
Metabolomic Characterization of Commercial, Old, and Red-Fleshed Apple Varieties

 , ,
, ,  and
and
Abstract
1. Introduction
2. Results and Discussion
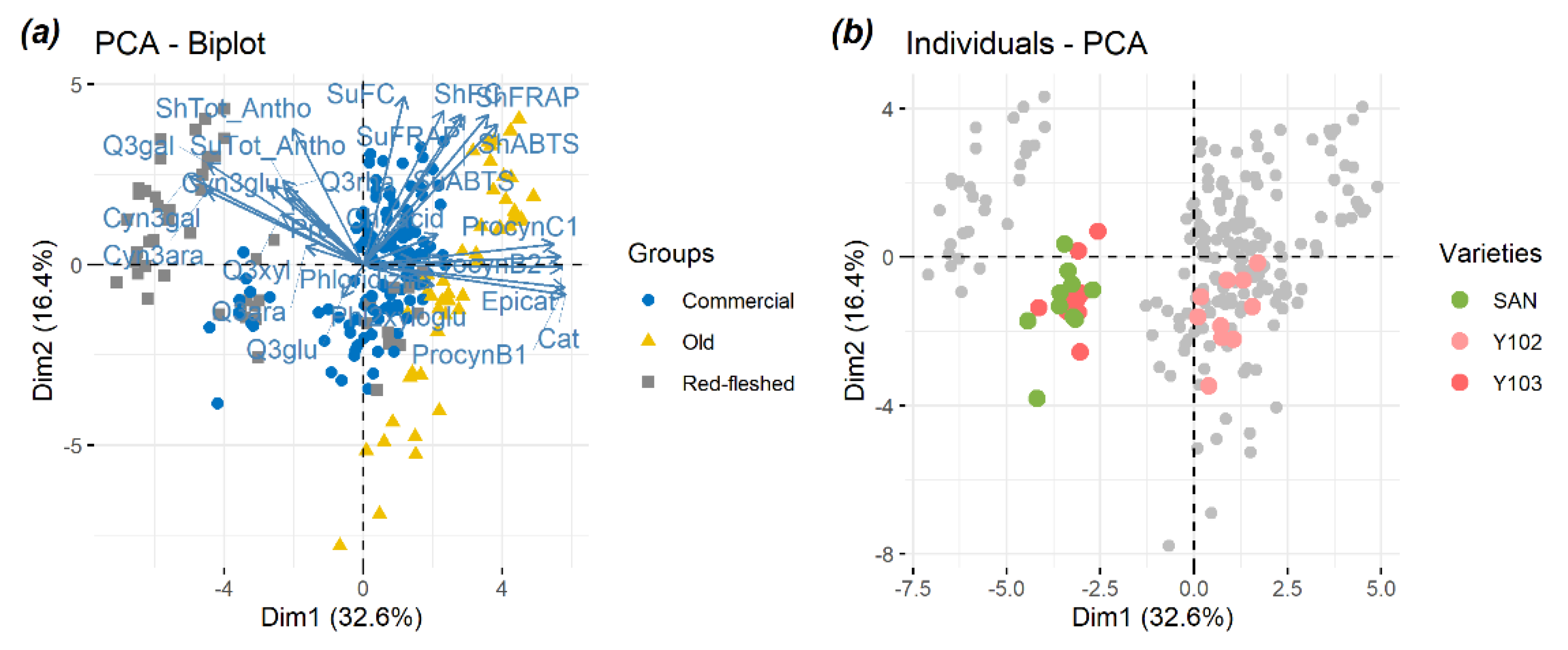
2.1. Principal Component Analysis (PCA) Used as an Exploratory Tool to Evaluate the Overall Apple Sample Distribution
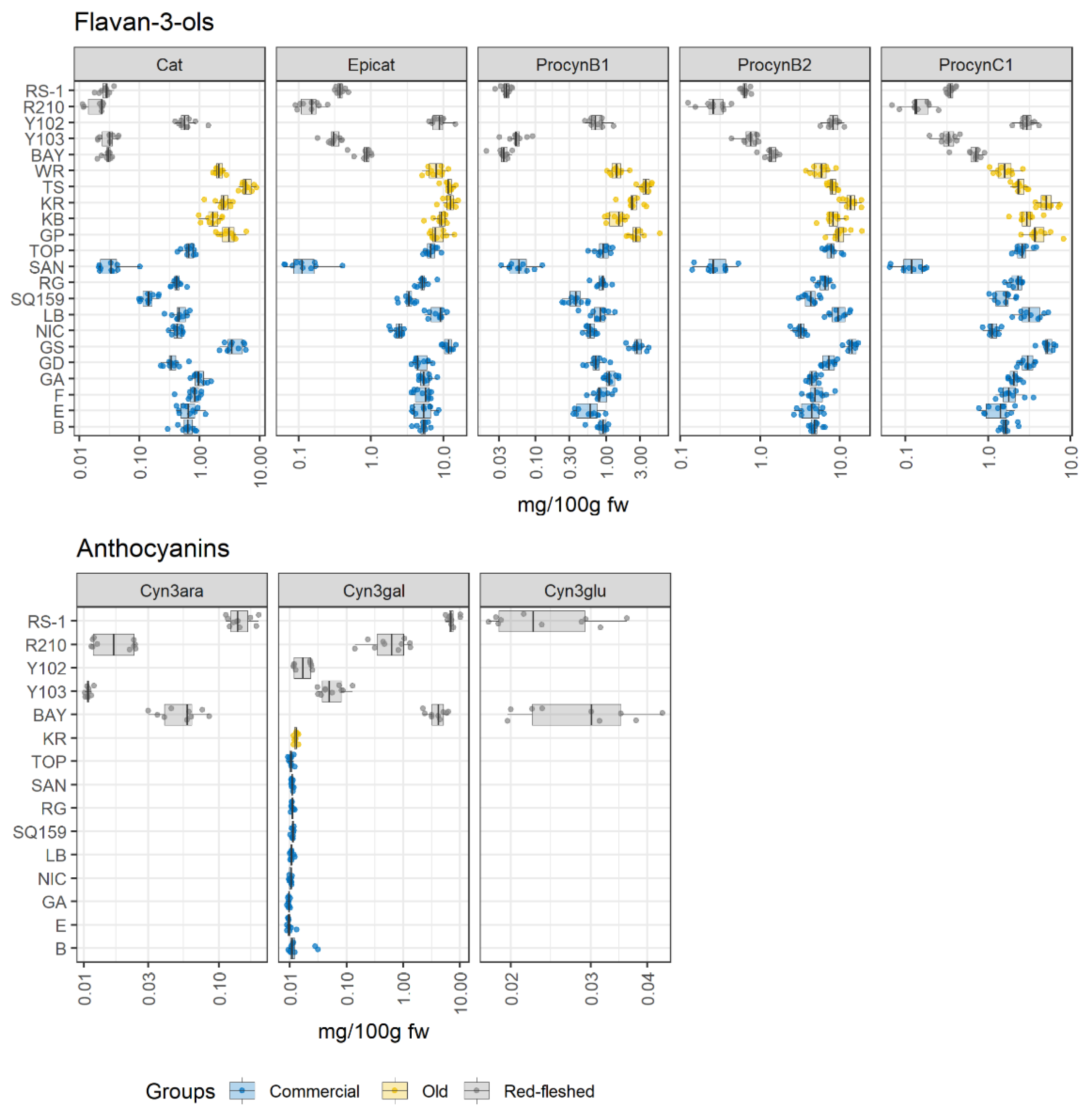
2.2. Apple Pulp: The Polyphenolic Profile
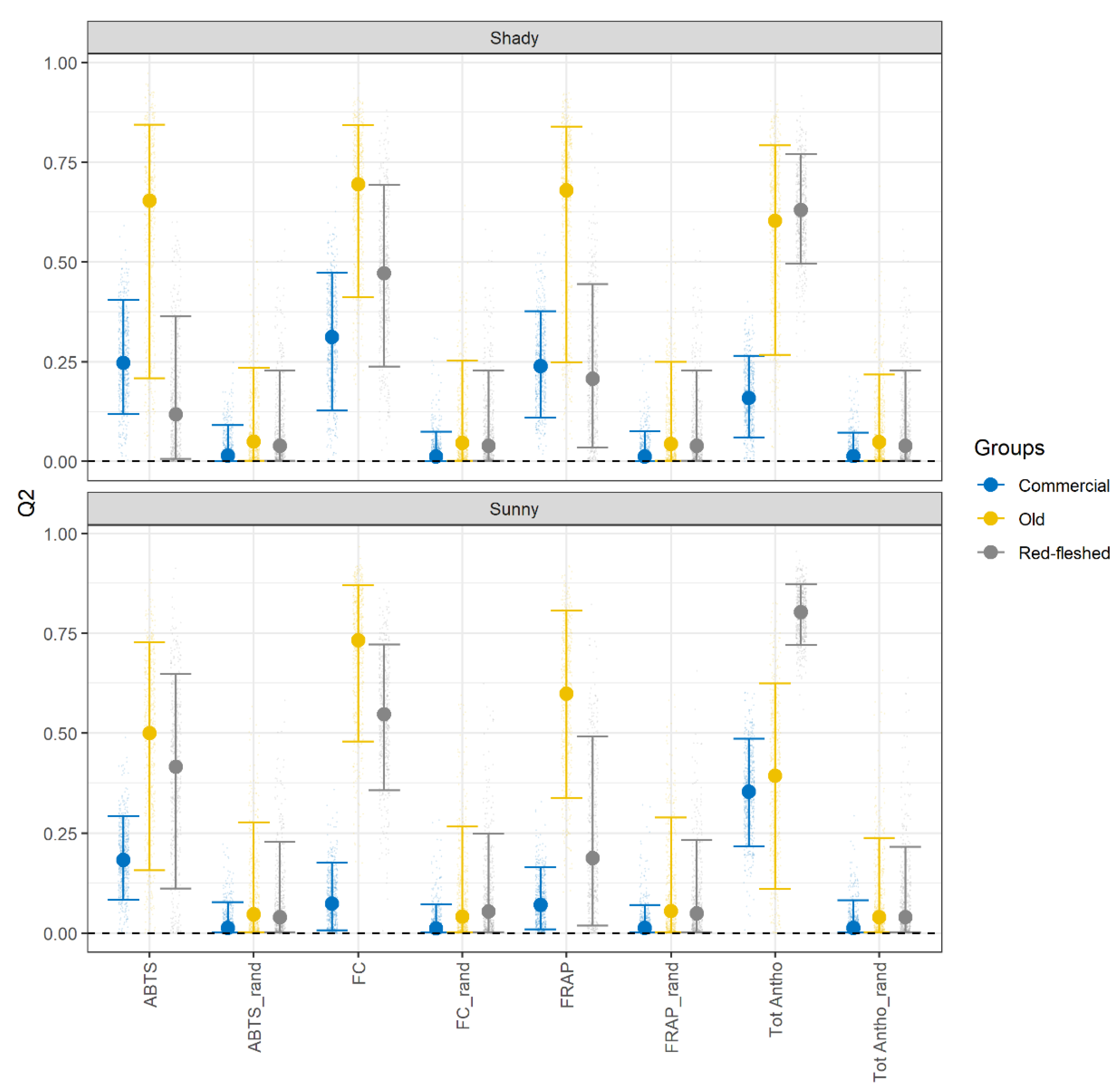
2.3. Apple Skin: The Antioxidant Activity (AA), Total Polyphenolic Content (TPC), and Total Anthocyanins Content (Tot Antho)
2.4. Prediction of the Antioxidant Activity in Apple Skin Based on the Polyphenolic Composition of Apple Pulp
3. Materials and Methods
3.1. Chemical and Reagents
3.2. Fruits Samples
3.3. Extraction of Samples
3.4. Analysis of Polyphenolic Profile of Apple Pulps
3.5. Spectrophotometric Assays of Apple Peels
3.5.1. Total Polyphenolic Content (TPC)
3.5.2. Total Anthocyanins Content (Tot Antho)
3.5.3. Antioxidant Capacity Measurements (AA)
Antioxidant Activity, ABTS Assay
Antioxidant Activity, FRAP Assay
3.6. Statistical Analysis
3.6.1. The Quantification Analysis
3.6.2. The Exploratory Tool: PCA
3.6.3. The PLS Regression Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Available online: https://www.who.int/health-topics/nutrition (accessed on 3 March 2021).
- Romagnolo, D.F.; Selmin, O.I. Mediterranean Diet and Prevention of Chronic Diseases. Nutr. Today 2017, 52, 208–222. [Google Scholar] [CrossRef]
- Saura-Calixto, F.; Goñi, I. Definition of the Mediterranean diet based on bioactive compounds. Crit. Rev. Food Sci. Nutr. 2009, 49, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Shoji, T.; Miura, T. Apple Polyphenols in Cancer Prevention. In Polyphenols in Human Health and Disease; Acad Press: London, UK, 2014; Volume 2, pp. 1373–1383. [Google Scholar]
- Liu, T.; Bassi, M.; Sadar, N.; Lubes, G.; Agnolet, S.; Stürz, B.; Guerra, W.; Robatscher, P.; Zanella, A.; Oberhuber, M. Non-destructive assessment of ascorbic acid in apples using near-infrared (NIR) spectroscopy together with partial least squares (PLS) regression. Acta Hortic. 2018, 447–454. [Google Scholar] [CrossRef]
- Denis, M.C.; Furtos, A.; Dudonné, S.; Montoudis, A.; Garofalo, C.; Desjardins, Y.; Delvin, E.; Levy, E. Apple peel polyphenols and their beneficial actions on oxidative stress and inflammation. PLoS ONE 2013, 8, e53725. [Google Scholar] [CrossRef]
- Persic, M.; Mikulic-Petkovsek, M.; Slatnar, A.; Veberic, R. Chemical composition of apple fruit, juice and pomace and the correlation between phenolic content, enzymatic activity and browning. LWT Food Sci. Technol. 2017, 82, 23–31. [Google Scholar] [CrossRef]
- Koutsos, A.; Riccadonna, S.; Ulaszewska, M.M.; Franceschi, P.; Trošt, K.; Galvin, A.; Braune, T.; Fava, F.; Perenzoni, D.; Mattivi, F.; et al. Two apples a day lower serum cholesterol and improve cardiometabolic biomarkers in mildly hypercholesterolemic adults: A randomized, controlled, crossover trial. Am. J. Clin. Nutr. 2020, 111, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Boyer, J.; Liu, R.H. Apple phytochemicals and their health benefits. Nutr. J. 2004, 3, 1–15. [Google Scholar] [CrossRef]
- Hyson, D.A. A Comprehensive Review of Apples and Apple Components and Their Relationship to Human Health. Adv. Nutr. 2011, 2, 408–420. [Google Scholar] [CrossRef]
- Tsao, R.; Yang, R.; Young, J.C.; Zhu, H. Polyphenolic profiles in eight apple cultivars using high-performance liquid chromatography (HPLC). J. Agric. Food Chem. 2003, 51, 6347–6353. [Google Scholar] [CrossRef]
- Vrhovsek, U.; Rigo, A.; Tonon, D.; Mattivi, F. Quantitation of polyphenols in different apple varieties. J. Agric. Food Chem. 2004, 52, 6532–6538. [Google Scholar] [CrossRef]
- Jacobo-Velázquez, D.A.; Cisneros-Zevallos, L. Correlations of antioxidant activity against phenolic content revisited: A new approach in data analysis for food and medicinal plants. J. Food Sci. 2009, 74, R107–R113. [Google Scholar] [CrossRef] [PubMed]
- Jokioja, J.; Percival, J.; Philo, M.; Yang, B.; Kroon, P.A.; Linderborg, K.M. Phenolic Metabolites in the Urine and Plasma of Healthy Men After Acute Intake of Purple Potato Extract Rich in Methoxysubstituted Monoacylated Anthocyanins. Mol. Nutr. Food Res. 2021, 65, 2000898. [Google Scholar] [CrossRef]
- Li, M.-J.; Ma, F.-W.; Zhang, M.; Pu, F. Distribution and metabolism of ascorbic acid in apple fruits (Malus domestica Borkh cv. Gala). Plant Sci. 2008, 174, 606–612. [Google Scholar] [CrossRef]
- Cuthbertson, D.; Andrews, P.K.; Reganold, J.P.; Davies, N.M.; Lange, B.M. Utility of metabolomics toward assessing the metabolic basis of quality traits in apple fruit with an emphasis on antioxidants. J. Agric. Food Chem. 2012, 60, 8552–8560. [Google Scholar] [CrossRef] [PubMed]
- Simmonds, M.S.; Howes, M.-J.R. Chapter 1-Profile of Compounds in Different Cultivars of Apple (Malus x domestica). In Nutritional Composition of Fruit Cultivars; Simmonds, M.S., Preedy, V.R., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 1–18. ISBN 978-0-12-408117-8. [Google Scholar]
- Baric, S.; Storti, A.; Hofer, M.; Guerra, W.; Dalla Via, J. Molecular Genetic Identification of Apple Cultivars Based on Microsatellite DNA Analysis. I. The Database of 600 Validated Profiles. Erwerbs Obstbau 2020, 62, 117–154. [Google Scholar] [CrossRef]
- Belviso, S.; Scursatone, B.; Re, G.; Zeppa, G. Novel Data on the Polyphenol Composition of Italian Ancient Apple Cultivars. Int. J. Food Prop. 2013, 16, 1507–1515. [Google Scholar] [CrossRef]
- Kissabel. Available online: https://kissabel.com/ (accessed on 11 May 2021).
- Farneti, B.; Masuero, D.; Costa, F.; Magnago, P.; Malnoy, M.; Costa, G.; Vrhovsek, U.; Mattivi, F. Is there room for improving the nutraceutical composition of apple? J. Agric. Food Chem. 2015, 63, 2750–2759. [Google Scholar] [CrossRef]
- Pojer, E.; Mattivi, F.; Johnson, D.; Stockley, C.S. The Case for Anthocyanin Consumption to Promote Human Health: A Review. Compr. Rev. Food Sci. Food Saf. 2013, 12, 483–508. [Google Scholar] [CrossRef]
- Bassi, M.; Lubes, G.; Bianchi, F.; Agnolet, S.; Ciesa, F.; Brunner, K.; Guerra, W.; Robatscher, P.; Oberhuber, M. Ascorbic acid content in apple pulp, peel, and monovarietal cloudy juices of 64 different cultivars. Int. J. Food Prop. 2017, 20, S2626–S2634. [Google Scholar] [CrossRef]
- Wojdyło, A.; Oszmiański, J.; Laskowski, P. Polyphenolic compounds and antioxidant activity of new and old apple varieties. J. Agric. Food Chem. 2008, 56, 6520–6530. [Google Scholar] [CrossRef] [PubMed]
- Jakobek, L.; Ištuk, J.; Buljeta, I.; Voća, S.; Žlabur, J.Š.; Babojelić, M.S. Traditional, Indigenous Apple Varieties, a Fruit with Potential for Beneficial Effects: Their Quality Traits and Bioactive Polyphenol Contents. Foods 2020, 9, 52. [Google Scholar] [CrossRef] [PubMed]
- Kschonsek, J.; Wolfram, T.; Stöckl, A.; Böhm, V. Polyphenolic Compounds Analysis of Old and New Apple Cultivars and Contribution of Polyphenolic Profile to the In Vitro Antioxidant Capacity. Antioxidants 2018, 7, 20. [Google Scholar] [CrossRef] [PubMed]
- Bars-Cortina, D.; Macià, A.; Iglesias, I.; Romero, M.P.; Motilva, M.J. Phytochemical Profiles of New Red-Fleshed Apple Varieties Compared with Traditional and New White-Fleshed Varieties. J. Agric. Food Chem. 2017, 65, 1684–1696. [Google Scholar] [CrossRef]
- Henry-Kirk, R.A.; McGhie, T.K.; Andre, C.M.; Hellens, R.P.; Allan, A.C. Transcriptional analysis of apple fruit proanthocyanidin biosynthesis. J. Exp. Bot. 2012, 63, 5437–5450. [Google Scholar] [CrossRef]
- Lancaster, J.E.; Dougall, D.K. Regulation of skin color in apples. Crit. Rev. Plant Sci. 1992, 10, 487–502. [Google Scholar] [CrossRef]
- Chagné, D.; Lin-Wang, K.; Espley, R.V.; Volz, R.K.; How, N.M.; Rouse, S.; Brendolise, C.; Carlisle, C.M.; Kumar, S.; De Silva, N.; et al. An ancient duplication of apple MYB transcription factors is responsible for novel red fruit-flesh phenotypes. Plant Physiol. 2013, 161, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Espley, R.V.; Bovy, A.; Bava, C.; Jaeger, S.R.; Tomes, S.; Norling, C.; Crawford, J.; Rowan, D.; McGhie, T.K.; Brendolise, C.; et al. Analysis of genetically modified red-fleshed apples reveals effects on growth and consumer attributes. Plant Biotechnol. J. 2013, 11, 408–419. [Google Scholar] [CrossRef] [PubMed]
- Giomaro, G.; Karioti, A.; Bilia, A.R.; Bucchini, A.; Giamperi, L.; Ricci, D.; Fraternale, D. Polyphenols profile and antioxidant activity of skin and pulp of a rare apple from Marche region (Italy). Chem. Cent. J. 2014, 8, 1–10. [Google Scholar] [CrossRef]
- McClure, K.A.; Gong, Y.; Song, J.; Vinqvist-Tymchuk, M.; Palmer, L.C.; Fan, L.; Burgher-MacLellan, K.; Zhang, Z.; Celton, J.-M.; Forney, C.F.; et al. Genome-wide association studies in apple reveal loci of large effect controlling apple polyphenols. Hortic. Res. 2019, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Vanzo, A.; Jenko, M.; Vrhovsek, U.; Stopar, M. Metabolomic profiling and sensorial quality of ‘Golden Delicious’, ‘Liberty’, ‘Santana’, and ‘Topaz’ apples grown using organic and integrated production systems. J. Agric. Food Chem. 2013, 61, 6580–6587. [Google Scholar] [CrossRef]
- Simonato, B.; Marangon, M.; Vincenzi, S.; Vegro, M.; Pasini, G. Evaluation of the phenolic profile and immunoreactivity of Mal d 3 allergen in ancient apple cultivars from Italy. J. Sci. Food Agric. 2020, 100, 4978–4986. [Google Scholar] [CrossRef]
- Kootstra, H.S.; Vlieg-Boerstra, B.J.; Dubois, A.E.J. Assessment of the reduced allergenic properties of the Santana apple. Ann. Allergy Asthma Immunol. 2007, 99, 522–525. [Google Scholar] [CrossRef]
- Li, P.; Ma, F.; Cheng, L. Primary and secondary metabolism in the sun-exposed peel and the shaded peel of apple fruit. Physiol. Plant. 2013, 148, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Biosüedtirol. Available online: https://biosuedtirol.com/en/apple/nicoter-kanzi (accessed on 15 May 2021).
- Pomiferous. Available online: https://pomiferous.com/applebyname/rosa-di-caldaro-id-5552 (accessed on 11 May 2021).
- Zhang, X.; Xu, J.; Xu, Z.; Sun, X.; Zhu, J.; Zhang, Y. Analysis of Antioxidant Activity and Flavonoids Metabolites in Peel and Flesh of Red-Fleshed Apple Varieties. Molecules 2020, 25, 1968. [Google Scholar] [CrossRef] [PubMed]
- Kalinowska, M.; Bielawska, A.; Lewandowska-Siwkiewicz, H.; Priebe, W.; Lewandowski, W. Apples: Content of phenolic compounds vs. variety, part of apple and cultivation model, extraction of phenolic compounds, biological properties. Plant Physiol. Biochem. 2014, 84, 169–188. [Google Scholar] [CrossRef]
- Lim, V.; Gorji, S.G.; Daygon, V.D.; Fitzgerald, M. Untargeted and Targeted Metabolomic Profiling of Australian Indigenous Fruits. Metabolites 2020, 10, 114. [Google Scholar] [CrossRef]
- Sahin, S.; Demir, C. Determination of Antioxidant Properties of Fruit Juice by Partial Least Squares and Principal Component Regression. Int. J. Food Prop. 2016, 19, 1455–1464. [Google Scholar] [CrossRef]
- Zheng, H.; Jiang, L.; Lou, H.; Hu, Y.; Kong, X.; Lu, H. Application of artificial neural network (ANN) and partial least-squares regression (PLSR) to predict the changes of anthocyanins, ascorbic acid, Total phenols, flavonoids, and antioxidant activity during storage of red bayberry juice based on fractal analysis and red, green, and blue (RGB) intensity values. J. Agric. Food Chem. 2011, 59, 592–600. [Google Scholar] [CrossRef]
- Vanzani, P.; Rossetto, M.; Rigo, A.; Vrhovsek, U.; Mattivi, F.; D’Amato, E.; Scarpa, M. Major phytochemicals in apple cultivars: Contribution to peroxyl radical trapping efficiency. J. Agric. Food Chem. 2005, 53, 3377–3382. [Google Scholar] [CrossRef]
- Csepregi, K.; Neugart, S.; Schreiner, M.; Hideg, É. Comparative Evaluation of Total Antioxidant Capacities of Plant Polyphenols. Molecules 2016, 21, 208. [Google Scholar] [CrossRef]
- Tsao, R.; Yang, R.; Xie, S.; Sockovie, E.; Khanizadeh, S. Which polyphenolic compounds contribute to the total antioxidant activities of apple? J. Agric. Food Chem. 2005, 53, 4989–4995. [Google Scholar] [CrossRef]
- Rossetto, M.; Vanzani, P.; Mattivi, F.; Lunelli, M.; Scarpa, M.; Rigo, A. Synergistic antioxidant effect of catechin and malvidin 3-glucoside on free radical-initiated peroxidation of linoleic acid in micelles. Arch. Biochem. Biophys. 2002, 408, 239–245. [Google Scholar] [CrossRef]
- Solovchenko, A.; Schmitz-Eiberger, M. Significance of skin flavonoids for UV-B-protection in apple fruits. J. Exp. Bot. 2003, 54, 1977–1984. [Google Scholar] [CrossRef] [PubMed]
- Valls, J.; Agnolet, S.; Haas, F.; Struffi, I.; Ciesa, F.; Robatscher, P.; Oberhuber, M. Valorization of Lagrein grape pomace as a source of phenolic compounds: Analysis of the contents of anthocyanins, flavanols and antioxidant activity. Eur. Food Res. Technol. 2017, 243, 2211–2224. [Google Scholar] [CrossRef]
- Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. R Package Version 2010, 1, 1–8. [Google Scholar]
- Wei, R.; Wang, J.; Su, M.; Jia, E.; Chen, S.; Chen, T.; Ni, Y. Missing Value Imputation Approach for Mass Spectrometry-based Metabolomics Data. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Granato, D.; Santos, J.S.; Escher, G.B.; Ferreira, B.L.; Maggio, R.M. Use of principal component analysis (PCA) and hierarchical cluster analysis (HCA) for multivariate association between bioactive compounds and functional properties in foods: A critical perspective. Trends Food Sci. Technol. 2018, 72, 83–90. [Google Scholar] [CrossRef]
- Feng, C.; Wang, H.; Lu, N.; Chen, T.; He, H.; Lu, Y.; Tu, X.M. Log-transformation and its implications for data analysis. Shanghai Arch. Psychiatry 2014, 26, 105–109. [Google Scholar] [CrossRef]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Extract and Visualize the Results of Multivariate Data Analyses factoextra (datanovia.com). Available online: https://rpkgs.datanovia.com/factoextra/index.html (accessed on 9 June 2021).
- Aznar, M.; López, R.; Cacho, J.; Ferreira, V. Prediction of aged red wine aroma properties from aroma chemical composition. Partial least squares regression models. J. Agric. Food Chem. 2003, 51, 2700–2707. [Google Scholar] [CrossRef]
- Cozzolino, D.; Kwiatkowski, M.J.; Dambergs, R.G.; Cynkar, W.U.; Janik, L.J.; Skouroumounis, G.; Gishen, M. Analysis of elements in wine using near infrared spectroscopy and partial least squares regression. Talanta 2008, 74, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Hauksson, J.; Bergqvist, G.; Bergsten, U.; Sjostrom, M.; Edlund, U. Prediction of basic wood properties for Norway spruce. Interpretation of Near Infrared Spectroscopy data using partial least squares regression. Wood Sci. Technol. 2001, 35, 475–485. [Google Scholar] [CrossRef]
- Kuhn, M. Building Predictive Models in R Using the caret Package. J. Stat. Softw. 2008, 28, 1–26. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Apple Cultivars | Code | Groups | Harvest Date | +10 d CS | 10 d CS + 3 d SL |
|---|---|---|---|---|---|
| Goldparmäne | GP | Old | 20 August 2015 | 30 August 2015 | 2 September 2015 |
| Kalterer Böhmer | KB | Old | 15 September 2015 | 25 September 2015 | 28 September 2015 |
| Kanada Renette | KR | Old | 15 September 2015 | 25 September 2015 | 28 September 2015 |
| Tiroler | TS | Old | 3 November 2015 | 13 November 2015 | 16 November 2015 |
| Spitzlederer | |||||
| Weisser | WR | Old | 28 September 2015 | 8 October 2015 | 11 October 2015 |
| Rosmarin | |||||
| Braeburn | B | Commercial | 2 October 2015 | 12 October 2015 | 15 October 2015 |
| Fuji (Brak) | F | Commercial | 6 October 2015 | 16 October 2015 | 19 October 2015 |
| Elstar | E | Commercial | 20 August 2015 | 30 August 2015 | 2 September 2015 |
| Gala (Simmons) | GA | Commercial | 12 August 2015 | 22 August 2015 | 25 August 2015 |
| Golden | GD | Commercial | 14 September 2015 | 24 September 2015 | 27 September 2015 |
| Delicious | |||||
| Granny | GS | Commercial | 24 September 2015 | 4 October 2015 | 7 October 2015 |
| Smith | |||||
| Nicoter | NIC | Commercial | 16 September 2015 | 26 September 2015 | 29 September 2015 |
| Lb 17906 | LB | Commercial | 27 October 2015 | 6 November 2015 | 9 November 2015 |
| SQ159 | SQ159 | Commercial | 17 September 2015 | 27 September 2015 | 30 September 2015 |
| Rosy Glow | RG | Commercial | 27 October 2015 | 6 November 2015 | 9 November 2015 |
| Santana | SAN | Commercial | 12 August 2015 | 22 August 2015 | 25 August 2015 |
| Topaz | TOP | Commercial | 21 September 2015 | 1 October 2015 | 4 October 2015 |
| Bay 3484 | BAY | Red-fleshed (type 1) | 13 August 2015 | 23 August 2015 | 26 August 2015 |
| Red-fleshed 2/Y103 | Y103 | Red-fleshed (type 2) | 13 August 2015 | 23 August 2015 | 26 August 2015 |
| Red-fleshed 3/Y102 | Y102 | Red-fleshed (type 2) | 1 September 2015 | 11 September 2015 | 14 September 2015 |
| Red-fleshed 4/R201 | R210 | Red-fleshed (type 1) | 30 September 2015 | 10 October 2015 | 13 October 2015 |
| RS-1 | RS-1 | Red-fleshed (type 1) | 20 August 2015 | 30 August 2015 | 2 September 2015 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ceci, A.T.; Bassi, M.; Guerra, W.; Oberhuber, M.; Robatscher, P.; Mattivi, F.; Franceschi, P. Metabolomic Characterization of Commercial, Old, and Red-Fleshed Apple Varieties. Metabolites 2021, 11, 378. https://doi.org/10.3390/metabo11060378
Ceci AT, Bassi M, Guerra W, Oberhuber M, Robatscher P, Mattivi F, Franceschi P. Metabolomic Characterization of Commercial, Old, and Red-Fleshed Apple Varieties. Metabolites. 2021; 11(6):378. https://doi.org/10.3390/metabo11060378
Chicago/Turabian StyleCeci, Adriana Teresa, Michele Bassi, Walter Guerra, Michael Oberhuber, Peter Robatscher, Fulvio Mattivi, and Pietro Franceschi. 2021. "Metabolomic Characterization of Commercial, Old, and Red-Fleshed Apple Varieties" Metabolites 11, no. 6: 378. https://doi.org/10.3390/metabo11060378
APA StyleCeci, A. T., Bassi, M., Guerra, W., Oberhuber, M., Robatscher, P., Mattivi, F., & Franceschi, P. (2021). Metabolomic Characterization of Commercial, Old, and Red-Fleshed Apple Varieties. Metabolites, 11(6), 378. https://doi.org/10.3390/metabo11060378