
Holobiont Rebalancing by a Natural Gentian Extract on a Skin Dehydration Model

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Fermented Extract Production (In-House Experiments)
2.2. In Vitro Evaluation of Biological Activity (In-House Experiments)
2.2.1. Hyaluronic Acid Dosage
2.2.2. Epifluorescence Microscopy for CD44 and Lipids Quantification
2.3. In Vivo Evaluation of Biological Activity
2.3.1. Skin Benefit Evaluation
2.3.2. Metaproteomic Analysis
2.4. Statistical Analyses
3. Results
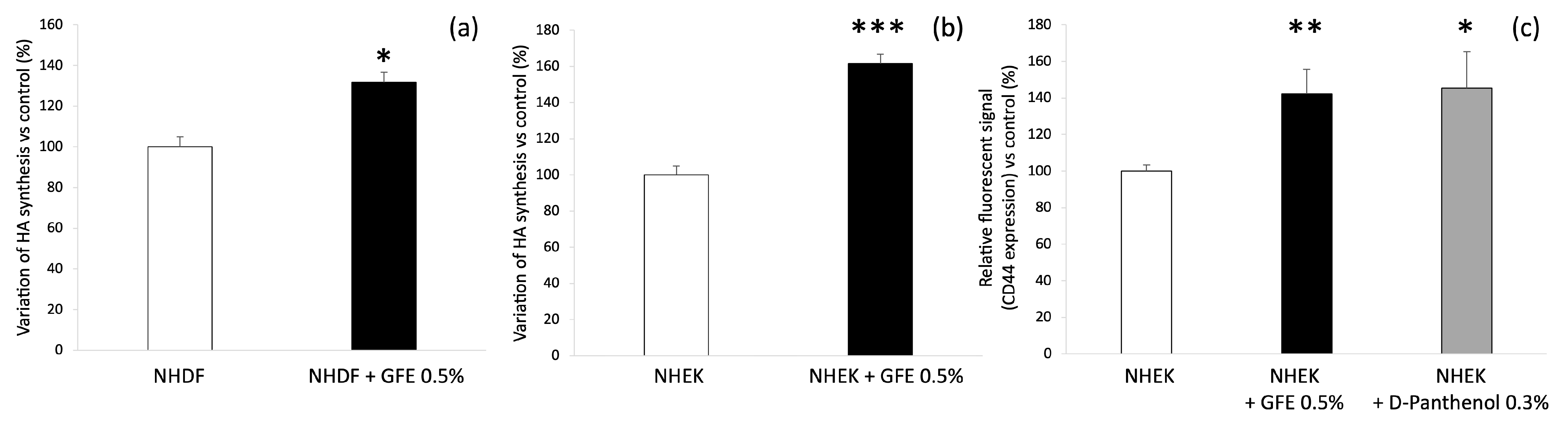
3.1. GFE Improves Hyaluronic Acid Pathway In Vitro
3.2. GFE Increases Lipid Synthesis In Vitro
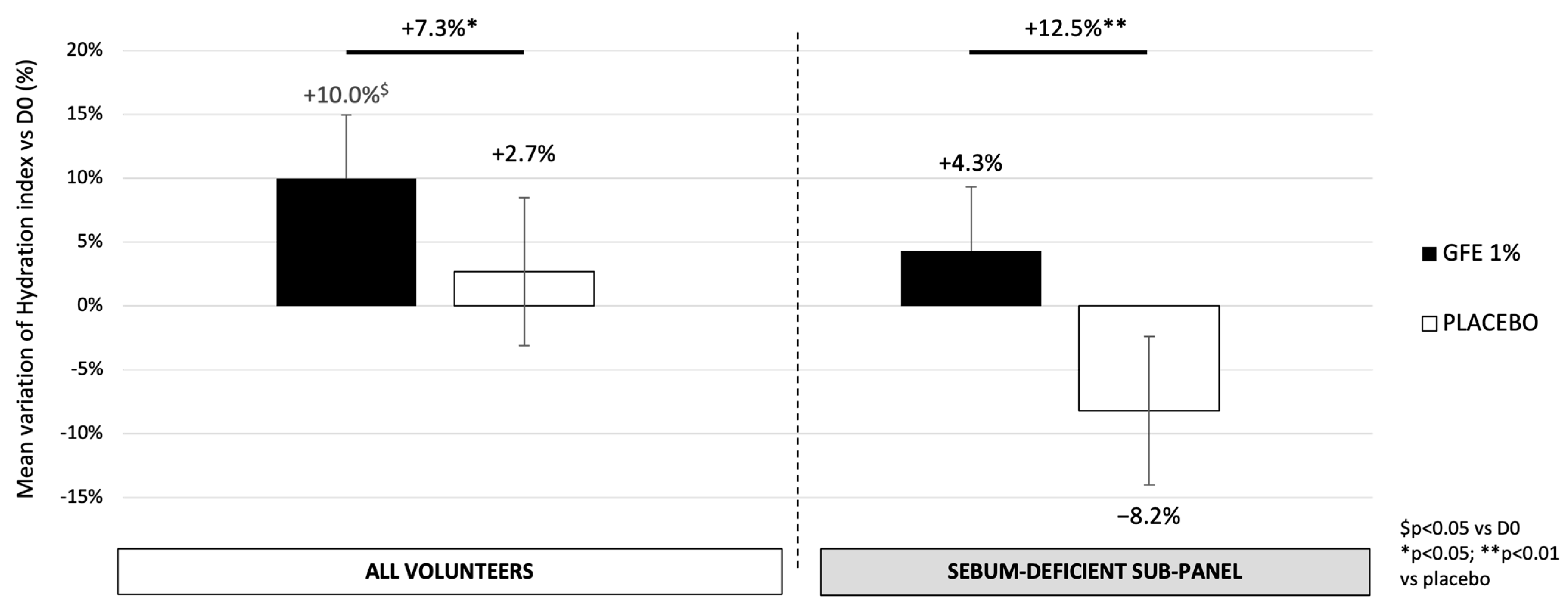
3.3. GFE Stimulates Hydration on Dry Skins
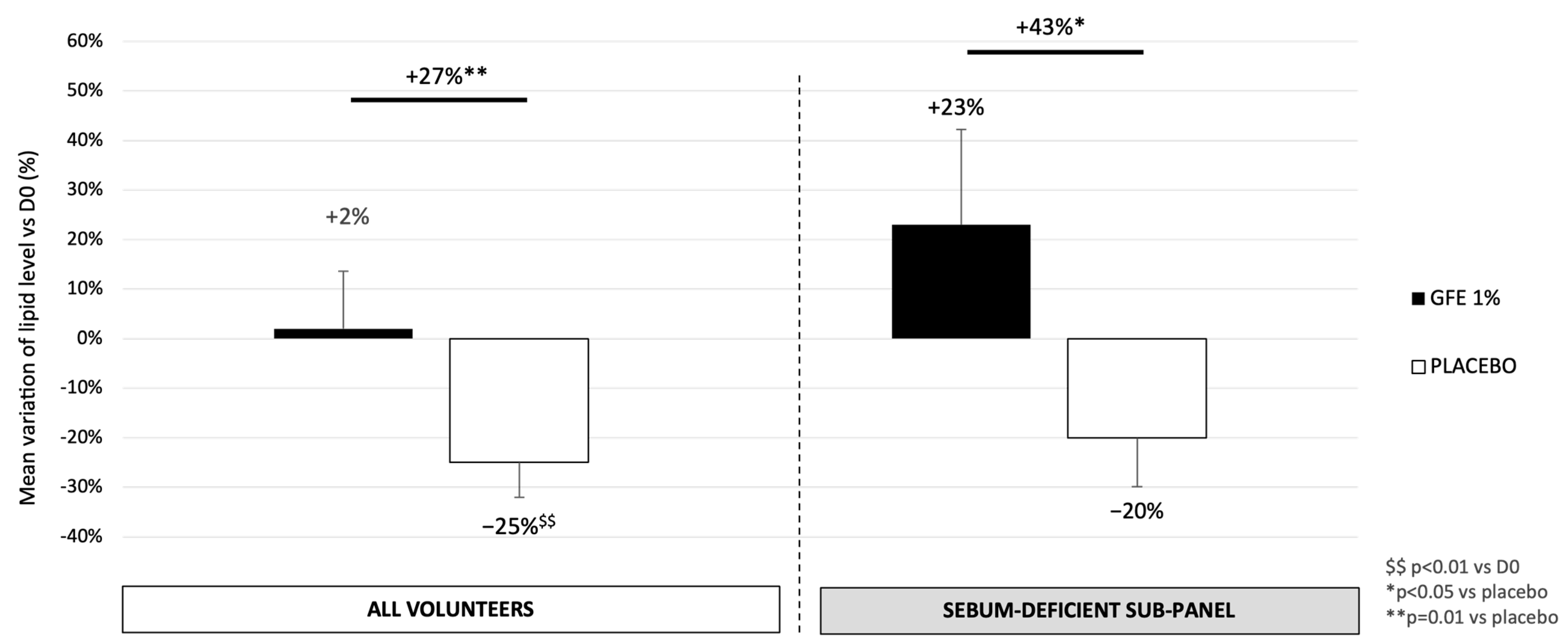
3.4. GFE Protects or Restores the Skin Surface Lipids According to Skin Condition
3.5. GFE Restores in Older Dehydrated Skin the Holobiotic Characteristics of Younger Skin and Stimulates Beneficial Bacteria Species
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Skowron, K.; Bauza-Kaszewska, J.; Kraszewska, Z.; Wiktorczyk-Kapischke, N.; Grudlewska-Buda, K.; Kwiecińska-Piróg, J.; Wałecka-Zacharska, E.; Radtke, L.; Gospodarek-Komkowska, E. Human Skin Microbiome: Impact of Intrinsic and Extrinsic Factors on Skin Microbiota. Microorganisms 2021, 9, 543. [Google Scholar] [CrossRef]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The Human Skin Microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- Oh, J.; Byrd, A.L.; Park, M.; NISC Comparative Sequencing Program; Kong, H.H.; Segre, J.A. Temporal Stability of the Human Skin Microbiome. Cell 2016, 165, 854–866. [Google Scholar] [CrossRef] [PubMed]
- Bouslimani, A.; Porto, C.; Rath, C.M.; Wang, M.; Guo, Y.; Gonzalez, A.; Berg-Lyon, D.; Ackermann, G.; Moeller Christensen, G.J.; Nakatsuji, T.; et al. Molecular Cartography of the Human Skin Surface in 3D. Proc. Natl. Acad. Sci. USA 2015, 112, E2120–E2129. [Google Scholar] [CrossRef]
- Guerrero, R.; Margulis, L.; Berlanga, M. Symbiogenesis: The Holobiont as a Unit of Evolution. Int. Microbiol. Off. J. Span. Soc. Microbiol. 2013, 16, 133–143. [Google Scholar] [CrossRef]
- Douladiris, N.; Vakirlis, E.; Vassilopoulou, E. Atopic Dermatitis and Water: Is There an Optimum Water Intake Level for Improving Atopic Skin? Child. Basel Switz. 2023, 10, 273. [Google Scholar] [CrossRef]
- Gidado, I.M.; Qassem, M.; Triantis, I.F.; Kyriacou, P.A. Review of Advances in the Measurement of Skin Hydration Based on Sensing of Optical and Electrical Tissue Properties. Sensors 2022, 22, 7151. [Google Scholar] [CrossRef] [PubMed]
- Masson, F. Skin hydration and hyaluronic acid. Ann. Dermatol. Venereol. 2010, 137 (Suppl. 1), S23–S25. [Google Scholar] [CrossRef]
- Bourguignon, L.Y.W.; Ramez, M.; Gilad, E.; Singleton, P.A.; Man, M.-Q.; Crumrine, D.A.; Elias, P.M.; Feingold, K.R. Hyaluronan-CD44 Interaction Stimulates Keratinocyte Differentiation, Lamellar Body Formation/Secretion, and Permeability Barrier Homeostasis. J. Investig. Dermatol. 2006, 126, 1356–1365. [Google Scholar] [CrossRef]
- González, O.; Mayo, S.; Rodríguez, A.; Casquero, P. Effect of Soil in the Growth of Gentiana Lutea Radical System in North Mountain of Léon (Spain). Acta Hortic. 2012, 955, 281–285. [Google Scholar] [CrossRef]
- Renard, P.; Canet, I.; Sancelme, M.; Wirgot, N.; Deguillaume, L.; Delort, A.-M. Screening of Cloud Microorganisms Isolated at the Puy de Dôme (France) Station for the Production of Biosurfactants. Atmos. Chem. Phys. Discuss. 2016, 16, 12347–12358. [Google Scholar] [CrossRef]
- Aberham, A.; Schwaiger, S.; Stuppner, H.; Ganzera, M. Quantitative Analysis of Iridoids, Secoiridoids, Xanthones and Xanthone Glycosides in Gentiana Lutea L. Roots by RP-HPLC and LC-MS. J. Pharm. Biomed. Anal. 2007, 45, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Muhamad Fadzil, N.S.; Sekar, M.; Gan, S.H.; Bonam, S.R.; Wu, Y.S.; Vaijanathappa, J.; Ravi, S.; Lum, P.T.; Dhadde, S.B. Chemistry, Pharmacology and Therapeutic Potential of Swertiamarin—A Promising Natural Lead for New Drug Discovery and Development. Drug Des. Devel. Ther. 2021, 15, 2721–2746. [Google Scholar] [CrossRef]
- Dzydzan, O.; Brodyak, I.; Sokół-Łętowska, A.; Kucharska, A.Z.; Sybirna, N. Loganic Acid, an Iridoid Glycoside Extracted from Cornus Mas L. Fruits, Reduces of Carbonyl/Oxidative Stress Biomarkers in Plasma and Restores Antioxidant Balance in Leukocytes of Rats with Streptozotocin-Induced Diabetes Mellitus. Life 2020, 10, 349. [Google Scholar] [CrossRef] [PubMed]
- Soltés, L.; Mendichi, R.; Kogan, G.; Schiller, J.; Stankovska, M.; Arnhold, J. Degradative Action of Reactive Oxygen Species on Hyaluronan. Biomacromolecules 2006, 7, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Busse, H.-J.; Denner, E.B.M.; Buczolits, S.; Salkinoja-Salonen, M.; Bennasar, A.; Kämpfer, P. Sphingomonas Aurantiaca Sp. Nov., Sphingomonas Aerolata Sp. Nov. and Sphingomonas Faeni Sp. Nov., Air- and Dustborne and Antarctic, Orange-Pigmented, Psychrotolerant Bacteria, and Emended Description of the Genus Sphingomonas. Int. J. Syst. Evol. Microbiol. 2003, 53, 1253–1260. [Google Scholar] [CrossRef] [PubMed]
- Hassan, N.; Anesio, A.M.; Rafiq, M.; Holtvoeth, J.; Bull, I.; Haleem, A.; Shah, A.A.; Hasan, F. Temperature Driven Membrane Lipid Adaptation in Glacial Psychrophilic Bacteria. Front. Microbiol. 2020, 11, 824. [Google Scholar] [CrossRef]
- Ebner, F.; Heller, A.; Rippke, F.; Tausch, I. Topical Use of Dexpanthenol in Skin Disorders. Am. J. Clin. Dermatol. 2002, 3, 427–433. [Google Scholar] [CrossRef]
- Zuang, V.; Rona, C.; Distante, F.; Berdasca, E. The Use of a Capacitance Device to Evaluate the Hydration of Human Skin. J. Appl. Cosmetol. 1997, 15, 95–102. [Google Scholar]
- Luebberding, S.; Krueger, N.; Kerscher, M. Skin Physiology in Men and Women: In Vivo Evaluation of 300 People Including TEWL, SC Hydration, Sebum Content and Skin Surface pH. Int. J. Cosmet. Sci. 2013, 35, 477–483. [Google Scholar] [CrossRef]
- Shannon, C.E. Communication Theory of Secrecy Systems. Bell Syst. Tech. J. 1949, 28, 656–715. [Google Scholar] [CrossRef]
- Simpson, E. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Bray, J.; Curtis, J. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Albouy, M.; Aubailly, S.; Jeanneton, O.; Marteau, C.; Sobilo, L.; Boulgana, R.; Bru, G.; Bellanger, M.; Leblanc, E.; Dos Santos, M.; et al. Skin-Protective Biological Activities of Bio-Fermented Aframomum Angustifolium Extract by a Consortium of Microorganisms. Front. Pharmacol. 2023, 14, 1303198. [Google Scholar] [CrossRef] [PubMed]
- Stanek-Wandzel, N.; Zarębska, M.; Wasilewski, T.; Hordyjewicz-Baran, Z.; Zajszły-Turko, E.; Tomaka, M.; Bujak, T.; Ziemlewska, A.; Nizioł-Łukaszewska, Z. Kombucha Fermentation as a Modern Way of Processing Vineyard By-Products into Cosmetic Raw Materials. Int. J. Cosmet. Sci. 2023, 45, 834–850. [Google Scholar] [CrossRef]
- Berthon, J.-Y.; Cabannes, M.; Bouton, C.; Carre, M.; Bridon, E.; Filaire, E. In Vitro, Ex Vivo and Clinical Approaches to Evaluate the Potential Effect of Gentiana Lutea Extract on Skin. Int. J. Cosmet. Sci. 2023, 45, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.A.; Bouchard, T.; St John, T.; Wayner, E.; Carter, W.G. Human Keratinocytes Express a New CD44 Core Protein (CD44E) as a Heparan-Sulfate Intrinsic Membrane Proteoglycan with Additional Exons. J. Cell Biol. 1991, 113, 207–221. [Google Scholar] [CrossRef]
- Wang, C.; Tammi, M.; Tammi, R. Distribution of Hyaluronan and Its CD44 Receptor in the Epithelia of Human Skin Appendages. Histochemistry 1992, 98, 105–112. [Google Scholar] [CrossRef]
- Kirschner, N.; Haftek, M.; Niessen, C.M.; Behne, M.J.; Furuse, M.; Moll, I.; Brandner, J.M. CD44 Regulates Tight-Junction Assembly and Barrier Function. J. Investig. Dermatol. 2011, 131, 932–943. [Google Scholar] [CrossRef]
- Gruber, J.V.; Holtz, R.; Riemer, J. Hyaluronic Acid (HA) Stimulates the in Vitro Expression of CD44 Proteins but Not HAS1 Proteins in Normal Human Epidermal Keratinocytes (NHEKs) and Is HA Molecular Weight Dependent. J. Cosmet. Dermatol. 2022, 21, 1193–1198. [Google Scholar] [CrossRef]
- Pasonen-Seppänen, S.; Karvinen, S.; Törrönen, K.; Hyttinen, J.M.T.; Jokela, T.; Lammi, M.J.; Tammi, M.I.; Tammi, R. EGF Upregulates, Whereas TGF-Beta Downregulates, the Hyaluronan Synthases Has2 and Has3 in Organotypic Keratinocyte Cultures: Correlations with Epidermal Proliferation and Differentiation. J. Investig. Dermatol. 2003, 120, 1038–1044. [Google Scholar] [CrossRef] [PubMed]
- Valachová, K.; Hassan, M.E.; Šoltés, L. Hyaluronan: Sources, Structure, Features and Applications. Molecules 2024, 29, 739. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; He, D.; Wang, J.; Xiao, Y.; Gong, L.; Tang, C.; Peng, H.; Qiu, X.; Liu, R.; Zhang, T.; et al. Swertiamarin Relieves Radiation-Induced Intestinal Injury by Limiting DNA Damage. Mol. Cell. Biochem. 2024. [CrossRef]
- Zhou, Q.; Zhou, Q.; Xia, R.; Zhang, P.; Xie, Y.; Yang, Z.; Khan, A.; Zhou, Z.; Tan, W.; Liu, L. Swertiamarin or Heat-Transformed Products Alleviated APAP-Induced Hepatotoxicity via Modulation of Apoptotic and Nrf-2/NF-κB Pathways. Heliyon 2023, 9, e18746. [Google Scholar] [CrossRef]
- Prakash, A.N.; Prasad, N.; Puppala, E.R.; Panda, S.R.; Jain, S.; Ravichandiran, V.; Singh, M.; Naidu, V.G.M. Loganic Acid Protects against Ulcerative Colitis by Inhibiting TLR4/NF-κB Mediated Inflammation and Activating the SIRT1/Nrf2 Anti-Oxidant Responses in-Vitro and in-Vivo. Int. Immunopharmacol. 2023, 122, 110585. [Google Scholar] [CrossRef]
- Imokawa, G.; Kuno, H.; Kawai, M. Stratum Corneum Lipids Serve as a Bound-Water Modulator. J. Investig. Dermatol. 1991, 96, 845–851. [Google Scholar] [CrossRef] [PubMed]
- Camilion, J.V.; Khanna, S.; Anasseri, S.; Laney, C.; Mayrovitz, H.N. Physiological, Pathological, and Circadian Factors Impacting Skin Hydration. Cureus 2022, 14, e27666. [Google Scholar] [CrossRef]
- Jang, S.I.; Han, J.; Lee, M.; Seo, J.; Kim, B.J.; Kim, E. A Study of Skin Characteristics According to Humidity during Sleep. Skin Res. Technol. 2019, 25, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Duplan, H.; Nocera, T. Skin hydration and hydrating products. Ann. Dermatol. Venereol. 2018, 145, 376–384. [Google Scholar] [CrossRef]
- Egawa, M.; Oguri, M.; Kuwahara, T.; Takahashi, M. Effect of Exposure of Human Skin to a Dry Environment. Skin Res. Technol. 2002, 8, 212–218. [Google Scholar] [CrossRef]
- Wang, Z.; Man, M.-Q.; Li, T.; Elias, P.M.; Mauro, T.M. Aging-Associated Alterations in Epidermal Function and Their Clinical Significance. Aging 2020, 12, 5551–5565. [Google Scholar] [CrossRef] [PubMed]
- Radner, F.P.; Grond, S.; Haemmerle, G.; Lass, A.; Zechner, R. Fat in the Skin: Triacylglycerol Metabolism in Keratinocytes and Its Role in the Development of Neutral Lipid Storage Disease. Dermatoendocrinol 2011, 3, 77–83. [Google Scholar] [CrossRef]
- Sakai, S.; Yasuda, R.; Sayo, T.; Ishikawa, O.; Inoue, S. Hyaluronan Exists in the Normal Stratum Corneum. J. Investig. Dermatol. 2000, 114, 1184–1187. [Google Scholar] [CrossRef] [PubMed]
- Lefèvre-Utile, A.; Braun, C.; Haftek, M.; Aubin, F. Five Functional Aspects of the Epidermal Barrier. Int. J. Mol. Sci. 2021, 22, 11676. [Google Scholar] [CrossRef] [PubMed]
- Evrard, C.; Lambert de Rouvroit, C.; Poumay, Y. Epidermal Hyaluronan in Barrier Alteration-Related Disease. Cells 2021, 10, 3096. [Google Scholar] [CrossRef]
- Karaduta, O.; Dvanajscak, Z.; Zybailov, B. Metaproteomics-An Advantageous Option in Studies of Host-Microbiota Interaction. Microorganisms 2021, 9, 980. [Google Scholar] [CrossRef]
- Zheng, Y.; Hunt, R.L.; Villaruz, A.E.; Fisher, E.L.; Liu, R.; Liu, Q.; Cheung, G.Y.C.; Li, M.; Otto, M. Commensal Staphylococcus Epidermidis Contributes to Skin Barrier Homeostasis by Generating Protective Ceramides. Cell Host Microbe 2022, 30, 301–313.e9. [Google Scholar] [CrossRef]
- Ma, L.; Niu, Y.; Yuan, C.; Bai, T.; Yang, S.; Wang, M.; Li, Y.; Shao, L. The Characteristics of the Skin Physiological Parameters and Facial Microbiome of “Ideal Skin” in Shanghai Women. Clin. Cosmet. Investig. Dermatol. 2023, 16, 325–337. [Google Scholar] [CrossRef]
- Wang, D.-Q.; Li, X.; Zhang, R.-Y.; Yuan, C.; Yan, B.; Humbert, P.; Quan, Z.-X. Effects of Investigational Moisturizers on the Skin Barrier and Microbiome Following Exposure to Environmental Aggressors: A Randomized Clinical Trial and Ex Vivo Analysis. J. Clin. Med. 2023, 12, 6078. [Google Scholar] [CrossRef]
- Zhu, Y.; Yu, X.; Cheng, G. Human Skin Bacterial Microbiota Homeostasis: A Delicate Balance between Health and Disease. mLife 2023, 2, 107–120. [Google Scholar] [CrossRef]
- Wu, J.; Li, L.; Zhang, T.; Lu, J.; Tai, Z.; Zhu, Q.; Chen, Z. The Epidermal Lipid-Microbiome Loop and Immunity: Important Players in Atopic Dermatitis. J. Adv. Res. 2024, S2090-1232(24)00088-2. [Google Scholar] [CrossRef] [PubMed]
- Filatov, V.; Sokolova, A.; Savitskaya, N.; Olkhovskaya, M.; Varava, A.; Ilin, E.; Patronova, E. Synergetic Effects of Aloe Vera Extract with Trimethylglycine for Targeted Aquaporin 3 Regulation and Long-Term Skin Hydration. Mol. Basel Switz. 2024, 29, 1540. [Google Scholar] [CrossRef] [PubMed]
- Kosik-Kozioł, A.; Nakielski, P.; Rybak, D.; Frączek, W.; Rinoldi, C.; Lanzi, M.; Grodzik, M.; Pierini, F. Adhesive Antibacterial Moisturizing Nanostructured Skin Patch for Sustainable Development of Atopic Dermatitis Treatment in Humans. ACS Appl. Mater. Interfaces 2024, 16, 32128–32146. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roux, L.N.; Dreux-Zigha, A.; Rey, C.; Boutot, C.; Laurent, Y.; Beytur, S.; Metton, I.; Abraham, J.-D.; Berthon, J.-Y. Holobiont Rebalancing by a Natural Gentian Extract on a Skin Dehydration Model. Cosmetics 2024, 11, 132. https://doi.org/10.3390/cosmetics11040132
Roux LN, Dreux-Zigha A, Rey C, Boutot C, Laurent Y, Beytur S, Metton I, Abraham J-D, Berthon J-Y. Holobiont Rebalancing by a Natural Gentian Extract on a Skin Dehydration Model. Cosmetics. 2024; 11(4):132. https://doi.org/10.3390/cosmetics11040132
Chicago/Turabian StyleRoux, Lauriane N., Assia Dreux-Zigha, Célia Rey, Carine Boutot, Yoan Laurent, Sercan Beytur, Isabelle Metton, Jean-Daniel Abraham, and Jean-Yves Berthon. 2024. "Holobiont Rebalancing by a Natural Gentian Extract on a Skin Dehydration Model" Cosmetics 11, no. 4: 132. https://doi.org/10.3390/cosmetics11040132
APA StyleRoux, L. N., Dreux-Zigha, A., Rey, C., Boutot, C., Laurent, Y., Beytur, S., Metton, I., Abraham, J.-D., & Berthon, J.-Y. (2024). Holobiont Rebalancing by a Natural Gentian Extract on a Skin Dehydration Model. Cosmetics, 11(4), 132. https://doi.org/10.3390/cosmetics11040132