distinct metabolic adjustments arise from acclimation to constant hypoxia and intermittent hypoxia in estuarine killifish (Fundulus heteroclitus). J. Exp. Biol. 2018, 221, jeb190900. [Google Scholar] [CrossRef] [PubMed]
- Stecyk, J.A.W.; Farrell, A.P. Regulation of the Cardiorespiratory System of Common Carp (Cyprinus carpio) during Severe Hypoxia at Three Seasonal Acclimation Temperatures. Physiol. Biochem. Zool. 2006, 79, 614–627. [Google Scholar] [CrossRef] [PubMed]
- Lague, S.L.; Speers-Roesch, B.; Richards, J.G.; Farrell, A.P. Exceptional cardiac anoxia tolerance in tilapia (Oreochromis hybrid). J. Exp. Biol. 2012, 215, 1354–1365. [Google Scholar] [CrossRef]
- Claireaux, G.; Webber, D.M.; Lagardère, J.-P.; Kerr, S.R. Influence of water temperature and oxygenation on the aerobic metabolic scope of Atlantic cod (Gadus morhua). J. Sea Res. 2000, 44, 257–265. [Google Scholar] [CrossRef]
- Lefrançois, C.; Claireaux, G. Influence of ambient oxygenation and temperature on metabolic scope and scope for heart rate in the common sole Solea solea. Mar. Ecol. Prog. Ser. 2003, 259, 273–284. [Google Scholar] [CrossRef]
- Zhang, Y.; Gilbert, M.J.H.; Farrell, A.P. Measuring maximum oxygen uptake with an incremental swimming test and by chasing rainbow trout to exhaustion inside a respirometry chamber yields the same results. J. Fish Biol. 2020, 97, 28–38. [Google Scholar] [CrossRef]
- Zhang, Y.; Timmerhaus, G.; Anttila, K.; Mauduit, F.; Jørgensen, S.M.; Kristensen, T.; Claireaux, G.; Takle, H.; Farrell, A.P. Domestication compromises athleticism and respiratory plasticity in response to aerobic exercise training in Atlantic salmon (Salmo salar). Aquaculture 2016, 463, 79–88. [Google Scholar] [CrossRef]
- Zhang, Y.; Gilbert, M.J.H.; Farrell, A.P. Finding the peak of dynamic oxygen uptake during fatiguing exercise in fish. J. Exp. Biol. 2019, 222, jeb196568. [Google Scholar] [CrossRef]
- Andersson, M.L.; Sundberg, F.; Eklöv, P. Chasing away accurate results: Exhaustive chase protocols underestimate maximum metabolic rate estimates in European perch Perca fluviatilis. J. Fish Biol. 2020, 97, 1644–1650. [Google Scholar] [CrossRef] [PubMed]
- Gj∅Saeter, J. Mesopelagic fish, a large potential resource in the Arabian Sea. Deep Sea Res. Part A Oceanogr. Res. Pap. 1984, 31, 1019–1035. [Google Scholar] [CrossRef]
- MacKenzie, B.R.; Mariani, P. Spawning of Bluefin Tuna in the Black Sea: Historical Evidence, Environmental Constraints and Population Plasticity. PLoS ONE 2012, 7, e39998. [Google Scholar] [CrossRef]
- Petersen, L.H.; Gamperl, A.K. Effect of acute and chronic hypoxia on the swimming performance, metabolic capacity and cardiac function of Atlantic cod (Gadus morhua). J. Exp. Biol. 2010, 213, 808–819. [Google Scholar] [CrossRef]
- Petersen, L.H.; Gamperl, A.K. Cod (Gadus morhua) Cardiorespiratory Physiology and Hypoxia Tolerance following Acclimation to Low-Oxygen Conditions. Physiol. Biochem. Zool. 2011, 84, 18–31. [Google Scholar] [CrossRef]
- Timmerman, C.M.; Chapman, L.J. Behavioral and Physiological Compensation for Chronic Hypoxia in the Sailfin Molly (Poecilia latipinna). Physiol. Biochem. Zool. 2004, 77, 601–610. [Google Scholar] [CrossRef]
- Fu, S.-J.; Brauner, C.J.; Cao, Z.-D.; Richards, J.G.; Peng, J.-L.; Dhillon, R.; Wang, Y.-X. The effect of acclimation to hypoxia and sustained exercise on subsequent hypoxia tolerance and swimming performance in goldfish (Carassius auratus). J. Exp. Biol. 2011, 214, 2080–2088. [Google Scholar] [CrossRef]
- Remen, M.; Oppedal, F.; Imsland, A.K.; Olsen, R.; Torgersen, T. Hypoxia tolerance thresholds for post-smolt Atlantic salmon: Dependency of temperature and hypoxia acclimation. Aquaculture 2013, 416-417, 41–47. [Google Scholar] [CrossRef]
- Cook, D.G.; Iftikar, F.I.; Baker, D.W.; Hickey, A.J.R.; Herbert, N.A. Low O2 acclimation shifts the hypoxia avoidance behaviour of snapper (Pagrus auratus) with only subtle changes in aerobic and anaerobic function. J. Exp. Biol. 2013, 216, 369–378. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; So, B.E.; Farrell, A.P. Hypoxia Performance Curve: Assess a Whole-Organism Metabolic Shift from a Maximum Aerobic Capacity towards a Glycolytic Capacity in Fish. Metabolites 2021, 11, 447. https://doi.org/10.3390/metabo11070447
Zhang Y, So BE, Farrell AP. Hypoxia Performance Curve: Assess a Whole-Organism Metabolic Shift from a Maximum Aerobic Capacity towards a Glycolytic Capacity in Fish. Metabolites. 2021; 11(7):447. https://doi.org/10.3390/metabo11070447
Chicago/Turabian StyleZhang, Yangfan, Bog E. So, and Anthony P. Farrell. 2021. "Hypoxia Performance Curve: Assess a Whole-Organism Metabolic Shift from a Maximum Aerobic Capacity towards a Glycolytic Capacity in Fish" Metabolites 11, no. 7: 447. https://doi.org/10.3390/metabo11070447
APA StyleZhang, Y., So, B. E., & Farrell, A. P. (2021). Hypoxia Performance Curve: Assess a Whole-Organism Metabolic Shift from a Maximum Aerobic Capacity towards a Glycolytic Capacity in Fish. Metabolites, 11(7), 447. https://doi.org/10.3390/metabo11070447