COVID-19 in Light of Seasonal Respiratory Infections




Abstract
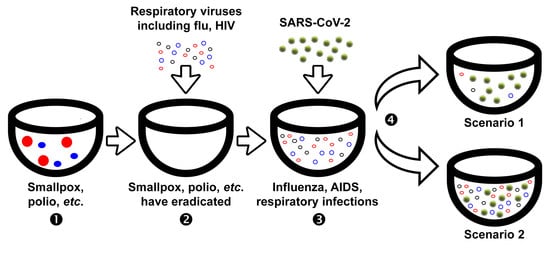
:
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Impact of Influenza Pandemics on the Circulation of Seasonal Respiratory Viruses
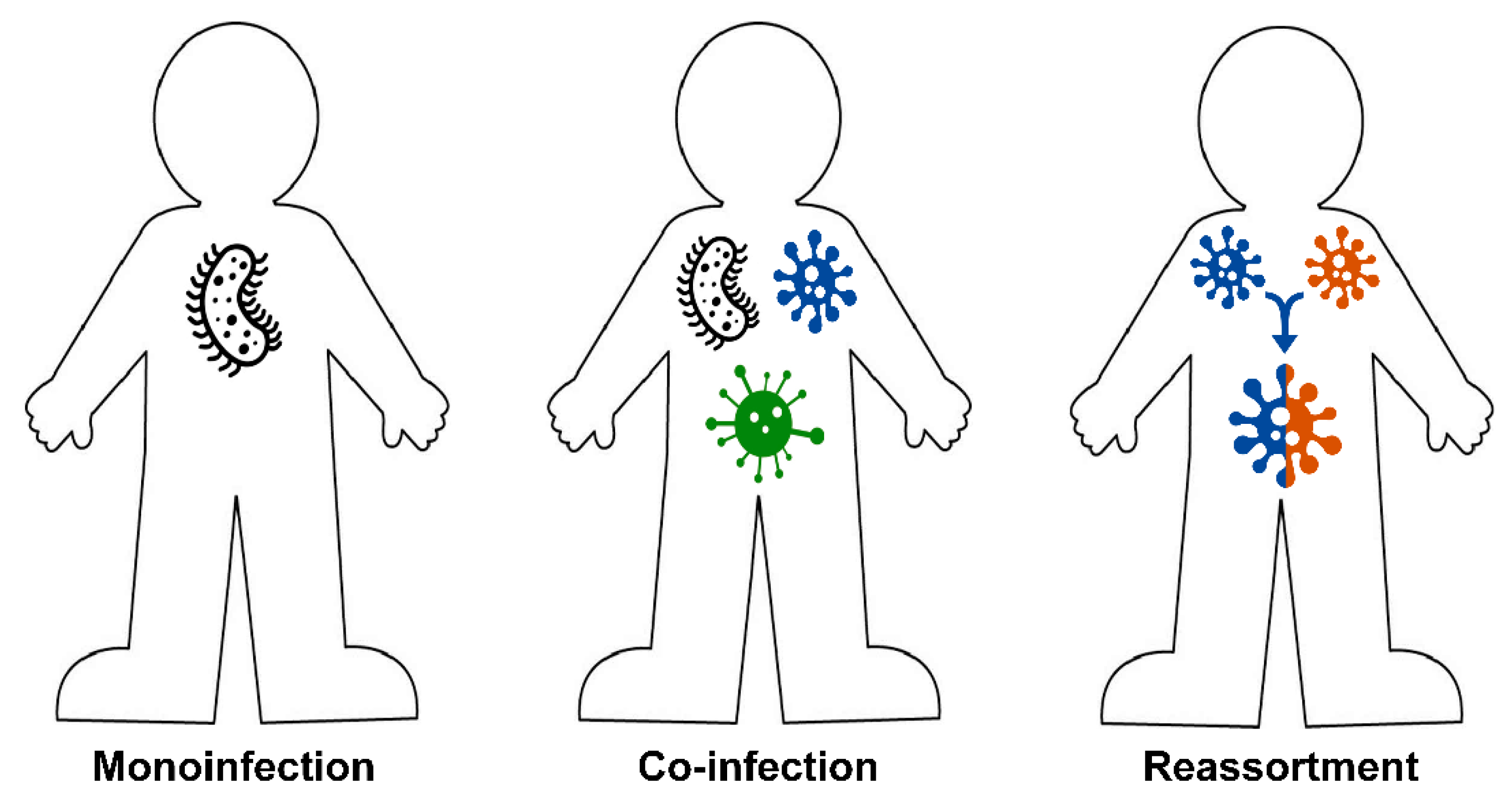
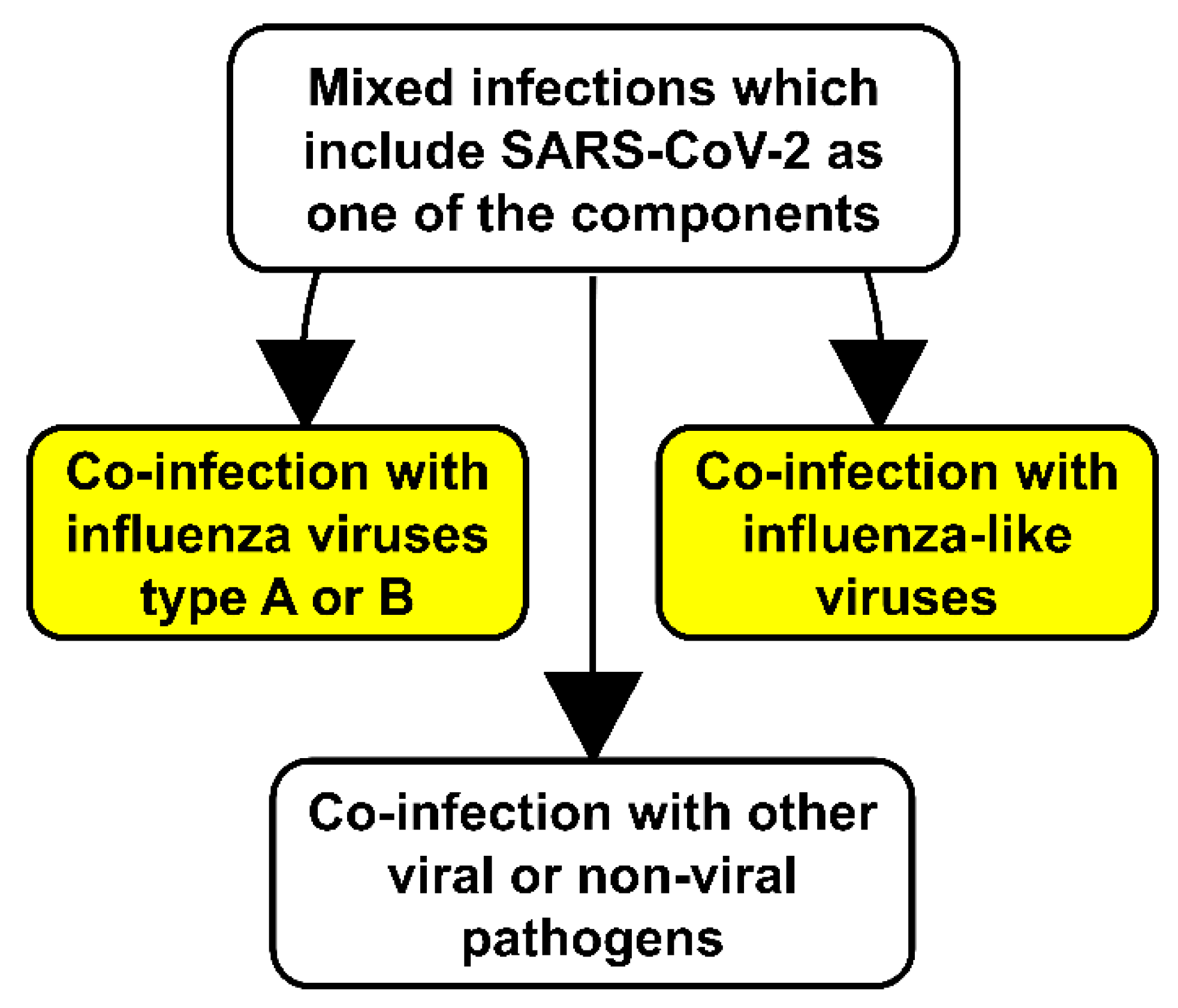
3. The Role of Mixed Infections in Etiology of Pulmonary Diseases
4. Impact of the COVID-19 Pandemic on the Circulation of Seasonal Respiratory Viruses
4.1. Co-Infection of SARS-CoV-2 with Influenza Viruses
4.2. Co-Infection of SARS-CoV-2 with Influenza-like Viruses
4.3. Co-Infection of SARS-CoV-2 with Other Viral and Nonviral Pathogens
5. On the Way to the Anti-COVID-19 Vaccine
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Belser, J.A. Assessment of SARS-CoV-2 replication in the context of other respiratory viruses. Lancet. Respir. Med. 2020. [Google Scholar] [CrossRef]
- Lowen, A.C.; Mubareka, S.; Tumpey, T.M.; Garcia-Sastre, A.; Palese, P. The guinea pig as a transmission model for human influenza viruses. Proc. Natl. Acad. Sci. USA 2006, 103, 9988–9992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bengoechea, J.A.; Bamford, C.G.G. SARS-CoV-2, bacterial co-infections, and AMR: The deadly trio in COVID-19? EMBO Mol. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- WHO. Coronavirus Disease (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 12 August 2020).
- Neumann, G.; Noda, T.; Kawaoka, Y. Emergence and pandemic potential of swine-origin H1N1 influenza virus. Nature 2009, 459, 931–939. [Google Scholar] [CrossRef] [Green Version]
- Flahault, A.; Zylberman, P. Influenza pandemics: Past, present and future challenges. Public Health Rev. 2010, 32, 319–340. [Google Scholar] [CrossRef] [Green Version]
- Potter, C.W. A history of influenza. J. Appl. Microbiol. 2001, 91, 572–579. [Google Scholar] [CrossRef]
- Krammer, F.; Smith, G.J.D.; Fouchier, R.A.M.; Peiris, M.; Kedzierska, K.; Doherty, P.C.; Palese, P.; Shaw, M.L.; Treanor, J.; Webster, R.G.; et al. Influenza. Nat. Rev. Dis. Primers 2018, 4, 3. [Google Scholar] [CrossRef]
- Krammer, F.; Palese, P. Influenza virus hemagglutinin stalk-based antibodies and vaccines. Curr Opin Virol 2013, 3, 521–530. [Google Scholar] [CrossRef] [Green Version]
- Palese, P.; Wang, T.T. Why do influenza virus subtypes die out? A hypothesis. mBio 2011, 2. [Google Scholar] [CrossRef] [Green Version]
- Dawood, F.S.; Iuliano, A.D.; Reed, C.; Meltzer, M.I.; Shay, D.K.; Cheng, P.Y.; Bandaranayake, D.; Breiman, R.F.; Brooks, W.A.; Buchy, P.; et al. Estimated global mortality associated with the first 12 months of 2009 pandemic influenza A H1N1 virus circulation: A modelling study. Lancet. Infect. Dis. 2012, 12, 687–695. [Google Scholar] [CrossRef] [Green Version]
- Simonsen, L.; Spreeuwenberg, P.; Lustig, R.; Taylor, R.J.; Fleming, D.M.; Kroneman, M.; Van Kerkhove, M.D.; Mounts, A.W.; Paget, W.J. Global mortality estimates for the 2009 influenza pandemic from the GLaMOR project: A modeling study. PLoS Med. 2013, 10, e1001558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meningher, T.; Hindiyeh, M.; Regev, L.; Sherbany, H.; Mendelson, E.; Mandelboim, M. Relationships between A(H1N1)pdm09 influenza infection and infections with other respiratory viruses. Influenza Other Respir. Viruses 2014, 8, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Marí, J.M.; Pérez-Ruiz, M.; Galán Montemayor, J.C.; Marcos Maeso, M.; Reina, J.; de Oña Navarro, M.; Cilla Eguiluz, C.G. Circulation of other respiratory viruses and viral co-infection during the 2009 pandemic influenza. Enferm. Infecc. Y Microbiol. Clin. 2012, 30 (Suppl. 4), 25–31. [Google Scholar] [CrossRef]
- Casalegno, J.S.; Ottmann, M.; Bouscambert-Duchamp, M.; Valette, M.; Morfin, F.; Lina, B. Impact of the 2009 influenza A(H1N1) pandemic wave on the pattern of hibernal respiratory virus epidemics, France, 2009. Eur. Commun. Dis. Bull. 2010, 15. [Google Scholar]
- Mak, G.C.; Wong, A.H.; Ho, W.Y.; Lim, W. The impact of pandemic influenza A (H1N1) 2009 on the circulation of respiratory viruses 2009-2011. Influenza Other Respir. Viruses 2012, 6, e6–e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ånestad, G.; Nordbø, S.A. Virus interference. Did rhinoviruses activity hamper the progress of the 2009 influenza A (H1N1) pandemic in Norway? Med Hypotheses 2011, 77, 1132–1134. [Google Scholar] [CrossRef]
- Diaz-Munoz, S.L. Viral coinfection is shaped by host ecology and virus-virus interactions across diverse microbial taxa and environments. Virus Evol. 2017, 3, vex011. [Google Scholar] [CrossRef] [Green Version]
- Alcaide, C.; Rabadan, M.P.; Moreno-Perez, M.G.; Gomez, P. Implications of mixed viral infections on plant disease ecology and evolution. Adv. Virus Res. 2020, 106, 145–169. [Google Scholar] [CrossRef]
- Maurice, J.B.; Patel, A.; Scott, A.J.; Patel, K.; Thursz, M.; Lemoine, M. Prevalence and risk factors of nonalcoholic fatty liver disease in HIV-monoinfection. Aids 2017, 31, 1621–1632. [Google Scholar] [CrossRef]
- Nowak, M.D.; Sordillo, E.M.; Gitman, M.R.; Paniz Mondolfi, A.E. Co-infection in SARS-CoV-2 infected patients: Where are influenza virus and rhinovirus/enterovirus? J. Med Virol. 2020. [Google Scholar] [CrossRef]
- Ou, X.; Zhou, L.; Huang, H.; Lin, Y.; Pan, X.; Chen, D. A severe case with co-infection of SARS-CoV-2 and common respiratory pathogens. Travel Med. Infect. Dis. 2020, 101672. [Google Scholar] [CrossRef] [PubMed]
- Ozaras, R.; Cirpin, R.; Duran, A.; Duman, H.; Arslan, O.; Bakcan, Y.; Kaya, M.; Mutlu, H.; Isayeva, L.; Kebanlı, F.; et al. Influenza and COVID-19 co-infection: Report of 6 cases and review of the Literature. J. Med Virol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Rawson, T.M.; Moore, L.S.P.; Zhu, N.; Ranganathan, N.; Skolimowska, K.; Gilchrist, M.; Satta, G.; Cooke, G.; Holmes, A. Bacterial and fungal co-infection in individuals with coronavirus: A rapid review to support COVID-19 antimicrobial prescribing. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Rodriguez, J.A.; Rubio-Gomez, H.; Roa, A.A.; Miller, N.; Eckardt, P.A. Co-Infection with SARS-COV-2 and parainfluenza in a young adult patient with pneumonia: Case report. IDCases 2020, 20, e00762. [Google Scholar] [CrossRef] [PubMed]
- Touzard-Romo, F.; Tapé, C.; Lonks, J.R. Co-infection with SARS-CoV-2 and human metapneumovirus. Rhode Island Med. J. 2020, 103, 75–76. [Google Scholar]
- Steel, J.; Lowen, A.C. Influenza A virus reassortment. Curr. Top. Microbiol. Immunol. 2014, 385, 377–401. [Google Scholar] [CrossRef]
- Klempa, B. Reassortment events in the evolution of hantaviruses. Virus Genes 2018, 54, 638–646. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, Y.; Sugawara, K.; Takashita, E.; Muraki, Y.; Hongo, S.; Katsushima, N.; Mizuta, K.; Nishimura, H. Genetic diversity of influenza B virus: The frequent reassortment and cocirculation of the genetically distinct reassortant viruses in a community. J. Med. Virol. 2004, 74, 132–140. [Google Scholar] [CrossRef]
- Brealey, J.C.; Sly, P.D.; Young, P.R.; Chappell, K.J. Viral bacterial co-infection of the respiratory tract during early childhood. FEMS Microbiol. Lett. 2015, 362. [Google Scholar] [CrossRef] [Green Version]
- Babiuk, L.A.; Lawman, M.J.; Ohmann, H.B. Viral-bacterial synergistic interaction in respiratory disease. Adv. Virus Res. 1988, 35, 219–249. [Google Scholar] [CrossRef]
- Jia, L.; Xie, J.; Zhao, J.; Cao, D.; Liang, Y.; Hou, X.; Wang, L.; Li, Z. Mechanisms of severe mortality-associated bacterial co-infections following influenza virus infection. Front. Cell. Infect. Microbiol. 2017, 7, 338. [Google Scholar] [CrossRef] [PubMed]
- Nickbakhsh, S.; Mair, C.; Matthews, L.; Reeve, R.; Johnson, P.C.D.; Thorburn, F.; von Wissmann, B.; Reynolds, A.; McMenamin, J.; Gunson, R.N.; et al. Virus-virus interactions impact the population dynamics of influenza and the common cold. Proc. Natl. Acad. Sci. USA 2019, 116, 27142–27150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloyd-Smith, J.O. Vacated niches, competitive release and the community ecology of pathogen eradication. Philos. Trans. R. Soc. Lond. Ser. BBiol. Sci. 2013, 368, 20120150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosch, A.A.; Biesbroek, G.; Trzcinski, K.; Sanders, E.A.; Bogaert, D. Viral and bacterial interactions in the upper respiratory tract. PLoS Pathog. 2013, 9, e1003057. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, S.; Foxman, B.; Weinberger, D.M.; Steiner, C.; Viboud, C.; Rohani, P. Identifying the interaction between influenza and pneumococcal pneumonia using incidence data. Sci. Transl. Med. 2013, 5, 191ra184. [Google Scholar] [CrossRef] [Green Version]
- Isaacs, A.; Burke, D.C. Viral interference and interferon. Br. Med Bull. 1959, 15, 185–188. [Google Scholar] [CrossRef]
- Dianzani, F. Viral interference and interferon. La Ric. Clin. E Lab. 1975, 5, 196–213. [Google Scholar] [CrossRef]
- Hirsh, S.; Hindiyeh, M.; Kolet, L.; Regev, L.; Sherbany, H.; Yaary, K.; Mendelson, E.; Mandelboim, M. Epidemiological changes of respiratory syncytial virus (RSV) infections in Israel. PLoS ONE 2014, 9, e90515. [Google Scholar] [CrossRef]
- Konala, V.M.; Adapa, S.; Gayam, V.; Naramala, S.; Daggubati, S.R.; Kammari, C.B.; Chenna, A. Co-infection with influenza A and COVID-19. Eur. J. Case Rep. Intern. Med. 2020, 7, 001656. [Google Scholar] [CrossRef]
- WHO. Overview of Influenza Activity Globally. Influenza Update N° 373 of 3 August 2020. Available online: http://www.who.int/influenza/surveillance_monitoring/updates/latest_update_GIP_surveillance (accessed on 12 August 2020).
- Li, Y.; Wang, J.; Wang, C.; Yang, Q.; Xu, Y.; Xu, J.; Li, Y.; Yu, X.; Zhu, H.; Liu, J. Characteristics of respiratory virus infection during the outbreak of 2019 novel coronavirus in Beijing. Int. J. Infect. Dis. 2020, 96, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Lu, P.; Fan, Y.; Xia, Y.; Liu, M. The clinical characteristics of pneumonia patients coinfected with 2019 novel coronavirus and influenza virus in Wuhan, China. J. Med Virol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Cuadrado-Payán, E.; Montagud-Marrahi, E.; Torres-Elorza, M.; Bodro, M.; Blasco, M.; Poch, E.; Soriano, A.; Piñeiro, G.J. SARS-CoV-2 and influenza virus co-infection. Lancet 2020, 395, e84. [Google Scholar] [CrossRef]
- Wu, D.; Lu, J.; Ma, X.; Liu, Q.; Wang, D.; Gu, Y.; Li, Y.; He, W. Coinfection of influenza virus and severe acute respiratory syndrome coronavirus 2 (SARS-COV-2). Pediatric Infect. Dis. J. 2020, 39, e79. [Google Scholar] [CrossRef]
- Wu, X.; Cai, Y.; Huang, X.; Yu, X.; Zhao, L.; Wang, F.; Li, Q.; Gu, S.; Xu, T.; Li, Y.; et al. Co-infection with SARS-CoV-2 and influenza A virus in patient with pneumonia, China. Emerg. Infect. Dis. 2020, 26, 1324–1326. [Google Scholar] [CrossRef]
- Azekawa, S.; Namkoong, H.; Mitamura, K.; Kawaoka, Y.; Saito, F. Co-infection with SARS-CoV-2 and influenza A virus. IDCases 2020, 20, e00775. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, S.A.; Safamanesh, S.; Ghafouri, M.; Taghavi, M.R.; Mohajer Zadeh Heydari, M.S.; Namdar Ahmadabad, H.; Ghasem Zadeh-Moghaddam, H.; Azimian, A. Co-infection with COVID-19 and influenza A virus in two died patients with acute respiratory syndrome, Bojnurd, Iran. J. Med Virol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Khodamoradi, Z.; Moghadami, M.; Lotfi, M. Co-infection of coronavirus disease 2019 and influenza A: A report from Iran. Arch. Iran. Med. 2020, 23, 239–243. [Google Scholar] [CrossRef] [Green Version]
- Wehl, G.; Laible, M.; Rauchenzauner, M. Co-infection of SARS CoV-2 and influenza A in a pediatric patient in Germany. Klin. Padiatr. 2020. [Google Scholar] [CrossRef]
- Wu, Q.; Xing, Y.; Shi, L.; Li, W.; Gao, Y.; Pan, S.; Wang, Y.; Wang, W.; Xing, Q. Coinfection and other clinical characteristics of COVID-19 in children. Pediatrics 2020. [Google Scholar] [CrossRef]
- Osterhaus, A.D.M.E. Influenza and RSV in a COVID-19 world. The First ESWI Webinar 11 June 2020. Available online: https://eswi.org/eswi-tv/influenza-and-rsv-in-a-covid-19-world-full-webinar/ (accessed on 25 June 2020).
- Jones, N. How Coronavirus Lockdowns Stopped Flu in its Tracks. Nature News. Available online: https://www.nature.com/articles/d41586-020-01538-8 (accessed on 18 June 2020).
- WHO. Coronavirus Disease (COVID-19) Pandemic. Press Conference of 15 June 2020. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019 (accessed on 1 August 2020).
- Lansbury, L.; Lim, B.; Baskaran, V.; Lim, W.S. Co-infections in people with COVID-19: A systematic review and meta-analysis. J. Infect. 2020. [Google Scholar] [CrossRef]
- Sánchez-Duque, J.A.; Orozco-Hernández, J.P.; Marín-Medina, D.S.; Cvetkovic-Vega, A.; Aveiro-Róbalo, T.R.; Mondragon-Cardona, A.; Failoc-Rojas, V.E.; Gutiérrez-Ocampo, E.; Villamizar-Peña, R.; Henao-Martínez, J.F.; et al. Are we now observing an increasing number of coinfections between SARS-CoV-2 and other respiratory pathogens? J. Med. Virol. 2020. [Google Scholar] [CrossRef]
- Kim, D.; Quinn, J.; Pinsky, B.; Shah, N.H.; Brown, I. Rates of co-infection between SARS-CoV-2 and other respiratory pathogens. JAMA 2020, 323, 2085–2086. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.T.; Chen, Z.M.; Chen, L.D.; Zhan, Y.Q.; Li, S.Q.; Cheng, J.; Zhu, A.R.; Chen, L.Y.; Zhong, N.S.; Li, S.Y.; et al. Coinfection with SARS-CoV-2 and other respiratory pathogens in COVID-19 patients in Guangzhou, China. J. Med. Virol. 2020. [Google Scholar] [CrossRef]
- Kakuya, F.; Okubo, H.; Fujiyasu, H.; Wakabayashi, I.; Syouji, M.; Kinebuchi, T. The first pediatric patients with coronavirus disease 2019 (COVID-19) in Japan; The risk of co-infection with other respiratory viruses. Jpn. J. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Costa, L.F.; Queiróz, D.A.; Lopes da Silveira, H.; Bernardino Neto, M.; de Paula, N.T.; Oliveira, T.F.; Tolardo, A.L.; Yokosawa, J. Human rhinovirus and disease severity in children. Pediatrics 2014, 133, e312–e321. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Pillai, P.; Miyake, F.; Nair, H. The role of viral co-infections in the severity of acute respiratory infections among children infected with respiratory syncytial virus (RSV): A systematic review and meta-analysis. J. Glob. Health 2020, 10. [Google Scholar] [CrossRef]
- Osterhaus, A.D.M.E. Clinical Manifestations of COVID-19, Influenza and RSV. The Second ESWI Webinar 18 June 2020. Available online: https://eswi.org/eswi-tv/clinical-manifestations-of-covid-19-influenza-and-rsv-full-webinar/ (accessed on 20 June 2020).
- Osterhaus, A.D.M.E. Intervention Strategies for COVID-19, Influenza and RSV. The Third ESWI Webinar 23 June 2020. Available online: https://player.vimeo.com/video/431944051 (accessed on 30 June 2020).
- Fan, B.E.; Lim, K.G.E.; Chong, V.C.L.; Chan, S.S.W.; Ong, K.H.; Kuperan, P. COVID-19 and mycoplasma pneumoniae coinfection. Am. J. Hematol. 2020, 95, 723–724. [Google Scholar] [CrossRef]
- Gayam, V.; Konala, V.M.; Naramala, S.; Garlapati, P.R.; Merghani, M.A.; Regmi, N.; Balla, M.; Adapa, S. Presenting characteristics, comorbidities, and outcomes of patients coinfected with COVID-19 and Mycoplasma pneumoniae in the USA. J. Med. Virol. 2020. [Google Scholar] [CrossRef]
- Stochino, C.; Villa, S.; Zucchi, P.; Parravicini, P.; Gori, A.; Raviglione, M.C. Clinical characteristics of COVID-19 and active tuberculosis co-infection in an Italian reference hospital. Eur. Respir. J. 2020. [Google Scholar] [CrossRef]
- Tadolini, M.; Codecasa, L.R.; García-García, J.M.; Blanc, F.X.; Borisov, S.; Alffenaar, J.W.; Andréjak, C.; Bachez, P.; Bart, P.A.; Belilovski, E.; et al. Active tuberculosis, sequelae and COVID-19 co-infection: First cohort of 49 cases. Eur. Respir. J. 2020. [Google Scholar] [CrossRef]
- Togun, T.; Kampmann, B.; Stoker, N.G.; Lipman, M. Anticipating the impact of the COVID-19 pandemic on TB patients and TB control programmes. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 21. [Google Scholar] [CrossRef]
- Arashiro, T.; Nakamura, S.; Asami, T.; Mikuni, H.; Fujiwara, E.; Sakamoto, S.; Miura, R.; Shionoya, Y.; Honda, R.; Furukawa, K.; et al. SARS-CoV-2 and Legionella co-infection in a person returning from a Nile cruise. J. Travel Med. 2020, 27. [Google Scholar] [CrossRef] [Green Version]
- van Arkel, A.L.E.; Rijpstra, T.A.; Belderbos, H.N.A.; van Wijngaarden, P.; Verweij, P.E.; Bentvelsen, R.G. COVID-19 associated pulmonary aspergillosis. Am. J. Respir. Crit. Care Med. 2020. [Google Scholar] [CrossRef]
- Blaize, M.; Mayaux, J.; Nabet, C.; Lampros, A.; Marcelin, A.G.; Thellier, M.; Piarroux, R.; Demoule, A.; Fekkar, A. Fatal invasive aspergillosis and coronavirus disease in an immunocompetent patient. Emerg. Infect. Dis. 2020, 26. [Google Scholar] [CrossRef]
- D’Ardes, D.; Boccatonda, A.; Schiavone, C.; Santilli, F.; Guagnano, M.T.; Bucci, M.; Cipollone, F. A case of coinfection with SARS-COV-2 and cytomegalovirus in the era of COVID-19. Eur. J. Case Rep. Intern. Med. 2020, 7, 001652. [Google Scholar] [CrossRef]
- Benkovic, S.; Kim, M.; Sin, E. 4 Cases: HIV and SARS-CoV-2 Co-infection in patients from Long Island, New York. J. Med. Virol. 2020. [Google Scholar] [CrossRef]
- WHO. Draft Landscape of COVID-19 Candidate Vaccines. Update of 10 August 2020. Available online: https://www.who.int/publications/m/item/draft-landscape-of-covid-19-candidate-vaccines (accessed on 12 August 2020).
- Burton, D.R.; Walker, L.M. Rational vaccine design in the time of COVID-19. Cell Host Microbe 2020, 27, 695–698. [Google Scholar] [CrossRef]
- Lauerman, J.; Paton, J. The First COVID Vaccines May Not Prevent COVID Infection. Prognosis of 15 June 2020. Available online: https://www.bloomberg.com/news/articles/2020-06-15/the-first-covid-vaccines-may-not-prevent-you-from-getting-covid (accessed on 2 August 2020).
- Graham, B.S. Rapid COVID-19 vaccine development. Science 2020, 368, 945–946. [Google Scholar] [CrossRef]
- Coperchini, F.; Chiovato, L.; Croce, L.; Magri, F.; Rotondi, M. The Cytokine Storm in COVID-19: An Overview of the Involvement of the Chemokine/Chemokine-Receptor System. Cytokine Growth Factor Rev. 2020, 53, 25–32. [Google Scholar] [CrossRef]
- Eyal, N.; Lipsitch, M.; Smith, P.G. Human challenge studies to accelerate coronavirus vaccine licensure. J. Infect. Dis. 2020, 221, 1752–1756. [Google Scholar] [CrossRef] [Green Version]
- Morris, K.V. The improbability of the rapid development of a vaccine for SARS-CoV-2. Mol. Ther. J. Am. Soc. Gene Ther. 2020. [Google Scholar] [CrossRef]
- Peeples, L. News feature: Avoiding pitfalls in the pursuit of a COVID-19 vaccine. Proc. Natl. Acad. Sci. USA 2020, 117, 8218–8221. [Google Scholar] [CrossRef] [Green Version]
- Hotez, P.J.; Corry, D.B.; Bottazzi, M.E. COVID-19 vaccine design: The Janus face of immune enhancement. Nat. Rev. Immunol. 2020, 20, 347–348. [Google Scholar] [CrossRef]
- Lyons-Weiler, J. Pathogenic priming likely contributes to serious and critical illness and mortality in COVID-19 via autoimmunity. J. Transl. Autoimmun. 2020, 3, 100051. [Google Scholar] [CrossRef]
- Ozdemir, C.; Kucuksezer, U.C.; Tamay, Z.U. Is BCG vaccination affecting the spread and severity of COVID-19? Allergy 2020. [Google Scholar] [CrossRef]
- Curtis, N.; Sparrow, A.; Ghebreyesus, T.A.; Netea, M.G. Considering BCG vaccination to reduce the impact of COVID-19. Lancet 2020, 395, 1545–1546. [Google Scholar] [CrossRef]
- Meena, J.; Yadav, A.; Kumar, J. BCG vaccination policy and protection against COVID-19. Indian J. Pediatrics 2020, 1. [Google Scholar] [CrossRef]
- Macedo, A.; Febra, C. Relation between BCG coverage rate and COVID-19 infection worldwide. Med. Hypotheses 2020, 142, 109816. [Google Scholar] [CrossRef]
- Kumar, J.; Meena, J. Demystifying BCG vaccine and COVID-19 relationship. Indian Pediatrics 2020, 57, 588–589. [Google Scholar] [CrossRef]
- Riccò, M.; Gualerzi, G.; Ranzieri, S.; Bragazzi, N.L. Stop playing with data: There is no sound evidence that Bacille Calmette-Guérin may avoid SARS-CoV-2 infection (for now). Acta Bio-Med. Atenei Parm. 2020, 91, 207–213. [Google Scholar] [CrossRef]
- WHO. Bacille Calmette-Guérin (BCG) Vaccination and COVID-19. Scientific Brief of 12 April 2020. Available online: https://www.who.int/news-room/commentaries/detail/bacille-calmette-gu%C3%A9rin-(bcg)-vaccination-and-covid-19 (accessed on 1 August 2020).
- O’Connor, E.; Teh, J.; Kamat, A.M.; Lawrentschuk, N. Bacillus Calmette Guérin (BCG) Vaccination Use in the Fight Against COVID-19—What’s Old is New Again? Future Oncol. 2020. [Google Scholar] [CrossRef]
- Redelman-Sidi, G. Could BCG be used to protect against COVID-19? Nat. Rev. Urol. 2020, 17, 316–317. [Google Scholar] [CrossRef]
- Dunning, J.; Thwaites, R.S.; Openshaw, P.J.M. Seasonal and pandemic influenza: 100 years of progress, still much to learn. Mucosal Immunol. 2020, 1–8. [Google Scholar] [CrossRef]
- Antony, S.J.; Almaghlouth, N.K.; Heydemann, E.L. Are co-infections with COVID-19 and Influenza low or underreported? An observational study examining current published literature including three new unpublished cases. J. Med. Virol. 2020. [Google Scholar] [CrossRef]
- Salem, M.L.; El-Hennawy, D. The possible beneficial adjuvant effect of influenza vaccine to minimize the severity of COVID-19. Med Hypotheses 2020, 140, 109752. [Google Scholar] [CrossRef]
- Hanney, S.R.; Wooding, S.; Sussex, J.; Grant, J. From COVID-19 research to vaccine application: Why might it take 17 months not 17 years and what are the wider lessons? Health Res. Policy Syst. 2020, 18, 61. [Google Scholar] [CrossRef]
- Mukherjee, R. Global efforts on vaccines for COVID-19: Since, sooner or later, we all will catch the coronavirus. J. Biosci. 2020, 45. [Google Scholar] [CrossRef]
- Merante, D. SARS-CoV-2, from its current highly contagious spreading toward the global development of an effective and safe vaccine: Challenges and uncertainties. Expert Opin. Drug Saf. 2020, 1–3. [Google Scholar] [CrossRef]
- WHO. Recommendations on the Composition of Influenza Virus Vaccines. Available online: https://www.who.int/influenza/vaccines/virus/recommendations/en/ (accessed on 1 August 2020).
- Bogaert, D.; Dockrell, D.H. 100 years of influenza research seen through the lens of Covid-19. Mucosal Immunol. 2020, 1–2. [Google Scholar] [CrossRef]
- Ozawa, M.; Kawaoka, Y. Cross talk between animal and human influenza viruses. Annu Rev Anim Biosci 2013, 1, 21–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiseleva, I.; Grigorieva, E.; Larionova, N.; Al Farroukh, M.; Rudenko, L. COVID-19 in Light of Seasonal Respiratory Infections. Biology 2020, 9, 240. https://doi.org/10.3390/biology9090240
Kiseleva I, Grigorieva E, Larionova N, Al Farroukh M, Rudenko L. COVID-19 in Light of Seasonal Respiratory Infections. Biology. 2020; 9(9):240. https://doi.org/10.3390/biology9090240
Chicago/Turabian StyleKiseleva, Irina, Elena Grigorieva, Natalie Larionova, Mohammad Al Farroukh, and Larisa Rudenko. 2020. "COVID-19 in Light of Seasonal Respiratory Infections" Biology 9, no. 9: 240. https://doi.org/10.3390/biology9090240