
Codonopsis pilosula Extract Protects Melanocytes against H2O2-Induced Oxidative Stress by Activating Autophagy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Preparation of the Codonopsis Pilosula Extract
2.3. Cell Culture and Stock Preparation of CPE
2.4. Measurement of Cell Viability
2.5. In Vitro Melanin Estimation
2.6. Protein Isolation and Western Blot Assay
2.7. Estimation of Intracellular ROS by DCFH-DA Staining
2.8. Statistical Processing
3. Results
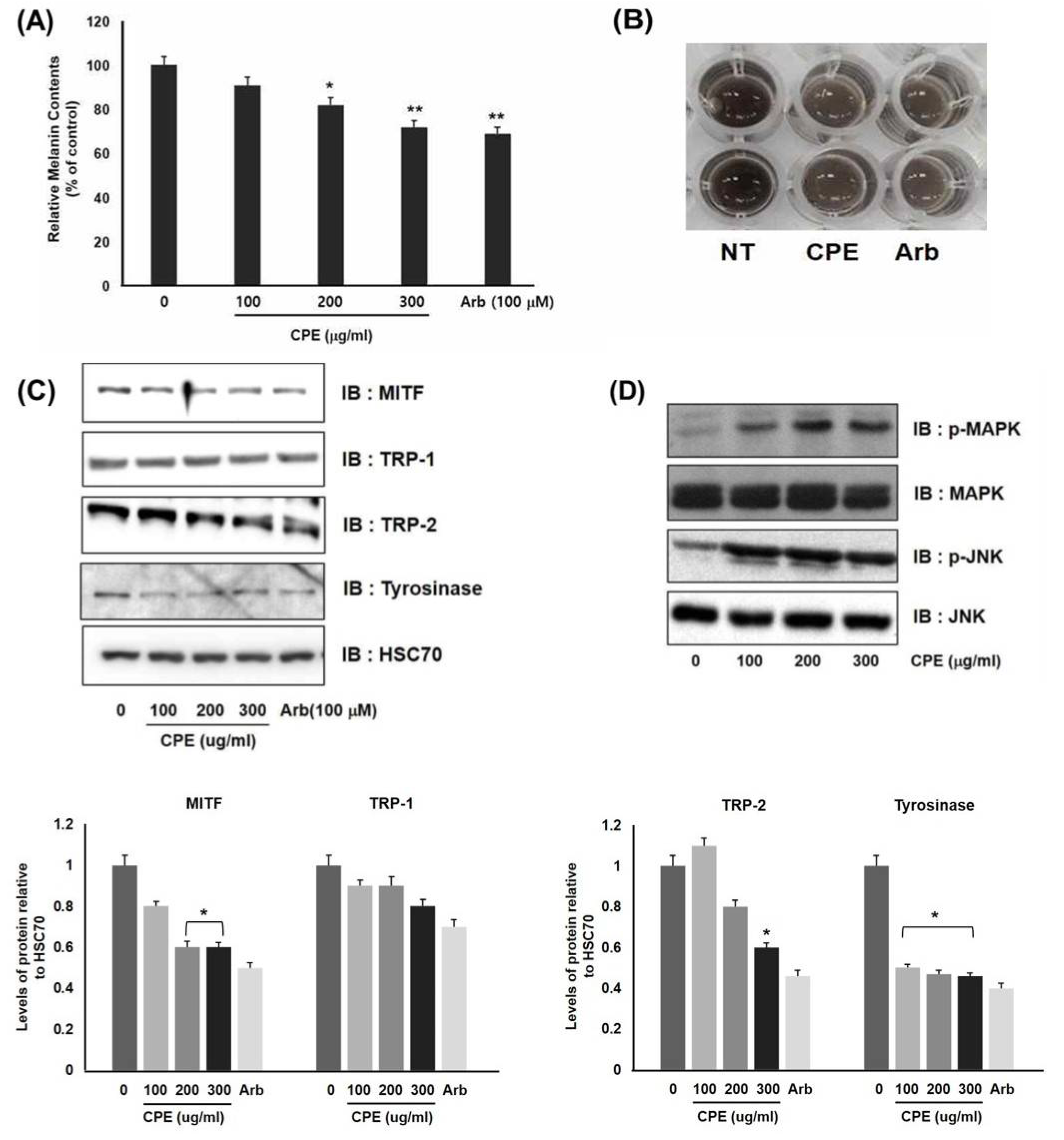
3.1. Downregulation of Melanogenesis in Melanocytes by CPE
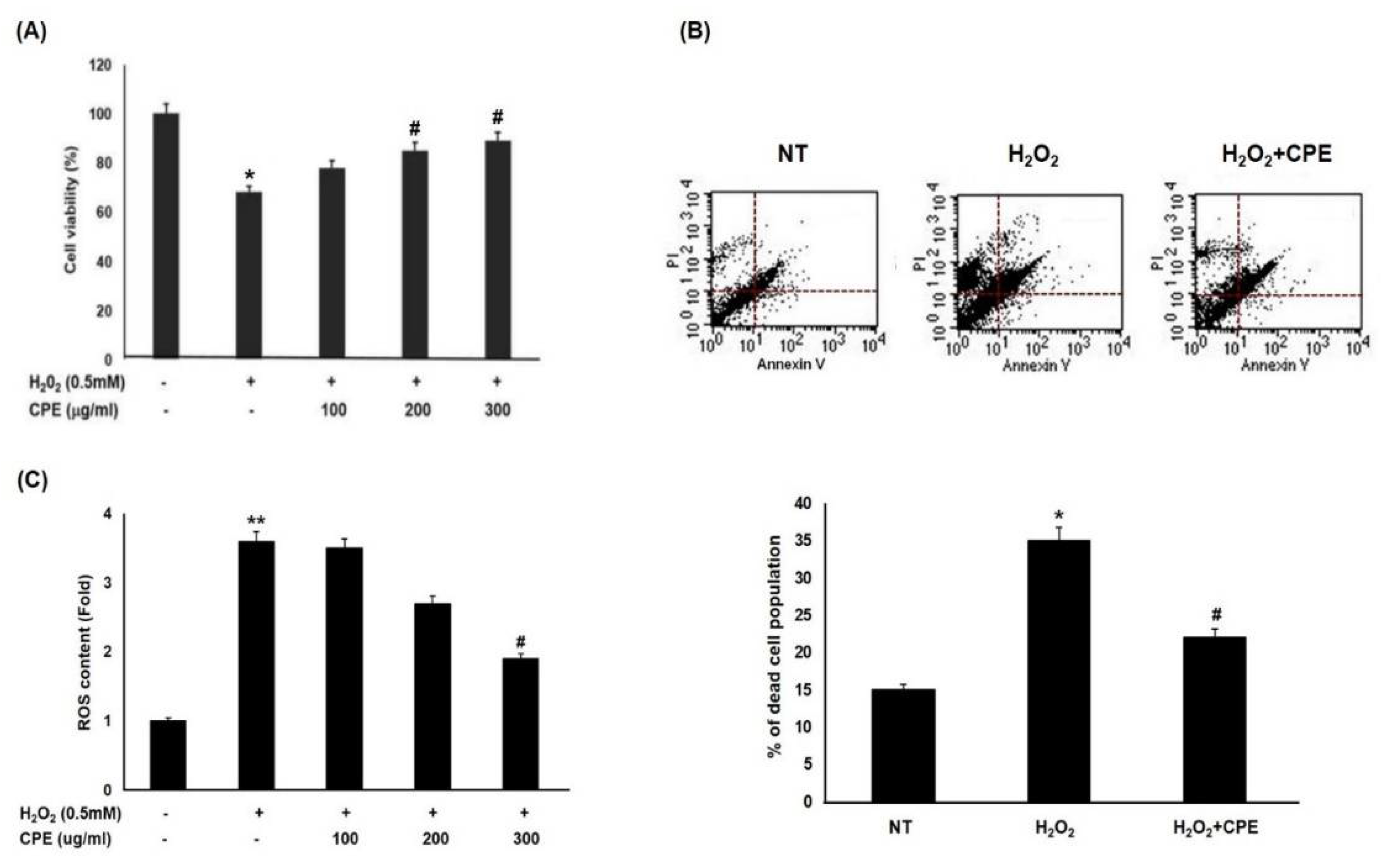
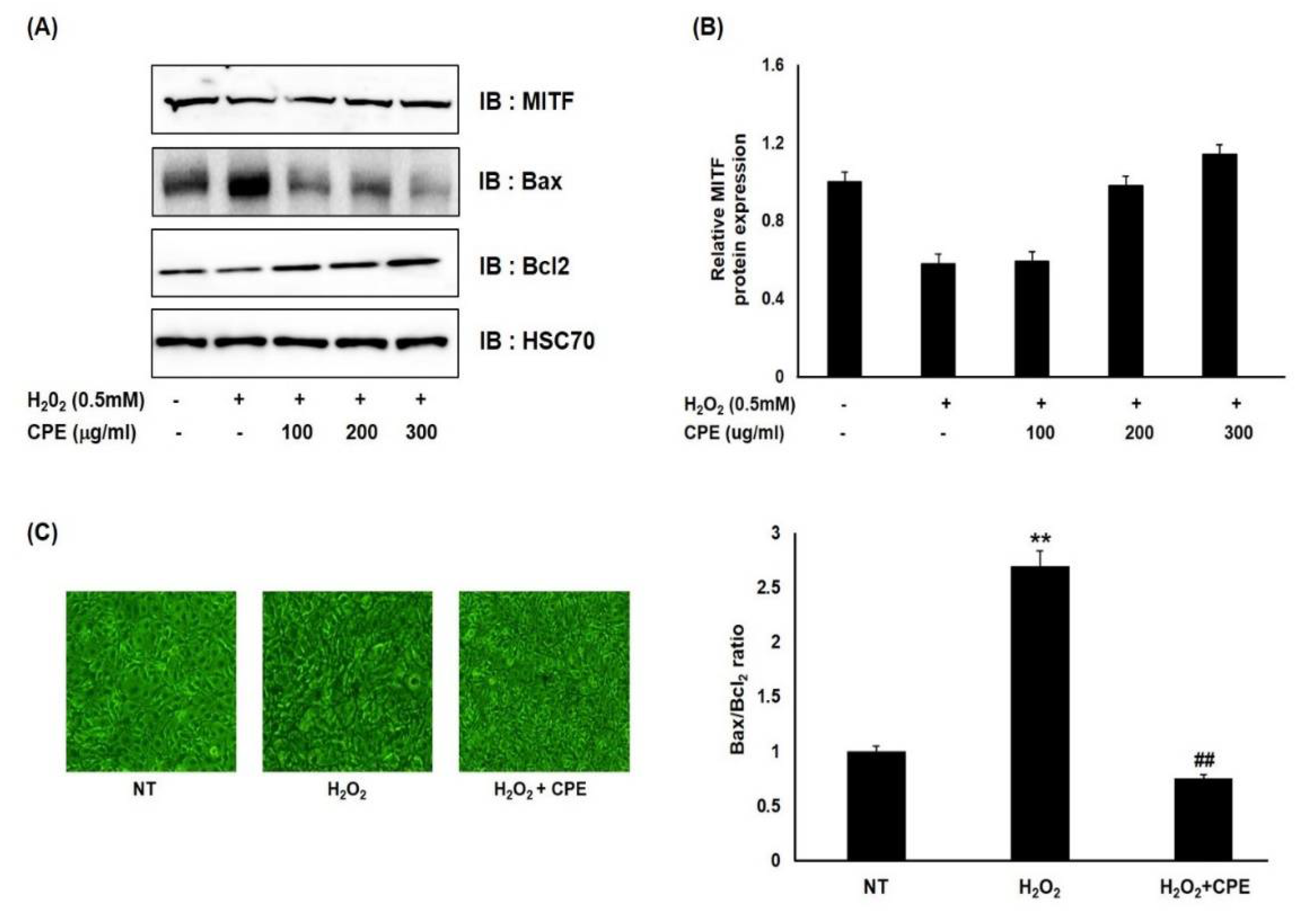
3.2. Effect of CPE on H2O2-Induced Cell Death in Melanocytes
3.3. Effect of CPE on H2O2-Induced Cell Death in Melanocytes
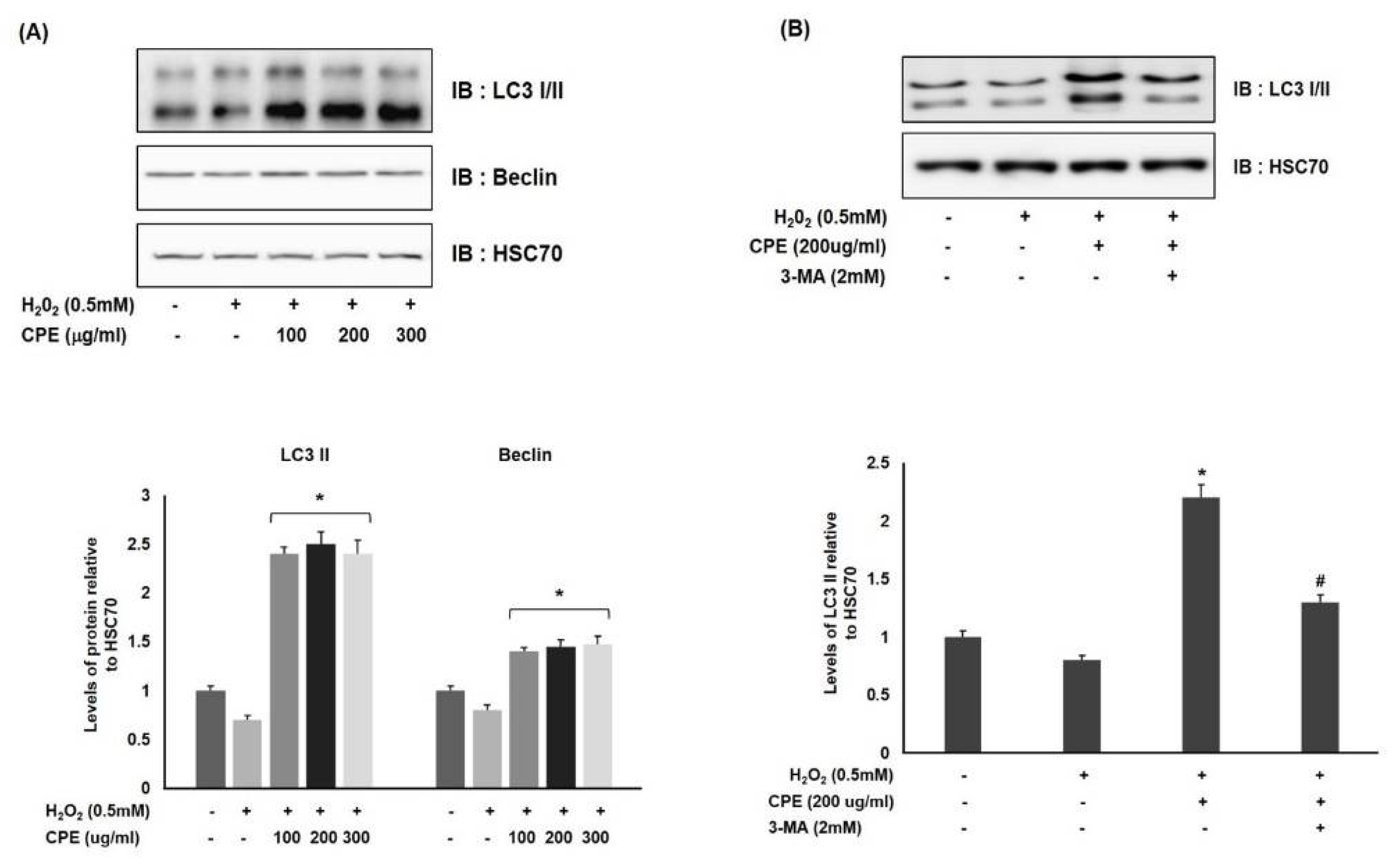
3.4. Effect of CPE on Autophagy Activation in Oxidative-Stress-Induced Melanocytes
4. Discussion
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ichihashi, M.; Ueda, M.; Budiyanto, A.; Bito, T.; Oka, M.; Fukunage, M.; Tsuru, K.; Horikawa, T. UV-induced skin damage. Toxicology 2003, 189, 21–39. [Google Scholar] [CrossRef]
- Lukiewicz, S. The biological role of melanin. I. New concepts and methodical approaches. Folia Histochem. Cytochem. (Krakow) 1972, 10, 93–108. [Google Scholar]
- Hearing, V.J. Biogenesis of pigment granules: A sensitive way to regulate melanocyte function. J. Dermatol. Sci. 2005, 37, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, A.K.; Ho, H.; Bodemann, B.; Petersen, S.; Aruri, J.; Koshy, S.; White, M.A. Genome-wide siRNA-based functional genomics of pigmentation identifies novel genes and pathways that impact melanogenesis in human cells. PLoS Genet. 2008, 4, e1000298. [Google Scholar] [CrossRef] [PubMed]
- Ko, M.S.; Lee, H.J.; Kang, M.J. Antioxidant activities and whitening effects of extracts from Hippophae rhamnoides L. J. East Asian Soc. Diet Life 2012, 22, 812–817. [Google Scholar]
- Ahn, J.H.; Jin, S.H.; Kang, H.Y. LPS induces melanogenesis through p38 MAPK activation in human melanocytes. Arch. Dermatol. Res. 2008, 300, 325–329. [Google Scholar] [CrossRef]
- Navone, F.; Genevini, P.; Borgese, N. Autophagy and neurodegeneration: Insights from a cultured cell model of ALS. Cells 2015, 4, 354–386. [Google Scholar] [CrossRef]
- Zhou, Y.Y.; Li, Y.; Jiang, W.Q.; Zhou, L.F. MAPK/JNK signalling: A potential autophagy regulation pathway. Biosci. Rep. 2015, 35, e00199. [Google Scholar] [CrossRef]
- Raposo, G.; Marks, M.S. Melanosomes—Dark organelles enlighten endosomal membrane transport. Nat. Rev. Mol. Cell Biol. 2007, 8, 786–797. [Google Scholar] [CrossRef]
- Ho, H.; Ganesan, A.K. The pleiotropic roles of autophagy regulators in melanogenesis. Pigment Cell Melanoma Res. 2011, 24, 595–604. [Google Scholar] [CrossRef]
- Rubinsztein, D.C.; Gestwicki, J.E.; Murphy, L.O.; Klionsky, D.J. Potential therapeutic applications of autophagy. Nat. Rev. Drug Discov. 2007, 6, 304–312. [Google Scholar] [CrossRef]
- Hu, X.Y.; Qin, F.Y.; Lu, X.F.; Zhang, L.S.; Cheng, Y.X. Three new polyynes from Codonopsis pilosula and their activities on lipid metabolism. Molecules 2018, 23, 887. [Google Scholar] [CrossRef]
- He, J.Y.; Ma, N.; Zhu, S.; Komatsu, K.; Li, Z.Y.; Fu, W.M. The genus Codonopsis (Campanulaceae): A review of phytochemistry, bioactivity and quality control. J. Nat. Med. 2015, 69, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Tong, X.; Li, P.; Liu, M.; Peng, W.; Cao, H.; Su, W. Bioactive components on immuno-enhancement effects in the traditional Chinese medicine Shenqi Fuzheng injection based on relevance analysis between chemical HPLC fingerprints and in vivo biological effects. J. Ethnopharmacol. 2014, 155, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.S.; Jung, K.J.; Lee, K.S.; Lee, H.S.; Park, Y.K. Effects of the ethanol extract of Codonopsis Pilosula Radix on ovalbumin-induced allergic responses in mice. Korea J. Herbol. 2013, 28, 9–15. [Google Scholar] [CrossRef][Green Version]
- He, K.; Li, X.; Chen, X.; Ye, X.; Huang, J.; Li, Y.; Deng, Y.; Jin, Q.; Shi, Q.; Shu, H. Evaluation of antidiabetic potential of selected traditional Chinese medicines in STZ-induced diabetic mice. J. Ethnophamacol. 2011, 137, 1135–1142. [Google Scholar] [CrossRef]
- Ng, T.B.; Liu, F.; Wang, H.X. The antioxidant effects of aqueous and organic extracts of panax quinquefolium, panax notoginseng, Codonopsis pilosula, Pseudostellaria heterophylla and Glehnia littoralis. J. Enthnopharmacol. 2004, 93, 285–288. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Ahn, S.H.; Choi, W.Y.; Lee, G.W.; Kwon, M.J.; Cho, Y.H. Changes in whitening activity of Codonopsis pilosula extracts according to extraction solvents. KSBB J. 2019, 34, 1–9. [Google Scholar] [CrossRef]
- Park, H.J. Anti-inflammatory mechanisms of gamma-irradiated fibroin protein. Master’s Thesis, Konyang University, Daejeon, Korea, 2020. [Google Scholar]
- Xin, Q.; Kwon, M.J.; Lee, J.W.; Kim, K.S.; Chen, H.; Campos, M.G.; Tundis, R.; Cui, C.B.; Cho, Y.H.; Cao, H. Gamma irradiated Rhodiola sachalinensis extract ameliorates testosterone-induced benign prostatic hyperplasia by downregulating 5-alpha reductase and restoring testosterone in rats. Molecules 2019, 24, 3981. [Google Scholar] [CrossRef]
- Kim, H.; Xue, X. Detection of total reactive oxygen species in adherent cells by 2′,7′-dichlorodihydrofluorescein diacetate staining. J. Vis. Exp. 2020, 160. [Google Scholar] [CrossRef] [PubMed]
- Murase, D.; Hachiya, A.; Amano, Y.; Ohuchi, A.; Kitahara, T.; Takema, Y. The essential role of p53 in hyperpigmentation of the skin via regulation of paracrine melanogenic cytokine receptor signaling. J. Biol. Chem. 2009, 284, 4343–4353. [Google Scholar] [CrossRef]
- Niwano, T.; Terazawa, S.; Nakajima, H.; Wakabayashi, Y.; Imokawa, G. Astaxanthin and withaferin a block paracrine cytokine interactions between uvb-exposed human keratinocytes and human melanocytes via the attenuation of endothelin-1 secretion and its downstream intracellular signaling. Cytokine 2015, 73, 184–197. [Google Scholar] [CrossRef] [PubMed]
- Trachootham, D.; Lu, W.; Ogasawara, M.A.; Valle, N.R.-D.; Huang, P. Redox regulation of cell survival. Antioxid. Redox Signal 2008, 10, 1343–1374. [Google Scholar] [CrossRef]
- Halliwell, B. Free radicals and antioxidants—Quo vadis? Trends Pharmacol. Sci. 2011, 32, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Z.; Xu, Z.; Xiao, Q.; Yang, Y.; Ying, J.; Xiang, L.; Zhang, C. Dysfunction of ATG7-dependent autophagy dysregulates the antioxidant response and contributes to oxidative stress-induced biological impairments in human epidermal melanocytes. Cell Death Discov. 2020, 6, 31. [Google Scholar] [CrossRef] [PubMed]
- Korsmeyer, S.J.; Shuttler, J.R.; Veis, D.J. Bcl2/Bax: A rheostat that regulates an anti-oxidant pathway and cell death. Semin. Cancer Biol. 1993, 4, 327–332. [Google Scholar]
- Khodapasand, E.; Jafarzadeh, N.; Farrokhi, F.; Kamalidehghan, B.; Houshmand, M. Is Bax/Bcl-2 ratio considered as a prognostic marker with age and tumor location in colorectal cancer? Iran. Biomed. J. 2015, 19, 69–75. [Google Scholar] [PubMed]
- Larsen, C.J. The Bcl2 gene is the prototype of a gene family that controls programmed cell death (apoptosis). Ann. Genet. 1994, 37, 121–134. [Google Scholar]
- Emanuele, S.; Notaro, A.; Piccionello, A.P.; Maggio, A.; Lauricella, M.; D’Anneo, A.; Cernigliaro, C.; Calvaruso, G.; Giuliano, M. Sicilian litchi fruit extracts induce autophagy versus apoptosis switch in human colon cancer cells. Nutrients 2018, 10, 1490. [Google Scholar] [CrossRef]
- Feng, Y.; Jiang, C.; Yang, F.; Chen, Z.; Li, Z. Apocynum venetum leaf extract protects against H2O2-induced oxidative stress by increasing autophagy in PC12 cells. Biomed. Rep. 2020, 6. [Google Scholar] [CrossRef]
- Han, J.S.; Sung, J.H.; Lee, S.K. Anti-melanogenesis activity of hydrolyzed ginseng extract (GINST) via inhibition of JNK mitogen-activated protein kinase in B16F10 cells. J. Food Sci. 2016, 81, H2085–H2092. [Google Scholar] [CrossRef]
- Haq, R.; Shoag, J.; Andreu-Perez, P.; Yokoyama, S.; Edelman, H.; Rowe, G.C.; Frederick, D.T.; Hurley, A.D.; Nellore, A.; Kung, A.L.; et al. Oncogenic BRAF regulates oxidative metabolism via PGC1α and MITF. Cancer Cell 2013, 23, 302–315. [Google Scholar] [CrossRef]
- Garraway, L.A.; Widlund, H.R.; Rubin, M.A.; Getz, G.; Berger, A.J.; Ramaswamy, S.; Beroukhim, R.; Milner, D.A.; Granter, S.R.; Du, J.; et al. Integrative genomic analyses identify MITF as a lineage survival oncogene amplified in malignant melanoma. Nature 2005, 436, 117–122. [Google Scholar] [CrossRef]
- Wu, M.; Hemesath, T.J.; Takemoto, C.M.; Horstmann, M.A.; Wells, A.G.; Price, E.R.; Fisher, D.Z.; Fisher, D.E. c-Kit triggers dual phosphorylations, which couple activation and degradation of the essential melanocyte factor Mi. Genes Dev. 2000, 4, 301–312. [Google Scholar]
- Kim, J.H.; Hong, A.R.; Kim, Y.H.; Yoo, H.J.; Kang, S.W.; Chang, S.E.; Song, Y.S. JNK suppresses melanogenesis by interfering with CREB-regulated transcription coactivator 3-dependent MITF expression. Theranostics 2020, 10, 4017–4029. [Google Scholar] [CrossRef]
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin pigmentation in mammalian skin and its hormonal regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [CrossRef] [PubMed]
- D’Orazio, J.; Jarrett, S.; Amar-Ortiz, A.; Scott, T. UV radiation and the skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [PubMed]
- Meyskens, F.L., Jr.; Farmer, P.; Fruehauf, J.P. Redox regulation in human melanocytes and melanoma. Pigment Cell Res. 2001, 14, 148–154. [Google Scholar] [CrossRef]
- Hoek, K.S.; Schlegel, N.C.; Eichhoff, O.M.; Widmer, D.S.; Praetorius, C.; Einarsson, S.O.; Valgeirsdottir, S.; Bergsteinsdottir, K.; Schepsky, A.; Dummer, R.; et al. Novel MITF targets identified using a two-step DNA microarray strategy. Pigment Cell Melanoma Res. 2008, 21, 665–676. [Google Scholar] [CrossRef] [PubMed]
- Strub, T.; Giuliano, S.; Ye, T.; Bonet, C.; Keime, C.; Kobi, D.; Gras, S.L.; Cormont, M.; Ballotti, R.; Bertolotto, C.; et al. Essential role of microphthalmia transcription factor for DNA replication, mitosis and genomic stability in melanoma. Oncogene 2011, 30, 2319–2332. [Google Scholar] [CrossRef]
- Kawakami, A.; Fisher, D. The master role of microphthalmia-associated transcription factor in melanocyte and melanoma biology. Lab. Investig. 2017, 97, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Steingrímsson, E.; Moore, K.J.; Lamoreux, M.L.; Ferré-D’Amaré, A.R.; Burley, S.K.; Sanders Zimring, D.C.; Skow, L.C.; Hodgkinson, C.A.; Arnheiter, H.; Copeland, N.G.; et al. Molecular basis of mouse microphthalmia (mi) mutations helps explain their developmental and phenotypic consequences. Nat. Genet. 1994, 8, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Liu, J.; Chen, D.; Kang, J.; Huang, Y.; Li, D.; Duan, Y.; Wang, J. Integration of lncRNA and mRNA profiles to reveal the protective effects of Codonopsis pilosula extract on the gastrointestinal tract of mice subjected to D-galactose-induced aging. Int. J. Mol. Med. 2021, 47, 1. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N.; Komatsu, M. Autophagy: Renovation of cells and tissues. Cell 2011, 147, 728–741. [Google Scholar] [CrossRef] [PubMed]
- Kabeya, Y.; Mizushima, N.; Ueno, T.; Yamamoto, A.; Kirisako, T.; Noda, T.; Kominami, E.; Ohsumi, Y.; Yoshimori, T. LC3, a mammalian homologue of yeast Apg8p, is localized in autophagosome membranes after processing. EMBO J. 2000, 19, 5720–5728. [Google Scholar] [CrossRef]
- Chifenti, B.; Locci, M.T.; Lazzeri, G.; Guagnozzi, M.; Dinucci, D.; Chiellini, F.; Filice, M.E.; Salerno, M.G.; Battini, L. Autophagy-related protein LC3 and Beclin-1 in the first trimester of pregnancy. Clin. Exp. Reprod. Med. 2013, 40, 33–37. [Google Scholar] [CrossRef]
- Zhang, C.F.; Gruber, F.; Ni, C.; Mildner, M.; Koenig, U.; Karner, S.; Barresi, C.; Rossiter, H.; Narzt, M.S.; Nagelreiter, I.M.; et al. Suppression of autophagy dysregulates the antioxidant response and causes premature senescence of melanocytes. J. Investig. Dermatol. 2015, 135, 1348–1357. [Google Scholar] [CrossRef] [PubMed]
- Eskelinen, E.L.; Saftig, P. Autophagy: A lysosomal degradation pathway with a central role in health and disease. Biochim. Biophys. Acta 2009, 1793, 664–673. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, Y.H. Codonopsis pilosula Extract Protects Melanocytes against H2O2-Induced Oxidative Stress by Activating Autophagy. Cosmetics 2021, 8, 67. https://doi.org/10.3390/cosmetics8030067
Cho YH. Codonopsis pilosula Extract Protects Melanocytes against H2O2-Induced Oxidative Stress by Activating Autophagy. Cosmetics. 2021; 8(3):67. https://doi.org/10.3390/cosmetics8030067
Chicago/Turabian StyleCho, Young Ho. 2021. "Codonopsis pilosula Extract Protects Melanocytes against H2O2-Induced Oxidative Stress by Activating Autophagy" Cosmetics 8, no. 3: 67. https://doi.org/10.3390/cosmetics8030067
APA StyleCho, Y. H. (2021). Codonopsis pilosula Extract Protects Melanocytes against H2O2-Induced Oxidative Stress by Activating Autophagy. Cosmetics, 8(3), 67. https://doi.org/10.3390/cosmetics8030067