The Bovine Metabolome

 , ,
, , 
Abstract
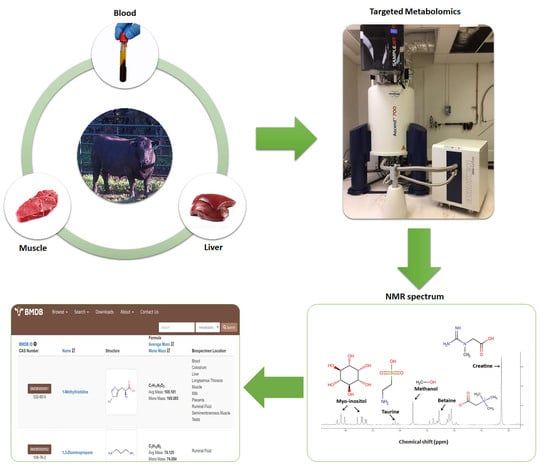
1. Introduction
2. Results
2.1. Water-Soluble Compound Identification and Quantification by NMR and LC–MS/MS
2.2. Lipid-Like Compound Identification and Quantification by LC–MS/MS
2.3. Trace Element Identification and Quantification by ICP–MS
2.4. The Chemical Composition of Bovine Biofluids and Tissues (Experimental Data)
2.5. Literature Survey of Bovine Biofluids and Tissues Metabolites
2.6. The BMDB Website
3. Discussion
3.1. Comparisons to Other Studies
3.2. Comparisons Across Platforms
4. Materials and Methods
4.1. Ethics Approvals
4.2. Animal Selection
4.3. Sample Collection
4.4. Biofluid Sample Preparation for NMR
4.5. Tissue Sample Preparation for NMR
4.6. NMR Spectroscopy
NMR Compound Identification and Quantification
4.7. LC–MS/MS Compound Identification and Quantification
4.8. Trace Elemental Analyses Using ICP–MS
4.9. Literature Research on Bovine Biofluid and Tissue Metabolites
4.10. Genome Scale Inference of Expected Bovine Metabolites
4.11. Construction of the BMDB
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Market Research Report. 2019. Beef Market Size, Share & Trends Analysis Report by Cut (Brisket, Shank, Loin), by Slaughter Method (Kosher, Halal), by Region (North America, Europe, APAC, MEA, CSA), and Segment Forecasts, 2019–2025. Available online: https://www.grandviewresearch.com/industry-analysis/beef-market-analysis (accessed on 17 November 2019).
- Statista. 2019. Production of Meat Worldwide from 2016 to 2019, by Type (in Million Metric Tons). Available online: https://www.statista.com/statistics/237632/production-of-meat-worldwide-since-1990/ (accessed on 17 November 2019).
- Statista. 2019. Milk Products. Available online: https://www.statista.com/outlook/40010000/100/milk-products/worldwide (accessed on 17 November 2019).
- Food and Agriculture Organization. 2017. Dairy Market Review. Available online: http://www.fao.org/3/I9210EN/i9210en.pdf/ (accessed on 21 September 2018).
- Food and Agriculture Organization. 2017. Gateway to Dairy Production and Products. Available online: http://www.fao.org/dairy-production-products/production/dairy-animals/en/ (accessed on 21 September 2018).
- Albenzio, M.; Santillo, A.; Ciliberti, M.G.; Figliola, L.; Caroprese, M.; Marino, R.; Polito, A.N. Milk from different species: Relationship between protein fractions and inflammatory response in infants affected by generalized epilepsy. J. Dairy Sci. 2016, 99, 5032–5038. [Google Scholar] [CrossRef]
- Jung, Y.; Lee, J.; Kwon, J.; Lee, K.S.; Ryu, D.H.; Hwang, G.S. Discrimination of the geographical origin of beef by (1)H NMR-based metabolomics. J. Agric. Food Chem. 2010, 58, 10458–10466. [Google Scholar] [CrossRef] [PubMed]
- Muroya, S.; Oe, M.; Ojima, K.; Watanabe, A. Metabolomic approach to key metabolites characterizing postmortem aged loin muscle of Japanese Black (Wagyu) cattle. Asian-Australas, J. Anim. Sci. 2019, 32, 1172–1185. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.; Curley, D.; O’Callaghan, T.F.; O’Shea, C.; Dempsey, E.M.; O’Toole, P.W.; Ross, R.P.; Ryan, C.A.; Stanton, C. The composition of human milk and infant faecal microbiota over the first three months of life: A pilot study. Sci. Rep. 2017, 7, 40597. [Google Scholar] [CrossRef] [PubMed]
- Rozenberg, S.; Body, J.; Bruyère, O.; Bergmann, P.; Brandi, M.L.; Cooper, C.; Devogelaer, J.; Gielen, E.; Goemaere, S.; Kaufman, J.; et al. Effects of dairy products consumption on health: Benefits and beliefs--a commentary from the Belgian bone club and the European society for clinical and economic aspects of osteoporosis, osteoarthritis and musculoskeletal diseases. Calcif. Tissue Int. 2016, 98, 1–17. [Google Scholar] [CrossRef] [PubMed]
- McCluskey, J.J.; Wahl, T.I.; Li, Q.; Wandschneider, P.R.U.S. grass-fed beef: Marketing health benefits. J. Food Distrib. Res. 2005, 36, 1. [Google Scholar]
- Saleem, F.; Bouatra, S.; Guo, A.C.; Psychogios, N.; Mandal, R.; Dunn, S.M.; Ametaj, B.N.; Wishart, D.S. The bovine ruminal fluid metabolome. Metabolomics. 2013, 9, 360–378. [Google Scholar] [CrossRef]
- Foroutan, A.; Guo, A.C.; Vazquez-Fresno, R.; Lipfert, M.; Zhang, L.; Zheng, J.; Badran, H.; Budinski, Z.; Mandal, R.; Ametaj, B.N.; et al. Chemical composition of commercial cow’s milk. J. Agric. Food Chem. 2019, 67, 4897–4914. [Google Scholar] [CrossRef] [PubMed]
- Dervishi, E.; Zhang, G.; Dunn, S.M.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. GC–MS metabolomics identifies metabolite alterations that precede subclinical mastitis in the blood of transition dairy cows. J. Proteome Res. 2017, 16, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Dervishi, E.; Zhang, G.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. Targeted metabolomics: New insights into pathobiology of retained placenta in dairy cows and potential risk biomarkers. Animal 2018, 12, 1050–1059. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Deng, Q.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. DI/LC-MS/MS-based metabolic profiling for identification of early predictive serum biomarkers of metritis in transition dairy cows. J. Agric. Food Chem. 2017, 65, 8510–8521. [Google Scholar] [CrossRef] [PubMed]
- Dervishi, E.; Zhang, G.; Hailemariam, D.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. Urine metabolic fingerprinting can be used to predict the risk of metritis and highlight the pathobiology of the disease in dairy cows. Metabolomics 2018, 14, 83. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Jewison, T.; Guo, A.C.; Wilson, M.; Knox, C.; Liu, Y.; Djoumbou, Y.; Mandal, R.; Aziat, F.; Dong, E.; et al. HMDB 3.0-the human metabolome database in 2013. Nucleic Acids Res. 2013, 41, 801. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2017, 46, D608–D617. [Google Scholar] [CrossRef]
- Wishart, D.S.; Knox, C.; Guo, A.C.; Eisner, R.; Young, N.; Gautam, B.; Hau, D.D.; Psychogios, N.; Dong, E.; Bouatra, S.; et al. HMDB: A knowledgebase for the human metabolome. Nucleic Acids Res. 2009, 37, 603. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Tzur, D.; Knox, C.; Eisner, R.; Guo, A.C.; Young, N.; Cheng, D.; Jewell, K.; Arndt, D.; Sawhney, S.; et al. HMDB: The human metabolome database. Nucleic Acids Res. 2007, 35, 521. [Google Scholar] [CrossRef]
- Ramirez-Gaona, M.; Marcu, A.; Pon, A.; Guo, A.C.; Sajed, T.; Wishart, N.A.; Karu, N.; Djoumbou Feunang, Y.; Arndt, D.; Wishart, D.S. YMDB 2.0: A significantly expanded version of the yeast metabolome database. Nucleic Acids Res. 2017, 45, D440–D445. [Google Scholar] [CrossRef]
- Jewison, T.; Knox, C.; Neveu, V.; Djoumbou, Y.; Guo, A.C.; Lee, J.; Liu, P.; Mandal, R.; Krishnamurthy, R.; Sinelnikov, I.; et al. YMDB: The yeast metabolome database. Nucleic Acids Res. 2012, 40, 815. [Google Scholar] [CrossRef]
- Sajed, T.; Marcu, A.; Ramirez, M.; Pon, A.; Guo, A.C.; Knox, C.; Wilson, M.; Grant, J.R.; Djoumbou, Y.; Wishart, D.S. ECMDB 2.0: A richer resource for understanding the biochemistry of E. coli. Nucleic Acids Res. 2016, 44, 495. [Google Scholar] [CrossRef]
- Guo, A.C.; Jewison, T.; Wilson, M.; Liu, Y.; Knox, C.; Djoumbou, Y.; Lo, P.; Mandal, R.; Krishnamurthy, R.; Wishart, D.S. ECMDB: The, E. coli metabolome database. Nucleic Acids Res. 2013, 41, 625. [Google Scholar] [CrossRef]
- Kind, T.; Scholz, M.; Fiehn, O. How large is the metabolome? A critical analysis of data exchange practices in chemistry. PLoS ONE 2009, 4, e5440. [Google Scholar] [CrossRef] [PubMed]
- Grennan, A.K. MoTo DB: A metabolic database for tomato. Plant. Physiol. 2009, 151, 1701–1702. [Google Scholar] [CrossRef] [PubMed]
- FooDB. Available online: https://foodb.ca/ (accessed on 28 February 2020).
- O’Callaghan, T.F.; Vazquez-Fresno, R.; Serra-Cayuela, A.; Dong, E.; Mandal, R.; Hennessy, D.; McAuliffe, S.; Dillon, P.; Wishart, D.S.; Stanton, C.; et al. Pasture feeding changes the bovine rumen and milk metabolome. Metabolites 2018, 8, 27. [Google Scholar] [CrossRef] [PubMed]
- Noaman, V.; Rasti, M.; Ranjbari, A.R.; Shirvani, E. Copper, zinc, and iron concentrations in blood serum and diet of dairy cattle on semi-industrial farms in central Iran. Trop. Anim. Health Prod. 2012, 44, 407–411. [Google Scholar] [CrossRef]
- Lee, H.J.; Jung, J.Y.; Oh, Y.K.; Lee, S.S.; Madsen, E.L.; Jeon, C.O. Comparative survey of rumen microbial communities and metabolites across one caprine and three bovine groups, using bar-coded pyrosequencing and (1)H nuclear magnetic resonance spectroscopy. Appl. Environ. Microbiol. 2012, 78, 5983–5993. [Google Scholar] [CrossRef] [PubMed]
- Brown-Woodman, P.D.; White, I.G. Amino acid composition of semen and the secretions of the male reproductive tract. Aust. J. Biol. Sci. 1974, 27, 415–422. [Google Scholar] [CrossRef]
- Raun, B.M.; Kristensen, N.B. Metabolic effects of feeding ethanol or propanol to postpartum transition Holstein cows. J. Dairy Sci. 2011, 94, 2566–2580. [Google Scholar] [CrossRef]
- Sato, H. Increased blood concentration of isopropanol in ketotic dairy cows and isopropanol production from acetone in the rumen. Anim. Sci. J. 2009, 80, 381–386. [Google Scholar] [CrossRef]
- Sato, H.; Matsumoto, M.; Hanasaka, S. Relations between plasma acetate, 3-hydroxybutyrate, FFA, glucose levels and energy nutrition in lactating dairy cows. J. Vet. Med. Sci. 1999, 61, 447–451. [Google Scholar] [CrossRef]
- Goldansaz, S.A.; Guo, A.C.; Sajed, T.; Steele, M.A.; Plastow, G.S.; Wishart, D.S. Livestock metabolomics and the livestock metabolome: A systematic review. PLoS ONE 2017, 12, e0177675. [Google Scholar] [CrossRef]
- Zhou, Z.; Vailati-Riboni, M.; Luchini, D.N.; Loor, J.J. Methionine and choline supply during the periparturient period alter plasma amino acid and one-carbon metabolism profiles to various extents: Potential role in hepatic metabolism and antioxidant status. Nutrients 2017, 9, 10. [Google Scholar] [CrossRef] [PubMed]
- Bovine Metabolome Database. Available online: http://www.bovinedb.ca/ (accessed on 28 February 2020).
- Wishart, D.S.; Li, C.; Marcu, A.; Badran, H.; Pon, A.; Budinski, Z.; Patron, J.; Lipton, D.; Cao, X.; Oler, E.; et al. A comprehensive pathway database for model organisms. Nucleic Acids Res. 2020, 48, D470–D478. [Google Scholar] [CrossRef] [PubMed]
- Motyl, T.; Barej, W. Plasma amino acid indices and urinary 3-methyl histidine excretion in dairy cows in early lactation. Ann. Rech. Vet. 1986, 17, 153–157. [Google Scholar] [PubMed]
- Sadri, H.; von Soosten, D.; Meyer, U.; Kluess, J.; Danicke, S.; Saremi, B.; Sauerwein, H. Plasma amino acids and metabolic profiling of dairy cows in response to a bolus duodenal infusion of leucine. PLoS ONE 2017, 12, e0176647. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, R.H.; Titgemeyer, E.C.; Stokka, G.L.; Drouillard, J.S.; Loest, C.A. Effects of L-carnitine on nitrogen retention and blood metabolites of growing steers and performance of finishing steers. J. Anim. Sci. 2001, 79, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.R.; Boger, R.H.; Bode-Boger, S.M.; Tangphao, O.; Tsao, P.S.; Blaschke, T.F.; Cooke, J.P. Asymmetric dimethylarginine increases mononuclear cell adhesiveness in hypercholesterolemic humans. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 1040–1046. [Google Scholar] [CrossRef]
- Consolo, N.R.B.; Munro, J.C.; Bourgon, S.L.; Karrow, N.A.; Fredeen, A.H.; Martell, J.E.; Montanholi, Y.R. Associations of blood analysis with feed efficiency and developmental stage in grass-fed beef heifers. Animals 2018, 8, 133. [Google Scholar] [CrossRef]
- Van der Drift, S.G.A.; Houweling, M.; Schonewille, J.T.; Tielens, A.G.M.; Jorritsma, R. Protein and fat mobilization and associations with serum beta-hydroxybutyrate concentrations in dairy cows. J. Dairy Sci. 2012, 95, 4911–4920. [Google Scholar] [CrossRef]
- Hernandez-Castellano, L.E.; Hernandez, L.L.; Weaver, S.; Bruckmaier, R.M. Increased serum serotonin improves parturient calcium homeostasis in dairy cows. J. Dairy Sci. 2017, 100, 1580–1587. [Google Scholar] [CrossRef]
- Padilla, L.; Matsui, T.; Kamiya, Y.; Kamiya, M.; Tanaka, M.; Yano, H. Heat stress decreases plasma vitamin C concentration in lactating cows. Livest. Sci. 2006, 101, 300–304. [Google Scholar] [CrossRef]
- Kenny, D.A.; Humpherson, P.G.; Leese, H.J.; Morris, D.G.; Tomos, A.D.; Diskin, M.G.; Sreenan, J.M. Effect of elevated systemic concentrations of ammonia and urea on the metabolite and ionic composition of oviductal fluid in cattle. Biol. Reprod. 2002, 66, 1797–1804. [Google Scholar] [CrossRef] [PubMed]
- Artegoitia, V.M.; Middleton, J.L.; Harte, F.M.; Campagna, S.R.; de Veth, M.J. Choline and choline metabolite patterns and associations in blood and milk during lactation in dairy cows. PLoS ONE 2014, 9, e103412. [Google Scholar] [CrossRef] [PubMed]
- Liker, B.; Bačar-Huskić, L.; Knežević, M.; Rupić, V.; Vranešić, N.; Romić, Ž.; Grbesa, D.; Macesic, D.; Leto, J.; Zvonko, A.; et al. Blood metabolites and haematological indices of pregnant beef cows fed rumen-protected methionine. J. Anim. Feed Sci. 2005, 14, 625–638. [Google Scholar] [CrossRef]
- Nozad, S.; Ramin, A.G.; Moghadam, G.; Asri-Rezaei, S.; Babapour, A.; Ramin, S. Relationship between blood urea, protein, creatinine, triglycerides and macro-mineral concentrations with the quality and quantity of milk in dairy Holstein cows. Vet. Res. Forum. 2012, 3, 55–59. [Google Scholar] [PubMed]
- Macdonald, A.G.C.; Bourgon, S.L.; Palme, R.; Miller, S.P.; Montanholi, Y.R. Evaluation of blood metabolites reflects presence or absence of liver abscesses in beef cattle. Vet. Rec. Open 2017, 4, e000170. [Google Scholar] [CrossRef] [PubMed]
- Waldner, C.; Campbell, J.; Jim, G.K.; Guichon, P.T.; Booker, C. Comparison of 3 methods of selenium assessment in cattle. Can. Vet. J. 1998, 39, 225–231. [Google Scholar] [PubMed]
- Sexton, T.J.; Amann, R.P.; Flipse, R.J. Free amino acids and protein in rete testis fluid, vas deferens plasma, accessory sex gland fluid, and seminal plasma of the conscious bull. J. Dairy Sci. 1971, 54, 412–416. [Google Scholar] [CrossRef]
- Velho, A.L.C.; Menezes, E.; Dinh, T.; Kaya, A.; Topper, E.; Moura, A.A.; Memili, E. Metabolomic markers of fertility in bull seminal plasma. PLoS ONE 2018, 13, e0195279. [Google Scholar] [CrossRef]
- Menezes, E.B.; Velho, A.L.C.; Santos, F.; Dinh, T.; Kaya, A.; Topper, E.; Moura, A.A.; Memili, E. Uncovering sperm metabolome to discover biomarkers for bull fertility. BMC Genom. 2019, 20, 714–716. [Google Scholar] [CrossRef]
- Kim, Y.H.; Kemp, R.; Samuelsson, L.M. Effects of dry-aging on meat quality attributes and metabolite profiles of beef loins. Meat Sci. 2016, 111, 168–176. [Google Scholar] [CrossRef]
- Kodani, Y.; Miyakawa, T.; Komatsu, T.; Tanokura, M. NMR-based metabolomics for simultaneously evaluating multiple determinants of primary beef quality in Japanese Black cattle. Sci. Rep. 2017, 7, 1297–1298. [Google Scholar] [CrossRef] [PubMed]
- Miles, E.D.; McBride, B.W.; Jia, Y.; Liao, S.F.; Boling, J.A.; Bridges, P.J.; Matthews, J.C. Glutamine synthetase and alanine transaminase expression are decreased in livers of aged vs. young beef cows and GS can be upregulated by 17beta-estradiol implants. J. Anim. Sci. 2015, 93, 4500–4509. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.Z.; Zhou, M.; Wang, O.; Chen, Y.; Liu, J.X.; Guan, L.L. Multi-omics reveals functional genomic and metabolic mechanisms of milk production and quality in dairy cows. Bioinformatics 2020, 36, 2530–2537. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, K.; Lopreiato, V.; Liang, Y.; Trevisi, E.; Osorio, J.S.; Xu, C.; Loor, J.J. Hepatic metabolomics and transcriptomics to study susceptibility to ketosis in response to prepartal nutritional management. J. Anim. Sci. Biotechnol. 2019, 10, 96. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Lewis, M.J.; Morrissey, J.A.; Flegel, M.D.; Jeroncic, K.; Xiong, Y.; Cheng, D.; Eisner, R.; Gautam, B.; Tzur, D.; et al. The human cerebrospinal fluid metabolome. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2008, 871, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Yin, P.; Lehmann, R.; Xu, G. Effects of pre-analytical processes on blood samples used in metabolomics studies. Anal. Bioanal Chem. 2015, 407, 4879–4892. [Google Scholar] [CrossRef] [PubMed]
- Gogiashvili, M.; Edlund, K.; Gianmoena, K.; Marchan, R.; Brik, A.; Andersson, J.T.; Lambert, J.; Madjar, K.; Hellwig, B.; Rahnenfuhrer, J.; et al. Metabolic profiling of ob/ob mouse fatty liver using HR-MAS (1)H-NMR combined with gene expression analysis reveals alterations in betaine metabolism and the transsulfuration pathway. Anal. Bioanal Chem. 2017, 409, 1591–1606. [Google Scholar] [CrossRef]
- Soga, T.; Baran, R.; Suematsu, M.; Ueno, Y.; Ikeda, S.; Sakurakawa, T.; Kakazu, Y.; Ishikawa, T.; Robert, M.; Nishioka, T.; et al. Differential metabolomics reveals ophthalmic acid as an oxidative stress biomarker indicating hepatic glutathione consumption. J. Biol. Chem. 2006, 281, 16768–16776. [Google Scholar] [CrossRef]
- Soga, T.; Ohashi, Y.; Ueno, Y.; Naraoka, H.; Tomita, M.; Nishioka, T. Quantitative metabolome analysis using capillary electrophoresis mass spectrometry. J. Proteome Res. 2003, 2, 488–494. [Google Scholar] [CrossRef]
- CCAC. Guide to the Care and Use of Experimental Animals; Canadian Council on Animal Care: Ottawa, ON, Canada, 1993. [Google Scholar]
- Johnson, C.; Fitzsimmons, C.; Colazo, M.; Li, C.; Kastelic, J.; Thundathil, J. Impacts of residual feed intake and pre-natal diet on reproductive potential of bulls. Anim. Prod. Sci. 2019, 59, 1827–1836. [Google Scholar] [CrossRef]
- Foroutan, A.; Goldansaz, S.A.; Lipfert, M.; Wishart, D.S. Protocols for NMR analysis in livestock metabolomics. Methods Mol. Biol. 2019, 1996, 311–324. [Google Scholar] [PubMed]
- Psychogios, N.; Hau, D.D.; Peng, J.; Guo, A.C.; Mandal, R.; Bouatra, S.; Sinelnikov, I.; Krishnamurthy, R.; Eisner, R.; Gautam, B.; et al. The human serum metabolome. PLoS ONE 2011, 6, e16957. [Google Scholar] [CrossRef] [PubMed]
- Saude, E.; Slupsky, C.; Sykes, B. Optimization of NMR analysis of biological fluids for quantitative accuracy. Metabolomics 2006, 2, 113–123. [Google Scholar] [CrossRef]
- Ametaj, B.N.; Zebeli, Q.; Saleem, F.; Psychogios, N.; Lewis, M.J.; Dunn, S.M.; Xia, J.; Wishart, D.S. Metabolomics reveals unhealthy alterations in rumen metabolism with increased proportion of cereal grain in the diet of dairy cows. Metabolomics 2010, 6, 583–594. [Google Scholar] [CrossRef]
- Minerbi, A.; Gonzalez, E.; Brereton, N.J.B.; Anjarkouchian, A.; Dewar, K.; Fitzcharles, M.A.; Chevalier, S.; Shir, Y. Altered microbiome composition in individuals with fibromyalgia. Pain 2019, 160, 2589–2602. [Google Scholar] [CrossRef] [PubMed]
- Magan, J.B.; O’Callaghan, T.F.; Zheng, J.; Zhang, L.; Mandal, R.; Hennessy, D.; Fenelon, M.A.; Wishart, D.S.; Kelly, A.L.; McCarthy, N.A. Impact of bovine diet on metabolomic profile of skim milk and whey protein ingredients. Metabolites 2019, 9, 305. [Google Scholar] [CrossRef] [PubMed]
- Google Scholar. Available online: https://scholar.google.ca/ (accessed on 28 February 2020).
- Pubmed. Available online: http://www.ncbi.nlm.nih.gov/pubmed (accessed on 28 February 2020).
- ScienceDirect. Available online: http://www.sciencedirect.com/ (accessed on 28 February 2020).
- Wishart, D.S. Quantitative metabolomics using NMR. Trends Anal. Chem. 2008, 27, 228–237. [Google Scholar] [CrossRef]
- Cheng, D.; Knox, C.; Young, N.; Stothard, P.; Damaraju, S.; Wishart, D.S. PolySearch: A web-based text mining system for extracting relationships between human diseases, genes, mutations, drugs and metabolites. Nucleic Acids Res. 2008, 36, W399–W405. [Google Scholar] [CrossRef]
- Liu, Y.; Liang, Y.; Wishart, D. PolySearch2: A significantly improved text-mining system for discovering associations between human diseases, genes, drugs, metabolites, toxins and more. Nucleic Acids Res. 2015, 43, 535. [Google Scholar] [CrossRef]
- UniProt. Available online: https://www.uniprot.org/ (accessed on 28 February 2020).


{kind=link}
{kind=link}
| Metabolite | Platform | Concentration | Literature Value |
|---|---|---|---|
| WATER-SOLUBLE COMPOUNDS AMINO ACIDS | |||
| Alanine * | LC–MS/MS and NMR | 240 ± 30 | 151–222 a |
| Arginine | LC–MS/MS and NMR | 218 ± 33 | 135–182 a |
| Asparagine * | LC–MS/MS and NMR | 25 ± 4 | 20–33 b |
| Aspartate * | LC–MS/MS and NMR | 24 ± 11 | 14–16 c, 31–36 a |
| Beta-alanine * | NMR | 8 ± 1 | 8–9 d |
| Citrulline * | LC–MS/MS and NMR | 88 ± 15 | 71–84 d |
| Creatine | LC–MS/MS and NMR | 196 ± 28 | |
| Glutamate * | LC–MS/MS and NMR | 92 ± 19 | 35–39 d, 174–198 a |
| Glutamine * | LC–MS/MS and NMR | 330 ± 42 | 246–260 d |
| Glycine * | LC–MS/MS and NMR | 398 ± 65 | 405–428 d |
| Histidine * | LC–MS/MS | 78 ± 10 | 74–84 c |
| Isoleucine | LC–MS/MS and NMR | 153 ± 14 | 101–122 a |
| Leucine * | LC–MS/MS and NMR | 212 ± 24 | 205–264 c |
| Lysine * | LC–MS/MS and NMR | 88 ± 15 | 58–92 b |
| Methionine * | LC–MS/MS and NMR | 34 ± 4 | 22–29 c, 46–52 a |
| Ornithine * | LC–MS/MS and NMR | 61 ± 13 | 62–135 a |
| Phenylalanine * | LC–MS/MS and NMR | 71 ± 7 | 65–75 c |
| Proline * | LC–MS/MS and NMR | 103 ± 15 | 84–110 a |
| Serine * | LC–MS/MS and NMR | 85 ± 14 | 86–89 d |
| Threonine * | LC–MS/MS and NMR | 74 ± 12 | 58–76 c |
| Tryptophan * | LC–MS/MS | 47 ± 7 | 37–42 c |
| Tyrosine | LC–MS/MS and NMR | 91 ± 10 | 68–75 c |
| Valine * | LC–MS/MS and NMR | 356 ± 34 | 262–322 c |
| BIOGENIC AMINES | |||
| Acetyl-ornithine | LC–MS/MS | 3 ± 1 | |
| Asymmetric-dimethylarginine * | LC–MS/MS | 1.1 ± 0.2 | 1.3–2.1 e |
| Carnosine * | LC–MS/MS | 30 ± 12 | 15–20 d |
| Creatinine * | LC–MS/MS and NMR | 113 ± 17 | 109–140 f |
| Kynurenine * | LC–MS/MS | 7 ± 2 | 4–7 b |
| Methionine-sulfoxide | LC–MS/MS | 1.2 ± 0.3 | |
| Methylhistidine * | LC–MS/MS | 14 ± 2 | 2–12 g |
| Putrescine | LC–MS/MS | 0.04 ± 0.02 | |
| Sarcosine | LC–MS/MS and NMR | 3 ± 1 | 10–12 d |
| Serotonin * | LC–MS/MS | 9 ± 3 | 4–13 h |
| Spermidine | LC–MS/MS | 0.2 ± 0.1 | |
| Spermine | LC–MS/MS | 0.2 ± 0.2 | |
| Taurine | LC–MS/MS and NMR | 80 ± 20 | 33–47 d |
| Total-dimethylarginine | LC–MS/MS | 2.1 ± 0.3 | |
| Trans-hydroxyproline | LC–MS/MS | 25 ± 5 | |
| Trimethylamine N-oxide | LC–MS/MS | 6 ± 3 | |
| CARBOHYDRATES | |||
| Glucose * | LC–MS/MS and NMR | 3962 ± 443 | 3290–4070 i |
| ORGANIC ACIDS | |||
| 3-hydroxybutyrate * | NMR | 340 ± 145 | 250–2110 j |
| Acetate | NMR | 403 ± 199 | 920–1040 k |
| Alpha-aminoadipate | LC–MS/MS | 1.28 ± 0.52 | 7.3–8.1 d |
| Ascorbate (Vitamin C) * | NMR | 11 ± 3 | 8–18 l |
| Formate | NMR | 78 ± 12 | |
| Fumarate | NMR | 1.2 ± 0.2 | |
| Lactate | NMR | 4850 ± 2017 | 658–1600 m |
| Pyruvate | NMR | 150 ± 40 | |
| MISCELANEOUS | |||
| Acetone * | NMR | 70 ± 22 | 80–990 j |
| Betaine | LC–MS/MS and NMR | 169 ± 31 | 14–26 n |
| Choline | LC–MS/MS and NMR | 20 ± 4 | 4–5 n |
| Ethanol * | NMR | 8 ± 1 | 3–68 i |
| Glycerol | NMR | 314 ± 38 | |
| Isopropanol | NMR | 2 ± 1 | |
| Methanol | NMR | 32 ± 4 | |
| Myo-inositol | NMR | 45 ± 11 | |
| Urea | NMR | 1321 ± 282 | 1950–4080 o |
| Uridine | NMR | 3 ± 1 | |
| LIPID-LIKE COMPOUNDS PHOSPHATIDYLCHOLINES, ACYL-ALKYL | |||
| PC ae (36:0) | LC–MS/MS | 1.6 ± 0.4 | |
| PC ae (40:6) | LC–MS/MS | 0.46 ± 0.11 | |
| PHOSPHATIDYLCHOLINES, DIACYL | |||
| PC aa (32:2) | LC–MS/MS | 4 ± 1 | |
| PC aa (36:6) | LC–MS/MS | 0.7 ± 0.2 | |
| PC aa (36:0) | LC–MS/MS | 6 ± 2 | |
| PC aa (38:6) | LC–MS/MS | 1 ± 0.3 | |
| PC aa (38:0) | LC–MS/MS | 0.8 ± 0.2 | |
| PC aa (40:6) | LC–MS/MS | 1.6 ± 0.4 | |
| PC aa (40:2) | LC–MS/MS | 0.4 ± 0.1 | |
| PC aa (40:1) | LC–MS/MS | 0.21 ± 0.04 | |
| LYSOPHOSPHATIDYLCHOLINES, ACYL C | |||
| LysoPC(14:0) | LC–MS/MS | 0.8 ± 0.1 | |
| LysoPC(16:1) | LC–MS/MS | 0.6 ± 0.1 | |
| LysoPC(16:0) * | LC–MS/MS | 20 ± 4 | 15–58 n |
| LysoPC(17:0) | LC–MS/MS | 3 ± 1 | |
| LysoPC(18:2) | LC–MS/MS | 15 ± 3 | 30–186 n |
| LysoPC(18:1) | LC–MS/MS | 6 ± 1 | 18–69 n |
| LysoPC(18:0) * | LC–MS/MS | 30 ± 5 | 14–82 n |
| LysoPC(20:4) | LC–MS/MS | 0.48 ± 0.14 | |
| LysoPC(20:3) | LC–MS/MS | 1.6 ± 0.3 | |
| LysoPC(24:0) | LC–MS/MS | 0.051 ± 0.012 | |
| LysoPC(26:1) | LC–MS/MS | 0.1 ± 0.04 | |
| LysoPC(26:0) | LC–MS/MS | 0.7 ± 0.3 | |
| LysoPC(28:1) | LC–MS/MS | 0.3 ± 0.1 | |
| LysoPC(28:0) | LC–MS/MS | 0.28 ± 0.11 | |
| SPHINGOMYELINS | |||
| SM(16:1) | LC–MS/MS | 5 ± 1 | |
| SM(16:0) | LC–MS/MS | 68 ± 10 | |
| SM(18:1) | LC–MS/MS | 11 ± 3 | |
| SM(18:0) | LC–MS/MS | 12 ± 2 | |
| SM(20:2) | LC–MS/MS | 1.1 ± 0.3 | |
| HYDROXYSPHINGOMYELINS | |||
| SM(14:1(OH)) | LC–MS/MS | 5 ± 1 | |
| SM(16:1(OH)) | LC–MS/MS | 9 ± 2 | |
| SM(22:2(OH)) | LC–MS/MS | 4 ± 1 | |
| SM(22:1(OH)) | LC–MS/MS | 9 ± 1 | |
| SM(24:1(OH)) | LC–MS/MS | 2 ± 0.4 | |
| ACYLCARNITINES | |||
| C0 (Carnitine) | LC–MS/MS | 7 ± 1 | |
| C2 (Acetylcarnitine) * | LC–MS/MS | 2 ± 1 | 0.65–1.09 b |
| C3:1 (Propenoylcarnitine) | LC–MS/MS | 0.029 ± 0.004 | |
| C3 (Propionylcarnitine) | LC–MS/MS | 0.2 ± 0.04 | |
| C4:1 (Butenylcarnitine) | LC–MS/MS | 0.017 ± 0.002 | |
| C4 (Butyrylcarnitine) | LC–MS/MS | 0.2 ± 0.1 | |
| C3-OH (Hydroxypropionylcarnitine) * | LC–MS/MS | 0.027 ± 0.004 | 0.01–0.02 b |
| C5:1 (Tiglylcarnitine) | LC–MS/MS | 0.023 ± 0.004 | |
| C5 (Valerylcarnitine) * | LC–MS/MS | 0.09 ± 0.03 | 0.03–0.06 b |
| C4-OH (C3-DC) (Hydroxybutyrylcarnitine) | LC–MS/MS | 0.04 ± 0.01 | |
| C6:1 (Hexenoylcarnitine) | LC–MS/MS | 0.02 ± 0.01 | |
| C6 (C4:1-DC) (Hexanoylcarnitine) | LC–MS/MS | 0.05 ± 0.01 | 0.02–0.03 b |
| C5-OH (C3-DC-M) (hydroxyvalerylcarnitine) * | LC–MS/MS | 0.04 ± 0.01 | 0.05–0.06 b |
| C5:1-DC (Glutaconylcarnitine) | LC–MS/MS | 0.018 ± 0.003 | |
| C5-DC (C6-OH)(Glutarylcarnitine) | LC–MS/MS | 0.03 ± 0.01 | |
| C8 (Octanoylcarnitine) | LC–MS/MS | 0.02 ± 0.01 | |
| C5-M-DC (methylglutarylcarnitine) | LC–MS/MS | 0.0196 ± 0.0024 | |
| C9 (Nonaylcarnitine) | LC–MS/MS | 0.022 ± 0.003 | |
| C7-DC (Pimelylcarnitine) * | LC–MS/MS | 0.04 ± 0.04 | 0.01–0.02 b |
| C10:2 (Decadienylcarnitine) | LC–MS/MS | 0.06 ± 0.01 | |
| C10:1 (Decenoylcarnitine) | LC–MS/MS | 0.17 ± 0.03 | |
| C10 (Decanoylcarnitine) | LC–MS/MS | 0.18 ± 0.04 | |
| C12:1 (Dodecenoylcarnitine) | LC–MS/MS | 0.084 ± 0.014 | |
| C12 (Dodecanoylcarnitine) * | LC–MS/MS | 0.04 ± 0.01 | 0.02–0.03 b |
| C14:2 (Tetradecadienylcarnitine) | LC–MS/MS | 0.03 ± 0.01 | |
| C14:1 (Tetradecenoylcarnitine) | LC–MS/MS | 0.0518 ± 0.0103 | |
| C14 (Tetradecanoylcarnitine) * | LC–MS/MS | 0.02 ± 0.01 | 0.01–0.02 b |
| C12-DC (Dodecanedioylcarnitine) | LC–MS/MS | 0.018 ± 0.003 | |
| C14:2-OH (Hydroxytetradecadienylcarnitine) | LC–MS/MS | 0.008 ± 0.002 | |
| C14:1-OH (Hydroxytetradecenoylcarnitine) | LC–MS/MS | 0.009 ± 0.002 | |
| C16:2 (Hexadecadienylcarnitine) | LC–MS/MS | 0.012 ± 0.002 | |
| C16:1 (Hexadecenoylcarnitine) | LC–MS/MS | 0.029 ± 0.003 | |
| C16 (Hexadecanoylcarnitine) | LC–MS/MS | 0.02 ± 0.01 | |
| C16:2-OH (Hydroxyhexadecadienylcarnitine) | LC–MS/MS | 0.005 ± 0.001 | |
| C16:1-OH (Hydroxyhexadecenoylcarnitine) | LC–MS/MS | 0.0184 ± 0.0034 | |
| C16-OH (Hydroxyhexadecanoylcarnitine) * | LC–MS/MS | 0.008 ± 0.001 | 0.003–0.006 b |
| C18:2 (Octadecadienylcarnitine) | LC–MS/MS | 0.007 ± 0.001 | |
| C18:1 (Octadecenoylcarnitine) | LC–MS/MS | 0.0147 ± 0.0031 | |
| C18 (Octadecanoylcarnitine) | LC–MS/MS | 0.021 ± 0.008 | |
| C18:1-OH (Hydroxyoctadecenoylcarnitine) * | LC–MS/MS | 0.009 ± 0.001 | 0.008–0.009 b |
| TRACE ELEMENTAL COMPOUNDS | |||
| Sodium * | ICP–MS | 133,515 ± 13,658 | 107,400–108,600 p, 136,000–136,710 q |
| Magnesium * | ICP–MS | 931 ± 88 | 850–920 f |
| Phosphorus * | ICP–MS | 1298 ± 164 | 1350–1620 p |
| Potassium * | ICP–MS | 4296 ± 388 | 4060–4340 f |
| Calcium * | ICP–MS | 2228 ± 221 | 1400–2200 h |
| Iron * | ICP–MS | 52 ± 13 | 50–51 r |
| Copper * | ICP–MS | 9 ± 2 | 6–9 r |
| Zinc * | ICP–MS | 12 ± 2 | 14–18 r |
| Selenium * | ICP–MS | 1.4 ± 0.2 | 0.5–2.7 s |
| Rubidium | ICP–MS | 1.8 ± 0.2 | |
| Strontium | ICP–MS | 1 ± 0.1 | |
| Cesium | ICP–MS | 0.0017 ± 0.0003 | |
| Barium | ICP–MS | 0.2 ± 0.03 | |
| Tissue/Biofluid Location | Identified Metabolites or Metabolite Species | Identified Metabolites with Unique Structures | Quantified Metabolites with Unique Structures |
|---|---|---|---|
| BIOFLUID | |||
| Blood | 330 | 453 | 296 |
| Colostrum | 70 | 70 | 4 |
| Milk | 928 | 2350 | 1652 |
| Ruminal fluid | 328 | 769 | 728 |
| Semen | 76 | 76 | 0 |
| Urine | 62 | 62 | 0 |
| TISSUE | |||
| Adipose tissue | 199 | 199 | 71 |
| Brain | 557 | 1887 | 0 |
| Epidermis | 275 | 275 | 0 |
| Fibroblasts | 327 | 327 | 0 |
| Intestine | 253 | 253 | 0 |
| Kidney | 531 | 615 | 0 |
| Liver | 1056 | 1254 | 273 |
| Longissimus thoracis muscle | 153 | 267 | 267 |
| Mammary gland | 269 | 269 | 0 |
| Neuron | 322 | 322 | 0 |
| Pancreas | 114 | 114 | 0 |
| Placenta | 579 | 586 | 0 |
| Platelet | 204 | 204 | 0 |
| Prostate | 268 | 268 | 0 |
| Semimembranosus muscle | 153 | 267 | 267 |
| Skeletal muscle | 382 | 496 | 274 |
| Spleen | 168 | 168 | 0 |
| Testis | 328 | 442 | 277 |
| All tissues | 857 | 4464 | N/A * |
| Compound Name | NMR | LC–MS/MS | Average Difference (%) |
|---|---|---|---|
| Alanine | 252 ± 31 | 240 ± 30 | 4 |
| Arginine | 210 ± 28 | 218 ± 33 | 3 |
| Asparagine | 26 ± 5 | 25 ± 4 | 3 |
| Aspartate | Glucose overlap | 24 ± 11 | |
| Betaine | 180 ± 37 | 169 ± 31 | 6 |
| Choline | 19 ± 4 | 20 ± 4 | 5 |
| Citrulline | 89 ± 16 | 88 ± 15 | 1 |
| Creatine | 210 ± 30 | 196 ± 28 | 6 |
| Creatinine | 121 ± 17 | 113 ± 17 | 6 |
| Glucose | 4572 ± 588 | 3962 ± 443 | 14 |
| Glutamate | Proline overlap | 92 ± 19 | |
| Glutamine | 360 ± 47 | 330 ± 42 | 8 |
| Glycine | 438 ± 71 | 398 ± 65 | 9 |
| Isoleucine | 160 ± 22 | 153 ± 14 | 4 |
| Leucine | 216 ± 30 | 212 ± 24 | 1 |
| Lysine | Arginine overlap | 88 ± 15 | |
| Methionine | 35 ± 5 | 34 ± 4 | 2 |
| Ornithine | 67 ± 11 | 61 ± 13 | 9 |
| Phenylalanine | 67 ± 9 | 71 ± 7 | 5 |
| Proline | 94 ± 16 | 103 ± 15 | 9 |
| Sarcosine | 4 ± 1 | 3 ± 1 | 28 |
| Serine | 87 ± 18 | 85 ± 14 | 2 |
| Taurine | Glucose overlap | 80 ± 20 | |
| Threonine | 72 ± 11 | 74 ± 12 | 2 |
| Tyrosine | 84 ± 11 | 91 ± 10 | 8 |
| Valine | 390 ± 48 | 356 ± 34 | 9 |
© Her Majesty the Queen in Right of Canda as represented by the Minister of Agriculture and Agri-Food, 2020 and © author Carolyn Fitzsimmons, 2020. Submitted for possible open access publication under the terms and conditions of the Creative Commons Attribution(CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Foroutan, A.; Fitzsimmons, C.; Mandal, R.; Piri-Moghadam, H.; Zheng, J.; Guo, A.; Li, C.; Guan, L.L.; Wishart, D.S. The Bovine Metabolome. Metabolites 2020, 10, 233. https://doi.org/10.3390/metabo10060233
Foroutan A, Fitzsimmons C, Mandal R, Piri-Moghadam H, Zheng J, Guo A, Li C, Guan LL, Wishart DS. The Bovine Metabolome. Metabolites. 2020; 10(6):233. https://doi.org/10.3390/metabo10060233
Chicago/Turabian StyleForoutan, Aidin, Carolyn Fitzsimmons, Rupasri Mandal, Hamed Piri-Moghadam, Jiamin Zheng, AnChi Guo, Carin Li, Le Luo Guan, and David S. Wishart. 2020. "The Bovine Metabolome" Metabolites 10, no. 6: 233. https://doi.org/10.3390/metabo10060233
APA StyleForoutan, A., Fitzsimmons, C., Mandal, R., Piri-Moghadam, H., Zheng, J., Guo, A., Li, C., Guan, L. L., & Wishart, D. S. (2020). The Bovine Metabolome. Metabolites, 10(6), 233. https://doi.org/10.3390/metabo10060233