The MAPK Signaling Pathways as a Novel Way in Regulation and Treatment of Parasitic Diseases

Abstract
:1. Introduction
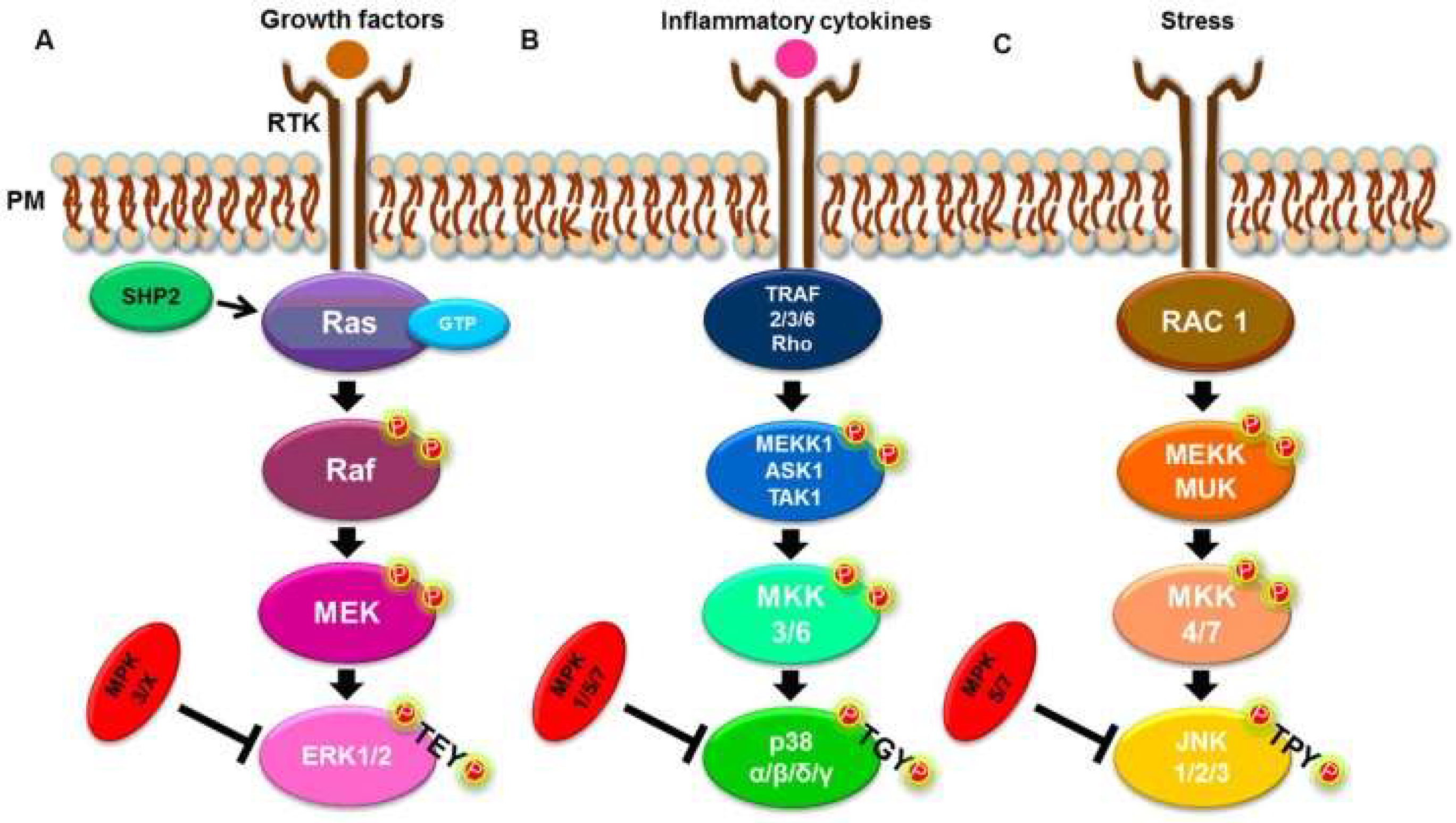
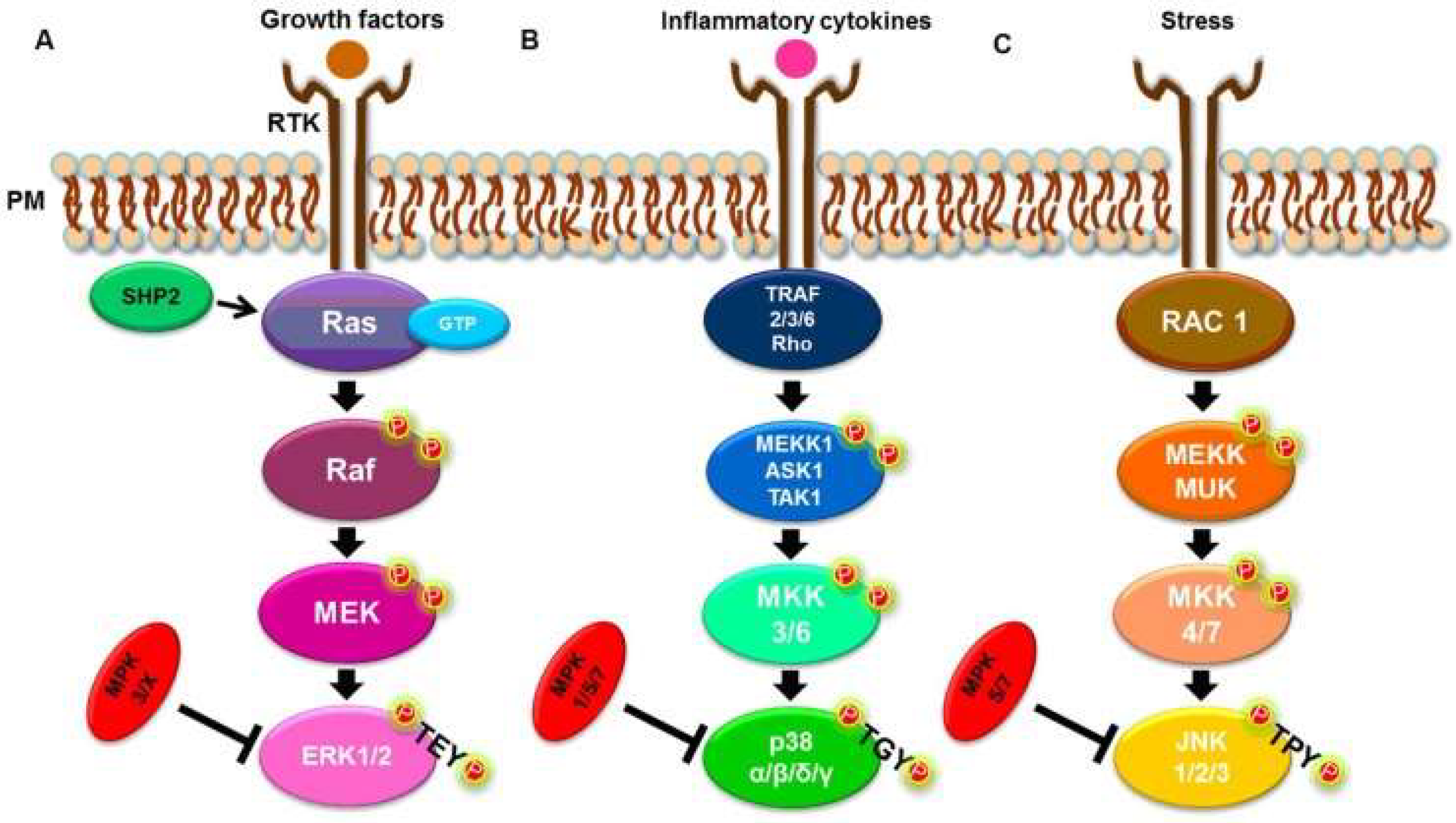
2. Research Directions and MAPK Mechanism of Action in Parasitic Diseases
2.1. The MAPK Interplay Between a Parasite and Host
2.2. Detection and Prevention of Parasites at MAPK Level
2.3. Treatment of MAPK-Dependent Parasitic Diseases
3. Conclusions
Funding
Conflicts of Interest
References
- Lu, Y.; Huang, Q.; Zhou, Z.; Lu, Y. An overview of parasitic diseases by analyzing publications at National Institute of Parasitic Diseases Control and Prevention in the last five years. Chin. J. Dis. Control. Prev. 2015, 19, 86–90. [Google Scholar]
- Zhu, H. Research Progress on Opportunistic Parasitic Infections. Chin. J. Parasitol. Parasit. Dis. 2015, 33, 450–455. [Google Scholar]
- Roy, C.R.; Mocarski, E.S. Pathogen subversion of cell-intrinsic innate immunity. Nat. Immunol. 2007, 8, 1179–1187. [Google Scholar] [CrossRef]
- Han, Z.G.; Brindley, P.J.; Wang, S.Y.; Chen, Z. Schistosoma genomics: New perspectives on schistosome biology and host-parasite interaction. Annu. Rev. Genomics Hum. Genet. 2009, 10, 211–240. [Google Scholar] [CrossRef] [PubMed]
- Brehm, K.; Kozio, U. Echinococcuse-Host Interactions at Cellular and Molecular Levels. Adv. Parasit. 2017, 95, 147–212. [Google Scholar] [CrossRef]
- Soares-Silva, M.; Diniz, F.F.; Gomes, G.N.; Bahia, D. The Mitogen-Activated Protein Kinase (MAPK) Pathway: Role in Immune Evasion by Trypanosomatids. Front. Microbiol. 2016, 7, 183. [Google Scholar] [CrossRef] [PubMed]
- Droguett, D.; Carrillo, I.; Castillo, C.; Gómez, F.; Negrete, M.; Liempi, A.; Muñoz, L.; Galanti, N.; Maya, J.D.; Kemmerling, U. Trypanosoma cruzi induces cellular proliferation in the trophoblastic cell line BeWo. Exp. Parasitol. 2017, 173, 9–17. [Google Scholar] [CrossRef]
- Guilherme, R.R.; Monteiro dos, S.; Marcio, F.; Felipe, A.; Pedro, L.O. Functional studies of TcRjl, a novel GTPase of Trypanosoma cruzi, reveals phenotypes related with MAPK activation during parasite differentiation and after heterologous expression in Drosophila model system. Biochem. Biophys. Res. Commun. 2015, 467, 115–120. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, J.; Lv, G.; Li, J.; Lu, X.; Mantion, G.; Vuitton, D.A.; Wen, H.; Lin, R. Hepatocyte proliferation/growth arrest balance in the liver of mice during E. multilocularis infection: A coordinated 3-stage course. PLoS ONE 2012, 7, 1–17. [Google Scholar] [CrossRef]
- Song, X. Construction and Biological Characteristics of MAPK1-Deficient Toxoplasma gondii. Master’s Thesis, Jilin Agricultural University, Changchun, China, March 2013. [Google Scholar]
- Chauhan, I.S.; SubbaRao, G.; Shankar, J.; Chauhan, L.K.S.; Kapadia, G.J.; Singh, N. Chemoprevention of Leishmaniasis: In vitro antiparasitic activity of dibenzalacetone, a synthetic curcumin analog leads to apoptotic cell death in Leishmania donovani. Parasitol. Int. 2018, 67, 627–636. [Google Scholar] [CrossRef]
- Zhou, L.; Shi, M.; Zhao, L.; Lin, Z.; Tang, Z.; Sun, H.; Chen, T.; Lv, Z.; Xu, J.; Huang, Y.; et al. Clonorchis sinensis lysophospholipase aupregulates IL-25 expression in macrophages as a potential pathway to liver fibrosis. Parasit. Vectors 2017, 10, 295. [Google Scholar] [CrossRef] [PubMed]
- Hassan, G.S.; Mukherjee, S.; Nagajyothi, F.; Louis, M.W.; Stefka, B.P.; Cecilia, J.; Huang, H.; Mahalia, S.D.; Boumediene, B.; Richard, G.P.; et al. Trypanosoma cruzi infection induces proliferation of vascular smooth muscle cells. Infect. Immun. 2006, 74, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Bouzahzah, B.; Nagajyothi, F.; Desruisseaux, M.S.; Mohan, K.; Stephen, M.F.; Alex, W.C.; Michael, P.L.; Stefka, B.P.; Richard, G.P.; Murray, W.; et al. Cell cycle regulatory proteins in the liver in murine Trypanosoma cruzi infection. Cell Cycle 2006, 5, 2396–2400. [Google Scholar] [CrossRef] [PubMed]
- DeSouza-Vieira, T.; Guimarães-Costa, A.; Rochael, N.C.; Lira, M.N.; Nascimento, M.T.; Lima-Gomez, P.S.; Mariante, R.M.; Persechini, P.M.; Saraiva, E.M. Neutrophil extracellular traps release induced by Leishmania: Role of PI3Kγ, ERK, PI3Kσ, PKC, and [Ca2+]. Leukoc Biol. 2016, 100, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, E.; Palencia, A.; Braun, L.; Kapp, U.; Bougdour, A.; Belrhali, H.; Bowler, M.W.; Hakimi, M.A. Structural Basis for the Subversion of MAP Kinase Signaling by an Intrinsically Disordered Parasite Secreted Agonist. Structure 2016, 25, 16–26. [Google Scholar] [CrossRef]
- Quan, J.H.; Chu, J.Q.; Kwon, J.; In-Wook, C.; HassanAhmedHassan, A.I.; Wei, Z.; Guang-Ho, C.; Yu, Z.; Jae-Min, Y.; Eun-Kyeong, J.; et al. Intracellular Networks of the PI3K/AKT and MAPK Pathways for Regulating Toxoplasma gondii-Induced IL-23 and IL-12 Production in Human THP-1 Cells. PLoS ONE 2015, 10, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Kong, X. Relationship between MAPK and parasites. Chin. J. Tradit. Vet. Sci. 2015, 9, 46. [Google Scholar]
- Gong, S.; Zhao, Q.; Liu, Q. Cloning and Prokaryotic Expression of MAPK1 Gene of Toxoplasma gondii. J. Jilin Agric. Univ. 2015, 37, 221–225, 236. [Google Scholar]
- Lucia, P.; Feng, C.; Omar, S.H.; Paul, H.D.; Daniel, P.B.; Catie, S.B.; Dinkorma, O.; David, S.R. Intergrative Genomic Approaches Highlight a Family of Parasite-Specific Kinases that Regulate Host Responses. Cell Host Microbe 2010, 8, 208–218. [Google Scholar] [CrossRef]
- Guo, L.; Zhao, X.; Zhang, J. Advances in the study of rhoptry proteins of Toxoplasma gondii and nucleic acid vaccines against toxoplasmosis. J. Pathog. Biol. 2015, 10, 1–4. [Google Scholar]
- Stuart, W.; Juliane, S.; Juliane, S.; Robin, P. MAP Kinase Phosphatase-2 Plays a Key Role in the Control of Infection with Toxoplasma gondii by Modulating iNOS and Arginase-1 Activities in Mice. PLoS Pathog. 2013, 9, 1–14. [Google Scholar] [CrossRef]
- Xu, P.; Cao, L.; Li, X.; Zeng, D.W.; Quan, Z.; Quan, L. Bioinformatic analysis of the structure and function of MAPK protein encoded by Toxoplasma gondii. Heilongjiang Anim. Sci. Vet. Med. 2015, 17, 1–4, 291. [Google Scholar]
- Sun, X. Proliferation of Toxoplasma gondii tachyzoites and its expression of mitogen activated protein kinases in host cells. J. Bengbu Med. Coll. 2004, 4, 283–284, 294. [Google Scholar]
- Sun, X. The blocking effect of mitogen activated protein kinases inhibitor to the cellular signaling pathway on Toxoplasma gondii tachyzoites invading to host cells. J. Trop. Dis. Parasitol. 2004, 2, 14–15, 28, 66. [Google Scholar]
- Yang, X.; Chen, X.; Sun, X.; Xia, H.; Fang, Q.; Hu, S.; Xu, S.; Wang, Y. The influence of mitogen activated protein kinases inhibitor on the Toxoplasma gondii tachyzoites invading to host cells. J. Trop. Dis. Parasitol. 2006, 4, 5–7, 30. [Google Scholar]
- Yang, X.; Chen, X.; Xu, J.; Sun, X.; Chang, X.; Wang, Y.; Hu, S.; Ge, X.; Li, B. Two mitogen activated protein kinases inhibitors on the Toxoplasma gondii tachyzoites invading to host cells. Chin. J. Zoonoses 2009, 25, 97–99. [Google Scholar]
- Wang, H. Cloning, Expression and Characterization of MAPK Genes from Echinococcus granulosus. Master’s Thesis, Xinjiang University, Urumchi, China, May 2010. [Google Scholar]
- Lv, H. The role of mitogen-activated protein kinase on the proliferation and apoptosis of Echinococcus granulosus. Ph.D. Thesis, Huazhong University of Science and Technology, Wuhan, China, October 2014. [Google Scholar]
- Zhe, C.; Fan, L.; Xiu, L.; Mengya, D.; Jianjian, W.; Xinrui, G.; Huimin, T.; Zhijie, H.; Ying, L.; Xiaoli, C.; et al. EGF-mediated EGFR/ERK signaling pathway promotes germinative cell proliferation in Echinococcus multilocularis that contributes to larval growth and development. PLoS Negl. Trop. Dis. 2017, 11, 1–18. [Google Scholar] [CrossRef]

{kind=link}
| Host Cell Infected by Parasite | Pathological Importance | The Role of the Pathway | Reference |
|---|---|---|---|
| Mice liver cells infected by E. multilocularis | Hepatocyte apoptosis after proliferation as well as damage of liver structure and function. | Early infection: Activation of ERK1/2 contribute to hypatocyte proliferation. Terminal infection: Activation of JNK contribute to hypatocyte apoptosis. | [9] |
| Mouse macrophage cell line (RAW264.7) cultured and treated with CsLysoPLA | Hepatic fibrosis during C. sinensis infection as well as damage of liver structure and function. | PKA-dependent B-Raf/ERK1/2 pathway in mouse macrophages was activated by CsLysoPLA→upregulated production of IL-25→activates HSCs. | [12] |
| BeWo trophoblastic cell line infected by T. cruzi | Cause trophoblast epithelium turnover, which is considered as part of innate immunity. | Participate Bevo trophoblast maintenance and differentiation. | [7] |
| Neutrophil infected by L. amazonensis | Neutrophils netosis can kill parasite. | L. amazonensis induce neutrophils netosis through ERK pathway, which acti-vation downstream of PI3Kg and upstream of ROS generation. | [15] |
| Macrophages infected by Leishmania spp. and T. cruzi | Highjack the immune response, and, in this manner, promote parasite maintenance in the host. | Parasite target on macrophages MAPK pathways to modulate host immune system and to favor it replication and survival. | [16] |
| Human THP-1 Cells infected by T. gondii | IL-23 and IL-12 regulate both innate and adaptive immunity. | IL-23 production in T. gondii-infected THP-1 cells was regulated by ERK1/2. IL-12 production in T. gondii-infected THP-1 cells was regulated by p38 MAPK and JNK. | [17] |
| Parasite | The Role of the Pathway | Reference |
|---|---|---|
| T. gondii | Play an important role in tachyzoite invasion and proliferation. | [10] |
| E. multilocularis | Promote growth of metacestodes and germinative cell proliferation. | [30] |
| L. donovani | Leads to apoptotic cell death in parasite. | [11] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Gui, W.; Niu, F.; Chong, S. The MAPK Signaling Pathways as a Novel Way in Regulation and Treatment of Parasitic Diseases. Diseases 2019, 7, 9. https://doi.org/10.3390/diseases7010009
Zhao Y, Gui W, Niu F, Chong S. The MAPK Signaling Pathways as a Novel Way in Regulation and Treatment of Parasitic Diseases. Diseases. 2019; 7(1):9. https://doi.org/10.3390/diseases7010009
Chicago/Turabian StyleZhao, Yumin, Weifeng Gui, Fuqiu Niu, and Shigui Chong. 2019. "The MAPK Signaling Pathways as a Novel Way in Regulation and Treatment of Parasitic Diseases" Diseases 7, no. 1: 9. https://doi.org/10.3390/diseases7010009
APA StyleZhao, Y., Gui, W., Niu, F., & Chong, S. (2019). The MAPK Signaling Pathways as a Novel Way in Regulation and Treatment of Parasitic Diseases. Diseases, 7(1), 9. https://doi.org/10.3390/diseases7010009