
Mechanism of Action of Topical Tranexamic Acid in the Treatment of Melasma and Sun-Induced Skin Hyperpigmentation


Abstract
1. Introduction
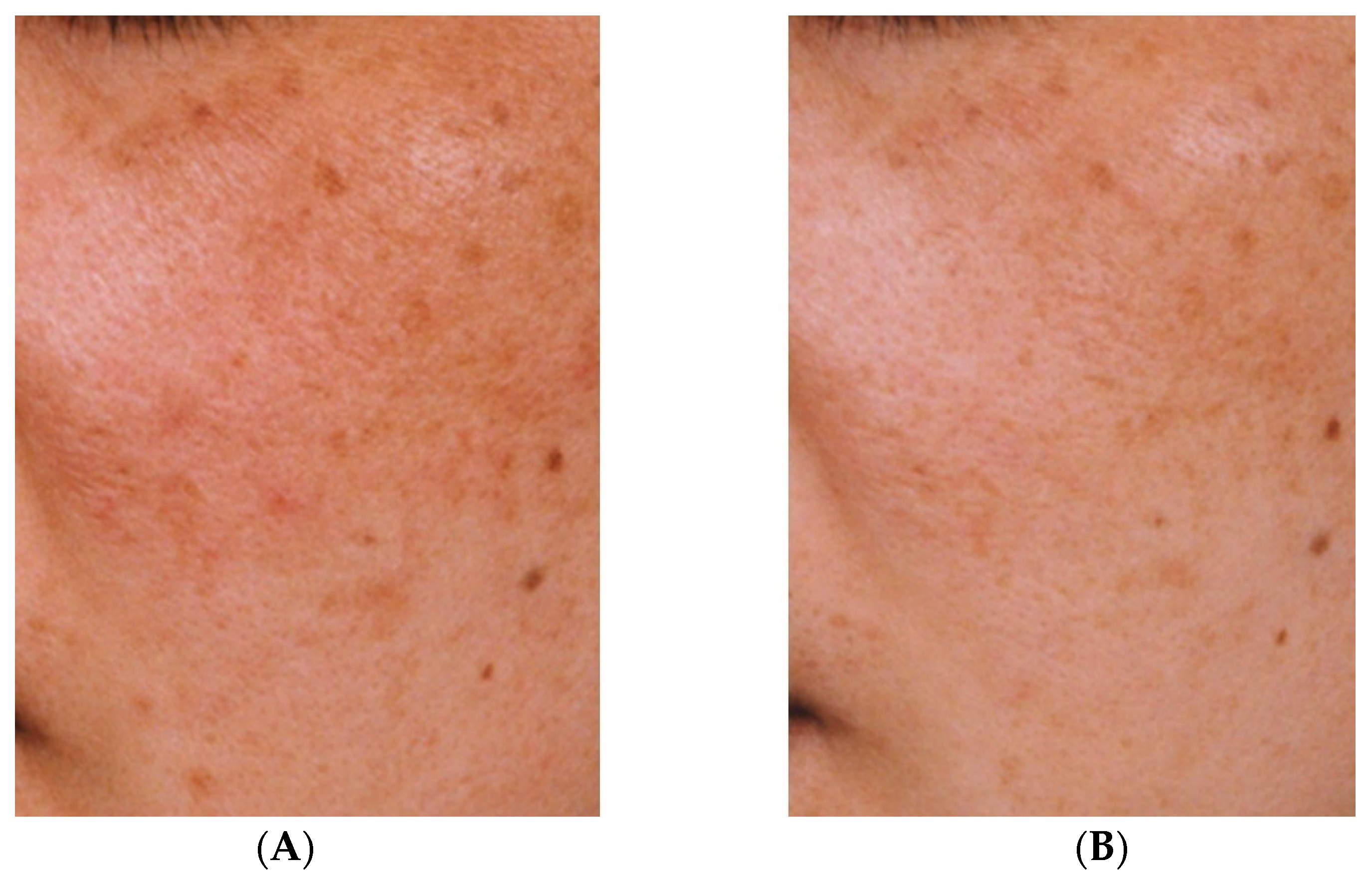
2. Effectiveness of TXA for Melasma
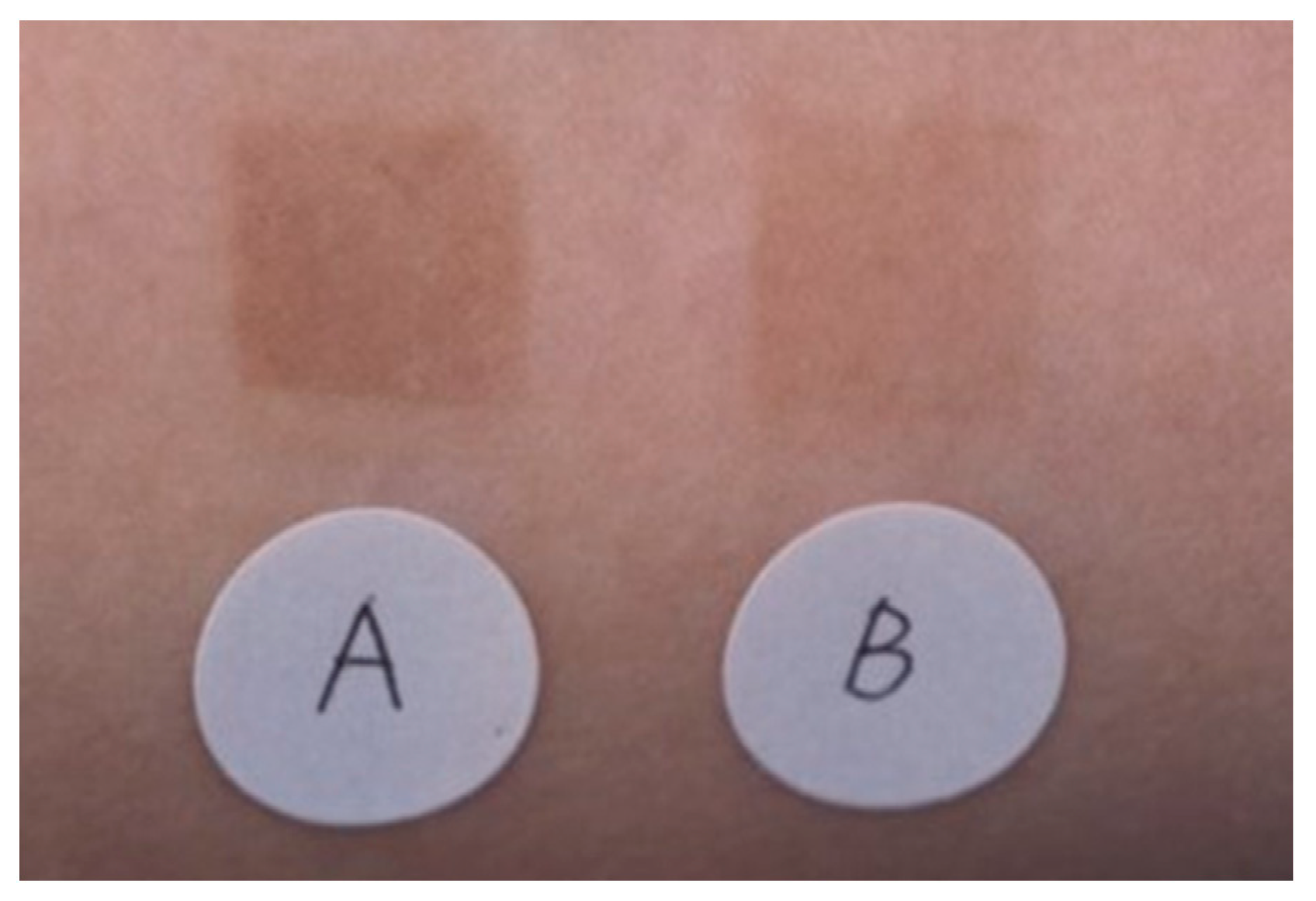
3. Effectiveness of TXA for Sun-Induced Hyperpigmentation
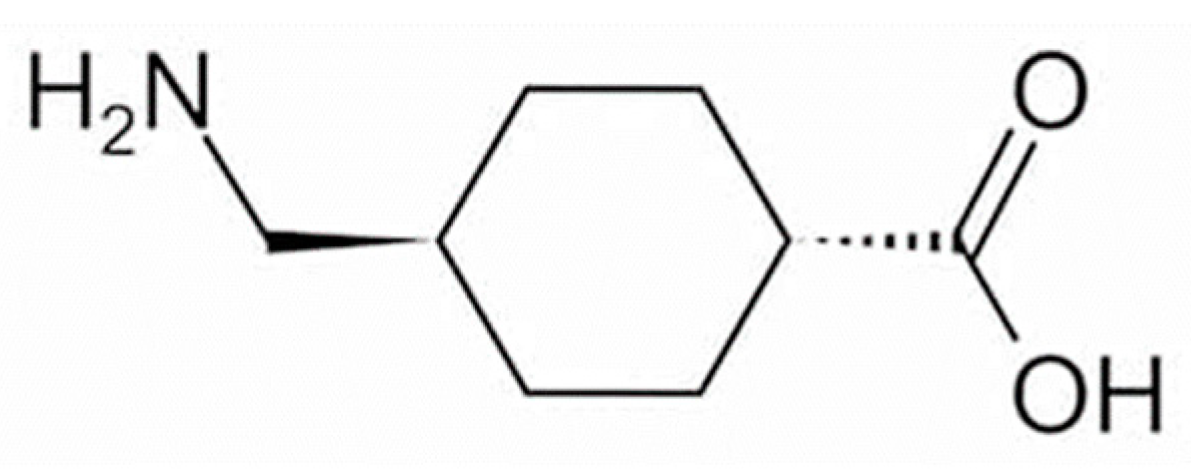
4. Mechanism of Action of TXA
4.1. Mechanism of Action of TXA on Melasma
4.2. Mechanism of Action of TXA on Sun-Induced Skin Hyperpigmentation
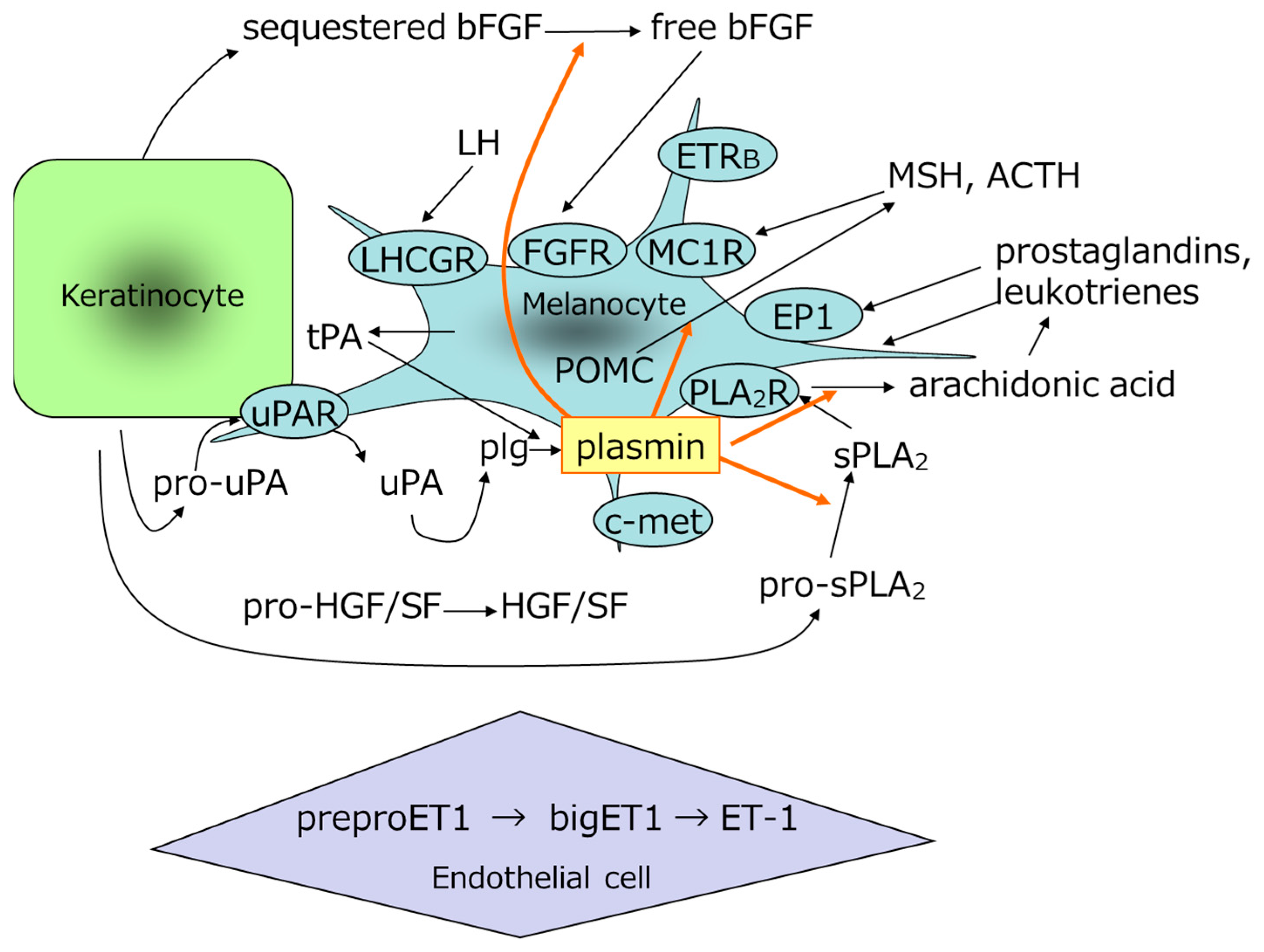
4.3. uPA/Plasminogen System in the Epidermis
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maeda, K. Large melanosome complex is increased in keratinocytes of solar lentigo. Cosmetics 2017, 4, 49. [Google Scholar] [CrossRef]
- Maeda, K. New method of measurement of epidermal turnover in humans. Cosmetics 2017, 4, 47. [Google Scholar] [CrossRef]
- Maeda, K. Timeline of the development of skin-lightening active ingredients in Japan. Molecules 2022, 27, 4774. [Google Scholar] [CrossRef] [PubMed]
- Abiko, Y.; Iwamoto, M. Plasminogen-plasmin system: VII. Potentiation of antifibrinolytic action of a synthetic inhibitor, tranexamic acid, by α2-macroglobulin antiplasmin. Biochim. Biophys. Acta 1970, 214, 411–418. [Google Scholar]
- Japanese Pharmacopoeia and Related Informations. The Japanese Pharmacopoeia 18th Edition, Tranexamic Acid, 1850~1851. Available online: https://jpdb.nihs.go.jp/kyokuhou/indexe.html (accessed on 15 September 2022).
- Dai, L.; Bevan, D.; Rangarajan, S.; Sørensen, B.; Mitchell, M. Stabilization of fibrin clots by activated prothrombin complex concentrate and tranexamic acid in FVIII inhibitor plasma. Haemophilia 2011, 17, e944–e948. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Yang, X.H.; Yang, H.; Yang, Y.P. Study of inhibitory effect of acidum tranexamicum on melanin synthesis. Chin. J. Dermatovenerol. Int. Tradit. West. Med. 2003, 2, 227–229. [Google Scholar]
- Kim, M.S.; Bang, S.H.; Kim, J.H.; Shin, H.J.; Choi, J.H.; Chang, S.E. Tranexamic acid diminishes laser-induced melanogenesis. Ann. Dermatol. 2015, 27, 250–256. [Google Scholar] [CrossRef]
- Lindgren, A.L.; Austin, A.H.; Welsh, K.M. The Use of Tranexamic acid to prevent and treat post-inflammatory hyperpigmentation. J. Drugs Dermatol. 2021, 20, 344–345. [Google Scholar] [CrossRef]
- Passeron, T. Melasma pathogenesis and influencing factors—An overview of the latest research. J. Eur. Acad. Dermatol. Venereol. 2013, 27 (Suppl. 1), 5–6. [Google Scholar] [CrossRef]
- Lee, A.Y. Recent progress in melasma pathogenesis. Pigment Cell Melanoma Res. 2015, 28, 648–660. [Google Scholar] [CrossRef]
- Hernández-Barrera, R.; Torres-Alvarez, B.; Castanedo-Cazares, J.P.; Oros-Ovalle, C.; Moncada, B. Solar elastosis and presence of mast cells as key features in the pathogenesis of melasma. Clin. Exp. Dermatol. 2008, 33, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.H.; Kim, Y.C.; Lee, E.S.; Kang, H.Y. The vascular characteristics of melasma. J. Dermatol. Sci. 2007, 46, 111–116. [Google Scholar] [CrossRef]
- Kim, S.J.; Park, J.Y.; Shibata, T.; Fujiwara, R.; Kang, H.Y. Efficacy and possible mechanisms of topical tranexamic acid in melasma. Clin. Exp. Dermatol. 2016, 41, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, M.; Kawada, A.; Takiwaki, H.; Mizuno, A.; Torii, H.; Hayashi, N.; Nogita, T.; Akiyoshi, E.; Yoshikawa, N.; Watanabe, C.; et al. Clinical efficacy of DH-4243 for Chloasma: A multi-center randomized controlled trial. Jpn. J. Clin. Dermatol. 2007, 61, 735–743. (In Japanese) [Google Scholar]
- Na, J.I.; Choi, S.Y.; Yang, S.H.; Choi, H.R.; Kang, H.Y.; Park, K.C. Effect of TXA on melasma: A clinical trial with histological evaluation. J. Eur. Acad. Dermatol. Venereol. 2013, 27, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.C.; Thing, T.G.; Goh, C.L. Oral Tranexamic acid (TA) in the treatment of melasma: A retrospective analysis. J. Am. Acad. Dermatol. 2016, 75, 385–392. [Google Scholar] [CrossRef]
- Del Rosario, E.; Florez-Pollack, S.; Zapata, L., Jr.; Hernandez, K.; Tovar-Garza, A.; Rodrigues, M. Randomized, placebo-controlled, double-blind study of oral tranexamic acid in the treatment of moderate to severe melasma. J. Am. Acad. Dermatol. 2018, 78, 63–369. [Google Scholar] [CrossRef]
- Tse, T.W.; Hui, E. Tranexamic acid: An important adjuvant in the treatment of melasma. J. Cosmet. Dermatol. 2013, 12, 57–66. [Google Scholar] [CrossRef]
- Cho, H.H.; Choi, M.; Cho, S.; Lee, J.H. Role of oral tranexamic acid in melasma patients treated with IPL and low fluence QS Nd: YAG laser. J. Dermatolog. Treat. 2013, 24, 292–296. [Google Scholar] [CrossRef]
- Wu, S.; Shi, H.; Wu, H.; Yan, S.; Guo, J.; Sun, Y.; Pan, L. Treatment of melasma with oral administration of tranexamic acid. Aesthetic Plast. Surg. 2012, 36, 964–970. [Google Scholar] [CrossRef]
- Kato, H.; Araki, J.; Eto, H.; Doi, K.; Hirai, R.; Kuno, S.; Higashino, T.; Yoshimura, K. A prospective randomized controlled study of oral tranexamic acid for preventing postinflammatory hyperpigmentation after Q-switched ruby laser. Dermatol. Surg. 2011, 37, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Kondo, S.; Okada, Y.; Tomita, Y. Clinical study of effect of tranexamic acid emulsion on melasma and freckles. Skin Res. 2007, 6, 309–315. (In Japanese) [Google Scholar]
- Ebrahimi, B.; Naeini, F.F. Topical tranexamic acid as a promising treatment for melasma. J. Res. Med. Sci. 2014, 19, 753–757. [Google Scholar] [PubMed]
- Banihashemi, M.; Zabolinejad, N.; Jaafari, M.R.; Salehi, M.; Jabari, A. Comparison of therapeutic effects of liposomal tranexamic acid and conventional hydroquinone on melasma. J. Cosmet. Dermatol. 2015, 14, 174–177. [Google Scholar] [CrossRef] [PubMed]
- Na Ayuthaya, P.K.; Niumphradit, N.; Manosroi, A.; Nakakes, A. Topical 5% tranexamic acid for the treatment of melasma in Asians: A double-blind randomized controlled clinical trial. J. Cosmet. Laser Ther. 2012, 14, 150–154. [Google Scholar] [CrossRef]
- Xu, Y.; Ma, R.; Juliandri, J.; Wang, X.; Xu, B.; Wang, D.; Lu, Y.; Zhou, B.; Luo, D. Efficacy of functional microarray of microneedles combined with topical tranexamic acid for melasma: A randomized, self-controlled, split-face study. Medicine 2017, 96, e6897. [Google Scholar] [CrossRef]
- Shihab, N.; Prihartono, J.; Tovar-Garza, A.; Agustin, T.; Legiawati, L.; Pandya, A.G. Randomised, controlled, double-blind study of combination therapy of oral tranexamic acid and topical hydroquinone in the treatment of melasma. Australas. J. Dermatol. 2020, 61, 237–242. [Google Scholar] [CrossRef]
- Igarashi, M.; Tomita, Y.; Seiji, M. Hydroquinone therapy for chloasma. Rinsho Derma 1977, 19, 761–765. (In Japanese) [Google Scholar]
- Shin, J.U.; Park, J.; Oh, S.H.; Lee, J.H. Oral tranexamic acid enhances the efficacy of low-fluence 1064-nm Q-switched neodymium-doped yttrium aluminum garnet laser treatment for melasma in Koreans: A randomized, prospective trial. Dermatol. Surg. 2013, 39, 435–442. [Google Scholar] [CrossRef]
- Qu, Y.; Wang, F.; Liu, J.; Xia, X. Clinical observation and dermoscopy evaluation of fractional CO2 laser combined with topical tranexamic acid in melasma treatments. J. Cosmet. Dermatol. 2021, 20, 1110–1116. [Google Scholar] [CrossRef]
- Agamia, N.; Apalla, Z.; Salem, W.; Abdallah, W. A comparative study between oral tranexamic acid versus oral tranexamic acid and Q-switched Nd-YAG laser in melasma treatment: A clinical and dermoscopic evaluation. J. Dermatolog. Treat. 2021, 32, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Maeda, K. Tranexamic acid. Mon. Book Derma 2005, 98, 35–42. (In Japanese) [Google Scholar]
- Higashi, N. Treatment of melasma with oral tranexamic acid. Ski. Res. 1988, 30, 676–680. (In Japanese) [Google Scholar]
- Kita, Y.; Sugai, T. Effect of bleach agents on chloasma. Ski. Res. 1992, 34, 142–146. (In Japanese) [Google Scholar]
- Zhu, C.Y.; Li, Y.; Sun, Q.N.; Takada, A.; Kawada, A. Analysis of the effect of different doses of oral tranexamic acid on melasma: A multicentre prospective study. Eur. J. Dermatol. 2019, 29, 55–58. [Google Scholar]
- Miescher, G.; Minder, H. Untersuchungen über die durch langwelliges Ultraviolett hervorgerufene Pigmetdunkelung. Strahlentherapie 1939, 66, 6–23. [Google Scholar]
- Pathak, M.A.; Stratton, K. Free radicals in human skin before and after exposure to light. Arch. Biochem. Biophys. 1968, 123, 468–476. [Google Scholar] [CrossRef]
- Mizuno, N. Behavior of melanocyte after single ultraviolet irradiation. Jpn. J. Clin. Dermatol. 1968, 22, 131–143. (In Japanese) [Google Scholar]
- Eller, M.S.; Yaar, S.M.; Gilchrest, B.A. DNA damage and melanogenesis. Nature 1994, 372, 413–414. [Google Scholar] [CrossRef]
- Abdel-Malek, Z.A. Endocrine factors as effectors of integumental pigmentation. In Dermatologic Clinics; Nordlund, J.J., Ed.; W.B. Saunders: New York, NY, USA, 1988; Volume 6, pp. 175–183. [Google Scholar]
- Nordlund, J.J.; Abdel-Malek, Z.A.; Boissy, R.E.; Rheins, L.A. Pigment cell biology: An historical review. J. Invest. Dermatol. 1989, 92, 53S–60S. [Google Scholar] [CrossRef]
- Gilchrest, B.A.; Park, H.-Y.; Eller, M.S.; Yaar, M. Mechanisms of ultraviolet light-induced pigmentation. Photochem. Photobiol. 1996, 63, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tomita, Y.; Maeda, K.; Tagami, H. Mechanisms for hyperpigmentation in postinflammatory pigmentation, ulticaria pigmentosa and sunburn. Dermatologica 1989, 179, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Nordlund, J.J.; Collins, C.E.; Rheins, L.A. Prostaglandin E2 and D2 but not MSH stimulate the proliferation of pigment cells in the pinnal epidermis of the DBA/2 mouse. J. Invest. Dermatol. 1986, 86, 433–437. [Google Scholar] [CrossRef] [PubMed]
- Tomita, Y.; Maeda, K.; Tagami, H. Melanocyte-stimulating properties of arachidonic acid metabolites: Possible role in postinflammatory pigmentation. Pigment. Cell Res. 1992, 5, 357–361. [Google Scholar] [CrossRef]
- Maeda, K.; Naganuma, M. Melanocyte-stimulating properties of secretory phospholipase A2. Photochem. Photobiol. 1997, 65, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Birchall, N.; Orlow, S.J.; Kupper, T.; Pawelek, J. Interactions between ultraviolet light and interleukin-1 on MSH binding in both mouse melanoma and human sequamous carcinoma cells. Biochem. Biophys. Res. Commun. 1991, 175, 839–845. [Google Scholar] [CrossRef]
- Schauer, E.; Trautinger, F.; Köck, A.; Schwarz, A.; Bhardwaj, R.; Simon, M.; Ansel, J.C.; Schwarz, T.; Luger, T.A. Proopiomelanocortin-derived peptides are synthesized and released by human keratinocytes. J. Clin. Invest. 1994, 93, 2258–2262. [Google Scholar] [CrossRef]
- Halaban, R.; Langdon, R.; Birchall, N.; Cuono, C.; Baird, A.; Scott, G.; Moellmann, G.; McGuire, J. Basic fibroblast growth factor from human keratinocytes is a natural mitogen for melanocytes. J. Cell Biol. 1988, 107, 1611–1619. [Google Scholar] [CrossRef]
- Imokawa, G.; Yada, Y.; Miyagishi, M. Endothelins secreted from human keratinocytes are intrinsic mitogens for human melanocytes. J. Biol. Chem. 1992, 267, 24675–24680. [Google Scholar] [CrossRef]
- Tomita, Y.; Maeda, K.; Tagami, H. Stimulatory effect of histamine on normal human melanocytes in vitro. Tohoku J. Exp. Med. 1998, 155, 209–210. [Google Scholar] [CrossRef]
- Chang, W.C.; Shi, G.Y.; Chow, Y.H.; Chang, L.C.; Hau, J.S.; Lin, M.T.; Jen, C.J.; Wing, L.Y.; Wu, H.L. Human plasmin induces a receptor-mediated arachidonate release coupled with G proteins in endothelial cells. Am. J. Physiol. 1993, 264, C271–C281. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, Y.; Kitamura, S.; Kosaka, K.; Harasawa, M. Tranexamic acid no prostaglandin gouseisogai ni kansuru kenkyu. Jpn. Pharmacol. Ther. 1978, 6, 398–402. (In Japanese) [Google Scholar]
- Weide, I.; Tippler, B.; Syrovets, T.; Simmet, T. Plasmin is a specific stimulus of the 5-lipoxygenase pathway of human peripheral monocytes. Thromb. Haemost. 1996, 76, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, H.; Akamatsu, H.; Matoba, Y.; Ri, S.; Ito, A.; Asada, Y. Effects of Tranexamic Acid on Neutrophil Chemotaxis, Phagocytosis and Reactive Oxygen Species Generation in vitro. Jpn. Pharmacol. Ther. 1994, 22, 1429–1435. (In Japanese) [Google Scholar]
- Toki, N.; Takasugi, S.; Fujii, K. Basic research of histaminergic drugs and antihistaminergic drugs. Med. Consult. New Remedies 1981, 18, 1195–1202. (In Japanese) [Google Scholar]
- Xing, X.; Xu, Z.; Chen, L.; Jin, S.; Zhang, C.; Xiang, L. Tranexamic acid inhibits melanogenesis partially via stimulation of TGF-β1 expression in human epidermal keratinocytes. Exp. Dermatol. 2022, 31, 633–640. [Google Scholar] [CrossRef]
- Zhu, J.W.; Ni, Y.J.; Tong, X.Y.; Guo, X.; Wu, X.P.; Lu, Z.F. Tranexamic acid inhibits angiogenesis and melanogenesis in vitro by targeting VEGF receptors. Int. J. Med. Sci. 2020, 17, 903–911. [Google Scholar] [CrossRef]
- Tomita, Y.; Maeda, K.; Tagami, H. Leukotrienes and thromboxane B2 stimulate normal human melanocytes in vitro: Possible inducers of postinflammatory pigmentation. Tohoku J. Exp. Med. 1988, 156, 303–304. [Google Scholar] [CrossRef]
- Morelli, J.G.; Hake, S.S.; Murphy, R.C.; Norris, D.A. Leukotriene B4-induced human melanocyte pigmentation and leukotriene C4-induced human melanocyte growth are inhibited by different isoquinolinesulfonamides. J. Invest. Dermatol. 1992, 98, 55–58. [Google Scholar] [CrossRef]
- Maeda, K.; Naganuma, M. Topical trans-4-aminomethylcyclohexanecarboxylic acid prevents ultraviolet radiation-induced pigmentation. J. Photochem. Photobiol. B Biol. 1998, 47, 136–141. [Google Scholar] [CrossRef]
- Nakano, T.; Fujita, H.; Kikuchi, N.; Arita, H. Plasmin converts pro-form of group I phospholipase A2 into receptor binding, active forms. Biochem. Biophys. Res. Commun. 1994, 198, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, N.P.; Pathak, M.A.; Sato, S.; Fitzpatrick, T.B.; Sanchez, J.L.; Mihm, M.C., Jr. Melasma: A clinical, light microscopic, ultrastructural, and immunofluorescence study. J. Am. Acad. Dermatol. 1981, 4, 698–710. [Google Scholar] [CrossRef]
- Kang, W.H.; Yoon, K.H.; Lee, E.S.; Kim, J.; Lee, K.B.; Yim, H.; Sohn, S.; Im, S. Melasma: Histopathological characteristics in 56 Korean patients. Br. J. Dermatol. 2002, 146, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Hunt, G.; Donatien, P.D.; Lunec, J.; Todd, C.; Kyne, S.; Thody, A.J. Cultured human melanocytes respond to MSH peptides and ACTH. Pigment Cell Res. 1994, 7, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Grabbe, J.; Welker, P.; Dippel, E.; Czarnetzki, B.M. Stem cell factor, a novel cutaneous growth factor for mast cells and melanocytes. Arch. Dermatol. Res. 1994, 287, 78–84. [Google Scholar] [CrossRef]
- Iversen, L.; Kristensen, P.; Grøn, B.; Ziboh, V.A.; Kragballe, K. Human epidermis transforms exogenous leukotriene A4 into peptide leukotrienes: Possible role in transcellular metabolism. Arch Dermatol. Res. 1994, 286, 261–266. [Google Scholar] [CrossRef]
- Man, M.Q.; Lin, T.K.; Santiago, J.L.; Celli, A.; Zhong, L.; Huang, Z.M.; Roelandt, T.; Hupe, M.; Sundberg, J.P.; Silva, K.A.; et al. Basis for enhanced barrier function of pigmented skin. J. Invest. Dermatol. 2014, 134, 2399–2407. [Google Scholar] [CrossRef]
- Yan, B.X.; Zheng, Y.X.; Li, W.; Chen, J.Q.; Zhou, J.; Cai, S.Q.; Zheng, M.; Man, X.Y. Comparative expression of PEDF and VEGF in human epidermal keratinocytes and dermal fibroblasts: From normal skin to psoriasis. Discov. Med. 2018, 25, 47–56. [Google Scholar]
- Khunger, N.; Kandhari, R.; Singh, A.; Ramesh, V. A clinical, dermoscopic, histopathological and immunohistochemical study of melasma and facial pigmentary demarcation lines in the skin of color. Dermatol. Ther. 2020, 33, e14515. [Google Scholar] [CrossRef]
- Bhardwaj, R.S.; Luger, T.A. Proopiomelanocortin production by epidermal cells: Evidence for an immune neuroendocrine network in the epidermis. Arch. Dermatol. Res. 1994, 287, 85–90. [Google Scholar] [CrossRef]
- Miot, L.D.; Miot, H.A.; Polettini, J.; Silva, M.G.; Marques, M.E. Morphologic changes and the expression of alpha-melanocyte stimulating hormone and melanocortin-1 receptor in melasma lesions: A comparative study. Am. J. Dermatopathol. 2010, 32, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Im, S.; Kim, J.; On, W.Y.; Kang, W.H. Increased expression of alpha-melanocyte-stimulating hormone in the lesional skin of melasma. Br. J. Dermatol. 2002, 146, 165–167. [Google Scholar] [PubMed]
- Kang, H.Y.; Hwang, J.S.; Lee, J.Y.; Ahn, J.H.; Kim, J.Y.; Lee, E.S.; Kang, W.H. The dermal stem cell factor and c-kit are overexpressed in melasma. Br. J. Dermatol. 2006, 154, 1094–1099. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.Y.; Suzuki, I.; Lee, D.J.; Ha, J.; Reiniche, P.; Aubert, J.; Deret, S.; Zugaj, D.; Voegel, J.J.; Ortonne, J.P. Transcriptional profiling shows altered expression of wnt pathway- and lipid metabolism-related genes as well as melanogenesis-related genes in melasma. J. Invest. Dermatol. 2011, 131, 1692–1700. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Zhang, L.; Miles, L.; Hoover-Plow, J. Plasminogen regulates pro-opiomelanocortin processing. J. Thromb. Haemost. 2004, 2, 785–796. [Google Scholar] [CrossRef]
- Falcone, D.J.; McCaffrey, T.A.; Haimovitz-Friedman, A.; Vergilio, J.A.; Nicholson, A.C. Macrophage and foam cell release of matrix-bound growth factors. Role of plasminogen activation. J. Biol. Chem. 1993, 268, 11951–11958. [Google Scholar] [CrossRef]
- Syrovets, T.; Jendrach, M.; Rohwedder, A.; Schüle, A.; Simmet, T. Plasmin-induced expression of cytokines and tissue factor in human monocytes involves AP-1 and IKKbeta-mediated NF-kappaB activation. Blood 2001, 97, 3941–3950. [Google Scholar] [CrossRef]
- Kamio, N.; Hashizume, H.; Nakao, S.; Matsushima, K.; Sugiya, H. Plasmin is involved in inflammation via protease-activated receptor-1 activation in human dental pulp. Biochem. Pharmacol. 2008, 75, 1974–1980. [Google Scholar] [CrossRef]
- Burysek, L.; Syrovets, T.; Simmet, T. The serine protease plasmin triggers expression of MCP-1 and CD40 in human primary monocytes via activation of p38 MAPK and janus kinase (JAK)/STAT signaling pathways. J. Biol. Chem. 2002, 277, 33509–33517. [Google Scholar] [CrossRef]
- Matsumura, Y.; Takada, K.; Murakami, A.; Takaoka, M.; Morimoto, S. Plasmin stimulates expression of endothelin-1 mRNA and endothelin-1 release in vascular endothelial cells. Life Sci. 1996, 58, 1067–1074. [Google Scholar] [CrossRef]
- Naldini, L.; Tamagnone, L.; Vigna, E.; Sachs, M.; Hartmann, G.; Birchmeier, W.; Daikuhara, Y.; Tsubouchi, H.; Blasi, F.; Comoglio, P.M. Extracellular proteolytic cleavage by urokinase is required for activation of hepatocyte growth factor/scatter factor. EMBO J. 1992, 11, 4825–4833. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.H.; Park, J.E.; Lim, D.S.; Lee, J.S. Tranexamic acid inhibits melanogenesis by activating the autophagy system in cultured melanoma cells. J. Dermatol. Sci. 2017, 88, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Tomita, Y.; Iwamoto, M.; Masuda, T.; Tagami, H. Stimulatory effect of prostaglandin E2 on the configulation of normal human melanocytes in vitro. J. Invest. Dermatol. 1987, 89, 299–301. [Google Scholar] [CrossRef]
- Abdel-Malek, Z.A.; Swope, V.B.; Amornsiripanitch, N.; Nordlund, J.J. In vitro modulation of proliferation and melanization of S91 melanoma cells by prostaglandins. Cancer Res. 1987, 47, 3141–3146. [Google Scholar] [PubMed]
- Imokawa, G.; Motegi, I. Skin organ culture model for examining epidermal melanization. J. Invest. Dermatol. 1993, 100, 47–54. [Google Scholar] [CrossRef]
- Takiwaki, H.; Shirai, S.; Kohno, H.; Soh, H.; Arase, S. The degrees of UVB-induced erythema and pigmentation correlate linealy and are reduced in a parallel manner by topical anti-inflammatory agents. J. Invest. Dermatol. 1994, 103, 642–646. [Google Scholar] [CrossRef] [PubMed]
- Black, A.K.; Greaves, M.W.; Hensby, C.N.; Plummer, N.A. Increased prostaglandins E2 and F2 alpha in human skin at 6 and 24 h after ultraviolet B irradiation (290–320 nm). Br. J. Clin. Pharmac. 1978, 5, 431–436. [Google Scholar] [CrossRef]
- Black, A.K.; Fincham, N.; Greaves, M.W.; Hensby, C.N. Time course changes in levels of arachidonic acid and prostaglandins D2, E2, F2 alpha in human skin following ultraviolet B irradiation. Br. J. Clin. Pharmac. 1980, 10, 453–457. [Google Scholar] [CrossRef]
- Hawk, J.L.M.; Black, A.K.; Jaenicke, K.F.; Barr, R.M.; Soter, N.A.; Mallett, A.I.; Gilchrest, B.A.; Hensby, C.N.; Parrish, J.A.; Greaves, M.W. Increased concentrations of arachidonic acid, prostaglandins E2, D2, and 6-oxo-F1 alpha, and histamine in human skin following UVA irradiation. J. Invest. Dermatol. 1983, 80, 496–499. [Google Scholar] [CrossRef]
- Mikoshiba, H.; Takei, M.; Takase, Y.; Nijo, S.; Shimosato, F.; Nomoto, S. Kanpan ni taisuru tranekisamsan naifuku ryouhou. Nishi Nihon Hifuka 1985, 47, 1101–1104. (In Japanese) [Google Scholar] [CrossRef]
- Li, D.; Shi, Y.; Li, M.; Liu, J.; Feng, X. Tranexamic acid can treat ultraviolet radiation-induced pigmentation in guinea pigs. Eur. J. Dermatol. 2010, 20, 89–92. [Google Scholar] [CrossRef]
- Maeda, K.; Tomita, Y. Mechanism of the inhibitory effect of tranexamic acid on melanogenesis in cultured human melanocytes in the presence of keratinocyte-conditioned medium. J. Health Sci. 2007, 53, 389–396. [Google Scholar] [CrossRef]
- Takada, A.; Takada, Y. Inhibition by tranexamic acid of the conversion of single-chain tissue plasminogen activator to its two chain form by plasmin: The presence on tissue plasminogen activator of a site to bind with lysine binding sites of plasmin. Thromb. Res. 1989, 55, 717–725. [Google Scholar] [CrossRef]
- Miles, L.A.; Dahlberg, C.M.; Plescia, J.; Felez, J.; Kato, K.; Plow, E.F. Role of cell-surface lysines in plasminogen binding to cells: Identification of alpha-enolase as a candidate plasminogen receptor. Biochemistry 1991, 30, 1682–1691. [Google Scholar] [CrossRef]
- Plow, E.F.; Herren, T.; Redlitz, A.; Miles, L.A.; Hoover-Plow, J.L. The cell biology of the plasminogen system. FASEB J. 1995, 9, 939–9455. [Google Scholar] [CrossRef]
- Bizik, J.; Stephens, R.W.; Grofova, M.; Vaheri, A. Binding of tissue-type plasminogen activator to human melanoma cells. J. Cell Biochem. 1993, 51, 326–335. [Google Scholar] [CrossRef]
- Isseroff, R.R.; Rifkin, D.B. Plasminogen is present in the basal layer of the epidermis. J. Invest. Dermatol. 1983, 80, 297–299. [Google Scholar] [CrossRef]
- Spiers, E.M.; Lazarus, G.S.; Lyons-Giordano, B. Expression of plasminogen activators in psoriatic epidermis. J. Invest. Dermatol. 1994, 102, 333–338. [Google Scholar] [CrossRef][Green Version]
- Loud, L.R.; Eriksen, J.; Ralfkiaer, E.; Romer, J. Differential expression of urokinase plasminogen activator, its receptor, and inhibitors in mouse skin after exposure to a tumor-promoting phorbol ester. J. Invest. Dermatol. 1996, 106, 622–630. [Google Scholar]
- Ichikawa, K.; Takashima, A.; Yasuda, S.; Mizuno, N. Enhanced rabbit skin plasmin activity by UV irradiation. Dermatologica 1989, 179 (Suppl. 1), 132. [Google Scholar] [CrossRef]
- Takashima, A.; Yasuda, S.; Mizuno, N. Determination of the action spectrum for UV-induced plasminogen activator synthesis in mouse keratinocytes in vitro. J. Dermatol. Sci. 1992, 4, 11–17. [Google Scholar] [CrossRef]
- Rotem, N.; Axelrod, J.H.; Miskin, R. Induction of urokinase-type plasminogen activator by UV light in human fetal fibroblasts is mediated through a UV-induced secreted protein. Mol. Cell Biol. 1987, 7, 622–631. [Google Scholar]
- Kang-Rotondo, C.H.; Miller, C.C.; Morrison, A.R.; Pentland, A.P. Enhanced keratinocyte prostaglandin synthesis after UV injury is due to increased phospholipase activity. Am. J. Physiol. 1993, 264, 396–401. [Google Scholar] [CrossRef]
- Grewe, M.; Trefzer, U.; Ballhorn, A.; Gyufko, K.; Henninger, H.; Krutmann, J. Analysis of the mechanism of ultraviolet (UV) B radiation-induced prostaglandin E2 synthesis by human epidermoid carcinoma cells. J. Invest. Dermatol. 1993, 101, 528–531. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Horikoshi, T.; Eguchi, H.; Onodera, H. The effects of tranexamic acid on the growth and melanogenesis of cultured human melanocytes. Jpn. J. Dermatol. 1994, 104, 641–646. [Google Scholar]
- Cario, M. How hormones may modulate human skin pigmentation in melasma: An in vitro perspective. Exp. Dermatol. 2019, 28, 709–718. [Google Scholar] [CrossRef]
- Peterson, R.A.; Krull, P.E.; Finley, P.; Ettinger, M.G. Changes in antithrombin 3 and plasminogen induced by oral contraceptives. Am. J. Clin. Pathol. 1970, 53, 468–473. [Google Scholar] [CrossRef] [PubMed]
- Zaitsu, M.; Narita, S.; Lambert, K.C.; Grady, J.J.; Estes, D.M.; Curran, E.M.; Brooks, E.G.; Watson, C.S.; Goldblum, R.M.; Midoro-Horiuti, T. Estradiol activates mast cells via a non-genomic estrogen receptor-alpha and calcium influx. Mol. Immunol. 2007, 44, 1977–1985. [Google Scholar] [CrossRef]
- Shoemaker, L.N.; Haigh, K.M.; Kuczmarski, A.V.; McGinty, S.J.; Welti, L.M.; Hobson, J.C.; Edwards, D.G.; Feinberg, R.F.; Wenner, M.M. ETB receptor-mediated vasodilation is regulated by estradiol in young women. Am. J. Physiol. Heart Circ. Physiol. 2021, 321, H592–H598. [Google Scholar] [CrossRef]
- Regazzetti, C.; De Donatis, G.M.; Ghorbel, H.H.; Cardot-Leccia, N.; Ambrosetti, D.; Bahadoran, P.; Chignon-Sicard, B.; Lacour, J.P.; Ballotti, R.; Mahns, A.; et al. Endothelial cells promote pigmentation through endothelin receptor B activation. J. Invest. Dermatol. 2015, 135, 3096–3104. [Google Scholar] [CrossRef]
- Pérez, M.; Sánchez, J.L.; Aguiló, F. Endocrinologic profile of patients with idiopathic melasma. J. Invest. Dermatol. 1983, 81, 543–545. [Google Scholar] [CrossRef] [PubMed]
- Hassan, I.; Kaur, I.; Sialy, R.; Dash, R.J. Hormonal milieu in the maintenance of melasma in fertile women. J. Dermatol. 1998, 25, 510–512. [Google Scholar] [CrossRef] [PubMed]
- Sialy, R.; Hassan, I.; Kaur, I.; Dash, R.J. Melasma in men: A hormonal profile. J. Dermatol. 2000, 27, 64–65. [Google Scholar] [CrossRef] [PubMed]
- Sato, N. Endocrine environment in adult females with chloasma. Jpn. J. Dermatol. 1987, 97, 937–943. (In Japanese) [Google Scholar]
- Reich, R.; Miskin, R.; Tsafriri, A. Follicular plasminogen activator: Involvement in ovulation. Endocrinology 1985, 116, 516–521. [Google Scholar] [CrossRef]
- Moraga, P.F.; Llanos, M.N.; Ronco, A.M. Arachidonic acid release from rat Leydig cells depends on the presence of luteinizing hormone/human chorionic gonadotrophin receptors. J. Endocrinol. 1997, 154, 201–209. [Google Scholar] [CrossRef]
- Maeda, K.; Naganuma, M.; Fukuda, M.; Matsunaga, J.; Tomita, Y. Effect of pituitary and ovarian hormones on human melanocytes in vitro. Pigment. Cell Res. 1996, 9, 204–212. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors Released or Activated by Plasmin | Literature |
|---|---|
| Release of kinin, kallikrein | 5 |
| Activation of phospholipase A2 | 63 |
| Release of arachidonic acid | 53 |
| Enhanced leukotriene formation | 55 |
| Processing of MSH from POMC | 77 |
| Release of bFGF | 78 |
| Activation of TGF-β | 78 |
| Increase in mRNA for IL-1α, IL-1β, and TNF α | 79 |
| Increase in IL-8 mRNA | 80 |
| Activation of NF-κB and AP-1 | 79 |
| Increase in MCP-1 mRNA | 81 |
| Increase phosphorylation of JAK1, STAT1, and STAT3 | 81 |
| Activation of P38 MAP kinase | 81 |
| Increase in preproendothelin-1 mRNA | 82 |
| Activation of HGF/SF | 83 |
| Possible Mechanisms of Hyperpigmentation | Literature | Involvement of Epidermal uPA/Plasminogen System |
|---|---|---|
| Activation of phospholipase A2 in epidermis | 69 | Yes |
| Release of arachidonic acid in epidermis | 43 | Yes |
| Increase PGE2 in epidermis | 45 | Yes |
| Enhance leukotriene formation in epidermis | 68 | Yes |
| Processing of MSH from POMC in epidermis | 43, 77 | Yes |
| Activation of TGF-β in keratinocytes | 58 | Yes |
| bFGF release in the epidermis | 43 | Yes |
| Increase vascular in dermis | 13 | No |
| Increase mast cell in dermis | 28 | No |
| Activation of HGF/SF | 83 | No |
| Increase preproendothelin-1 in vascular endothelial cells | 82 | No |
| Increased VEGF in epidermis | 70 | Unknown |
| Increase in autophagy system | 84 | Unknown |
| Increase Wnt inhibitory factor 1 and Wnt5a | 76 | Unknown |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maeda, K. Mechanism of Action of Topical Tranexamic Acid in the Treatment of Melasma and Sun-Induced Skin Hyperpigmentation. Cosmetics 2022, 9, 108. https://doi.org/10.3390/cosmetics9050108
Maeda K. Mechanism of Action of Topical Tranexamic Acid in the Treatment of Melasma and Sun-Induced Skin Hyperpigmentation. Cosmetics. 2022; 9(5):108. https://doi.org/10.3390/cosmetics9050108
Chicago/Turabian StyleMaeda, Kazuhisa. 2022. "Mechanism of Action of Topical Tranexamic Acid in the Treatment of Melasma and Sun-Induced Skin Hyperpigmentation" Cosmetics 9, no. 5: 108. https://doi.org/10.3390/cosmetics9050108
APA StyleMaeda, K. (2022). Mechanism of Action of Topical Tranexamic Acid in the Treatment of Melasma and Sun-Induced Skin Hyperpigmentation. Cosmetics, 9(5), 108. https://doi.org/10.3390/cosmetics9050108