
Potential of Slime Molds as a Novel Source for the Cosmetics Industry

 ,
,  ,
,
Abstract
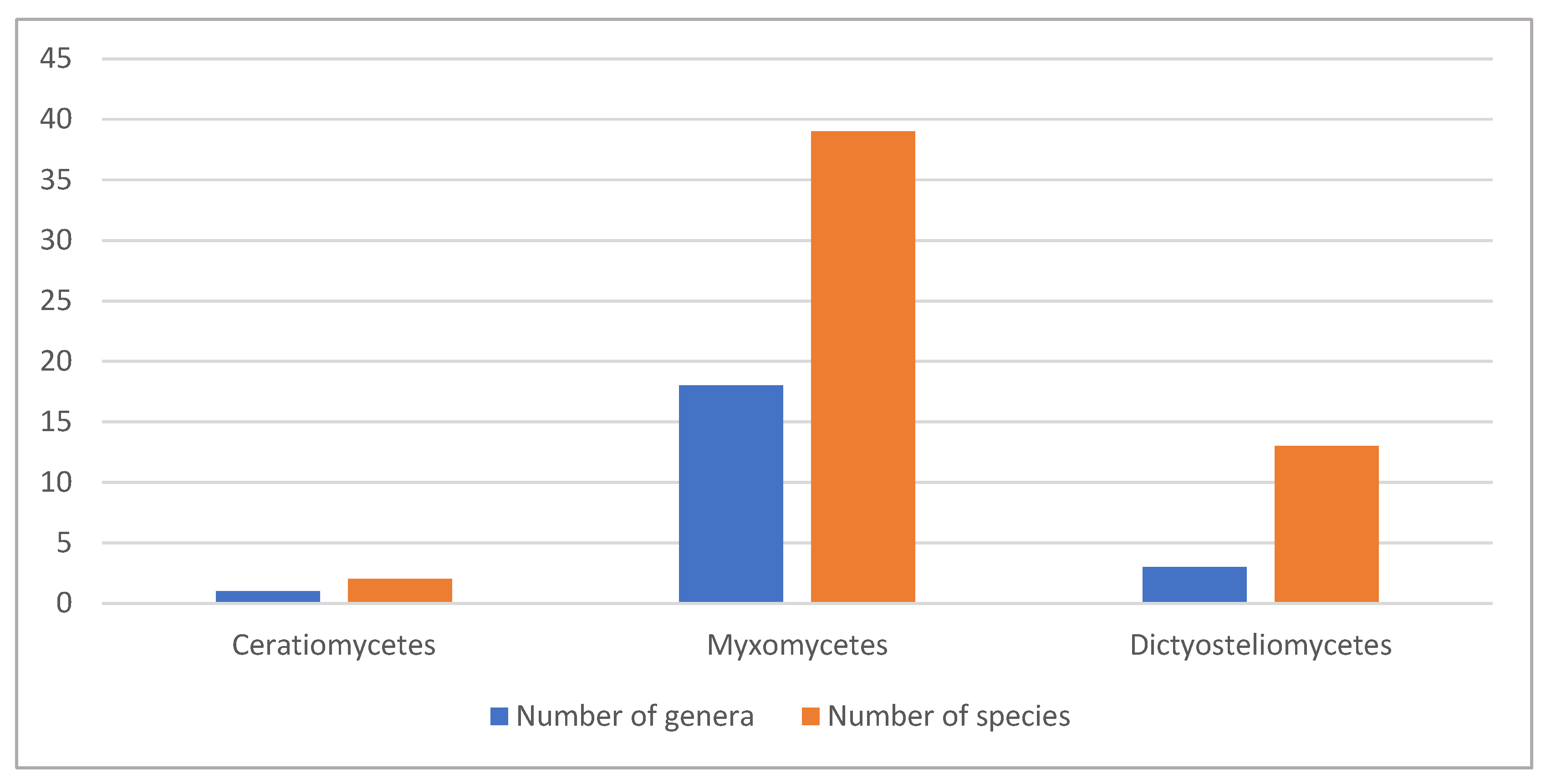
1. Introduction
2. Materials and Methods
3. Results
3.1. Pigments
3.1.1. Plasmodial Pigments
3.1.2. Pigments of Dormant Sclerotia
3.1.3. Pigments of Fruiting Bodies
3.1.4. Spore Pigments
3.2. Lipids
3.2.1. Fatty Acids
3.2.2. Sterols
3.2.3. Phospholipids
3.2.4. Glycerolipids
3.2.5. Esters and Lactones
3.3. Glycosides
3.4. Proteins
3.5. Exopolysaccharides
3.6. Aromatic Compounds
3.7. Safety Aspects
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Seca, A.M.L.; Moujir, L. Natural Compounds: A Dynamic Field of Applications. Appl. Sci. 2020, 10, 4025. [Google Scholar] [CrossRef]
- Khan, R.A. Natural products chemistry: The emerging trends and prospective goals. Saudi Pharm. J. 2018, 26, 739–753. [Google Scholar] [CrossRef] [PubMed]
- Amberg, N.; Fogarassy, C. Green Consumer Behavior in the Cosmetics Market. Resources 2019, 8, 137. [Google Scholar] [CrossRef]
- Desam, N.; Al-Rajab, A. The Importance of Natural Products in Cosmetics. In Bioactive Natural Products for Pharmaceutical Applications; Pal, D., Nayak, A.K., Eds.; Advanced Structured Materials; Springer: Cham, Switzerland, 2021; Volume 140. [Google Scholar] [CrossRef]
- Stoyneva-Gärtner, M.; Uzunov, B.; Gärtner, G. Enigmatic microalgae from aeroterrestrial and extreme habitats in cosmetics: The potential of the untapped natural sources. Cosmetics 2020, 7, 27. [Google Scholar] [CrossRef]
- Masamiishibashi. Search for Bioactive Natural Products from Unexploited Microbial Resources. In Studies in Natural Products Chemistry; Atta-ur-Rahman, Ed.; Elsevier: Amsterdam, The Netherlands, 2003; Volume 29, Part J, pp. 223–262. [Google Scholar] [CrossRef]
- Stephenson, S.L. Secretive Slime Molds. Myxomycetes of Australia; CSIRO Publishing: Melbourne, Australia, 2021. [Google Scholar]
- Stephenson, S.L.; Rojas, C. (Eds.) Myxomycetes: Biology, Systematics, Biogeography and Ecology; Elsevier Academic Press: London, UK, 2017. [Google Scholar]
- Rojas, C.; Stephenson, S.L. (Eds.) Myxomycetes: Biology, Systematics, Biogeography and Ecology, 2nd ed.; Elsevier Academic Press: London, UK, 2021. [Google Scholar]
- Sasaki, H.; Kubohara, Y.; Ishigaki, H.; Takahashi, K.; Eguchi, H.; Sugawara, A.; Oshima, Y.; Kikuchi, H. Two new terpenes isolated from Dictyostelium cellular slime molds. Molecules 2020, 25, 2895. [Google Scholar] [CrossRef] [PubMed]
- Pears, C.J.; Gross, J.D. Microbe Profile: Dictyostelium discoideum: Model system for development, chemotaxis and biomedical research. Microbiology 2021, 167, 001040. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, S.; Thulin, M.; Cavender, J.C.; Escalante, R.; Kawakami, S.; Lado, C.; Landolt, J.; Nanjundiah, V.; Queller, D.; Strassmann, J.; et al. A new classification of the Dictyostelids. Protists 2017, 169, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Boussard, A.; Fessel, A.; Oettmeier, C.; Briard, L.; Döbereiner, H.-G.; Dussutour, A. Adaptive behaviour andlearning in slime moulds: The role ofoscillations. Philos. Trans. R. Soc. 2021, B376, 20190757. [Google Scholar] [CrossRef] [PubMed]
- White, D.H. What is STEM education and why it is important? JFATE 2014, 1, 1–8. [Google Scholar]
- Baldauf, S.L.; Strassmann, J.E. Dictyostelia. In Handbook of the Protists, 2nd ed.; Simpson, A.G.B., Archibald, J.M., Slamovits, C.H., Eds.; Springer: Heidelberg/Germany, Germany, 2017; pp. 1433–1477. [Google Scholar]
- Alexopoulos, C.J.; Mims, C.W.; Blackwell, M. Introductory Mycology; Wiley India Pvt. Limited: New Delhi, India, 2007; pp. 1–880. [Google Scholar]
- Stephenson, S.L.; Stempen, H. Myxomycetes. A Handbook of Slime Molds, 1st ed.; Paperback Edition Printed 2000; Timber Press Inc.: Portland, OR, USA, 1994. [Google Scholar]
- Adl, S.M.; Bass, D.; Lane, C.E.; Lukeš, J.; Schoch, C.L.; Smirnov, A.; Agatha, S.; Berney, C.; Brown, M.W.; Burki, F.; et al. Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes. J. Eukaryot. Microbiol. 2018, 66, 4–119. [Google Scholar] [CrossRef]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K.; et al. International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code) Adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017 (Electronic ed.); Glashütten: International Association for Plant Taxonomy: Bratislava, Slovakia, 2018. [Google Scholar] [CrossRef]
- Keller, H.; Everhart, S. Importance of Myxomycetes in Biological Research and Teaching. Pap. Plant Pathol. 2010, 366, 16–27. [Google Scholar]
- Leontyev, D.V.; Schnittler, M.; Stephenson, S.L.; Novozhilov, Y.K.; Shchepin, O.N. Towards a phylogenetic classification of the Myxomycetes. Phytotaxa 2019, 399, 209–238. [Google Scholar] [CrossRef]
- Webster, J.; Weber, R.W.S. Introduction to Fungi, 3rd ed.; Cambridge University Press: Cambridge, UK, 2007; pp. 1–841. [Google Scholar] [CrossRef]
- Bonner, J. The Social Amoebae: The Biology of Cellular Slime Molds.; Princeton University Press: Princeton, NJ, USA; Oxford, UK, 2009; pp. 1–126. [Google Scholar]
- Stoyneva-Gärtner, M.; Uzunov, B. Bases of Systematics of Algae and Fungi; JAMG Books: Sofia, Bulgaria, 2017; pp. 1–189. [Google Scholar]
- Krzywda, A.; Petelenz, E.; Michalczyki, D.; Płonka, P. Sclerotia of the acellular (true) slime mould Fuligo septica as a model to study melanization and anabiosis. CMBL 2008, 13, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Woyzichovski, J.; Shchepin, O.N.; Schnittler, M. High environmentally induced plasticity in spore size and numbers of nuclei per spore in Physarum albescens (Myxomycetes). Protist 2022, 173, 125904. [Google Scholar] [CrossRef]
- Stephenson, S.L.; Fiore-Donno, A.M.; Schnittler, M. Myxomycetes in soil. Soil Biol. Biochem. 2011, 43, 2237–2242. [Google Scholar] [CrossRef]
- Loquin, M.; Prevot, A.R. Etude de quelque antibiotiques produits par les myxomycetes. Ann Inst Pasteur 1948, 75, 8–13. [Google Scholar]
- Tafakori, V. Slime molds as a valuable source of antimicrobial agents. AMB Express 2021, 11, 92. [Google Scholar] [CrossRef] [PubMed]
- Steglich, W. Slime molds (myxomycetes) as a source of new biological active metabolites. Pure Appl. Chem. 1989, 61, 281–288. [Google Scholar] [CrossRef]
- Dembitsky, V.; Rezanka, T.; Spızek, J.; Hanus, L. Secondary metabolites of slime molds (myxomycetes). Phytochemistry 2005, 66, 747–769. [Google Scholar] [CrossRef]
- Ishibashi, M. Study on myxomycetes as a new source of bioactive natural products. YakugakuZasshi 2007, 127, 1369–1381. [Google Scholar] [CrossRef]
- Baba, H.; Sevindik, M.; Dogan, M.; Akgul, H. Antioxidant, antimicrobial activities and heavy metal contents of some myxomycetes. Fresenius Environ. Bull. 2020, 9, 7840–7846. [Google Scholar]
- Kubohara, Y.; Kikuchi, H. Dictyostelium: An important source of structural and functional diversity in drug discovery. Cells 2019, 8, 6. [Google Scholar] [CrossRef]
- Kubohara, Y.; Oshima, Y.; Shiratsuchi, Y.; Ishigaki, H.; Takahashi, K.; Kikuchi, H. Antimicrobial activities of Dictyosteliumdiferentiation-inducing factors and their derivatives. Biomolecules 2019, 9, 163. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Lin, N.; Wu, B. Laboratory culture and bioactive natural products of myxomycetes. Fitoterapia 2020, 146, 104725. [Google Scholar] [CrossRef]
- Sevindik, M.; Baba, H.; Bal, C.; Colak, O.; Akgul, H. Antioxidant, oxidant and antimicrobial capacities of Physarum album (Bull.) Chevall. JBMOA 2018, 6, 317–320. [Google Scholar] [CrossRef][Green Version]
- Yokota, M.; Nitta, K. Purification and some properties of hemagglutinin from the myxomycete Physarum polycephalum. Experientia 1996, 52, 544–548. [Google Scholar] [CrossRef]
- Kikuchi, H.; Saito, Y.; Komiya, J.; Takaya, Y.; Honma, S.; Nakahata, N.; Ito, A.; Oshima, Y. Furanodictine A and B: Amino sugar analogues produced by cellular slime molds Dictyostelium discoideum showing neuronal differentiation activity. J. Org. Chem. 2001, 66, 6982–6987. [Google Scholar] [CrossRef]
- Jahan, A.; Ahmad, I.; Fatima, N.; Ansari, V.; Akhtar, J. Algal bioactive compounds in the cosmeceutical industry: A review. Phycologia 2017, 56, 410–422. [Google Scholar] [CrossRef]
- Ahmad, J. Lipid nanoparticles-based cosmetics with potential application in alleviating skin disorders. Cosmetics 2021, 8, 84. [Google Scholar] [CrossRef]
- Stoyneva-Gärtner, M.; Uzunov, B.; Gärtner, G. Aeroterrestrial and extremophilic microalgae as promising sources for lipids and lipid nanoparticles in dermal cosmetics. Cosmetics 2022, 9, 11. [Google Scholar] [CrossRef]
- PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/ (accessed on 9 November 2022).
- Taxonomy Browser. Available online: https://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi (accessed on 3 November 2022).
- Index Fungorum. Available online: http://www.indexfungorum.org/names/names.asp (accessed on 10 November 2022).
- Walimbe, S. Global Cosmetic Pigments Market Size Is Anticipated to Cross USD 1 Billion by 2026. Available online: https://www.hpci-events.com/global-cosmetic-pigments-market-size-is-anticipated-to-cross-usd-1-billion-by-2026/#:~:text=Pigments%20are%20used%20in%20various%20formulations&text=One%20of%20the%20most%20integral,personal%20care%20items%20like%20soap (accessed on 11 November 2022).
- Guerra, E.; Llompart, M.; Garcia-Jares, C. Analysis of Dyes in Cosmetics: Challenges and Recent Developments. Cosmetics 2018, 5, 47. [Google Scholar] [CrossRef]
- Wormington, W.M.; Weaver, R.F. Photoreceptor pigment that induces differentiation in the slime mold Physarum polycephalum. Proc. Natl. Acad. Sci. USA 1976, 73, 3896–3899. [Google Scholar] [CrossRef] [PubMed]
- Young, A.M. A Field Guide to the Fungi of Australia; University of New South Wales Press Ltd.: Sydney, Australia, 2005; ISBN 9780868407425. [Google Scholar]
- Velten, R.; Josten, I.; Steglich, W. Ungesättigte 6-Alkylpyrone aus dem Schleimpilz Ceratiomyxa fruticulosa (Myxomycetes). Liebigs Ann. 1995, 1, 81–85. [Google Scholar] [CrossRef]
- Kamata, K.; Kiyota, M.; Naoe, A.; Nakatani, S.; Yamamoto, Y.; Hayashi, M.; Komiyama, K.; Yamori, T.; Ishibashi, M. New bisindole alkaloids isolated from myxomycetes Arcyria cinerea and Lycogala epidendrum. Chem. Pharm. Bull. 2005, 53, 594–597. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, M. Isolation of bioactive natural products from myxomycetes. Med. Chem. 2005, 1, 575–590. [Google Scholar] [CrossRef] [PubMed]
- Shintani, A.; Toume, K.; Rifai, Y.; Arai, M.; Ishibashi, M. A bisindole alkaloid with hedgehog signal inhibitory activity from the myxomycete Perichaena chrysosperma. J. Nat. Prod. 2010, 73, 1711–1713. [Google Scholar] [CrossRef]
- Řezanka, T. Polyunsaturated and unusual fatty acids from slime moulds. Phytochemistry 1993, 33, 1441–1444. [Google Scholar] [CrossRef]
- Řezanka, T. Glycosides of polyenoic branched fatty acids from myxomycetes. Phytochemistry 2002, 60, 639–646. [Google Scholar] [CrossRef]
- Steglich, W.; Steffan, B.; Kopanski, L.; Eckhardt, G. Pilzpigmente, 36. Indolfarbstoffe aus Fruchtkörpern des Schleimpilzes Arcyria denudata. Angew. Chem. Int. Ed. 1980, 19, 459–460. [Google Scholar] [CrossRef]
- Mayer, G.; Wille, G.; Brenner, M.; Zeitler, K.; Steglich, W. Total synthesis of the slime mold alkaloids arcyroxocin A and B. Synth 2011, 2, 330–336. [Google Scholar] [CrossRef]
- Kamata, K.; Suetsugu, T.; Yamamoto, Y.; Hayashi, M.; Komiyama, K.; Ishibashi, M. Bisindole Alkaloids from Myxomycetes Arcyria denudata and Arcyria obvelata. J. Nat. Prod. 2006, 69, 1252–1254. [Google Scholar] [CrossRef]
- Nakatani, S.; Naoe, A.; Yamamoto, Y.; Yamauchi, T.; Yamaguchi, N.; Ishibashi, M. Isolation of bisindole alkaloids that inhibit the cell cycle from Myxomycetes Arcyria ferruginea and Tubifera casparyi. Bioorg. Med. Chem. Lett. 2003, 13, 2879–2881. [Google Scholar] [CrossRef]
- Kaniwa, K.; Arai, M.A.; Li, X.; Ishibashi, M. Synthesis, determination of stereochemistry, and evaluation of new bisindole alkaloids from the myxomycete Arcyria ferruginea: An approach for Wnt signal inhibitor. Bioorg. Med. Chem. Lett. 2007, 17, 4254–4257. [Google Scholar] [CrossRef]
- Iwata, D.; Ishibashi, M.; Yamamoto, Y. Cribrarione B, a new naphthoquinone pigment from the myxomycete Cribraria cancellata. J. Nat. Prod. 2003, 66, 1611–1612. [Google Scholar] [CrossRef]
- Misono, Y.; Ishikawa, Y.; Yamamoto, Y.; Hayashi, M.; Komiyama, K.; Ishibashi, M. Dihydrolindbladiones, three new naphthoquinone pigments from a myxomycete Lindbladia tubulina. J. Nat. Prod. 2003, 66, 999–1001. [Google Scholar] [CrossRef]
- Shintani, A.; Yamazaki, H.; Yamamoto, Y.; Ahmed, F.; Ishibashi, M. Cribrarione C, a naphthoquinone pigment from the myxomycete Cribraria meylanii. Chem. Pharm. Bull. 2009, 57, 894–895. [Google Scholar] [CrossRef] [PubMed]
- Naoe, A.; Ishibashi, M.; Yamamoto, Y. Cribrarione A, a new antimicrobial naphthoquinone pigment from a myxomycete Cribraria purpurea. Tetrahedron 2003, 59, 3433–3435. [Google Scholar] [CrossRef]
- Ishibashi, M.; Iwasaki, T.; Imai, S.; Sakamoto, S.; Yamaguchi, K.; Ito, A. Laboratory culture of the myxomycetes: Formation of fruiting bodies of Didymium bahiense and its plasmodial production of makaluvamine A. J. Nat. Prod. 2001, 64, 108–110. [Google Scholar] [CrossRef]
- Misono, Y.; Ishibashi, M.; Ito, A. Bahiensol, a new glycerolipid from a cultured myxomycete Didymium bahiense var. bahiense. Chem. Pharm. Bull. 2003, 51, 612–613. [Google Scholar] [CrossRef] [PubMed]
- Nakatani, S.; Kiyota, M.; Matsumoto, J.; Ishibashi, M. Pyrroloiminoquinone pigments from Didymium iridis. Biochem. Syst. Ecol. 2005, 33, 323–325. [Google Scholar] [CrossRef]
- Ishibashi, M.; Mitamura, M.; Ito, A. Laboratory culture of the myxomycete Didymium squamulosum and its production of clionasterol. Nat. Med. 1999, 53, 316–318. [Google Scholar] [CrossRef]
- Nakatani, S.; Yamamoto, Y.; Hayashi, M.; Komiyama, K.; Ishibashi, M. Cycloanthranilylproline-derived constituents from a myxomycete Fuligo candida. Chem. Pharm. Bull. 2004, 52, 368–370. [Google Scholar] [CrossRef]
- Hasegawa, H.; Yamada, Y.; Komiyama, K.; Hayashi, M.; Ishibashi, M.; Sunazuka, T.; Izuhara, T.; Sugahara, K.; Tsuruda, K.; Masuda, M.; et al. A novel natural compound, a cycloanthranilylproline derivative (Fuligocandin B), sensitizes leukemia cells to apoptosis induced by tumor necrosis factor related apoptosis-inducing ligand (TRAIL) through 15-deoxy-Delta 12, 14 prostaglandin J2 production. Blood 2007, 110, 1664–1674. [Google Scholar] [CrossRef] [PubMed]
- Řezanka, T.; Hanuš, L.O.; Dvořáková, R.; Dembitsky, V.M. The fulicineroside, a new unusual glycosidic dibenzofuran metabolite from the slime mold Fuligo cinerea. Wiggers Eur. J. Org. Chem. 2005, 13, 2708–2714. [Google Scholar] [CrossRef]
- Casser, I.; Steffan, B.; Steglich, W. The Chemistry of the Plasmodial Pigments of the Slime Mold Fuligo septica (Myxomycetes). Angew. Chem. 1987, 26, 586–587. [Google Scholar] [CrossRef]
- Shintani, A.; Toume, K.; Yamamoto, Y.; Ishbibashi, M. Dehydrofuligoic acid, a new yellow pigment isolated from the myxomycete Fuligo septica f. flava. Heterocycles 2010, 82, 839–842. [Google Scholar] [CrossRef]
- Sobels, J.C. Nutrition de quelque myxomycetes en cultures et associees et leurs proprietes antibiotiques. Antonie Van Leeuwenhoek J. Microbiol. Serol. 1950, 16, 123–243. [Google Scholar] [CrossRef]
- Ishikawa, Y.; Ishibashi, M.; Yamamoto, Y.; Hayashi, M.; Komiyama, K. Lindbladione and related naphthoquinone pigments from a myxomycete Lindbladia tubulina. Chem. Pharm. Bull. 2002, 50, 1126–1127. [Google Scholar] [CrossRef]
- Arai, M.A.; Makita, Y.; Yamaguchi, Y.; Kawano, H.; Suganami, A.; Tamura, Y.; Ishibashi, M. Total synthesis of lindbladione, a Hes1 dimerization inhibitor and neural stem cell activator isolated from Lindbladia tubulina. Sci. Rep. 2020, 10, 21433. [Google Scholar] [CrossRef] [PubMed]
- Hosoya, T.; Yamamoto, Y.; Uehara, Y.; Hayashi, M.; Komiyama, K.; Ishibashi, M. New cytotoxic bisindole alkaloids with protein tyrosine kinase inhibitory activity from a myxomycete Lycogala epidendrum. Bioorg. Med. Chem. Lett. 2005, 15, 2776–2780. [Google Scholar] [CrossRef]
- Fröde, R.; Hinze, C.; Josten, I.; Schmidt, B.; Steffan, B.; Steglich, W. Isolation and synthesis of 3,4-bis(indol-3-yl)pyrrole-2,5-dicarboxylic acid derivatives from the slime mould Lycogala epidendrum. Tetrahedron Lett. 1994, 35, 1689–1690. [Google Scholar] [CrossRef]
- Hashimoto, T.; Akazawa, A.; Tori, M.; Kan, Y.; Kusumi, T.; Takahashi, H.; Asakawa, Y. Three novel polyacetylene triglycerides, lycogarides A–C, from the myxomycetes Lycogala epidendrum. Chem. Pharm. Bull. 1994, 42, 1531–1533. [Google Scholar] [CrossRef]
- Buchanan, M.; Hashimoto, T.; Asakawa, Y. Acylglycerols from the slime mold, Lycogala epidendrum. Phytochemistry 1996, 41, 791–794. [Google Scholar] [CrossRef]
- Hashimoto, T.; Yasuda, A.; Akazawa, K.; Takaoka, S.; Tori, M.; Asakawa, Y. Three novel dimethyl pyrroledicarboxylate, lycogarubins A-C, from the myxomycetes Lycogala epidendrum. Tetrahedron Lett. 1994, 35, 2559–2560. [Google Scholar] [CrossRef]
- Yasuzawa, T.; Iida, T.; Yoshida, M.; Hirayama, N.; Takahashi, M.; Shirahata, K.; Sano, H. The structures of the novel protein kinase C inhibitors K-252a, b, c and d. J. Antibiot. 1986, 39, 1072–1078. [Google Scholar] [CrossRef] [PubMed]
- Řezanka, T.; Dvořáková, R. Polypropionate lactones of deoxysugars glycosides from slime mold Lycogala epidendrum. Phytochemistry 2003, 63, 945–952. [Google Scholar] [CrossRef]
- Kopanski, L.; Li, G.; Besl, H.; Steglich, W. Pilzpigmente, 41. Naphthochinon-Farbstoffe aus den Schleimpilzen Trichia floriformis und Metatrichia vesparium (Myxomycetes). Liebigs Ann. Chem. 1982, 1982, 1722–1729. [Google Scholar] [CrossRef]
- Kopanski, L.; Karbach, G.; Selbitschka, G.; Steglich, W. Pilzfarbstoffe, 53. Vesparion, ein Naphtho(2,3-b)pyrandion-Derivat aus dem Schleimpilz Metatrichia vesparium (Myxomycetes). Liebigs Ann. Chem. 1987, 1987, 793–796. [Google Scholar] [CrossRef]
- Steffan, B.; Praemassing, M.; Steglich, W. Physarochrome A, a plasmodial pigment from the slime mould Physarum polycephalum (Myxomycetes). Tetrahedron Lett. 1987, 28, 3667. [Google Scholar] [CrossRef]
- Huynh, T.; Phung, T.; Stephenson, S.; Tran, H. Biological activities and chemical compositions of slime tracks and crude exopolysaccharides isolated from plasmodia of Physarum polycephalum and Physarella oblonga. BMC Biotechnol. 2017, 17, 76–86. [Google Scholar] [CrossRef]
- Nakatani, S.; Kamata, K.; Sato, M.; Onuki, H.; Hirota, H.; Matsumoto, J.; Ishibashi, M. Melleumin A, a novel peptide lactone isolated from the cultured myxomycete Physarum melleum. Tetrahedron Lett. 2005, 46, 267–271. [Google Scholar] [CrossRef]
- Płonka, P.M.; Rakoczy, L. Electron paramagnetic resonance spectroscopy (EPR) as a method for studying the biology of the acellular slime moulds (Myxomycetes). Acta Physiol. Plant 1997, 19, 233. [Google Scholar]
- Nowak, A.; Steffan, B. Polycephalin B and C: Unusual tetramic acids from plasmodia of the slime mold Physarum polycephalum (Myxomycetes). Angew. Chem. Int. Ed. Engl. 1998, 37, 3139–3141. [Google Scholar] [CrossRef]
- Blumenthal, F.; Polborn, K.; Steffan, B. The absolute configuration of polycephalin C from the slime mold Physarum polycephalum (Myxomycetes). Tetrahedron 2002, 58, 8433–8437. [Google Scholar] [CrossRef]
- Longbottom, D.; Morrison, A.; Dixon, D.; Ley, S. Total synthesis of the polyenoyltetramic acid polycephalin C. Tetrahedron 2003, 59, 6955–6966. [Google Scholar] [CrossRef]
- Eisenbarth, S.; Steffan, B. Structure and biosynthesis of chrysophysarin A, a plasmodial pigment from the slime mould Physarum polycephalum (myxomycetes). Tetrahedron 2000, 56, 363–365. [Google Scholar] [CrossRef]
- Kobayashi, S.; Mukai, M.; Murofushi, H.; Tigyi, G.; Murakami-Murofushi, K.; Uchiyama, A.; Fujiwara, Y.; Kobayashi, T. Biological functions of a novel lipid mediator, cyclic phosphatidic acid. Biochim. Biophys. Acta 2002, 1582, 1–7. [Google Scholar] [CrossRef]
- Misono, Y.; Ito, A.; Matsumoto, J.; Sakamoto, S.; Yamaguchi, K.; Ishibashi, M. Physarigins A–C, three new yellow pigments from a cultured myxomycete Physarum rigidum. Tetrahedron Lett. 2003, 44, 4479–4481. [Google Scholar] [CrossRef]
- Loganathan, P.; Paramasivan, P.; Kalyanasundaram, I. Melanin as the spore wall pigment of some myxomycetes. Mycol. Res. 1989, 92, 286–292. [Google Scholar] [CrossRef]
- Řezanka, T.; Dvoráková, R.; Hanus, L.O.; Dembitsky, V.M. Enteridinines A and B from slime mold Enteridium lycoperdon. Phytochemistry 2004, 65, 455–462. [Google Scholar] [CrossRef]
- Loganathan, L.; Kalyanasundram, I. The melanin of the myxomycete Stemonitis herbatica. Acta Protozool. 1999, 38, 97–103. [Google Scholar]
- Kaniwa, K.; Ohtsuki, T.; Yamamoto, Y.; Ishibashi, M. Kehokorins A-C, novel cytotoxic dibenzofurans isolated from the myxomycete Trichia favoginea var. persimilis. Tetrahedron Lett. 2006, 47, 1505–1508. [Google Scholar] [CrossRef]
- Kamata, K.; Onuki, H.; Hirota, H.; Yamamoto, Y.; Hayashi, M.; Komiyama, K.; Sato, M.; Ishibashi, M. Tubiferal A, a backbone-rearranged triterpenoid lactone isolated from the myxomycete Tubifera dimorphotheca, possessing reversal of drug resistance activity. Tetrahedron 2004, 60, 9835–9839. [Google Scholar] [CrossRef]
- Kikuchi, H.; Saito, Y.; Sekiya, J.; Okano, Y.; Saito, M.; Nakahata, N.; Kubohara, Y.; Oshima, Y. Isolation and synthesis of a new aromatic compound, brefelamide, from Dictyostelium cellular slime molds and its inhibitory effect on the proliferation of astrocytoma cells. J. Org. Chem. 2005, 70, 8854–8858. [Google Scholar] [CrossRef]
- Takaya, Y.; Kikuchi, H.; Terui, Y.; Komiya, J.; Furukawa, K.I.; Seya, K.; Motomura, S.; Ito, A.; Oshima, Y. Novel acyl alpha-pyronoids, dictyopyrone A, B, and C, from Dictyostelium cellular slime molds. J. Org. Chem. 2000, 65, 985–989. [Google Scholar] [CrossRef]
- Staples, S.O.; Gregg, J.H. Carotenoid pigments in the cellular slime mold, Dictyostellium discoideum. Biol. Bull. 1967, 132, 413–422. [Google Scholar] [CrossRef]
- Davidoff, F.; Korn, E. Fatty acid and phospholipid composition of cellular slime mold, Dictyostelium discoideum—Occurrence of previously undescribed fatty acids. J. Biol. Chem. 1963, 238, 3199–3209. [Google Scholar] [CrossRef]
- Saito, T.; Ochiai, H. Identification of Delta5-fatty acid desaturase from the cellular slime mold Dictyostelium discoideum. Eur. J. Biochem. 1999, 265, 809–814. [Google Scholar] [CrossRef]
- Murata, C.; Ogura, T.; Narita, S.; Kondo, A.; Iwasaki, N.; Saito, T.; Usuki, T. Synthesis and SAR of 4-methyl-5-pentylbenzene-1,3-diol (MPBD), produced by Dictyostelium discoideum. Bioorganic Med. Chem. Lett. 2016, 26, 1428–1433. [Google Scholar] [CrossRef] [PubMed]
- Adelman, M.R.; Taylor, E.W. Further purification and characterization of slime mold myosin and slime mold actin. Biochemistry 1969, 8, 4976–4988. [Google Scholar] [CrossRef] [PubMed]
- Yumura, S.; Uyeda, T.Q. Myosins and cell dynamics in cellular slime molds. Int. Rev. Cytol. 2003, 224, 173–225. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, H.; Nakamura, K.; Kubohara, Y.; Gokan, N.; Hosaka, K.; Maeda, Y.; Oshima, Y. Dihydrodictyopyrone A and C: New members of dictyopyrone family isolated from Dictyostelium cellular slime molds. Tetrahedron Lett. 2007, 48, 5905–5909. [Google Scholar] [CrossRef]
- Nguyen, V.H.; Kikuchi, H.; Sasaki, H.; Iizumi, K.; Kubohara, Y.; Oshima, Y. Production of novel bispyrone metabolites in the cellular slime mold Dictyostelium giganteum induced by zinc (II) ion. Tetrahedron 2017, 73, 583–588. [Google Scholar] [CrossRef]
- Takaya, Y.; Kikuchi, H.; Terui, Y.; Komiya, J.; Maeda, Y.; Itoc, A.; Oshima, Y. Novel aromatic substances, dictyomedin A and B, from Dictyostelium cellular slime molds and their inhibitory effects on Dictyostelium development. Tetrahedron Lett. 2001, 42, 61–63. [Google Scholar] [CrossRef]
- Sawada, T.; Aono, M.; Asakawa, S.; Ito, A.; Awano, K. Structure determination and total synthesis of a novel antibacterial substance, AB0022A, produced by a cellular slime mold. J. Antibiot. 2000, 53, 959–966. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, H.; Kubohara, Y.; Nguyen, V.; Katou, Y.; Oshima, Y. Novel chlorinated dibenzofurans isolated from the cellular slime mold, Polysphondylium filamentosum, and their biological activities. Bioorg. Med. Chem. 2013, 21, 4628–4633. [Google Scholar] [CrossRef]
- Saito, T.; Ochiai, H. Identification of a novel all-cis-5,9,12- heptadecatrienoic acid in the cellular slime mold Polysphondylium pallidum. Lipids 1996, 31, 445–447. [Google Scholar] [CrossRef]
- Saito, T.; Ochiai, H. Fatty acid composition of the cellular slime mold Polysphondylium pallidum. Lipids 1998, 33, 327–332. [Google Scholar] [CrossRef]
- Kikuchi, H.; Ishiko, S.; Nakamura, K.; Kubohara, Y.; Oshima, Y. Novel prenylated and geranylated aromatic compounds isolated from Polysphondylium cellular slime molds. Tetrahedron 2010, 66, 6000–6007. [Google Scholar] [CrossRef]
- Ley, S.; Smith, S.; Woodward, P. Use of t-butyl 4-diethylphosphono-3-oxobutanethioate for tetramic acid synthesis: Total synthesis of the plasmodial pigment fuligorubin A. Tetrahedron Lett. 1988, 29, 5829–5832. [Google Scholar] [CrossRef]
- Physarum polycephalum. Available online: https://en.wikipedia.org/wiki/Physarum_polycephalum (accessed on 10 November 2022).
- Zhou, Z.-Y.; Liu, J.-K. Pigments of fungi (macromycetes). Nat. Prod. Rep. 2010, 11. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin pigmentation in mammalian skin and its hormonal regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.M.; Jimbow, K.; Boissy, R.E.; Slominski, A.; Plonka, P.M.; Slawinski, J.; Wortsman, J.; Tosk, J. What is the use of generating melanin? Exp. Dermatol. 1999, 8, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Plonka, P.M.; Grabacka, M. Melanin synthesis in microorganisms—Biotechnological and medical aspects. Acta Biochim. Pol. 2006, 53, 429–443. [Google Scholar] [CrossRef] [PubMed]
- Płonka, P.M.; Rakoczy, L. Heme and non-heme iron complexes of nitric oxide in the plasmodia of acellular slime moulds cultured in vitro. ZeszProbl. Post N. Roln. 2000, 473, 249–259. [Google Scholar] [CrossRef]
- Rakoczy, L.; Płonka, P.M. Accumulation of manganese in plasmodia of the acellular slime mould (Myxomycetes) Metatrichia vesparium. Ochr Środ Zas. Nat. 1997, 18, 299–308. [Google Scholar]
- Du, Y.L.; Ryan, K.S. Expansion of bisindole biosynthetic pathways by combinatorial construction. ACS Synth. Biol. 2015, 4, 682–688. [Google Scholar] [CrossRef]
- Chambers, G.E.; Sayan, A.E.; Brown, R.C.D. The synthesis of biologically active indolocarbazole natural products. Nat. Prod. Rep. 2021, 38, 1794. [Google Scholar] [CrossRef]
- Sartori, S.K.; Diaz, M.A.N.; Diaz-Muñoz, G. Lactones: Classification, synthesis, biological activities, and industrial applications. Tetrahedron 2021, 84, 132001. [Google Scholar] [CrossRef]
- Kim, Y.; Sengupta, S.; Sim, T. Natural and synthetic lactones possessing antitumor activities. Int. J. Mol. Sci. 2021, 22, 1052. [Google Scholar] [CrossRef]
- Ferraz, H.M.C.; Bombonato, F.I.; Sano, M.K.; Longo, L.S. Natural occurrence, biological activities and synthesis of eight-, nine-, and eleven membered ring lactones. Quim. Nova 2008, 31, 885–900. [Google Scholar] [CrossRef]
- Leontyev, D.V.; Schnittler, M.; Stephenson, S.L.; Novozhilov, Y.K. Systematic revision of the Tubifera casparyi–T. dictyoderma complex: Resurrection of the genus Siphoptychium and introduction of the new genus Thecotubifera. Mycologia 2019, 111, 981–997. [Google Scholar] [CrossRef]
- Lycogala epidendrum. Available online: https://en.wikipedia.org/wiki/Lycogala_epidendrum (accessed on 9 November 2022).
- Volvoikar, P.; Torne, P. An account of synthetic strategies towards indolocarbazole alkaloids: Arcyriaflavin A and staurosporinone. Tetrahedron 2021, 82, 131756. [Google Scholar] [CrossRef]
- Malsy, M.; Bitzinger, D.; Graf, B.; Bundscherer, A. Staurosporine induces apoptosis in pancreatic carcinoma cells PaTu 8988t and Panc-1 via the intrinsic signaling pathway. Eur. J. Med. Res. 2019, 24, 5. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, S.; Matsuda, Y.; Iwahashi, K.; Kase, H. K-252b, c and d, potent inhibitors of protein kinase C from microbial origin. J. Antibiot. 1986, 39, 1066–1071. [Google Scholar] [CrossRef]
- Arai, M.; Seto, J.; Ahmed, F.; Uchiyama, K.; Ishibashi, M. Practical Total Synthesis of Fuligocandins A and B. Synlett 2010, 16, 2498–2502. [Google Scholar] [CrossRef]
- Barrows, L.; Radisky, D.; Copp, B.; Swaffar, D.; Kramer, R.; Warters, R.; Ireland, C. Makaluvamines, marine natural products, are active anti-cancer agents and DNA topo II inhibitors. Anticancer Drug Des. 1993, 8, 333–347. [Google Scholar]
- Chen, Y.; Li, J.; Fan, K.; Du, Y.; Ren, Z.; Xu, J.; Zheng, J.; Liu, Y.; Fu, J.; Ren, D.; et al. Mutations in the maize zeta-carotene desaturase gene lead to viviparous kernel. PLoS ONE 2017, 12, e017427. [Google Scholar] [CrossRef]
- Płonka, P.M.; Rakoczy, L. Usefulness of the EPR method in the research on slime moulds. In Application of the In Vitro Cultures in Plant Physiology, 1st ed.; Dubert, F., Skoczowski, A., Eds.; The Franciszek Górski Department of Plant Physiology, Polish Academy of Science: Krakow, Poland, 1997; pp. 175–180. [Google Scholar]
- Rakoczy, L.; Panz, T. Melanin revealed in spores of true slime moulds using the electron paramagnetic resonance method. Acta Protozool. 1994, 33, 227–231. [Google Scholar]
- Yang, M.; Zhou, M.; Song, L. A review of fatty acids influencing skin condition. J. Cosmet. Dermatol. 2020, 19, 3199–3244. [Google Scholar] [CrossRef]
- Korn, E.D.; Greenblatt, C.L.; Lees, A.M. Synthesis of unsaturated fatty acids in the slime mold Physarum polycephalum and the zooflagellates Leishmania tarentolae, Trypanosoma lewisi, and Crithidia sp.: A comparative study. J. Lipid Res. 1965, 6, 43–45. [Google Scholar] [CrossRef] [PubMed]
- Abedin, M.R.; Barua, S. Isolation and purification of glycoglycerolipids to induce apoptosis in breast cancer cells. Sci. Rep. 2021, 11, 1298. [Google Scholar] [CrossRef] [PubMed]
- O’Lenick, A.J.; Garrison, M. Comparatively Speaking: Esters vs. Lactones. Cosmetics & Toiletries, 27 January 2009. Available online: https://www.cosmeticsandtoiletries.com/research/literature-data/article/21834303/comparatively-speaking-esters-vs-lactones (accessed on 15 November 2022).
- Enteridium lycoperdon. Available online: https://en.wikipedia.org/wiki/Enteridium_lycoperdon (accessed on 10 November 2022).
- Lycogalinoside, A. Available online: https://www.wikidata.org/wiki/Q57281678 (accessed on 11 November 2022).
- Wang, Q.; Li, Y.; Liu, P. Physiology and biochemistry of Myxomycetes. In Myxomycetes: Biology, Systematics, Biogeography, and Ecology; Elsevier Inc.: Amsterdam, The Netherlands, 2017; Volume 193, pp. 231–267. [Google Scholar] [CrossRef]
- Azelmat, J.; Fiorito, S.; Genovese, S.; Epifano, F.; Grenier, D. Antibacterial and anti-inflammatory activities of Ppc-1, active principle of the cellular slime mold Polysphondylium pseudocandidum. Med. Chem. 2015, 11, 666–669. [Google Scholar] [CrossRef] [PubMed]
- Czumaj, A.; Śledziński, T. Biological role of unsaturated fatty acid desaturases in health and disease. Nutrients 2020, 12, 356. [Google Scholar] [CrossRef]
- Liakath-Ali, K.; Vancollie, V.E.; Sequeira, I.; Lelliott, C.J.; Watt, F.M. Myosin 10 is involved in murine pigmentation. Exp. Dermatol. 2019, 28, 391–394. [Google Scholar] [CrossRef]
- Androutsopoulou, C.; Lamari, F.; Christopoulou, S.; Hahalis, P.; Kotsalou, C.; Vantarakis, A. Evaluation of essential oils and extracts of rose geranium and rose petals as natural preservatives in terms of toxicity, antimicrobial, and antiviral activity. Pathogens 2021, 10, 494. [Google Scholar] [CrossRef]
- Deitz, B. What are terpenes and how are they used in skin care? Coveteur, 27 September 2021. Available online: https://coveteur.com/terpenes-skin-benefits (accessed on 16 November 2022).
- Honma, S.; Kouno, K.; Takasaka, S.; Mitazaki, S.; Abe, S.; Kikuchi, H.; Oshima, Y.; Yoshida, M. Effect of brefelamide on proliferation of 1321N1 human astrocytoma cells induced by glial cell line-derived neurotrophic factor. Pharmazie 2018, 73, 22–28. [Google Scholar] [CrossRef]
- Honma, S.; Saito, M.; Kikuchi, H.; Saito, Y.; Oshima, Y.; Nakahata, N.; Yoshida, M. A reduction of epidermal growth factor receptor is involved in brefelamide-induced inhibition of phosphorylation of ERK in human astrocytoma cells. Eur. J. Pharmacol. 2009, 616, 38–42. [Google Scholar] [CrossRef]
- Honma, S.; Takasaka, S.; Ishikawa, T.; Shibuya, T.; Mitazaki, S.; Abe, S.; Yoshida, M. Effect of brefelamide on HGF-induced survival of 1321N1 human astrocytoma cells. In Vitro Cell Dev. Biol. Anim. 2016, 52, 705–711. [Google Scholar] [CrossRef]
- Lierl, M. Myxomycete (slime mold) spores: Unrecognized aeroallergens? Ann. Allergy Asthma Immunol. 2013, 111, 537–541. [Google Scholar] [CrossRef]
- Gianini, E.; Northy, W.; Leathers, C. The allergenic significance of certain fungi rarely reported as allergens. Ann. Allergy 1975, 35, 372–376. [Google Scholar]
- Santilli, J., Jr.; Rockwell, W.J.; Collins, R.P. The significance of the spores of the Basidiomycetes (mushrooms and their allies) in bronchial asthma and allergic rhinitis. Ann. Allergy 1985, 55, 469–471. [Google Scholar] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class/Species | Chemical Compounds | References |
|---|---|---|
| Ceratiomyxomycetes (Protostelids) | ||
| Ceratiomyxa fruticulosa T. Macbr. | ceratiopyrone | [30,50] |
| Ceratiomyxa sp. | arcyrioxocin A (?) | [43] |
| Myxomycetes (Myxogastrids) | ||
| Arcyria cinerea (Bull.) Pers. | cinereapyrroles A-B, arcyriarubin A; 6,9′-dihydroxystaurosporinone; 74 FA; 4 GL | [32,51,52,53,54,55] |
| Arcyria denudata Fr. | arcyriacyanin A, dihydroarcyrioxocin A, arcyroxocins A-B, arcyroxindole A, arcyriarubins A-C, arcyriaflavins A-C, arcyrioxepins A-B; arcyriabun B 6-O-sulfate; 74 FA; 4 GL | [30,31,54,55,56,57,58] |
| Arcyria ferruginea Saut. | arcyriarubin C, arcyriaflavin C, dihydroarcyriarubin C | [31,52,59,60] |
| Arcyria obvelata (Oeder) Obsberg | arcyriaflavins A-C, CDD of arcyriacyanin A and arcyrioxocin A; dihydroarcyriacyanin A; 74 FA; 4 GL | [30,31,43,54,55,58] |
| Arcyria sp. | 12 PHL | [31] |
| Cribraria cancellata (Batsch) Nann.-Bremek. | cribrarione B | [61] |
| Cribraria intricata Schrad. | lindbladione | [62] |
| Cribraria meylanii Brândză | cribrarione C | [63] |
| Cribraria purpurea (Fr.) Donk. | cribrarione A | [64] |
| Dictydiaethalium plumbeum (Schumach.) Rostaf. ex Lister | arcyriaflavin D | [30] |
| Didymium bahiense Gottsb. | makaluvamines A-B; bahiensol | [31,65,66] |
| Didymium iridis (Ditmar) Fr. | makaluvamine A, damirone C | [52,65,67] |
| Didymium squamulosum (Alb. et Schwein.) Fr. et Palmquist | clionasterol | [68] |
| Fuligo candida Pers. | fuligocandins A-B, FCB; 4-aminobenzoyltryptophan | [52,69,70] |
| Fuligo cinerea (Schwein.) Morgan | fulicineroside | [71] |
| Fuligo septica (L.) F. H. Wigg | fuligorubin A, fuligopyrone; fuligoic acid; 74 FA, 12 PHL; 4 GL | [28,30,31,43,54,55,72,73] |
| Leocarpus fragilis (Dicks.) Rostaf. | acyltetramic acids | [30] |
| Licea variabilis Schrad. | EPS | [29,74] |
| Lindbladia tubulina Fr. | lindbladione, lindbladiapyrone, 6,7-dimethoxydihydrolindbladione, dihydrolindbladione, 6-methoxydihydro-lindbladione, 7-methoxyl-lindbladione, 6,7-metoxylindbladione | [30,52,62,75,76] |
| Lycogala epidendrum (J.C. Buxb. ex L.) Fr. | 6-hydroxylstaurosporinone; 5,6-dihydroxyarcyriaflavin A; arcyriaflavins A-B; lycogaric acid; lycogarubins A-C with 2 derivatives, staurosporinone (K-252); (Z)-methyl-2-hydroxy-3-(1H-indol-3-yl)acrylate; lycogarides A-G, 74 FA; 4 GL; lycogalinosides A-B | [31,51,52,54,55,77,78,79,80,81,82,83] |
| Lycogala flavofuscum (Ehrenb.) Rostaf. | 74 FA | [31,54] |
| Metatrichia floriformis (Schwein.) Nann.-Bremek. | trichione, homotrichione; TF-1; 2,3,5-trihydroxynaphthoquinone | [30,84,85] |
| Metatrichia vesparia (Batsch) Nann.-Bremek. ex G.W. Martin et Alexop. | arcyriaflavin C; trichione; homotrichione; vesparione | [29,84,85,86] |
| Ophiotheca chrysosperma (Curr.) Lister. | arcyriaflavin B; arcyriaflavins A and C(?); 6-hydroxylstaurosporinone; 6-hydroxy-9′-methoxystaurosporinone | [43,53] |
| Physarella oblonga (Berk. et M.A. Curtis) Morgan | EPS | [87] |
| Physarum melleum (Berk. and Broome) Massee | melleumins A-B | [52,88] |
| Physarum nudum T. Macbr. | melanin | [25,89] |
| Physarum polycephalum Schwein. | Polycephalins B-C, physarorubinic acids A-B, chrysophysarin A; physarochrome A; FA; stigmasterol, b-sitosterol, stigmastanol, campesterol, campestanol, cholesterol, lanosterol, 24-merhylene dihydrolanosterol; 12 PHL; EPS | [31,54,55,86,87,88,90,91,92,93,94] |
| Physarum rigidum (G. Lister) G. Lister | physarigins A-C | [52,95] |
| Physarum sp. | 74 FA | [31,54] |
| Reticularia lycoperdon Bull. | Ethanol-extractable yellow-green pigment; enteridinines A-B | [96,97] |
| Siphoptychium casparyi (Rostaf.) T. Macbr. | Arcyriaflavins B-C | [59] |
| Stemonitis herbatica Peck | melanin | [98] |
| Trichia favoginea (Batsch) Pers. | 74 FA; all-cis 5,9–16:2, 5,9–18:2, 5,11,14–20:3, 5,11,14,17–20:4, 7,13–22:2, and 7,15–22:2 | [31,54] |
| Trichia persimilis P. Karst. | Kehokorins A-C | [99] |
| Trichia varia (Pers. Ex J.F. Gmel.) Pers. | 74 FA; all-cis 5,9–16:2, 5,9–18:2, 5,11,14–20:3, 5,11,14,17–20:4, 7,13–22:2, and 7,15–22:2; 4 GL | [31,54,55] |
| Trichia sp. | trichione; homotrichione | [84,85] |
| Tubifera dimorphotheca Nann.-Bremek. et Loer. | Tubiferals A-B | [52,100] |
| Dictyosteliomycetes (Dictyostelids) | ||
| Dictyostelium brefeldianum H. Hagiw. | Brefelamide | [101] |
| Dictyostelium discoideum Raper | dictyopyrones A-B, zeta-carotenes; unique dienoic acids 5,9-16:2; 5,9-18:2; 5,11-18:2; 12 FA; stigmast-22-en-3b-ol; Delta-5 desaturase, myosin, MPBD | [31,102,103,104,105,106,107,108] |
| Dictyostelium firmibasis H. Hagiw. | Dihydrodictyopyrones A, C; firmibasiol | [10,109] |
| Dictyostelium giganteum B. N. Singh | dictyobispyrones B, E; brefelamide | [110] |
| Dictyostelium longosporum H. Hagiw. | Dictyopyrone C | [102] |
| “Dictyostelium medium” (? err. typogr.) | dictyomedins A-B | [43,111] |
| Dictyostelium mucoroides Bref. | mucoroidiol | [10] |
| Dictyostelium purpureum Olive | AB0022A | [112] |
| Hagiwaraea rhizopodium (Raper and Fennell) S. Baldauf, S. Sheikh et Thulin | dictyopyrones A-B | [102] |
| Heterostelium filamentosum (F. Traub, H.R. Hohl and Cavender) S. Baldauf, S. Sheikh et Thulin | Pf-1, Pf-2 | [113] |
| Heterostelium pallidum (Olive) S. Baldauf, S. Sheikh et Thulin | all-cis-5,9,12-heptadecatrienoic acid | [114,115] |
| Heterostelium pseudocandidum (H. Hagiw.) S. Baldauf, S. Sheikh et Thulin | Ppc-1 | [116] |
| Heterostelium tenuissimum H. Hagiw. | Pt-1-5 | [116] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stoyneva-Gärtner, M.; Uzunov, B.; Androv, M.; Ivanov, K.; Gärtner, G. Potential of Slime Molds as a Novel Source for the Cosmetics Industry. Cosmetics 2023, 10, 3. https://doi.org/10.3390/cosmetics10010003
Stoyneva-Gärtner M, Uzunov B, Androv M, Ivanov K, Gärtner G. Potential of Slime Molds as a Novel Source for the Cosmetics Industry. Cosmetics. 2023; 10(1):3. https://doi.org/10.3390/cosmetics10010003
Chicago/Turabian StyleStoyneva-Gärtner, Maya, Blagoy Uzunov, Miroslav Androv, Kristian Ivanov, and Georg Gärtner. 2023. "Potential of Slime Molds as a Novel Source for the Cosmetics Industry" Cosmetics 10, no. 1: 3. https://doi.org/10.3390/cosmetics10010003
APA StyleStoyneva-Gärtner, M., Uzunov, B., Androv, M., Ivanov, K., & Gärtner, G. (2023). Potential of Slime Molds as a Novel Source for the Cosmetics Industry. Cosmetics, 10(1), 3. https://doi.org/10.3390/cosmetics10010003