
Serum Branched-Chain Amino Acid Metabolites Increase in Males When Aerobic Exercise Is Initiated with Low Muscle Glycogen

Abstract
:1. Introduction
2. Results
2.1. Muscle Glycogen and Substrate Oxidation
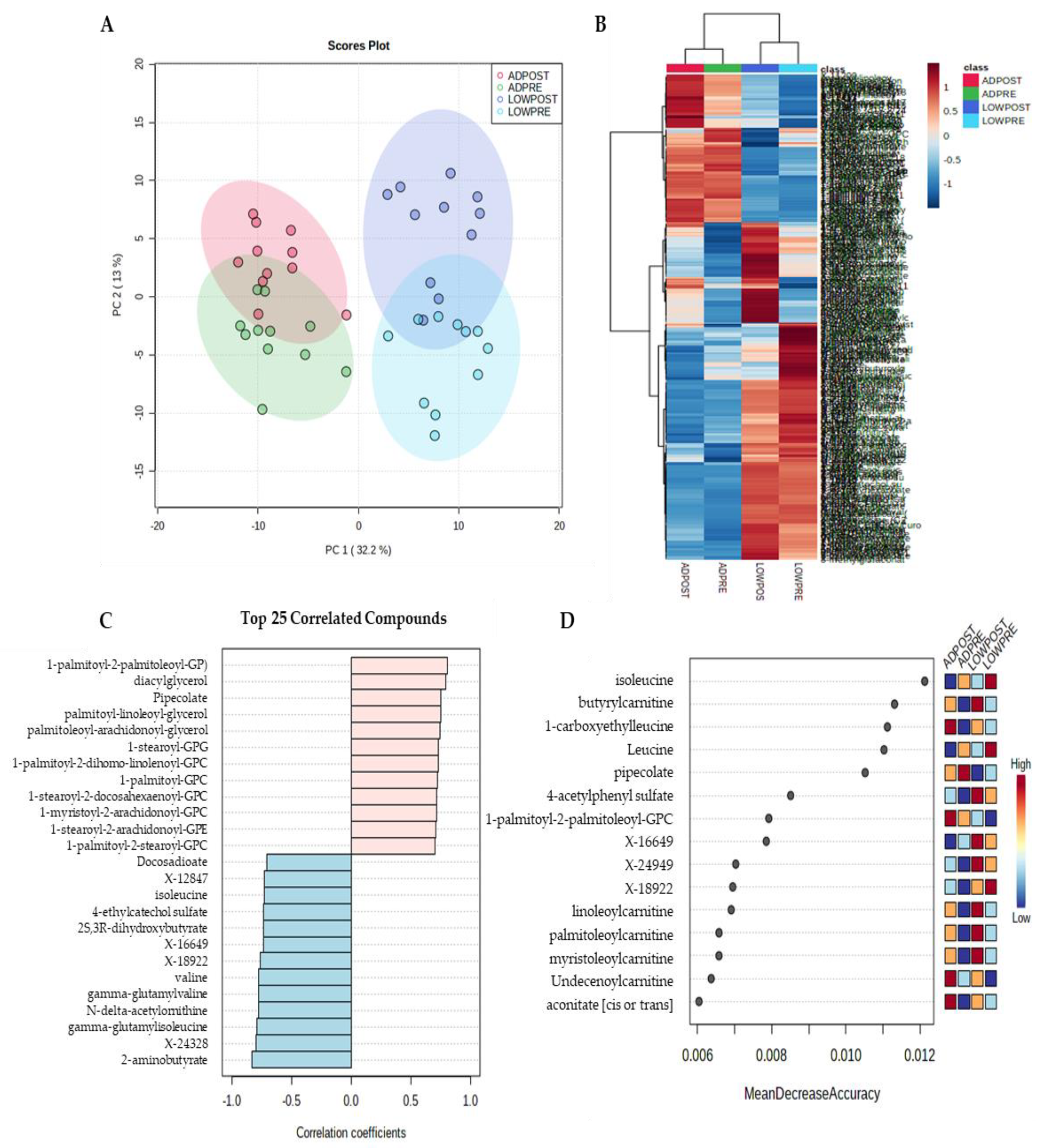
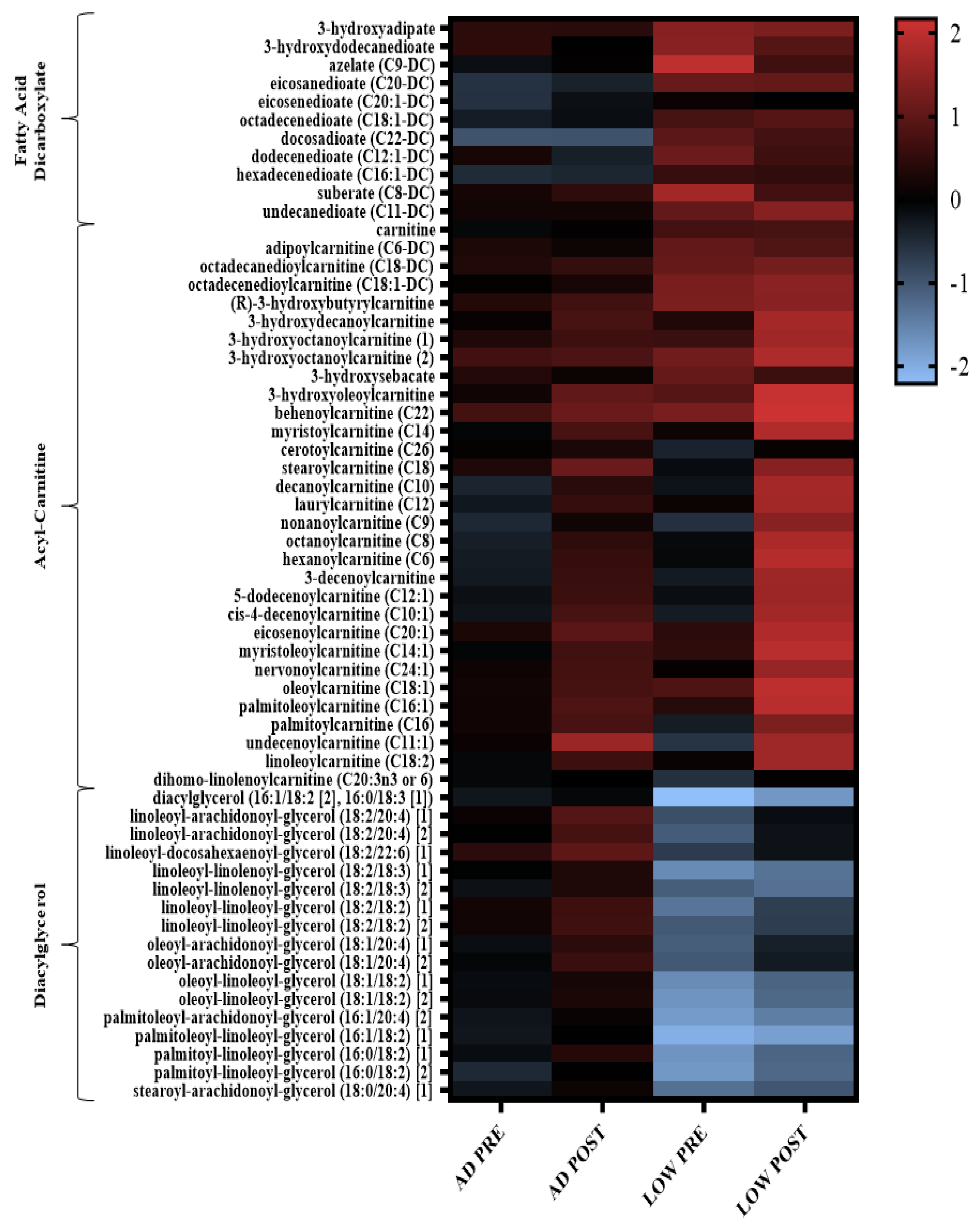
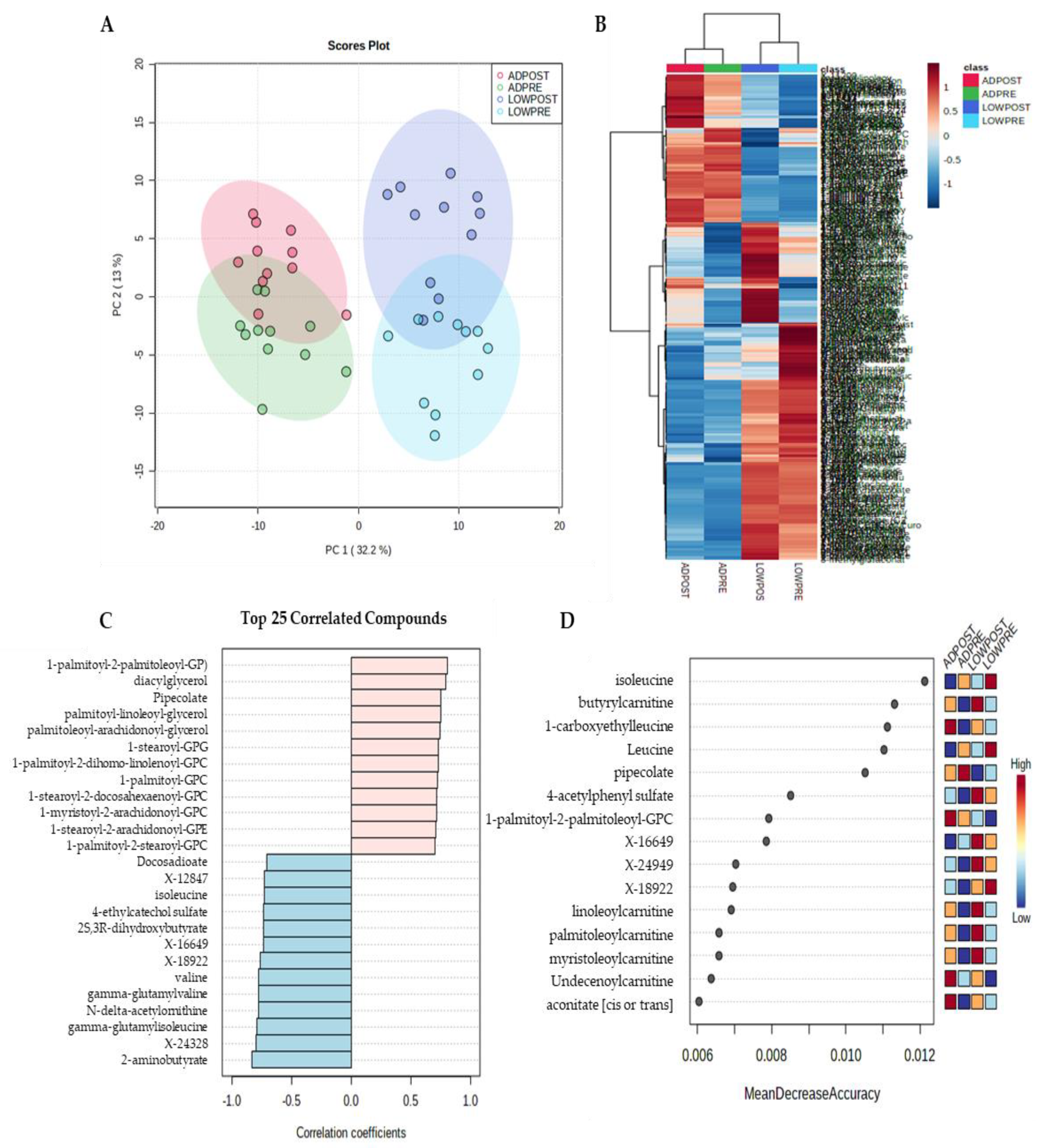
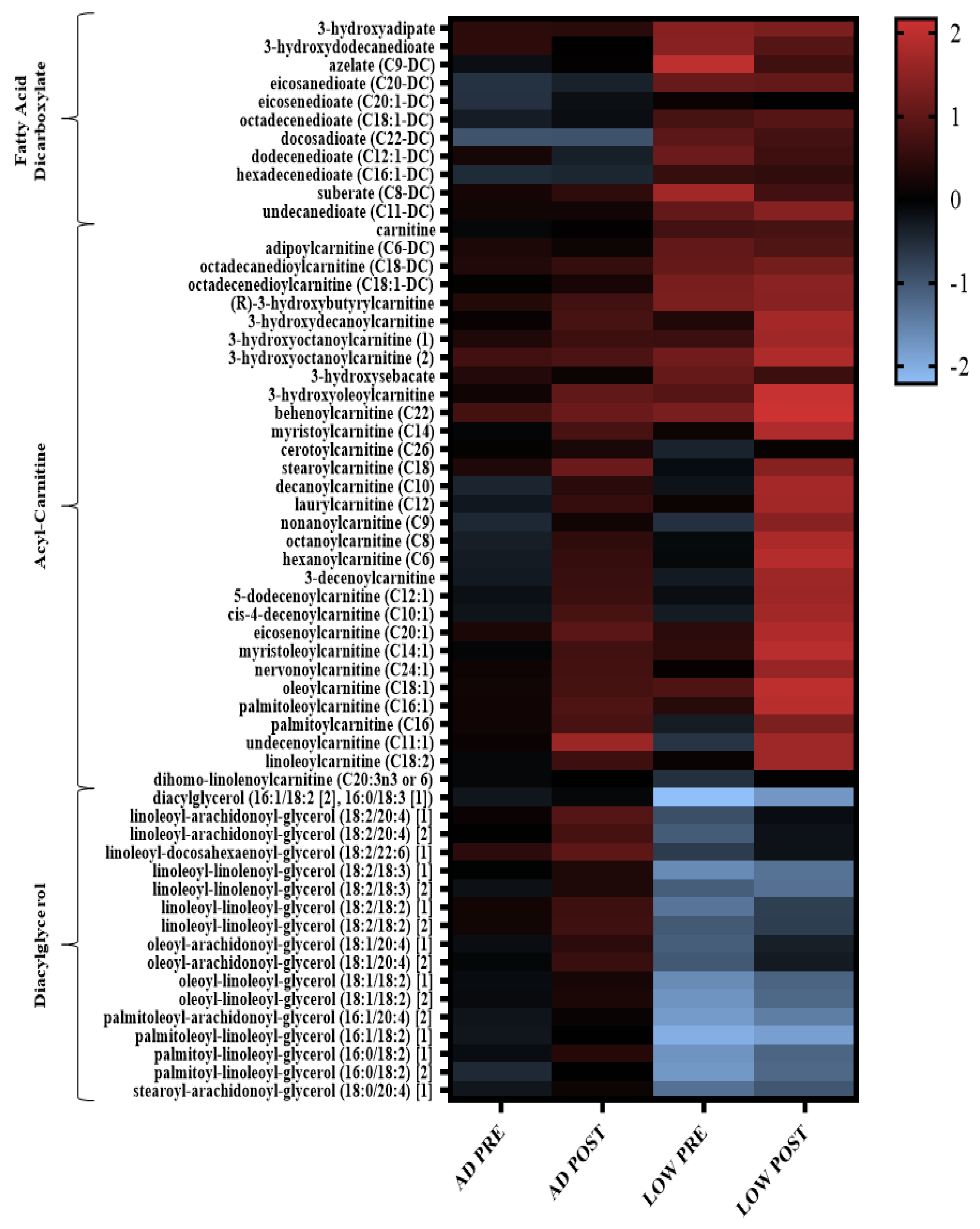
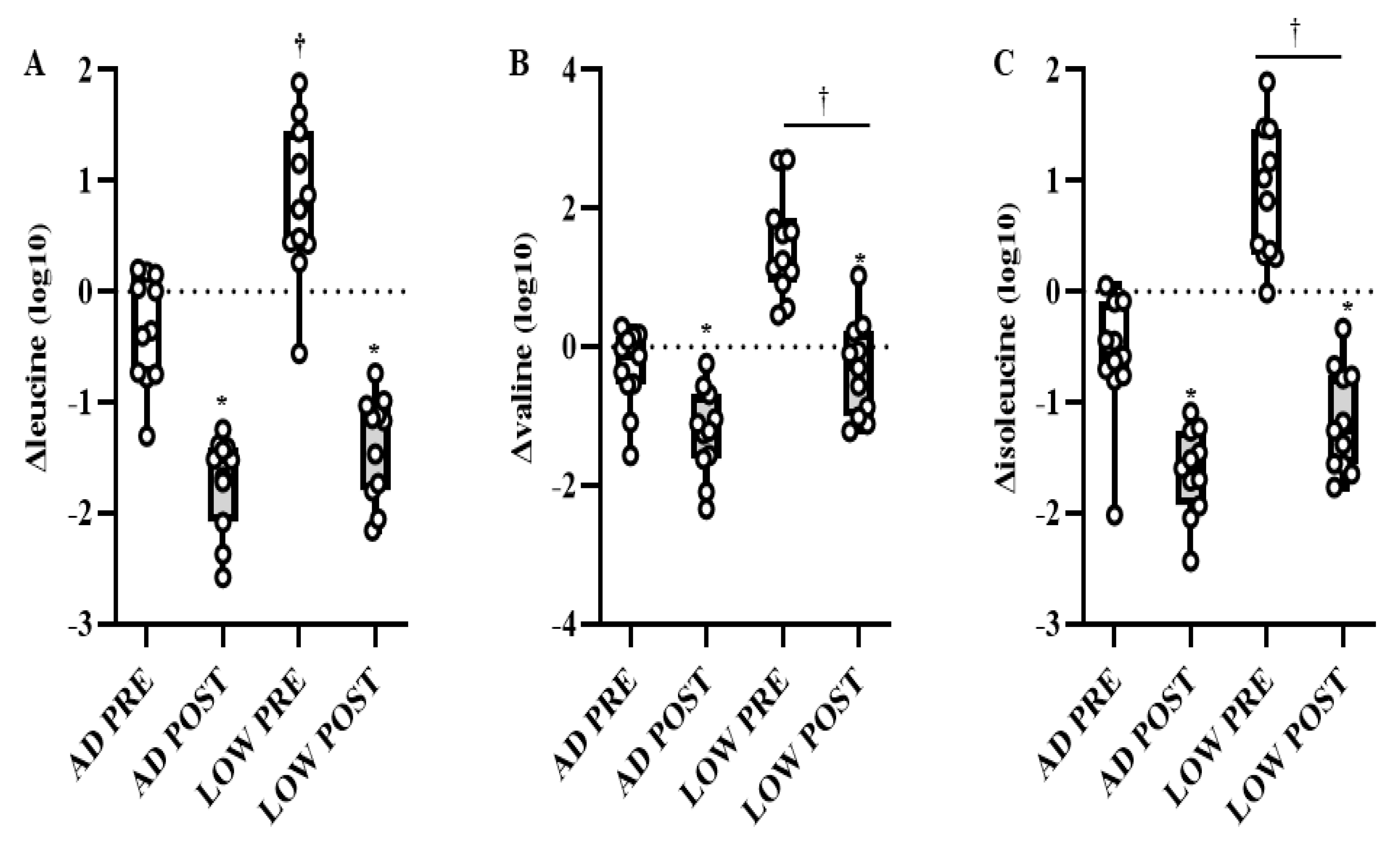
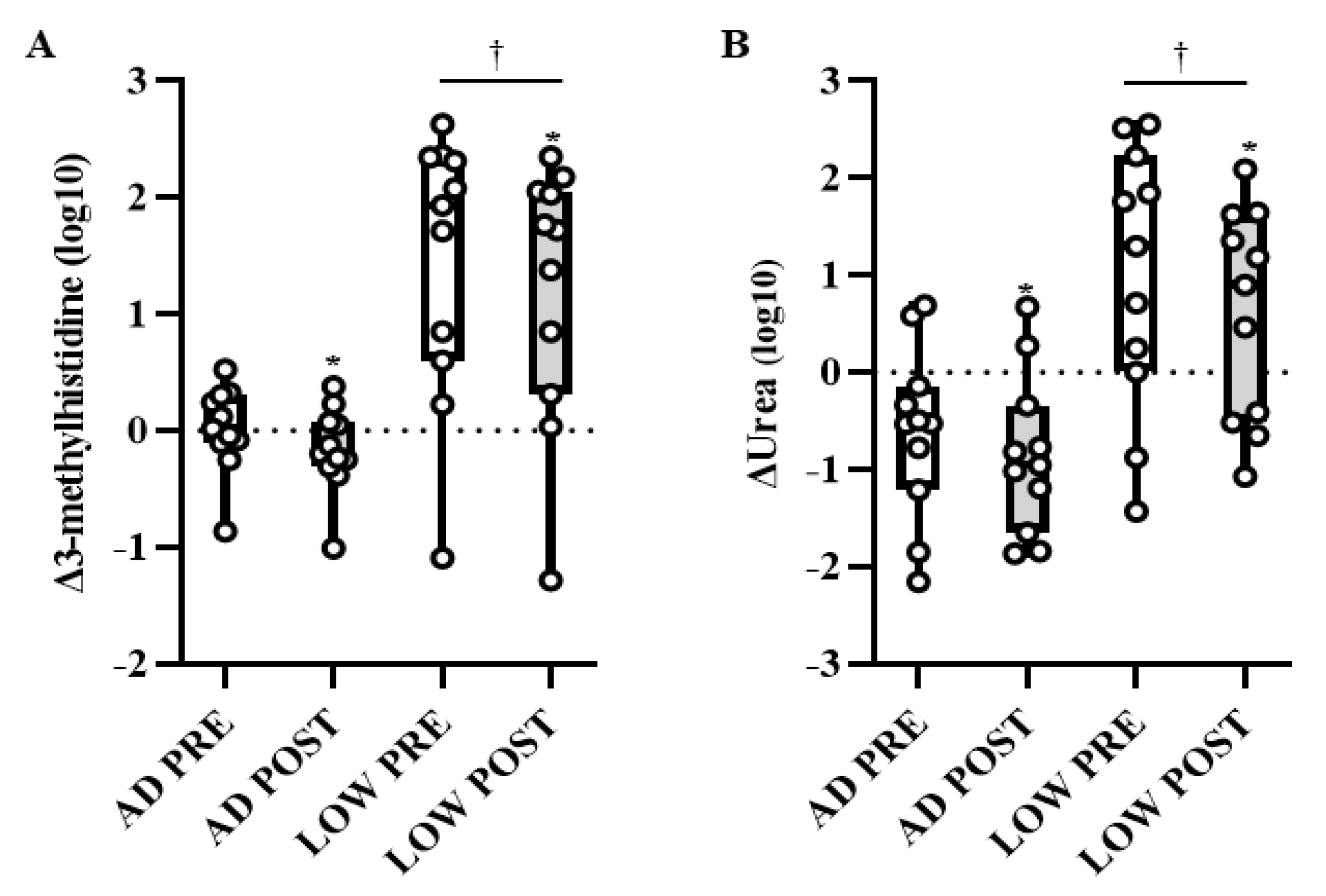
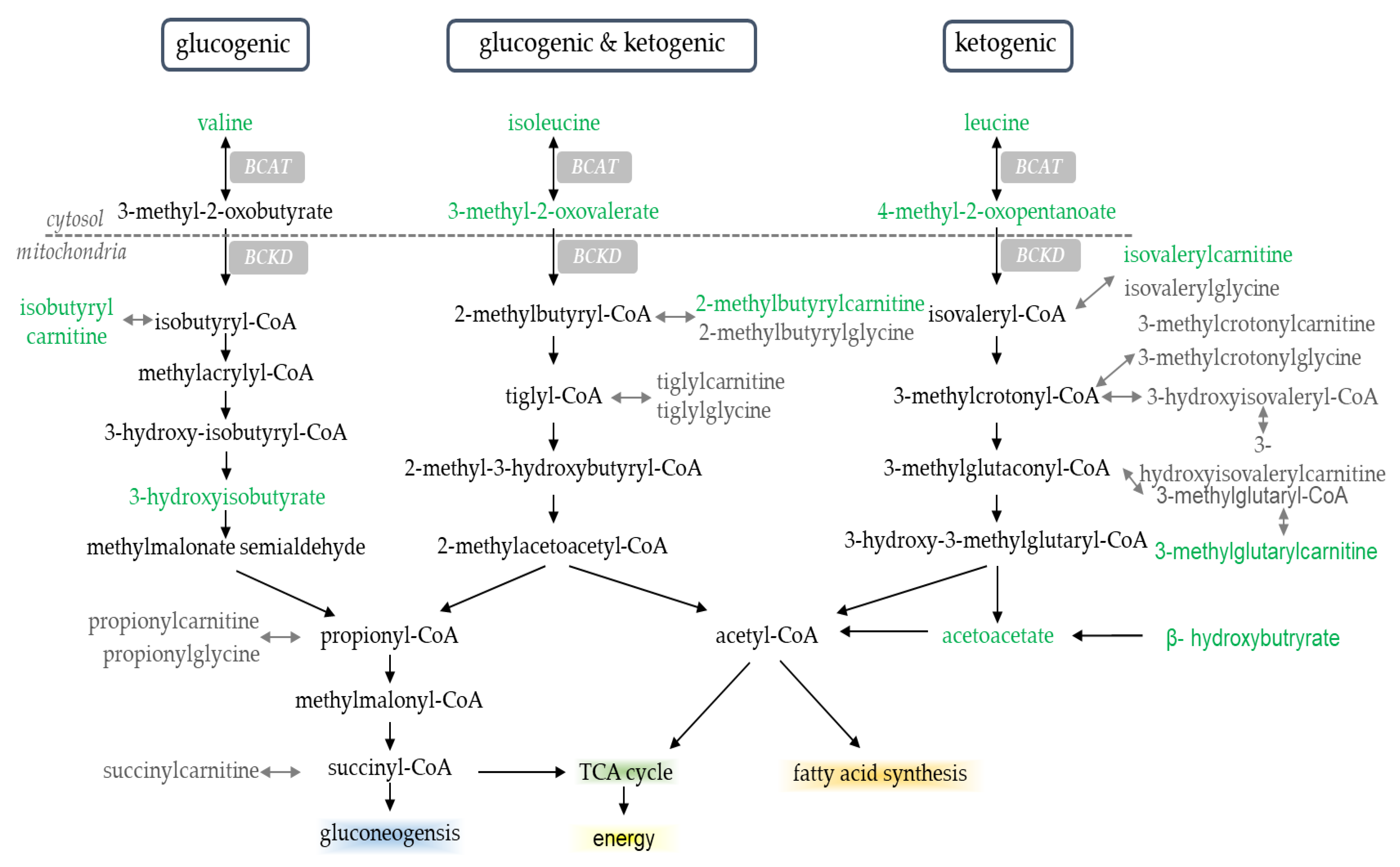
2.2. Serum Metabolites
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. Experimental Design
4.3. Glycogen Depletion Protocol
4.4. Study Diets
4.5. Steady-State Cycle Ergometry
4.6. Metabolomics
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Purdom, T.; Kravitz, L.; Dokladny, K.; Mermier, C. Understanding the factors that effect maximal fat oxidation. J. Int. Soc. Sports Nutr. 2018, 15, 3. [Google Scholar] [CrossRef] [Green Version]
- Arkinstall, M.J.; Bruce, C.R.; Clark, S.A.; Rickards, C.A.; Burke, L.M.; Hawley, J.A. Regulation of fuel metabolism by preexercise muscle glycogen content and exercise intensity. J. Appl. Physiol. 2004, 97, 2275–2283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hargreaves, M.; McConell, G.; Proietto, J. Influence of muscle glycogen on glycogenolysis and glucose uptake during exercise in humans. J. Appl. Physiol. 1995, 78, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Hearris, M.A.; Hammond, K.M.; Seaborne, R.A.; Stocks, B.; Shepherd, S.O.; Philp, A.; Sharples, A.P.; Morton, J.P.; Louis, J.B. Graded reductions in preexercise muscle glycogen impair exercise capacity but do not augment skeletal muscle cell signaling: Implications for cho periodization. J. Appl. Physiol. 2019, 126, 1587–1597. [Google Scholar] [CrossRef]
- Burke, L.M.; Angus, D.J.; Cox, G.R.; Cummings, N.K.; Febbraio, M.A.; Gawthorn, K.; Hawley, J.A.; Minehan, M.; Martin, D.T.; Hargreaves, M. Effect of fat adaptation and carbohydrate restoration on metabolism and performance during prolonged cycling. J. Appl. Physiol. 2000, 89, 2413–2421. [Google Scholar] [CrossRef] [Green Version]
- Burke, L.M.; Hawley, J.A. Effects of short-term fat adaptation on metabolism and performance of prolonged exercise. Med. Sci. Sports Exerc. 2002, 34, 1492–1498. [Google Scholar] [CrossRef]
- Stepto, N.K.; Carey, A.L.; Staudacher, H.M.; Cummings, N.K.; Burke, L.M.; Hawley, J.A. Effect of short-term fat adaptation on high-intensity training. Med. Sci. Sports Exerc. 2002, 34, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Webster, C.C.; Noakes, T.D.; Chacko, S.K.; Swart, J.; Kohn, T.A.; Smith, J.A. Gluconeogenesis during endurance exercise in cyclists habituated to a long-term low carbohydrate high-fat diet. J. Physiol. 2016, 594, 4389–4405. [Google Scholar] [CrossRef] [PubMed]
- Margolis, L.M.; Wilson, M.A.; Whitney, C.C.; Carrigan, C.T.; Murphy, N.E.; Hatch, A.M.; Montain, S.J.; Pasiakos, S.M. Exercising with low muscle glycogen content increases fat oxidation and decreases endogenous, but not exogenous carbohydrate oxidation. Metabolism 2019, 97, 1–8. [Google Scholar] [CrossRef]
- Howarth, K.R.; Phillips, S.M.; MacDonald, M.J.; Richards, D.; Moreau, N.A.; Gibala, M.J. Effect of glycogen availability on human skeletal muscle protein turnover during exercise and recovery. J. Appl. Physiol. 2010, 109, 431–438. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.S.; Holm, L.; Svart, M.V.; Hjelholt, A.J.; Bengtsen, M.B.; Dollerup, O.L.; Dalgaard, L.B.; Vendelbo, M.H.; van Hall, G.; Moller, N.; et al. Effects of protein intake prior to carbohydrate-restricted endurance exercise: A randomized crossover trial. J. Int. Soc. Sports Nutr. 2020, 17, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillen, J.B.; West, D.W.D.; Williamson, E.P.; Fung, H.J.W.; Moore, D.R. Low-carbohydrate training increases protein requirements of endurance athletes. Med. Sci. Sports Exerc. 2019, 51, 2294–2301. [Google Scholar] [CrossRef]
- Hammond, K.M.; Impey, S.G.; Currell, K.; Mitchell, N.; Shepherd, S.O.; Jeromson, S.; Hawley, J.A.; Close, G.L.; Hamilton, L.D.; Sharples, A.P.; et al. Postexercise high-fat feeding suppresses p70s6k1 activity in human skeletal muscle. Med. Sci. Sports Exerc. 2016, 48, 2108–2117. [Google Scholar] [CrossRef]
- Impey, S.G.; Hammond, K.M.; Shepherd, S.O.; Sharples, A.P.; Stewart, C.; Limb, M.; Smith, K.; Philp, A.; Jeromson, S.; Hamilton, D.L.; et al. Fuel for the work required: A practical approach to amalgamating train-low paradigms for endurance athletes. Physiol. Rep. 2016, 4, e12803. [Google Scholar] [CrossRef]
- Margolis, L.M.; Wilson, M.A.; Whitney, C.C.; Carrigan, C.T.; Murphy, N.E.; Hatch-McChesney, A.; Pasiakos, S.M. Initiating aerobic exercise with low glycogen content reduces markers of myogenesis but not mtorc1 signaling. J. Int. Soc. Sports Nutr. 2021, 18, 56. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Lindon, J.C.; Holmes, E. ‘Metabonomics’: Understanding the metabolic responses of living systems to pathophysiological stimuli via multivariate statistical analysis of biological nmr spectroscopic data. Xenobiotica 1999, 29, 1181–1189. [Google Scholar] [CrossRef]
- Sampson, D.L.; Broadbent, J.A.; Parker, A.W.; Upton, Z.; Parker, T.J. Urinary biomarkers of physical activity: Candidates and clinical utility. Expert Rev. Proteomics 2014, 11, 91–106. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, C.A.; Nieman, D.C.; Signini, E.F.; Abreu, R.M.; Catai, A.M. Metabolomics-based studies assessing exercise-induced alterations of the human metabolome: A systematic review. Metabolites 2019, 9, 164. [Google Scholar] [CrossRef] [Green Version]
- Schranner, D.; Kastenmuller, G.; Schonfelder, M.; Romisch-Margl, W.; Wackerhage, H. Metabolite concentration changes in humans after a bout of exercise: A systematic review of exercise metabolomics studies. Sports Med. Open 2020, 6, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stander, Z.; Luies, L.; Mienie, L.J.; Van Reenen, M.; Howatson, G.; Keane, K.M.; Clifford, T.; Stevenson, E.J.; Loots, D.T. The unaided recovery of marathon-induced serum metabolome alterations. Sci. Rep. 2020, 10, 11060. [Google Scholar] [CrossRef] [PubMed]
- Stander, Z.; Luies, L.; Mienie, L.J.; Keane, K.M.; Howatson, G.; Clifford, T.; Stevenson, E.J.; Loots, D.T. The altered human serum metabolome induced by a marathon. Metabolomics 2018, 14, 150. [Google Scholar] [CrossRef] [PubMed]
- Lewis, G.D.; Farrell, L.; Wood, M.J.; Martinovic, M.; Arany, Z.; Rowe, G.C.; Souza, A.; Cheng, S.; McCabe, E.L.; Yang, E.; et al. Metabolic signatures of exercise in human plasma. Sci. Transl. Med. 2010, 2, 33ra37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margolis, L.M.; Karl, J.P.; Wilson, M.A.; Coleman, J.L.; Ferrando, A.A.; Young, A.J.; Pasiakos, S.M. Metabolomic profiles are reflective of hypoxia-induced insulin resistance during exercise in healthy young adult males. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2021, 321, R1–R11. [Google Scholar] [CrossRef] [PubMed]
- Dhahbi, J.M.; Mote, P.L.; Wingo, J.; Rowley, B.C.; Cao, S.X.; Walford, R.L.; Spindler, S.R. Caloric restriction alters the feeding response of key metabolic enzyme genes. Mech. Ageing Dev. 2001, 122, 1033–1048. [Google Scholar] [CrossRef]
- Hagopian, K.; Ramsey, J.J.; Weindruch, R. Caloric re.estriction increases gluconeogenic and transaminase enzyme activities in mouse liver. Exp. Gerontol. 2003, 38, 267–278. [Google Scholar] [CrossRef]
- Howe, C.C.F.; Alshehri, A.; Muggeridge, D.; Mullen, A.B.; Boyd, M.; Spendiff, O.; Moir, H.J.; Watson, D.G. Untargeted metabolomics profiling of an 80.5 km simulated treadmill ultramarathon. Metabolites 2018, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Bester, R.; Stander, Z.; Mason, S.; Keane, K.M.; Howatson, G.; Clifford, T.; Stevenson, E.J.; Loots, D.T. Characterizing marathon-induced metabolic changes using 1h-nmr metabolomics. Metabolites 2021, 11, 656. [Google Scholar] [CrossRef]
- Neinast, M.; Murashige, D.; Arany, Z. Branched chain amino acids. Annu. Rev. Physiol. 2019, 81, 139–164. [Google Scholar] [CrossRef]
- Margolis, L.M.; Carbone, J.W.; Berryman, C.E.; Carrigan, C.T.; Murphy, N.E.; Ferrando, A.A.; Young, A.J.; Pasiakos, S.M. Severe energy deficit at high altitude inhibits skeletal muscle mtorc1-mediated anabolic signaling without increased ubiquitin proteasome activity. FASEB J. 2018, 32, 5955–5966. [Google Scholar] [CrossRef] [PubMed]
- Nieman, D.C.; Sha, W.; Pappan, K.L. Il-6 linkage to exercise-induced shifts in lipid-related metabolites: A metabolomics-based analysis. J. Proteome Res. 2017, 16, 970–977. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, R.; Zhao, X.; Weigert, C.; Simon, P.; Fehrenbach, E.; Fritsche, J.; Machann, J.; Schick, F.; Wang, J.; Hoene, M.; et al. Medium chain acylcarnitines dominate the metabolite pattern in humans under moderate intensity exercise and support lipid oxidation. PLoS ONE 2010, 5, e11519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karl, J.P.; Margolis, L.M.; Murphy, N.E.; Carrigan, C.T.; Castellani, J.W.; Madslien, E.H.; Teien, H.K.; Martini, S.; Montain, S.J.; Pasiakos, S.M. Military training elicits marked increases in plasma metabolomic signatures of energy metabolism, lipolysis, fatty acid oxidation, and ketogenesis. Physiol. Rep. 2017, 5, e13407. [Google Scholar] [CrossRef]
- Xu, G.; Hansen, J.S.; Zhao, X.J.; Chen, S.; Hoene, M.; Wang, X.L.; Clemmesen, J.O.; Secher, N.H.; Haring, H.U.; Pedersen, B.K.; et al. Liver and muscle contribute differently to the plasma acylcarnitine pool during fasting and exercise in humans. J. Clin. Endocrinol. Metab. 2016, 101, 5044–5052. [Google Scholar] [CrossRef] [PubMed]
- Lundsgaard, A.M.; Fritzen, A.M.; Kiens, B. Molecular regulation of fatty acid oxidation in skeletal muscle during aerobic exercise. Trends Endocrinol. Metab. 2018, 29, 18–30. [Google Scholar] [CrossRef]
- Krug, S.; Kastenmuller, G.; Stuckler, F.; Rist, M.J.; Skurk, T.; Sailer, M.; Raffler, J.; Romisch-Margl, W.; Adamski, J.; Prehn, C.; et al. The dynamic range of the human metabolome revealed by challenges. FASEB J. 2012, 26, 2607–2619. [Google Scholar] [CrossRef]
- Newgard, C.B. Interplay between lipids and branched-chain amino acids in development of insulin resistance. Cell Metab. 2012, 15, 606–614. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Shao, J.; Wu, C.Y.; Shu, L.; Dong, W.; Liu, Y.; Chen, M.; Wynn, R.M.; Wang, J.; Wang, J.; et al. Targeting bcaa catabolism to treat obesity-associated insulin resistance. Diabetes 2019, 68, 1730–1746. [Google Scholar] [CrossRef] [Green Version]
- Shao, D.; Villet, O.; Zhang, Z.; Choi, S.W.; Yan, J.; Ritterhoff, J.; Gu, H.; Djukovic, D.; Christodoulou, D.; Kolwicz, S.C., Jr.; et al. Glucose promotes cell growth by suppressing branched-chain amino acid degradation. Nat. Commun. 2018, 9, 2935. [Google Scholar] [CrossRef] [Green Version]
- Hsueh, T.Y.; Baum, J.I.; Huang, Y. Effect of eicosapentaenoic acid and docosahexaenoic acid on myogenesis and mitochondrial biosynthesis during murine skeletal muscle cell differentiation. Front. Nutr. 2018, 5, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gwin, J.A.; Church, D.D.; Wolfe, R.R.; Ferrando, A.A.; Pasiakos, S.M. Muscle protein synthesis and whole-body protein turnover responses to ingesting essential amino acids, intact protein, and protein-containing mixed meals with considerations for energy deficit. Nutrients 2020, 12, 2457. [Google Scholar] [CrossRef]
- Church, D.D.; Hirsch, K.R.; Park, S.; Kim, I.Y.; Gwin, J.A.; Pasiakos, S.M.; Wolfe, R.R.; Ferrando, A.A. Essential amino acids and protein synthesis: Insights into maximizing the muscle and whole-body response to feeding. Nutrients 2020, 12, 3717. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, R.R. Branched-chain amino acids and muscle protein synthesis in humans: Myth or reality? J. Int. Soc. Sports Nutr. 2017, 14, 30. [Google Scholar] [CrossRef] [Green Version]
- Areta, J.L.; Burke, L.M.; Camera, D.M.; West, D.W.; Crawshay, S.; Moore, D.R.; Stellingwerff, T.; Phillips, S.M.; Hawley, J.A.; Coffey, V.G. Reduced resting skeletal muscle protein synthesis is rescued by resistance exercise and protein ingestion following short-term energy deficit. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E989–E997. [Google Scholar] [CrossRef] [PubMed]
- Gwin, J.A.; Church, D.D.; Hatch-McChesney, A.; Howard, E.E.; Carrigan, C.T.; Murphy, N.E.; Wilson, M.A.; Margolis, L.M.; Carbone, J.W.; Wolfe, R.R.; et al. Effects of high versus standard essential amino acid intakes on whole-body protein turnover and mixed muscle protein synthesis during energy deficit: A randomized, crossover study. Clin. Nutr. 2021, 40, 767–777. [Google Scholar] [CrossRef]
- Nieman, D.C.; Scherr, J.; Luo, B.; Meaney, M.P.; Dreau, D.; Sha, W.; Dew, D.A.; Henson, D.A.; Pappan, K.L. Influence of pistachios on performance and exercise-induced inflammation, oxidative stress, immune dysfunction, and metabolite shifts in cyclists: A randomized, crossover trial. PLoS ONE 2014, 9, e113725. [Google Scholar] [CrossRef] [Green Version]
- Nieman, D.C.; Gillitt, N.D.; Sha, W.; Meaney, M.P.; John, C.; Pappan, K.L.; Kinchen, J.M. Metabolomics-based analysis of banana and pear ingestion on exercise performance and recovery. J. Proteome Res. 2015, 14, 5367–5377. [Google Scholar] [CrossRef] [PubMed]
- Nieman, D.C.; Shanely, R.A.; Luo, B.; Meaney, M.P.; Dew, D.A.; Pappan, K.L. Metabolomics approach to assessing plasma 13- and 9-hydroxy-octadecadienoic acid and linoleic acid metabolite responses to 75-km cycling. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, R68–R74. [Google Scholar] [CrossRef] [Green Version]
- Nieman, D.C.; Shanely, R.A.; Gillitt, N.D.; Pappan, K.L.; Lila, M.A. Serum metabolic signatures induced by a three-day intensified exercise period persist after 14 h of recovery in runners. J. Proteome Res. 2013, 12, 4577–4584. [Google Scholar] [CrossRef]
- Magkos, F. Basal very low-density lipoprotein metabolism in response to exercise: Mechanisms of hypotriacylglycerolemia. Prog. Lipid Res. 2009, 48, 171–190. [Google Scholar] [CrossRef]
- Miura, Y. The biological significance of omega-oxidation of fatty acids. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2013, 89, 370–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brennan, L.; Gibbons, H. Sex matters: A focus on the impact of biological sex on metabolomic profiles and dietary interventions. Proc. Nutr. Soc. 2020, 79, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Draper, C.F.; Duisters, K.; Weger, B.; Chakrabarti, A.; Harms, A.C.; Brennan, L.; Hankemeier, T.; Goulet, L.; Konz, T.; Martin, F.P.; et al. Menstrual cycle rhythmicity: Metabolic patterns in healthy women. Sci. Rep. 2018, 8, 14568. [Google Scholar] [CrossRef]
- Al-Khelaifi, F.; Diboun, I.; Donati, F.; Botre, F.; Alsayrafi, M.; Georgakopoulos, C.; Suhre, K.; Yousri, N.A.; Elrayess, M.A. A pilot study comparing the metabolic profiles of elite-level athletes from different sporting disciplines. Sports Med. Open 2018, 4, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McInerney, P.; Lessard, S.J.; Burke, L.M.; Coffey, V.G.; Lo Giudice, S.L.; Southgate, R.J.; Hawley, J.A. Failure to repeatedly supercompensate muscle glycogen stores in highly trained men. Med. Sci. Sports. Exerc. 2005, 37, 404–411. [Google Scholar] [CrossRef]
- Dehaven, C.D.; Evans, A.M.; Dai, H.; Lawton, K.A. Organization of gc/ms and lc/ms metabolomics data into chemical libraries. J. Cheminform. 2010, 2, 9. [Google Scholar] [CrossRef] [Green Version]
- Evans, A.M.; DeHaven, C.D.; Barrett, T.; Mitchell, M.; Milgram, E. Integrated, nontargeted ultrahigh performance liquid chromatography/electrospray ionization tandem mass spectrometry platform for the identification and relative quantification of the small-molecule complement of biological systems. Anal. Chem. 2009, 81, 6656–6667. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis chemical analysis working group (cawg) metabolomics standards initiative (msi). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Wishart, D.S. Using metaboanalyst 3.0 for comprehensive metabolomics data analysis. Curr. Protoc. Bioinform. 2016, 55, 14.10.11–14.10.91. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Low | Adequate | p Value | |
|---|---|---|---|
| Energy (kcal/d) | 3081 ± 374 | 3086 ± 347 | 0.77 |
| Carbohydrate (g/kg/d) | 1.5 ± 0.1 | 6.0 ± 0.2 | <0.01 |
| Fat (g/kg/d) | 3.0 ± 0.5 | 1.0 ± 0.5 | <0.01 |
| Protein (g/kg/d) | 1.3 ± 0.5 | 1.2 ± 0.5 | <0.01 |
| AD | LOW | p, Q Values | |||||
|---|---|---|---|---|---|---|---|
| Metabolite | PRE | POST | PRE | POST | Time | Treatment | T × T |
| 1-carboxyethylleucine | −0.47 ± 0.31 | 1.59 ± 0.40 | 0.11 ± 0.77 | 1.60 ± 0.66 | <0.01, <0.01 | 0.19, 0.20 | 0.01, 0.04 |
| 2-hydroxy-3-methylvalerate | −0.08 ± 0.31 | 0.13 ± 0.41 | 1.27 ± 0.61 | 1.00 ± 0.87 | 0.77, 0.79 | <0.01, <0.01 | 0.03, 0.11 |
| 3-methyl-2-oxovalerate | −0.42 ± 0.65 | −0.06 ± 0.59 | 0.59 ± 0.83 | 0.16 ± 0.82 | 0.70, 0.73 | 0.05, 0.06 | <0.01, <0.01 |
| 4-methyl-2-oxopentanoate | −0.27 ± 0.70 | 0.18 ± 0.70 | 0.50 ± 1.05 † | 0.00 ± 1.04 | 0.81, 0.83 | 0.43, 0.43 | <0.01, <0.01 |
| alpha-hydroxyisocaproate | −0.17 ± 0.48 | −0.32 ± 0.83 | 0.98 ± 0.74 † | 0.01 ± 1.00 * | <0.01, <0.01 | 0.03, 0.03 | 0.01, 0.04 |
| alpha-hydroxyisovalerate | 0.09 ± 0.51 | 0.26 ± 0.45 | 1.34 ± 0.57 | 1.30 ± 0.72 | 0.24, 0.29 | <0.01, <0.01 | 0.06, 0.19 |
| beta-hydroxyisovalerate | 0.03 ± 0.41 | 0.08 ± 0.35 | 0.91 ± 0.50 | 0.87 ± 0.54 | 0.93, 0.94 | <0.01, <0.01 | 0.26, 0.51 |
| ethylmalonate | 0.01 ± 0.29 | 0.46 ± 0.37 | 0.27 ± 0.22 | 1.01 ± 0.66 | <0.01, <0.01 | 0.02, 0.03 | 0.07, 0.22 |
| N-acetylisoleucine | −0.74 ± 0.54 | −0.09 ± 0.71 * | 0.88 ± 1.27 † | 0.35 ± 1.48 | 0.82, 0.84 | 0.01, 0.02 | 0.03, 0.11 |
| 2-ketocaprylate | −0.20 ± 0.37 | 0.06 ± 0.35 | 0.76 ± 0.68 | 1.26 ± 0.49 | <0.01, <0.01 | <0.01, <0.01 | 0.15, 0.35 |
| 2-methylbutyrylcarnitine (C5) | −0.28 ± 0.49 | 0.30 ± 0.63 | 0.69 ± 0.41 | 1.82 ± 0.62 | <0.01, <0.01 | <0.01, <0.01 | 0.02, 0.10 |
| 3-hydroxyisobutyrate | −0.06 ± 0.69 | 0.95 ± 0.69 | 0.88 ± 0.63 | 1.77 ± 0.78 | <0.01, <0.01 | <0.01, <0.01 | 0.74, 0.86 |
| 3-methylglutaconate | −0.04 ± 0.39 | 0.15 ± 0.46 | 0.67 ± 0.68 | 1.23 ± 0.65 | <0.01, <0.01 | <0.01, <0.01 | 0.05, 0.16 |
| 3-methylglutarylcarnitine (2) | 0.12 ± 0.48 | 0.12 ± 0.60 | −0.84 ± 0.40 | −0.49 ± 0.45 | 0.12, 0.16 | <0.01, <0.01 | 0.12, 0.30 |
| isobutyrylcarnitine (C4) | −0.26 ± 0.50 | 0.18 ± 0.68 | 0.57 ± 0.88 | 1.59 ± 0.71 | <0.01, <0.01 | <0.01, <0.01 | 0.02, 0.10 |
| isovalerylcarnitine (C5) | 0.02 ± 1.16 | 0.73 ± 0.94 | 0.41 ± 0.53 | 1.99 ± 0.74 †* | <0.01, <0.01 | 0.01, 0.02 | 0.06, 0.18 |
| n-acetylleucine | −0.83 ± 0.93 | 0.00 ± 0.91 | 0.36 ± 0.87 | 0.34 ± 1.19 | 0.02, 0.03 | 0.06, 0.07 | 0.01, 0.07 |
| butyrylcarnitine (C4) | −0.27 ± 0.23 | 0.47 ± 0.28 | −0.26 ± 0.18 | 1.04 ± 0.56 †* | 0.00, 0.00 | 0.02, 0.03 | 0.01, 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Margolis, L.M.; Karl, J.P.; Wilson, M.A.; Coleman, J.L.; Whitney, C.C.; Pasiakos, S.M. Serum Branched-Chain Amino Acid Metabolites Increase in Males When Aerobic Exercise Is Initiated with Low Muscle Glycogen. Metabolites 2021, 11, 828. https://doi.org/10.3390/metabo11120828
Margolis LM, Karl JP, Wilson MA, Coleman JL, Whitney CC, Pasiakos SM. Serum Branched-Chain Amino Acid Metabolites Increase in Males When Aerobic Exercise Is Initiated with Low Muscle Glycogen. Metabolites. 2021; 11(12):828. https://doi.org/10.3390/metabo11120828
Chicago/Turabian StyleMargolis, Lee M., J Philip Karl, Marques A. Wilson, Julie L. Coleman, Claire C. Whitney, and Stefan M. Pasiakos. 2021. "Serum Branched-Chain Amino Acid Metabolites Increase in Males When Aerobic Exercise Is Initiated with Low Muscle Glycogen" Metabolites 11, no. 12: 828. https://doi.org/10.3390/metabo11120828
APA StyleMargolis, L. M., Karl, J. P., Wilson, M. A., Coleman, J. L., Whitney, C. C., & Pasiakos, S. M. (2021). Serum Branched-Chain Amino Acid Metabolites Increase in Males When Aerobic Exercise Is Initiated with Low Muscle Glycogen. Metabolites, 11(12), 828. https://doi.org/10.3390/metabo11120828