1. Introduction
The metabolic diseases plaguing today’s civilization result from damage to normal metabolisms, especially in cases such as diabetes mellitus, in which the carbohydrate pathway is disrupted by inappropriate insulin secretion or abnormal responses by the body. With this disruption of the carbohydrate pathway, there is an elevated level of glucose in the blood, called hyperglycemia. The glucose consumption of most cells is regulated by insulin. Red blood cells (RBCs) do not need insulin; however, in the absence of this hormone, they are exposed to high glucose levels during hyperglycemia. Glucose has an effect on RBCs, changing their biochemical properties and the ability of these cells to aggregate. Changes in these processes depend on the period of time the cells are exposed to glucose and on the glucose concentration, but also on the composition of the solution in which the RBCs are suspended. Polymers are widely known among the substances that change the aggregation of RBCs. Thus, it is particularly important to understand the simultaneous effects of glucose and drug polymers on RBC aggregation.
Biochemical tests, which are the basis for the diagnosis of diabetes, are increasingly supplemented with analyses of the rheological properties of blood [
1,
2]. Many factors influence the rheology of whole blood, including RBC aggregation. One of the first documented studies on the changes in RBC aggregation caused by elevated glucose levels was conducted in 1956 [
3]. The in vivo effect of increased RBC aggregation under hyperglycemia was confirmed in the following years using various research methods and under various experimental conditions [
4,
5,
6,
7,
8]. It has been shown that increased aggregation in patients with elevated glucose levels in type 2 DM decreases after the application of glycemic control, and this effect is visible both for RBCs suspended in autologous plasma and cells suspended in dextran solutions [
8]. In general, the incubation of RBCs in glucose solutions in vitro causes a decrease in the aggregation of the cells. In order to demonstrate the effect in vitro, cells of healthy volunteers incubated in high glucose concentrations were studied [
9,
10,
11,
12]. RBC aggregation, as measured by the Aggregate Shape Parameter (ASP) method after two hours of incubation [
10] and by the Myrenne method after half an hour of incubation [
11], showed a downward trend.
In the case of blood, RBCs are suspended in the plasma, which may contain medicinal substances. One of these substances may be the nonionic surfactant poloxamer 188 (P188), which has many uses as a pharmaceutical excipient [
13,
14,
15]. Extensive research is ongoing into its potential use as a biological membrane sealant [
16,
17,
18], while other research is focused on its hemorheological effects [
19,
20,
21,
22]. In vitro and in vivo studies of the influence of P188 on hemorheological parameters in patients with acute myocardial infarction showed that the aggregation of RBCs in autologous plasma decreased after the application of this surfactant [
19]. Conversely, the aggregation of RBCs of healthy volunteers measured in dextran 70 solution after in vitro treatment with P188 decreased, and the effect was dependent on the concentration of P188 [
21]. The in vitro activity of P188 tested in dextran 70 solution was confirmed by studies of cells from healthy donors and donors with sickle cell anemia [
18]. Another study showed a reduction in RBC deformability by 10% only after 2 h of incubation in P188 [
22]. The determination of the influence of P188 on the aggregation of RBCs from healthy donors in dextran 70 solution, used as a medium for cells, allows for the exclusion of the effects of interindividual differences in plasma composition.
Knowledge of the possible joint effect of P188 and glucose is important for both the development of dosage forms and the prediction of enhancing effects in therapeutic applications that are still under development. There are studies investigating the synergistic effect of P188 and tinidazole on the solubility and drug release profile [
23] or on the treatment of certain diseases [
24], but the effect on RBC aggregation is a new area of research. Knowledge about the possible additional enhancement of the anti-aggregation effect of P188 is important, especially in light of reports about the need to administer P188 as soon as possible in order to support endogenous cell repair systems [
25]. In this work, the effect of P188 on RBC aggregation in the presence of glucose was investigated. We present the results of a new approach where RBC aggregation was tested during the incubation of cells in dextran solutions with glucose and with or without P188. The aggregation of RBCs is usually measured immediately after sample preparation. In this article, we use a working hypothesis that the aggregation of RBCs changes during the incubation of these cells in solutions containing glucose and P188. Thus, we propose the measurement of temporal changes in RBC aggregation. This working hypothesis may be useful as the basis for further research into the effects of various substances on erythrocyte aggregation.
3. Discussion
Metabolic processes are time dependent, and their courses are modified by the presence of drugs and other substances in the body. Both of these elements are taken into account in this article. The subject of this study was the time dependence of RBC aggregation parameters in solutions containing glucose and the influence of P188 added to these solutions on the aggregation parameters. Thus, in addition to the temporal effect, the joint effect of glucose and P188 on RBC aggregation was investigated.
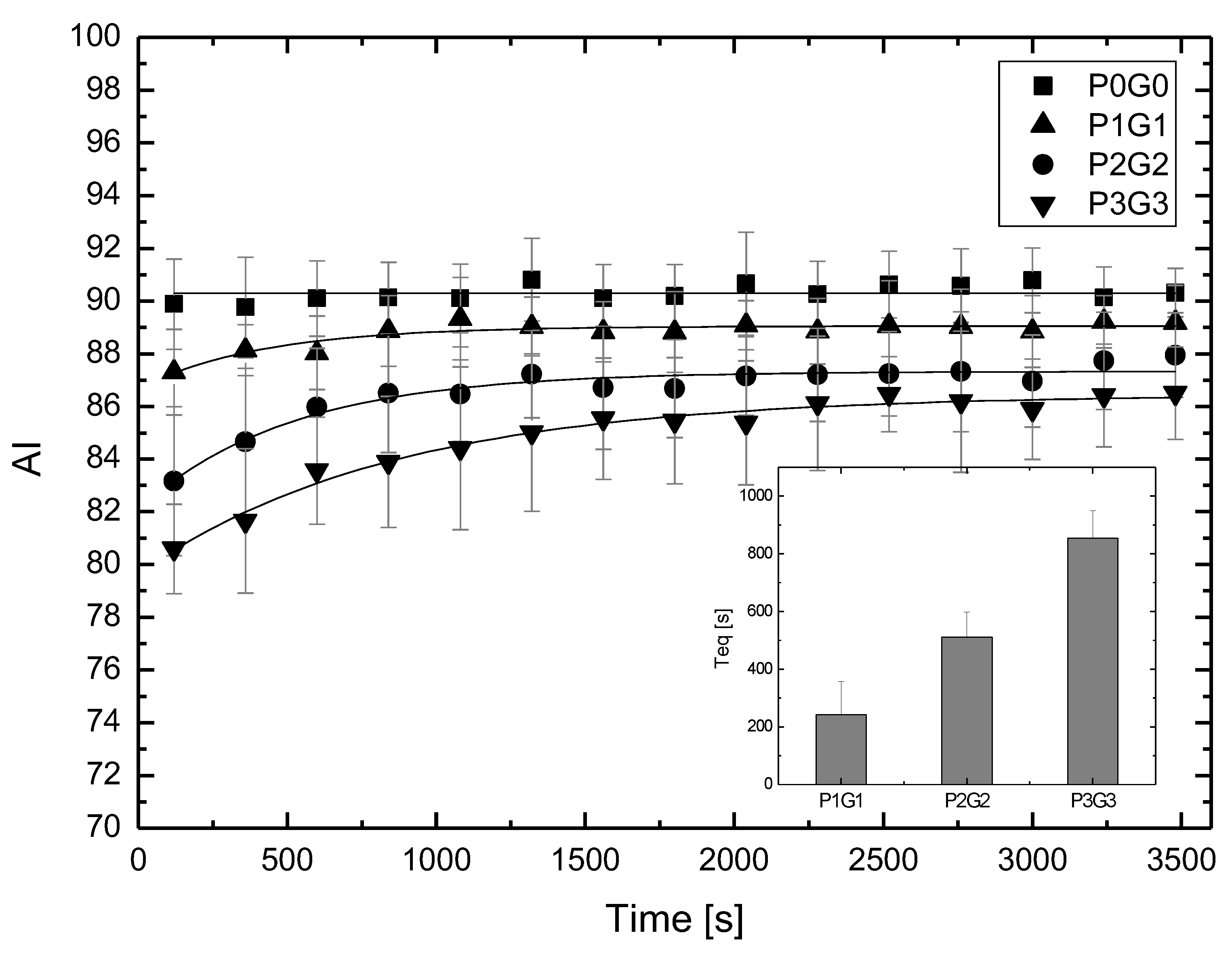
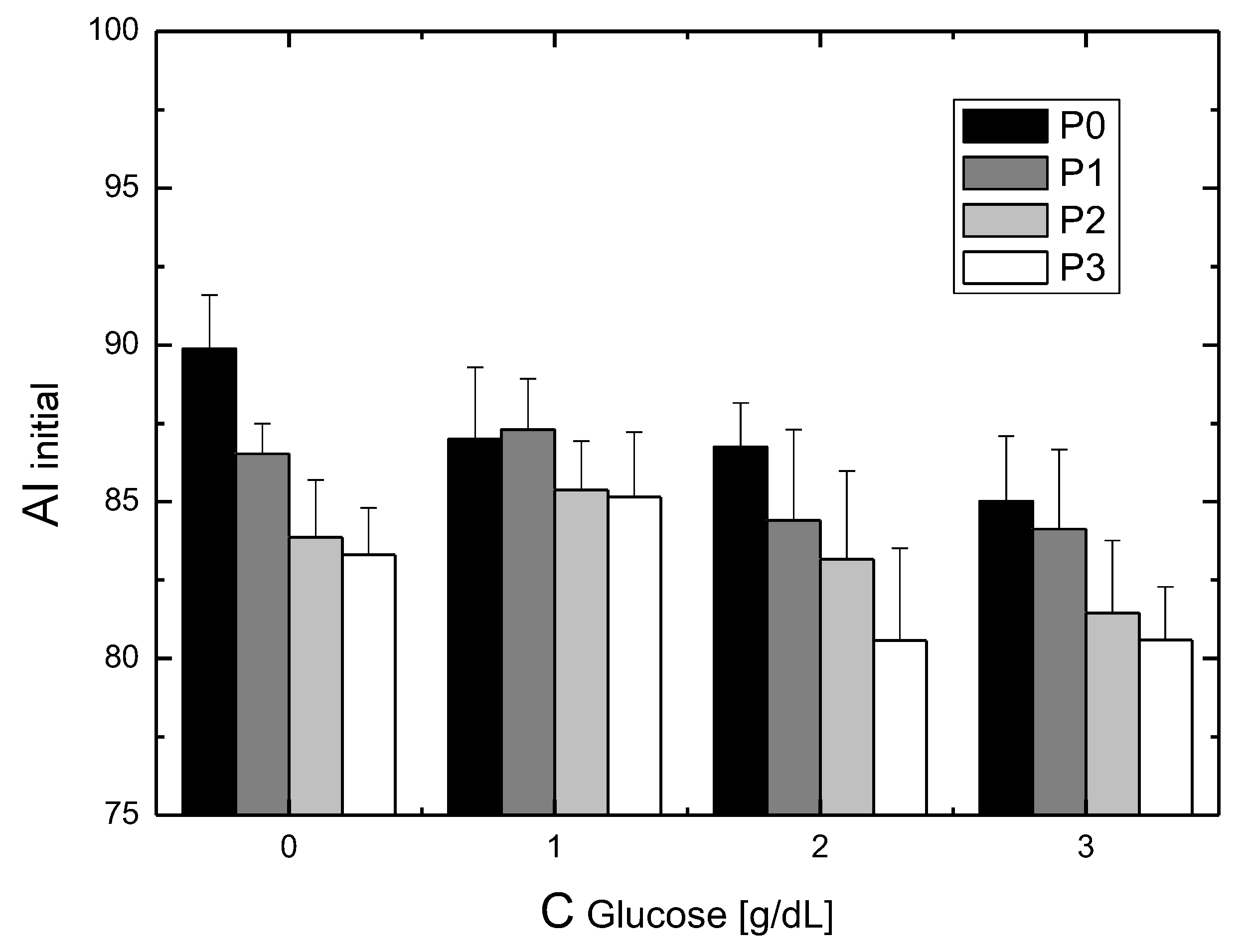
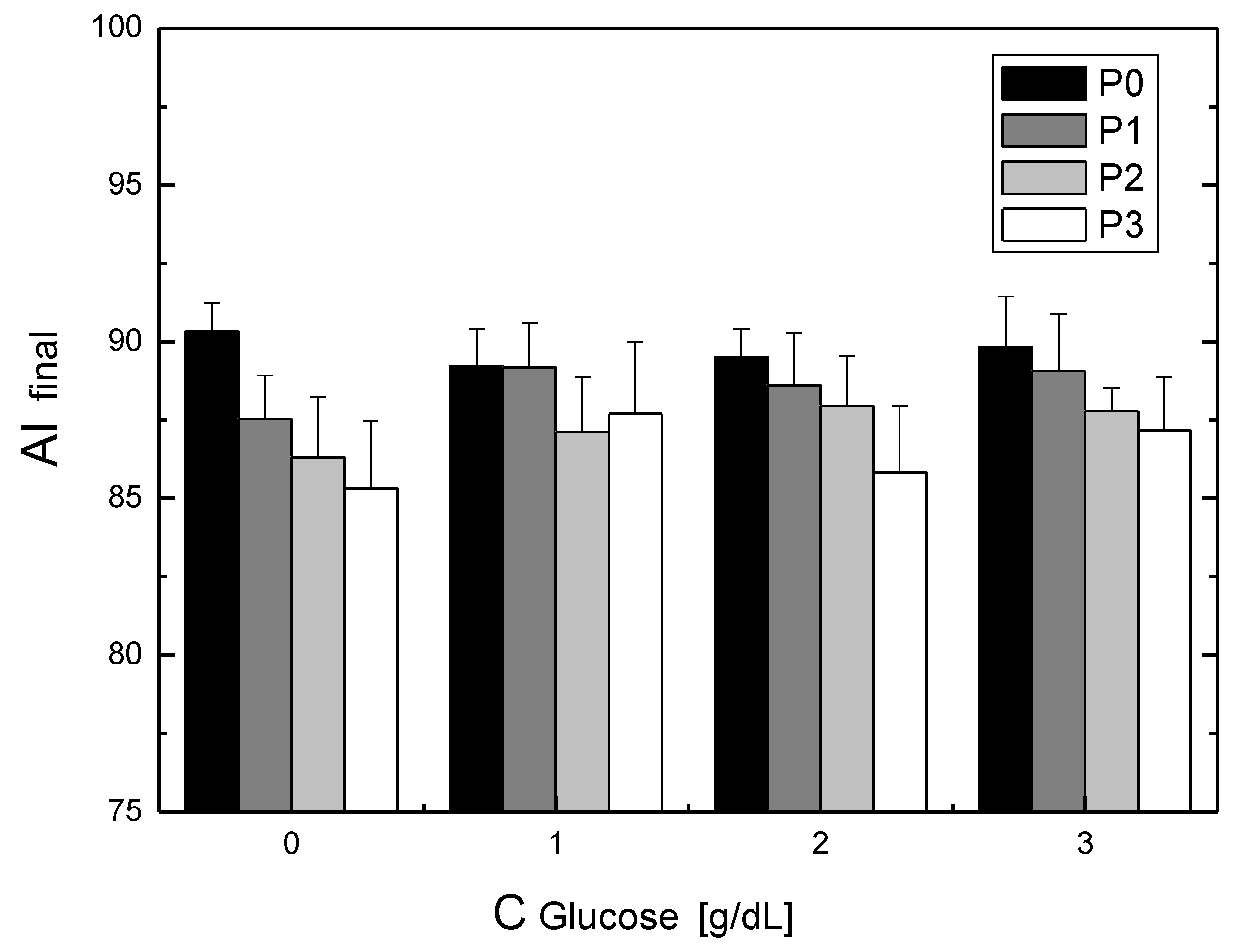
Studying the influence of glucose on the rheological properties of blood has a long history and results from the need to understand the mechanisms involved in diabetes [
26]. Research shows that incubation of RBCs in glucose solutions causes oxidation of membrane lipids and glycation of proteins, which leads to a reduction of RBC aggregation [
9,
12]. Shin et al. suggest that these changes occur even after 1 h of incubation. We conduct research depending on time. Thus, oxidation and glycation affect changes in RBC aggregation; however, the results presented in this paper suggest that glucose transport to erythrocytes plays a key role in the temporal changes in the aggregation of these cells. Elevated blood glucose also affects red-blood-cell aggregation, but the in vivo and in vitro effects are not the same and are still not fully understood; thus, this issue still needs to be investigated [
3,
4,
5,
6,
7,
8,
9,
10,
11]. The results presented in this study show that, for RBCs placed in glucose solutions, the aggregation index increases with the time of incubation in these solutions. After the growth period, the saturation of this parameter is observed. The time required for saturation increases as the glucose concentration in the solution increases. The aggregation index values measured immediately after placing the RBCs in solutions containing glucose decrease with the concentration of glucose. However, the aggregation index measured at the time of saturation does not show such a marked decrease as a function of the glucose concentration. This means that the time of the incubation of the RBCs in the glucose solution has a significant influence on the course of aggregation and may lead to differing results [
9,
10,
11,
12].
The phenomenon of transport occurs for RBCs placed in glucose solutions. Glucose enters the RBCs, and this transport does not require the presence of insulin [
27]. As a result, the concentration of glucose in the solution decreases while the concentration of glucose inside the RBCs increases. Lowering the glucose concentration in the solution in which the RBCs are suspended results in a lower viscosity of the solution [
28,
29]. This happens until equilibrium is achieved—when the transportation back and forth is the same. The study of this transport showed that the values of the parameters of this transport significantly differ depending on whether the transport takes place inside the RBC or in the solution [
12]. This immediately indicates that the issue of the incubation time and cell cleaning after incubation can cause major changes in aggregation parameters. Finally, a question arises about the mechanisms of the change in aggregation during incubation in glucose solutions. Research shows that the biochemical changes of RBCs incubated in glucose solutions occur slowly. Thus, it can be assumed that, for the results shown here, the biochemical changes had little effect on the changes in aggregation. This allows for a hypothesis that, in the cases considered here, mechanical changes in the cell membrane and changes in the viscosity of the substance in which the RBCs are dissolved determine the changes in the aggregation of these cells.
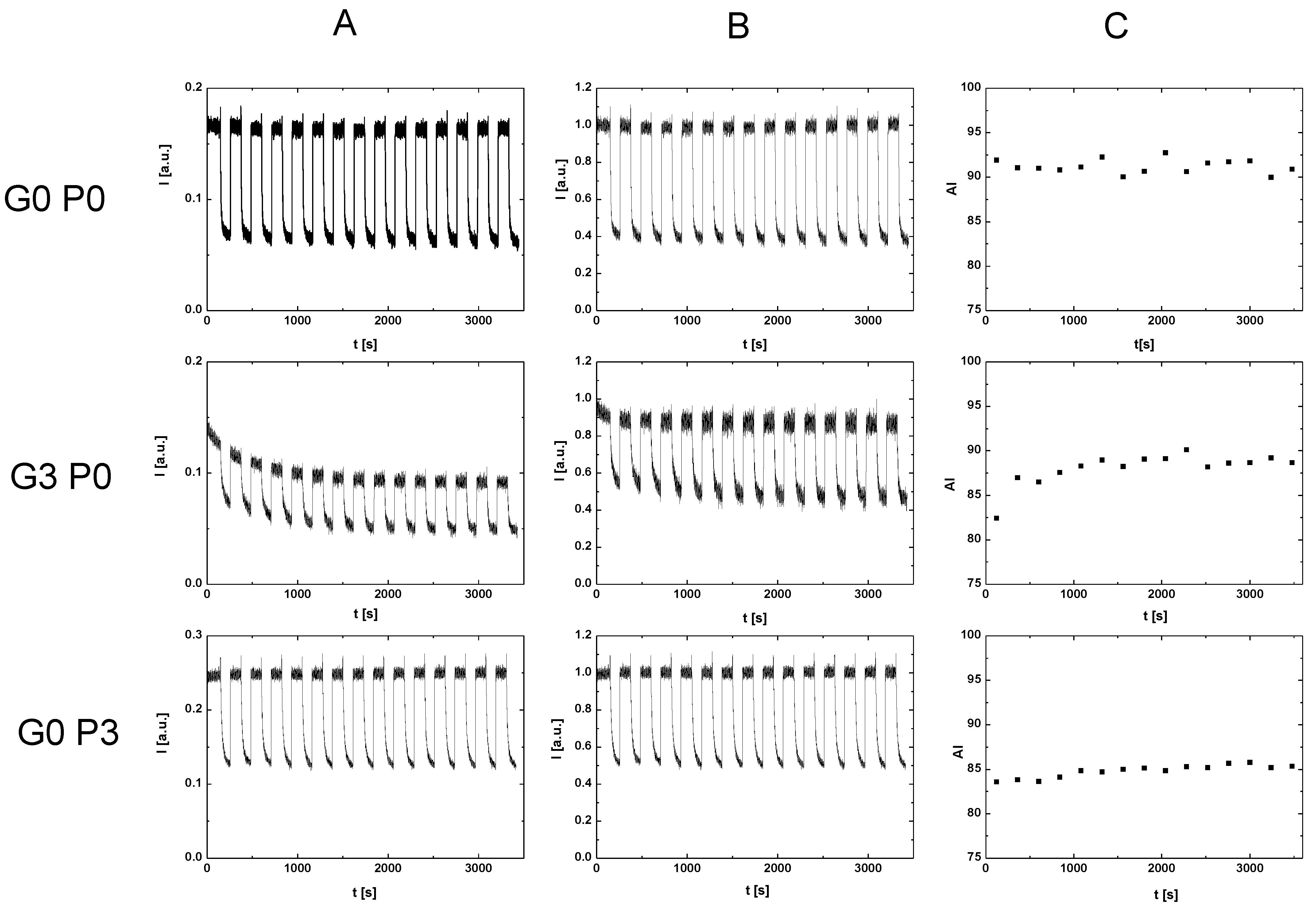
For RBCs placed in P188 solutions, as in the case of RBCs placed in solutions containing glucose, we observed an increase in the aggregation index with the incubation time. The saturation effect was also observed. In this case, both the initial and final aggregation index values decreased with increasing P188 concentrations in the solution. This RBC aggregation behavior corresponding to the initial aggregation index values studied here was observed using the microscopic aggregation index (MAI) and Myrenne method [
21]. As in the previous case, a question arises about the mechanism of the aggregation process in the case of RBCs in solutions containing P188. The reduction in the aggregation index appears to be due to the presence of a low-molecular-weight polymer in the solution. According to the depletion theory of aggregation, such polymers reduce the aggregation capacity of RBCs [
30]. The question remains open, however, of why the presence of polaxamer causes temporary changes in the RBC aggregation index similar to the temporal changes in the aggregation index of RBCs placed in glucose solutions.
Consider the problem of the synergistic effect of glucose and P188 on RBC aggregation. According to the definition, a synergistic effect takes place when the effect of processes interacting together is greater than the effects of the individual processes. The results presented in this study show that the presence of P188 enhances the process of suppressing glucose-dependent RBC aggregation. We showed this effect, expected by clinicians, only in vitro. The mechanism of lowering aggregation in this case results from the aggregation mechanisms for glucose solutions and the aggregation mechanism for P188 solutions. It is not certain, however, whether the mechanism of the synergistic effect is a simple assembly of partial mechanisms.
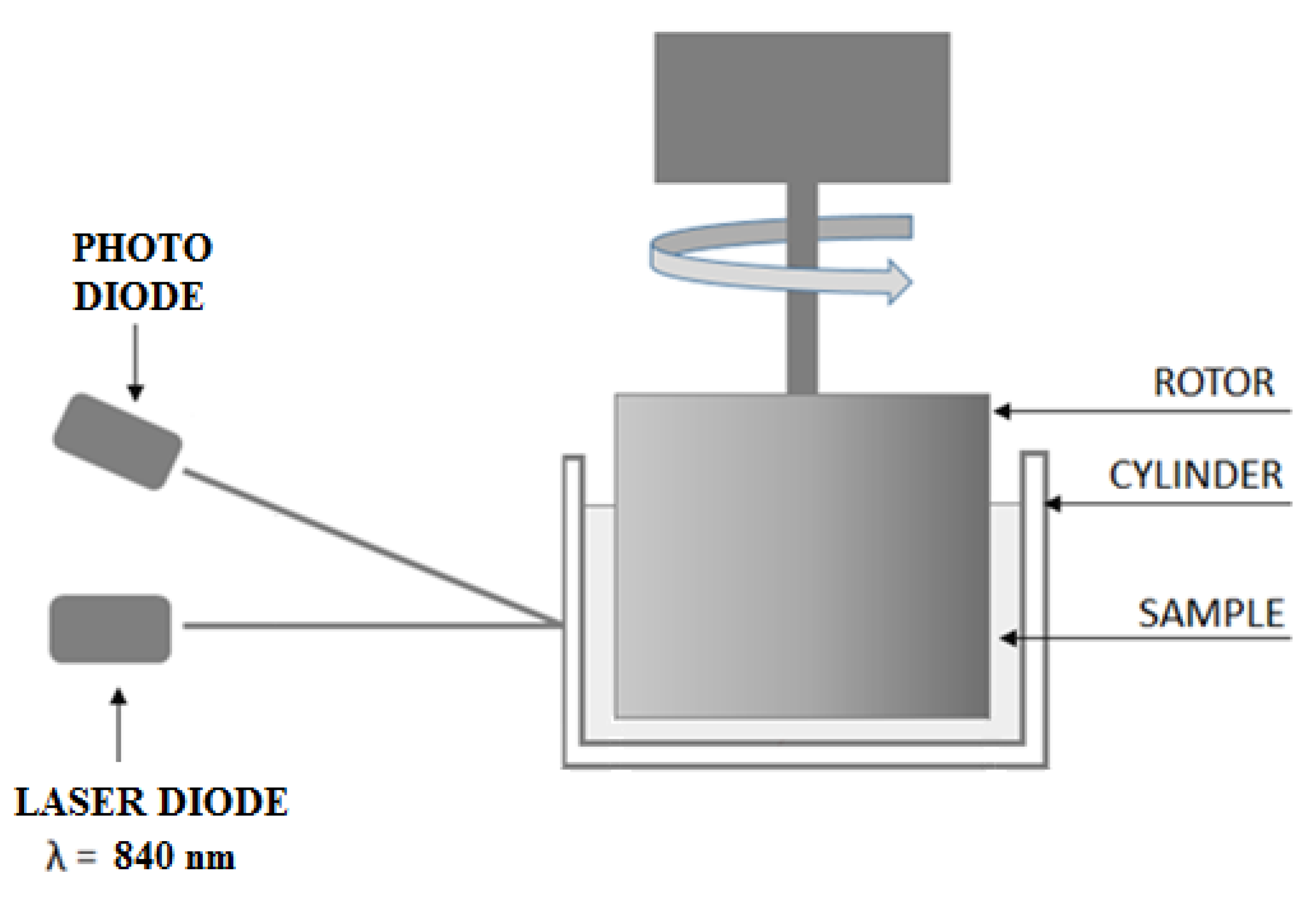
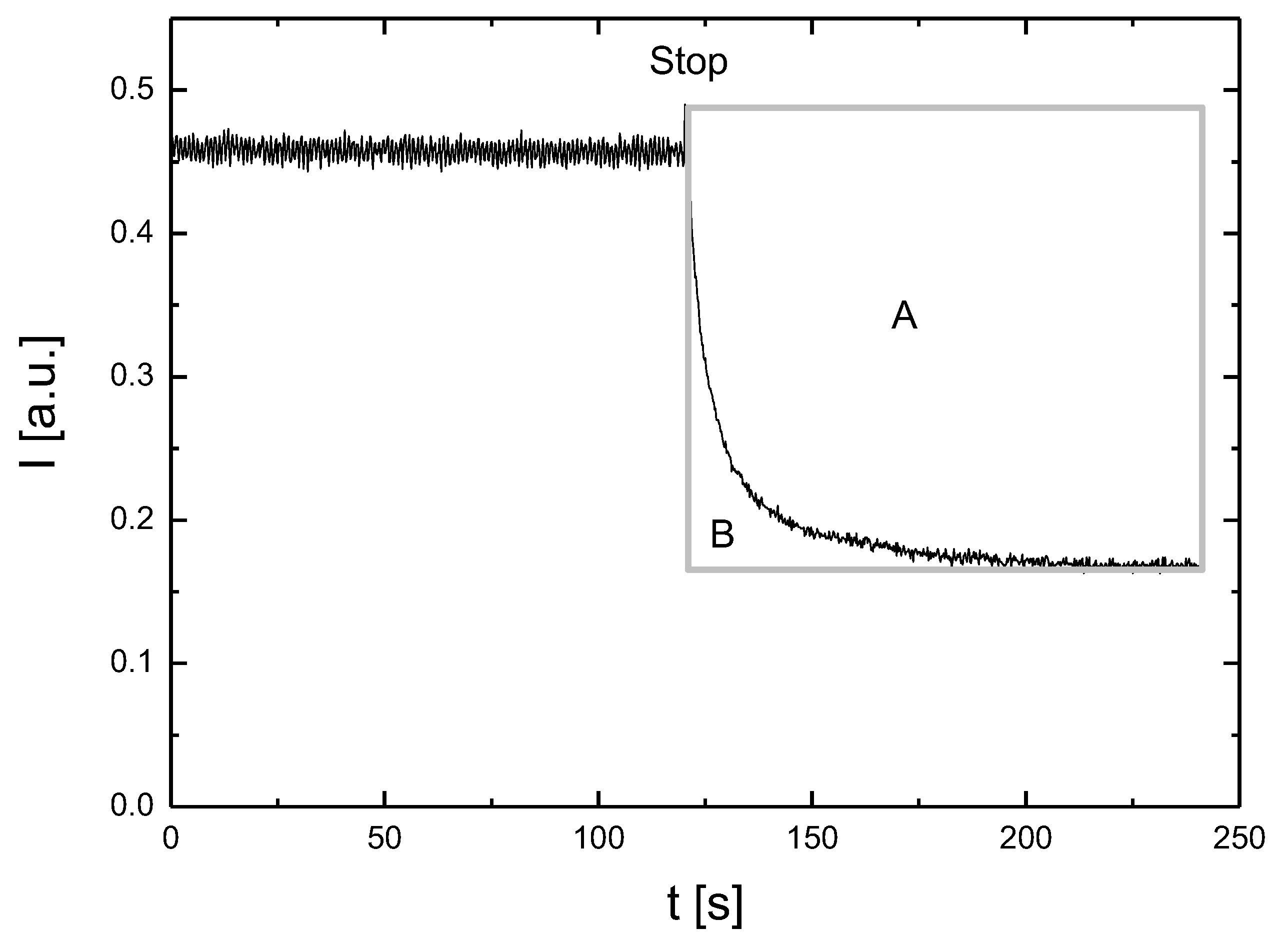
Each measurement technique has certain limitations. This is also the case for RBC aggregation measurements. When choosing the measurement technique, we were guided by the need to simultaneously measure the aggregation and incubation of cells. In addition, we were looking for a technique that would allow for relatively fast repeatable measurements. The choice fell on a photometric technique that meets the above-mentioned requirements. The approach proposed in this paper enables RBC aggregation measurements with a resolution of 4 min.
In conclusion, we performed a study of the effects of P188 in the presence of glucose on RBC aggregation. The aggregation was studied as a function of time. The study showed a synergistic effect of glucose and P188 on RBC aggregation and the temporary changes in this aggregation. To the best of our knowledge, the synergistic effect of P188 and glucose, as well as temporal changes, on aggregation are presented for the first time. The research method applied and the results obtained appear to be promising for further research in this area.
Author Contributions
Conceptualization, B.G. and A.S.-C.; methodology, B.G.; software, B.G.; validation, B.G. and A.S.-C.; formal analysis, B.G.; investigation, A.S.-C.; resources, B.G. and A.S.-C.; data curation, B.G. and A.S.-C.; writing—original draft preparation, A.S.-C.; writing—review and editing, B.G.; visualization, B.G. and A.S.-C.; supervision, B.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
This study was conducted according to the guidelines of the Declaration of Helsinki and was approved by the Bioethics Commission of Collegium Medicum Nicolaus Copernicus University (protocol code: KB 91/2003; date of approval: 25 October 2016).
Informed Consent Statement
Informed consent was obtained from all the subjects involved in the study.
Data Availability Statement
The data presented in this study are available on request from the corresponding author according to local policies.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Antonova, N.; Tsiberkin, K.; Podtaev, S.; Paskova, V.; Velcheva, I.; Chaushev, N. Comparative study between microvascular tone regulation and rheological properties of blood in patients with type 2 diabetes mellitus. Clin. Hemorheol. Microcirc. 2016, 64, 837–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Dévéhat, C.; Vimeux, M.; Khodabandehlou, T. Blood rheology in patients with diabetes mellitus. Clin. Hemorheol. Microcirc. 2004, 30, 297–300. [Google Scholar] [PubMed]
- Ditzel, J. Angioscopic Changes in the Smaller Blood Vessels in Diabetes Mellitus and their Relationship to Aging. Circulation 1956, 14, 386–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmid-Schönbein, H.; Volger, E. Red-cell aggregation and red-cell deformability in diabetes. Diabetes 1976, 25, 897–902. [Google Scholar]
- Van Haeringen, N.J.; Oosterhuis, J.A.; Terpstra, J.; Glasius, E. Erythrocyte aggregation in relation to diabetic retinopathy. Diabetologia 1973, 9, 20–24. [Google Scholar] [CrossRef]
- Demiroğlu, H.; Gürlek, A.; Barişta, I. Enhanced erythrocyte aggregation in type 2 diabetes with late complications. Exp. Clin. Endocrinol. Diabetes 1999, 107, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Babu, N.; Singh, M. Influence of hyperglycemia on aggregation, deformability and shape parameters of erythrocytes. Clin. Hemorheol. Microcirc. 2004, 31, 273–280. [Google Scholar]
- Chong-Martinez, B.; Buchanan, T.A.; Wenby, R.B.; Meiselman, H.J. Decreased red blood cell aggregation subsequent to improved glycaemic control in Type 2 diabetes mellitus. Diabet. Med. 2003, 20, 301–306. [Google Scholar] [CrossRef]
- Resmi, H.; Akhunlar, H.; Temiz Artmann, A.; Güner, G. In vitro effects of high glucose concentrations on membrane protein oxidation, G-actin and deformability of human erythrocytes. Cell. Biochem. Funct. 2005, 23, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Riquelme, B.; Foresto, P.; D’Arrigo, M.; Valverde, J.; Rasia, R. A dynamic and stationary rheological study of erythrocytes incubated in a glucose medium. J. Biochem. Biophys. Methods 2005, 62, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Ku, Y.-H.; Suh, J.-S.; Singh, M. Rheological characteristics of erythrocytes incubated in glucose media. Clin. Hemorheol. Microcirc. 2008, 38, 153–161. [Google Scholar] [PubMed]
- Viskupicova, J.; Blaskovic, D.; Galiniak, S.; Soszynski, M.; Bartosz, G.; Horakova, L.; Sadowska-Bartosz, I. Effect of high glucose concentrations on human erythrocytes in vitro. Redox Biol. 2015, 5, 381–387. [Google Scholar] [CrossRef] [Green Version]
- Shubhra, Q.T.H.; Tóth, J.; Gyenis, J.; Feczkó, T. Poloxamers for Surface Modification of Hydrophobic Drug Carriers and Their Effects on Drug Delivery. Polym. Rev. 2014, 54, 112–138. [Google Scholar] [CrossRef]
- Moghimi, S.; Hunter, A. Poloxamers and poloxamines in nanoparticle engineering and experimental medicine. Trends Biotechnol. 2000, 18, 412–420. [Google Scholar] [CrossRef]
- Patel, H.R.; Patel, R.P.; Patel, M.M. Poloxamers: A pharmaceutical excipients with therapeutic behaviors. Int. J. Pharmtech Res. 2009, 1, 299–303. [Google Scholar]
- Collins, J.M.; Despa, F.; Lee, R.C. Structural and functional recovery of electropermeabilized skeletal muscle in-vivo after treatment with surfactant poloxamer 188. Biochim. Biophys. Acta 2007, 1768, 1238–1246. [Google Scholar] [CrossRef] [Green Version]
- Maskarinec, S.; Hannig, J.; Lee, R.C.; Lee, K.Y.C. Direct Observation of Poloxamer 188 Insertion into Lipid Monolayers. Biophys. J. 2002, 82, 1453–1459. [Google Scholar] [CrossRef] [Green Version]
- Sandor, B.; Marin, M.; Lapoumeroulie, C.; Rabaï, M.; Lefevre, S.D.; Lemonne, N.; El Nemer, W.; Mozar, A.; Francais, O.; Le Pioufle, B.; et al. Effects of Poloxamer 188 on red blood cell membrane properties in sickle cell anaemia. Br. J. Haematol. 2016, 173, 145–149. [Google Scholar] [CrossRef]
- Toth, K.; Bogar, L.; Juricskay, I.; Keltai, M.; Yusuf, S.; Haywood, L.J.; Meiselman, H.J. The effect of RheothRx Injection on the hemorheological parameters in patients with acute myocardial infarction. Clin. Hemorheol. Microcirc. 1997, 17, 117–125. [Google Scholar]
- Lechmann, T.; Reinhart, W.H. The non-ionic surfactant Poloxamer 188 (RheothRx) increases plasma and whole blood viscosity. Clin. Hemorheol. Microcirc. 1998, 18, 31–36. [Google Scholar] [PubMed]
- Toth, K.; Wenby, R.B.; Meiselman, H.J. Inhibition of polymer-induced red blood cell aggregation by poloxamer 188. Biorheology 2000, 37, 301–312. [Google Scholar] [PubMed]
- Guzniczak, E.; Jimenez, M.; Irwin, M.; Otto, O.; Willoughby, N.; Bridle, H. Impact of poloxamer 188 (Pluronic F-68) additive on cell mechanical properties, quantification by real-time deformability cytometry. Biomicrofluidics 2018, 12, 044118. [Google Scholar] [CrossRef]
- Shaikh, A.; Yeole, P.G.; Iyer, D. Formulation and Evaluation of Stable Solid Dispersion of Anti-Protozoal Drug. Res. J. Pharm. Biol. Chem. Sci. 2013, 4, 1495–1509. [Google Scholar]
- Jain, D.; Gangshettiwar, A. Combination of lycopene, quercetin and poloxamer 188 alleviates anxiety and depression in 3-nitropropionic acid-induced Huntington’s disease in rats. J. Intercult. Ethnopharmacol. 2014, 3, 186–191. [Google Scholar] [CrossRef]
- Houang, E.M.; Bartos, J.; Hackel, B.J.; Lodge, T.P.; Yannopoulos, D.; Bates, F.S.; Metzger, J.M. Cardiac Muscle Membrane Stabilization in Myocardial Reperfusion Injury. JACC Basic Transl. Sci. 2019, 4, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Meiselman, H.J.; Neu, B. Red blood cell adhesion can be reduced by non-reactive macromolecules. Colloids Surf. B Biointerfaces 2019, 174, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Ebeling, P.; Koistinen, H.A.; Koivisto, V.A. Insulin-independent glucose transport regulates insulin sensitivity. FEBS Lett. 1998, 436, 301–303. [Google Scholar] [CrossRef]
- Brahm, J. Kinetics of glucose transport in human erythrocytes. J. Physiol. 1983, 339, 339–354. [Google Scholar] [CrossRef] [Green Version]
- Cloherty, E.K.; Heard, K.S.; Carruthers, A. Human Erythrocyte Sugar Transport is Incompatible with Available Carrier Models. Biochemistry 1996, 35, 10411–10421. [Google Scholar] [CrossRef]
- Baskurt, O.; Neu, B.; Meiselman, H.J. Red Blood Cell Aggregation; CRC Press: Boca Raton, FL, USA, 2012; Chapter 2.2.1; p. 12. [Google Scholar]
| Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






