Natural Flavones and Flavonols: Relationships among Antioxidant Activity, Glycation, and Metalloproteinase Inhibition


 and
and
Abstract
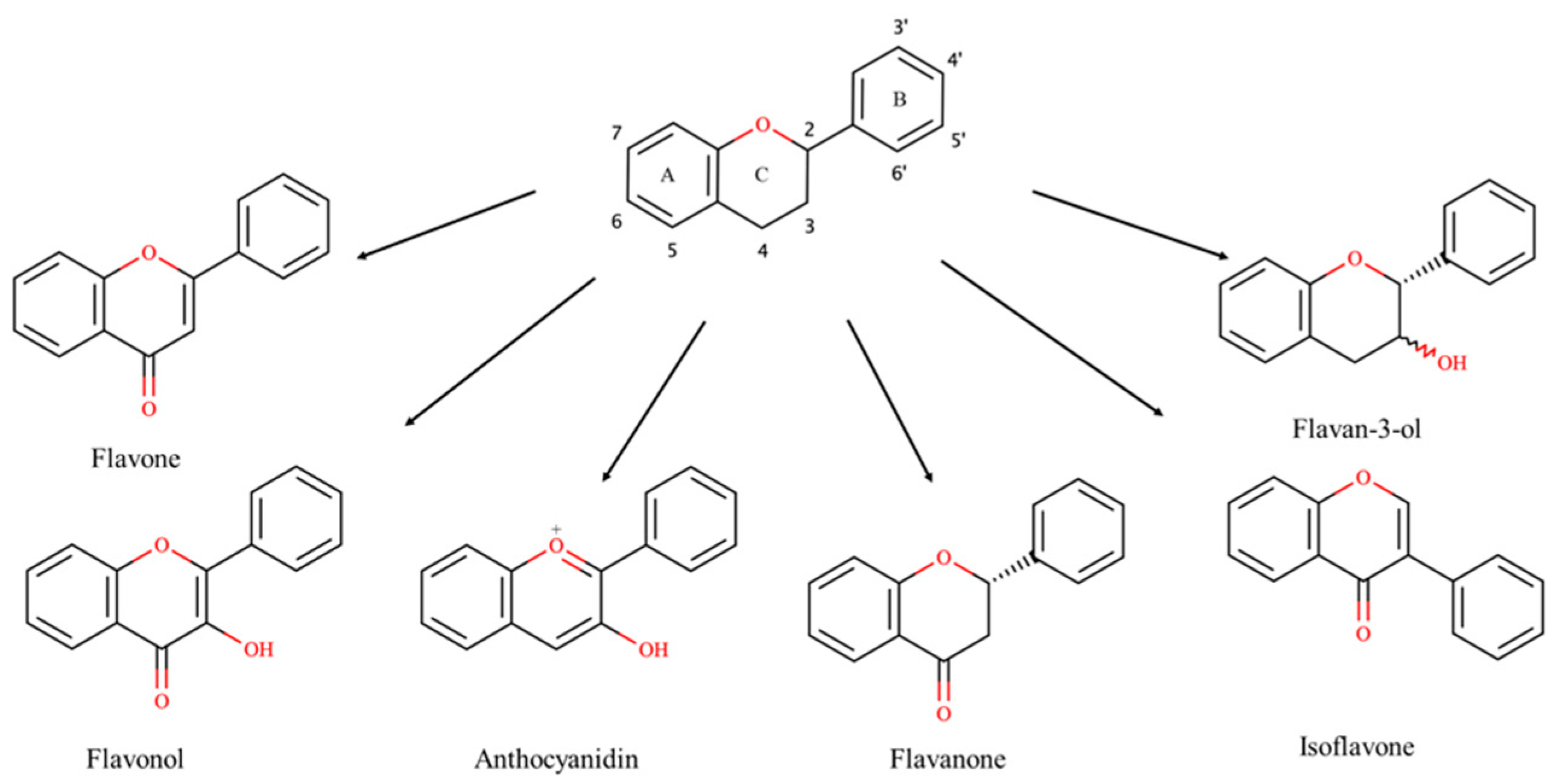
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Oxygen Radical Absorbance Capacity (Orac) Assay
2.3. NO Scavenger Assay
2.4. Anti-glycation Activity
2.5. MMP Inhibition Assay
2.6. QSAR Analysis
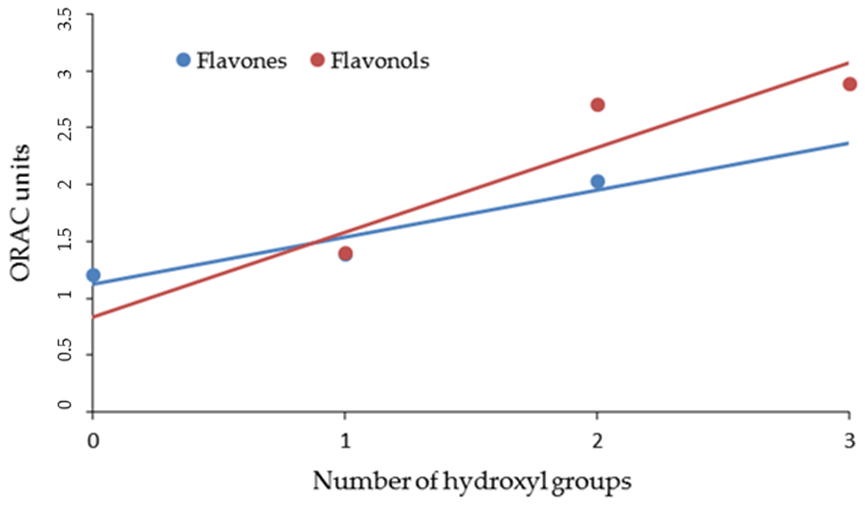
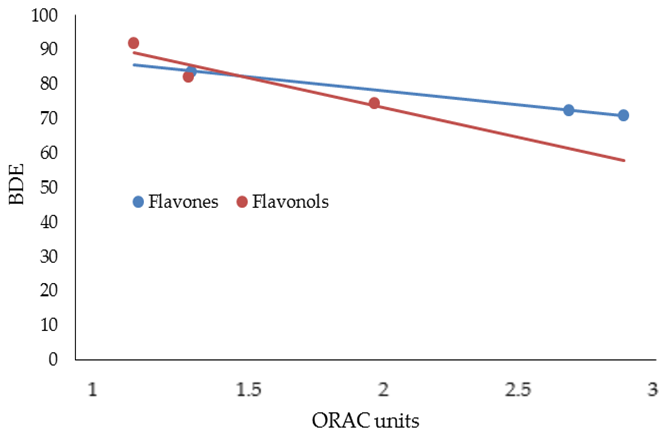
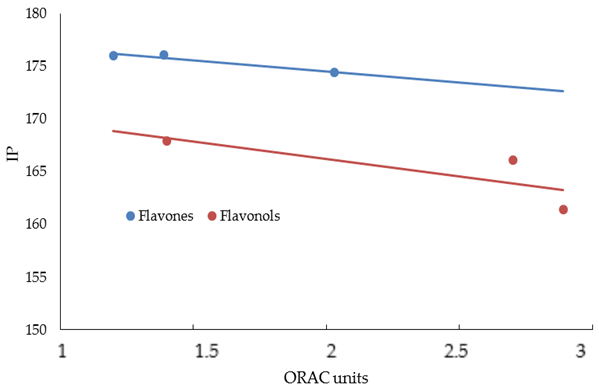
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kim, C.S.; Park, S.; Kim, J. The role of glycation in the pathogenesis of aging and its prevention through herbal products and physical exercise. J. Exerc. Nutr. Biochem. 2017, 21, 55–61. [Google Scholar] [CrossRef]
- Basta, G.; Schmidt, A.M.; De Caterina, R. Advanced glycation end products and vascular inflammation: Implications for accelerated atherosclerosisin diabetes. Cardiovasc. Res. 2004, 63, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Percik, R.; Stumvoll, M. Obesity and cancer. Exp. Clin. Endocr. Diab. 2009, 117, 563–566. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, P.J.; Carrington, A.L.; Frost, G.S.; Boulton, A.J. Neurovascular disease, antioxidants and glycation in diabetes. Diabetes Metab. Res. 2002, 18, 260–272. [Google Scholar] [CrossRef] [PubMed]
- Crascì, L.; Lauro, M.R.; Puglisi, G.; Panico, A.M. Natural antioxidant polyphenols on inflammation management: Anti-glycation activity vs metalloproteinases inhibition. Crit. Rev. Food Sci. 2018, 58, 893–904. [Google Scholar] [CrossRef]
- Sparvero, L.J.; Asafu-Adjei, D.; Kang, R.; Tang, D.; Amin, N.; Im, J.; Rutledge, R.; Lin, B.; Amoscato, A.A.; Zeh, H.J.; et al. RAGE (Receptor for Advanced Glycation Endproducts), RAGE ligands, and their role in cancer and inflammation. J. Transl. Med. 2009, 7, 1–17. [Google Scholar] [CrossRef]
- Panico, A.M.; Vicini, P.; Geronikaki, A.; Incerti, M.; Cardile, V.; Crascì, L.; Messina, R.; Ronsisvalle, S. Heteroarylimino-4-thiazolidinonesas inhibitors of cartilage degradation. Bioorgan. Chem. 2011, 39, 48–52. [Google Scholar] [CrossRef]
- Raeeszadeh-Sarmazdeh, M.; Do, L.D.; Hritz, B.G. Metalloproteinases and their inhibitors: Potential for the development of new therapeutics. Cells 2020, 9, 1313. [Google Scholar] [CrossRef]
- Levin, M.; Udi, Y.; Solomonov, I.; Sagi, I. Next generation matrix metalloproteinase inhibitors—Novel strategies bring new prospects. Biochim. Biophys. Acta 2017, 1864, 1927–1939. [Google Scholar] [CrossRef]
- Fields, G.B. The rebirth of matrix metalloproteinase inhibitors: Moving beyond the dogma. Cells 2019, 8, 984. [Google Scholar] [CrossRef]
- Lauro, M.R.; Crascì, L.; Carbone, C.; Aquino, R.P.; Panico, A.M.; Puglisi, G. Encapsulation of a citrus by-product extract: Development, characterization and stability studies of a nutraceutical with antioxidant and metalloproteinases inhibitory activity. LWT Food Sci Technol. 2015, 62, 169–176. [Google Scholar] [CrossRef]
- Pietta, P.G. Flavonoids as antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Galano, A. Free radicals induced oxidative stress at a molecular level: The current status, challenges and perspectives of computational chemistry based protocols. J. Mex. Chem. Soc. 2015, 59, 231–262. [Google Scholar] [CrossRef]
- Trouillas, P.; Marsal, P.; Siri, D.; Lazzaroni, R.; Duroux, J.L. A DFT study of the reactivity of OH groups in quercetin and taxifolin antioxidants: The specificity of the 3-OH site. Food Chem. 2006, 97, 679–688. [Google Scholar] [CrossRef]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef]
- Leopoldini, M.; Marino, T.; Russo, N.; Toscano, M. Antioxidant properties of phenolic compounds: H-atom versus electron transfer mechanism. J. Phys. Chem. A 2004, 108, 4916–4922. [Google Scholar] [CrossRef]
- Leopoldini, M.; Pitarch, I.P.; Russo, N.; Toscano, M. Structure, conformation, and electronic properties of apigenin, luteolin, and taxifolin antioxidants. A first principle theoretical study. J. Phys. Chem. A 2004, 108, 92–96. [Google Scholar] [CrossRef]
- Leopoldini, M.; Chiodo, S.G.; Russo, N.; Toscano, M. Detailed investigation of the OH radical quenching by natural antioxidant caffeic acid studied by quantum mechanical models. J. Chem. Theory Comput. 2011, 7, 4218–4233. [Google Scholar] [CrossRef]
- Moalin, M.; Van Strijdonck, G.P.; Beckers, M.; Hagemen, G.; Borm, P.; Bast, A.; Haenen, G.R. A planar conformation and the hydroxyl groups in the B and C rings play a pivotal role in the antioxidant capacity of quercetin and quercetin derivatives. Molecules 2011, 16, 9636–9650. [Google Scholar] [CrossRef]
- Jeong, J.M.; Choi, C.H.; Kang, S.K.; Lee, I.H.; Lee, J.Y.; Jung, H. Antioxidant and chemosensitizing effects of flavonoids with hydroxy and/or methoxy groups and structure-activity relationship. J. Pharm. Pharm. Sci. 2007, 10, 537–546. [Google Scholar] [CrossRef]
- Treml, J.; Smejkal, K. Flavonoids as potent scavengers of hydroxyl radicals. Compr. Rev. Food Sci. F 2016, 15, 720–738. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [PubMed]
- Xie, P.; Huang, L.; Zhang, C.; Zhang, Y. Phenolic compositions, and antioxidant performance of olive leaf and fruit (Olea europaea L.) extracts and their structure–activity relationships. J. Funct. Foods 2015, 16, 460–471. [Google Scholar] [CrossRef]
- Rafat Husain, S.; Cillard, J.; Cillard, P. Hydroxyl radical scavenging activity of flavonoids. Phytochemistry 1987, 26, 2489–2491. [Google Scholar] [CrossRef]
- Eto, M.; Masuoka, C.; Yamasaski, T.; Harano, K.; Ono, M. Molecular orbital analysis of antioxidative activity of phenolics from Tessaria integrifolia and Piper elongatum. Food Sci. Technol. Res. 2008, 14, 415–420. [Google Scholar] [CrossRef][Green Version]
- Wu, C.H.; Yen, G.C. Inhibitory effect of naturally occurring flavonoids on the formation of advanced glycation endproducts. J. Agric. Food Chem. 2005, 53, 3167–3173. [Google Scholar] [CrossRef] [PubMed]
- Ramkissoon, J.S.; Mahomoodally, M.F.; Ahmed, N.; Subratty, A.H. Antioxidant and anti-glycation activities correlates with phenolic composition of tropical medicinal herbs. Asian Pac. J. Trop. Med. 2013, 6, 561–569. [Google Scholar] [CrossRef]
- Sim, G.S.; Lee, B.C.; Cho, H.S.; Lee, J.W.; Kim, J.H.; Lee, D.H.; Kim, J.H.; Pyo, H.B.; Moon, D.C.; Oh, K.W.; et al. Structure activity relationship of antioxidative property of flavonoids and inhibitory effect on matrix metalloproteinase activity in UVA-irradiated human dermal fibroblast. Arch. Pharm. Res. 2007, 30, 290–298. [Google Scholar] [CrossRef]
- Ende, C.; Gebhardt, R. Inhibition of matrix metalloproteinase-2 and -9 activities by selected flavonoids. Planta Med. 2004, 70, 1006–1008. [Google Scholar] [CrossRef]
- Crascì, L.; Basile, L.; Panico, A.; Puglia, C.; Bonina, F.P.; Basile, P.M.; Rizza, L.; Guccione, S. Correlating in vitro target-oriented screening and docking: Inhibition of matrix metalloproteinases activities by flavonoids. Planta Med. 2017, 83, 901–911. [Google Scholar] [CrossRef]
- Montenegro, L.; Panico, A.; Santagati, L.; Siciliano, E.A.; Intagliata, S.; Modica, M. Solid lipid nanoparticles loading idebenone ester with pyroglutamic acid: In vitro antioxidant activity and in vivo topical efficacy. Nanomaterials 2019, 9, 43. [Google Scholar] [CrossRef] [PubMed]
- Crascì, L.; Cardile, V.; Longhitano, G.; Nanfitò, F.; Panico, A. Anti-degenerative effect of Apigenin, Luteolin and Quercetin on human keratinocyte and chondrocyte cultures: SAR evaluation. Drug Res. 2018, 68, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Derbré, S.; Gatto, J.; Pelleray, A.; Coulon, L.; Séraphin, D.; Richomme, P. Automating a 96-well microtiter plate assay for identification of AGEs inhibitors or inducers: Application to the screening of a small natural compounds library. Anal. Bioanal. Chem. 2010, 398, 1747–1758. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Rafter, J.; Jenner, A. Health promotion by flavonoids, tocopherols, tocotrienols, and other phenols: Direct or indirect effects? Antioxidant or not? Am. J. Clin. Nutr. 2005, 81, 268S–276S. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.F.; Ramasamym, R.; Schmidtm, A.M. Mechanisms of disease: Advanced glycation end-products and their receptor in inflammation and diabetes complications. Nat. Clin. Pract. Endocrinol. Metab. 2008, 4, 285–293. [Google Scholar] [CrossRef]
- Komosinska-Vassev, K.; Olczyk, P.; Winsz-Szczotka, K.; Klimek, K.; Olczyk, K. Plasma biomarkers of oxidative and AGE-mediated damage of proteins and glycosaminoglycans during healthy ageing: A possible association with ECM metabolism. Mech. Ageing Dev. 2012, 133, 538–548. [Google Scholar] [CrossRef]
- Nagai, R.; Murray, D.B.; Metz, T.O.; Baynes, J.W. Chelation: A fundamental mechanism of action of AGE inhibitors, AGE breakers, and other inhibitors of diabetes complications. Diabetes 2012, 61, 549–559. [Google Scholar] [CrossRef]
- Verma, R.P.; Hansch, C. Matrix metalloproteinases (MMPs): Chemical–biological functions and (Q) SARs. Bioorg. Med. Chem. 2007, 15, 2223–2268. [Google Scholar] [CrossRef]
- Kameda, K.; Matsunaga, T.; Abe, N.; Hanada, H.; Ishizaka, H.; Ono, H.; Saitoh, M.; Fukui, K.; Fukuda, I.; Osanai, T.; et al. Correlation of oxidative stress with activity of matrix metalloproteinase in patients with coronary artery disease. Eur. Heart J. 2003, 24, 2180–2185. [Google Scholar] [CrossRef]
- Oh, S.H.; Lim, S.C. A rapid and transient ROS generation by cadmium triggers apoptosis via caspase-dependent pathway in HepG2 cells and this is inhibited through N-acetylcysteine-mediated catalase upregulation. Toxicol. Appl. Pharmacol. 2006, 212, 212–223. [Google Scholar] [CrossRef]
- Kar, S.; Subbaram, S.; Carrico, P.M.; Melendez, J.A. Redox-control of matrix metalloproteinase-1: A critical link between free radicals, matrix remodeling and degenerative disease. Respir. Physiol. Neurobiol. 2010, 174, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Goldring, M.B.; Otero, M. Inflammation in osteoarthritis. Curr. Opin. Rheumatol. 2011, 23, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Aljada, A.; Ghanim, H.; Mohanty, P.; Syed, T.; Bandyopadhyay, A.; Dandona, P. Glucose intake induces an increase in activator protein 1 and early growth response 1 binding activities, in the expression of tissue factor and matrix metalloproteinase in mononuclear cells, and in plasma tissue factor and matrix metalloproteinase concentrations. Am. J. Clin. Nutr. 2004, 80, 51–57. [Google Scholar] [CrossRef]
- Amić, D.; Davidović-Amić, D.; Beslo, D.; Rastija, V.; Lucić, B.; Trinajstić, N. SAR and QSAR of the antioxidant activity of flavonoids. Curr. Med. Chem. 2007, 14, 827–845. [Google Scholar] [CrossRef]
- Takano-Ishikawa, Y.; Goto, M.; Yamaki, K. Structure–activity relations of inhibitory effects of various flavonoids on lipopolysaccharide-induced prostaglandin E 2 production in rat peritoneal macrophages: Comparison between subclasses of flavonoids. Phytomedicine 2006, 13, 310–317. [Google Scholar] [CrossRef]
- Nagula, R.L.; Wairkar, S. Recent advances in topical delivery of flavonoids: A review. J. Control. Release 2019, 296, 190–201. [Google Scholar] [CrossRef]
- Montenegro, L. Lipid-based nanoparticles as carriers for dermal delivery of antioxidants. Curr. Drug Metab. 2017, 18, 469–480. [Google Scholar] [CrossRef]
- Intagliata, S.; Modica, M.N.; Santagati, L.M.; Montenegro, L. Strategies to improve resveratrol systemic and topical bioavailability: An update. Antioxidants 2019, 8, 244. [Google Scholar] [CrossRef] [PubMed]
- Bilia, A.; Isacchi, B.; Righeschi, C.; Guccione, C.; Bergonzi, M. Flavonoids loaded in nanocarriers: An opportunity to increase oral bioavailability and bioefficacy. Food Nutr. Sci. 2014, 5, 1212–1227. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
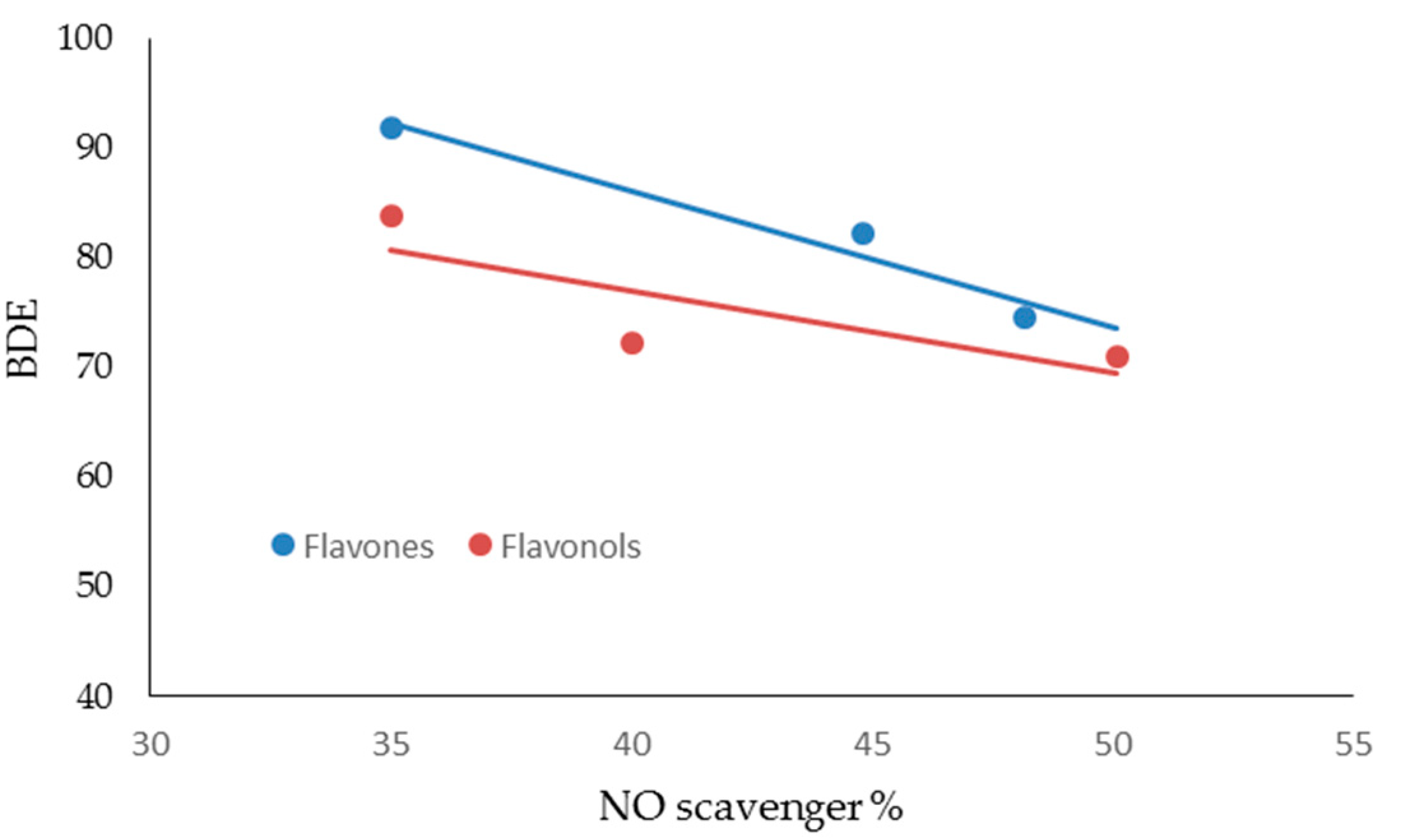
| FLAVONES | ORAC Units (µM) | NO Scavenger (%) | AGEs Inhibition (%) | MMP-1 (IC50) | MMP-13 (IC50) | MMP-2 (IC50) | MMP-9 (IC50) |
|---|---|---|---|---|---|---|---|
| LUTEOLIN | 2.03 ± 0.11 | 48.19 ± 0.18 | 60.04 ± 0.10 | 3.0 ± 0.11 | 1.0 ± 0.14 | 20.0 ±0.13 | 22.0 ± 0.23 |
| APIGENIN | 1.39 ± 0.13 | 44.81 ± 0.26 | 59.10 ± 0.12 | 6.0 ± 0.13 | 3.0 ± 0.12 | 16.0 ± 0.11 | 18.0 ± 0.15 |
| CHRYSIN | 1.20 ± 0.10 | 35.00 ± 0.28 | 45.08 ± 0.10 | 7.0 ± 0.22 | 10.0 ± 0.24 | 33.0 ± 0.18 | 31.0 ± 0.17 |
| FLAVONOLS | ORAC Units (µM) | NO Scavenger (%) | AGEs inhibition (%) | MMP-1 (IC50) | MMP-13 (IC50) | MMP-2 (IC50) | MMP-9 (IC50) |
|---|---|---|---|---|---|---|---|
| MIRYCETIN | 2.89 ± 0.09 | 50.10 ± 0.18 | 58.04 ± 0.11 | 3.0 ± 0.11 | 2.00 ± 0.13 | 17.00 ± 0.23 | 19.00 ± 0.31 |
| QUERCETIN | 2.70 ± 0.10 | 40.01 ± 0.09 | 52.04 ± 0.12 | 1.0 ± 0.16 | 3.00 ± 0.17 | 12.00 ± 0.19 | 22.00 ± 0.16 |
| KAEMPFEROL | 1.40 ± 0.11 | 35.00 ± 0.26 | 38.02 ± 0.17 | 4.0 ± 0.14 | 2.00 ± 0.15 | 24.00 ± 0.21 | 27.00 ± 0.23 |
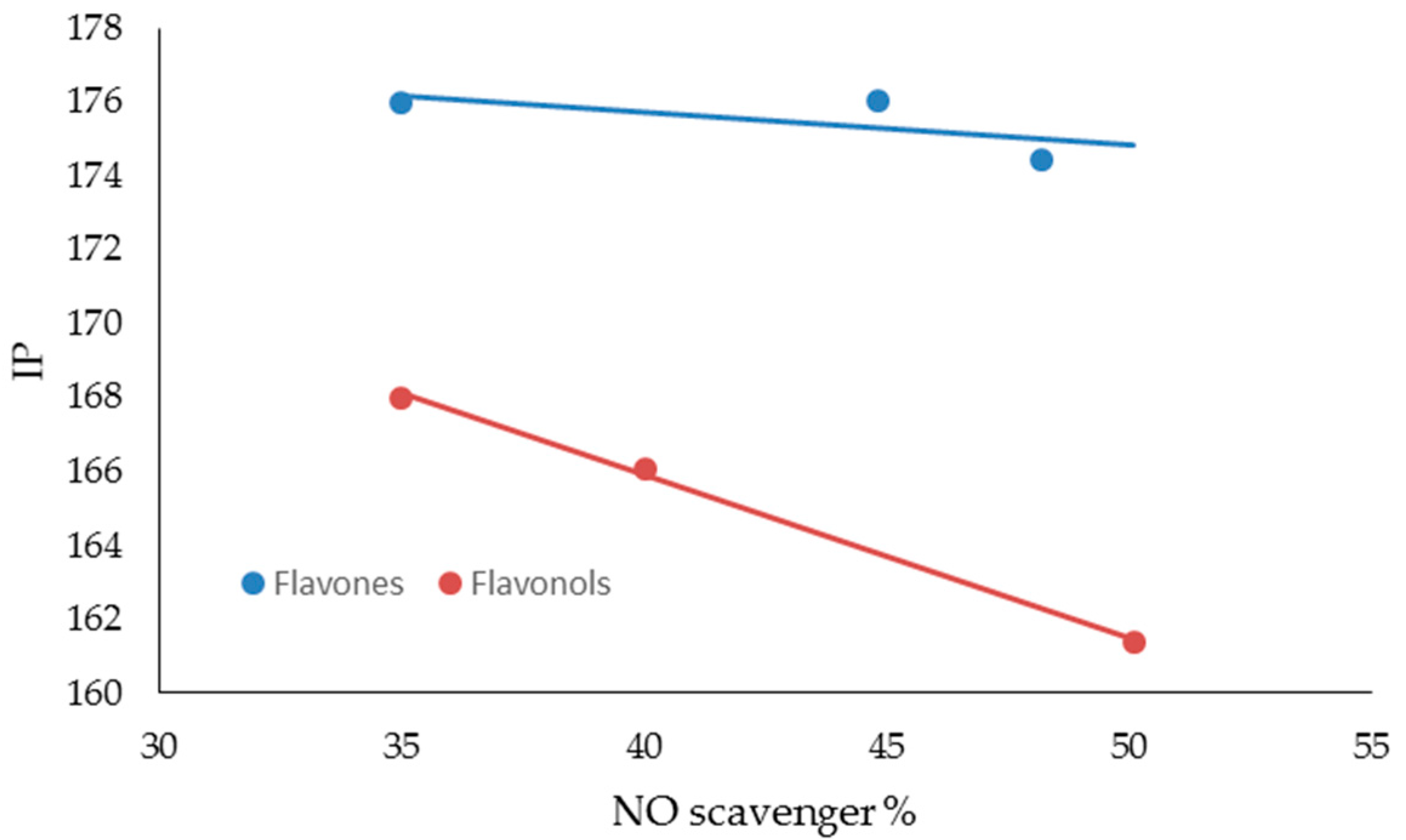
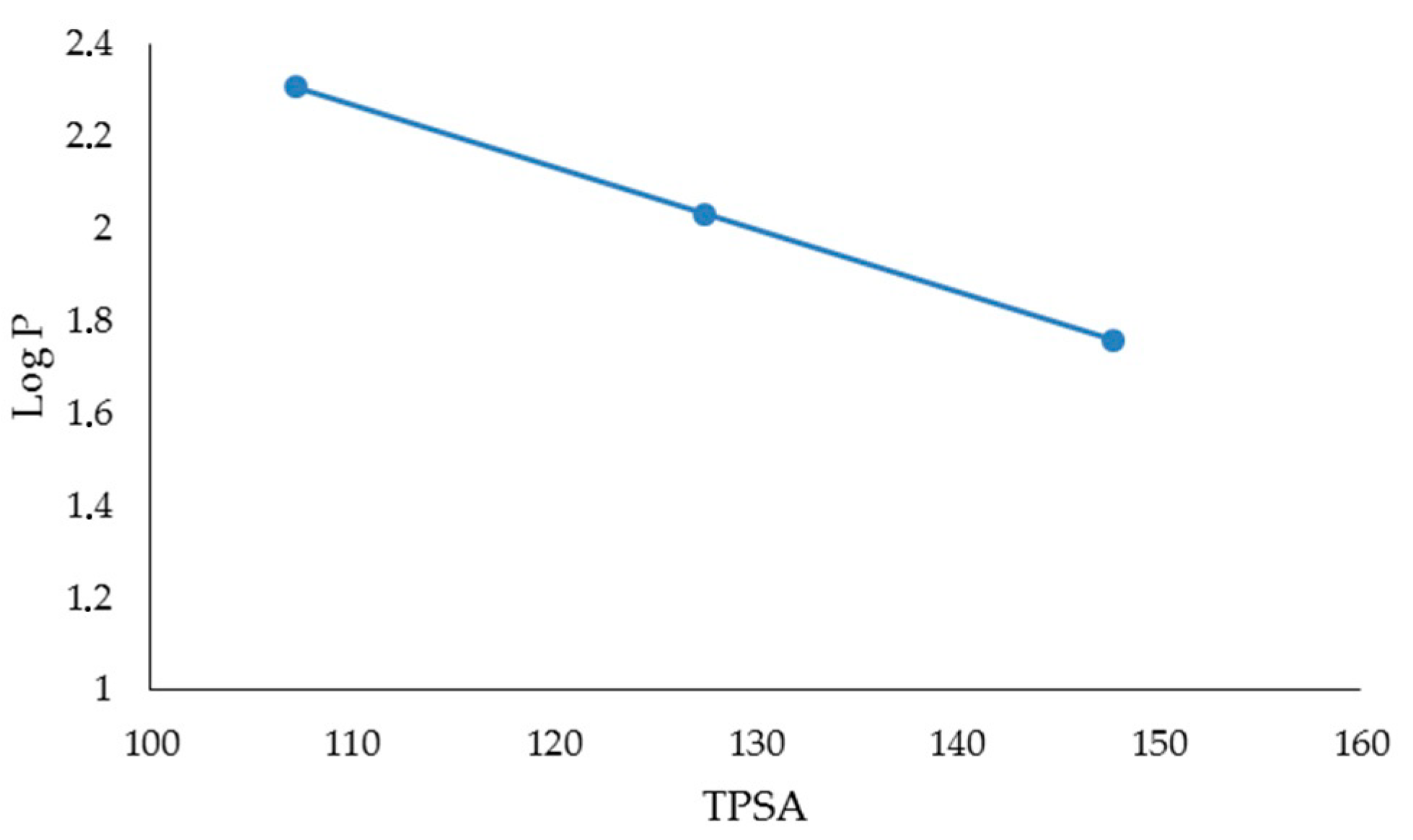
| FLAVONES | BDE (Kcal·mol−1) | IP (Kcal·mol−1) | TPSA | Log P |
|---|---|---|---|---|
| LUTEOLIN | 74.54 | 174.44 | 107.22 | 2.26 |
| APIGENIN | 82.20 | 176.05 | 86.99 | 2.20 |
| CHRYSIN | 91.85 | 176.00 | 66.76 | 2.50 |
| FLAVONOLS | BDE (Kcal·mol−1) | IP (Kcal·mol−1) | TPSA | Log P |
|---|---|---|---|---|
| MIRYCETIN | 71.08 | 161.40 | 147.68 | 1.76 |
| QUERCETIN | 72.35 | 166.08 | 127.45 | 2.03 |
| KAEMPFEROL | 83.80 | 167.99 | 107.22 | 2.31 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ronsisvalle, S.; Panarello, F.; Longhitano, G.; Siciliano, E.A.; Montenegro, L.; Panico, A. Natural Flavones and Flavonols: Relationships among Antioxidant Activity, Glycation, and Metalloproteinase Inhibition. Cosmetics 2020, 7, 71. https://doi.org/10.3390/cosmetics7030071
Ronsisvalle S, Panarello F, Longhitano G, Siciliano EA, Montenegro L, Panico A. Natural Flavones and Flavonols: Relationships among Antioxidant Activity, Glycation, and Metalloproteinase Inhibition. Cosmetics. 2020; 7(3):71. https://doi.org/10.3390/cosmetics7030071
Chicago/Turabian StyleRonsisvalle, Simone, Federica Panarello, Giusy Longhitano, Edy Angela Siciliano, Lucia Montenegro, and Annamaria Panico. 2020. "Natural Flavones and Flavonols: Relationships among Antioxidant Activity, Glycation, and Metalloproteinase Inhibition" Cosmetics 7, no. 3: 71. https://doi.org/10.3390/cosmetics7030071
APA StyleRonsisvalle, S., Panarello, F., Longhitano, G., Siciliano, E. A., Montenegro, L., & Panico, A. (2020). Natural Flavones and Flavonols: Relationships among Antioxidant Activity, Glycation, and Metalloproteinase Inhibition. Cosmetics, 7(3), 71. https://doi.org/10.3390/cosmetics7030071